

NADPH Oxidase 3: Beyond the Inner Ear


1
Institute for Medical Microbiology, Immunology and Hygiene, Faculty of Medicine, University Hospital Cologne, University of Cologne, 50935 Cologne, Germany
2
German Centre for Infection Research, Partner Site Bonn-Cologne, 50931 Cologne, Germany
3
Cologne Cluster of Excellence on Cellular Stress Responses in Aging-Associated Diseases (CECAD), 50931 Cologne, Germany
Antioxidants 2024, 13(2), 219; https://doi.org/10.3390/antiox13020219
Submission received: 13 January 2024
/
Revised: 2 February 2024
/
Accepted: 6 February 2024
/
Published: 8 February 2024
(This article belongs to the Special Issue Reactive Oxygen Species in Different Biological Processes—Second Edition)
Abstract
:Reactive oxygen species (ROS) were formerly known as mere byproducts of metabolism with damaging effects on cellular structures. The discovery and description of NADPH oxidases (Nox) as a whole enzyme family that only produce this harmful group of molecules was surprising. After intensive research, seven Nox isoforms were discovered, described and extensively studied. Among them, the NADPH oxidase 3 is the perhaps most underrated Nox isoform, since it was firstly discovered in the inner ear. This stigma of Nox3 as “being only expressed in the inner ear” was also used by me several times. Therefore, the question arose whether this sentence is still valid or even usable. To this end, this review solely focuses on Nox3 and summarizes its discovery, the structural components, the activating and regulating factors, the expression in cells, tissues and organs, as well as the beneficial and detrimental effects of Nox3-mediated ROS production on body functions. Furthermore, the involvement of Nox3-derived ROS in diseases progression and, accordingly, as a potential target for disease treatment, will be discussed.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. NADPH Oxidase 3
1.1. The Family of NADPH Oxidases
Reactive oxygen species (ROS) were once described as mere byproducts of metabolism and as an unavoidable harming effect that cells have to cope with [1,2,3,4,5]. ROS is the summative term for a group of molecules that all contain oxygen but show more reactivity toward biological molecules than molecular oxygen [6,7,8]. A few early studies have observed beneficial effects of ROS during egg fertilization processes, but only suggestions for ROS sources were made [9,10,11]. Intriguingly, the discovery of an enzyme family present in nearly every form of life [12,13,14,15,16], including bacteria [17,18,19,20], mammals [13,21,22,23,24,25,26,27,28,29,30,31,32], fish [33,34], insects [35], plants [36,37,38], fungi [39,40,41,42] and worms [43], namely, the family of NADPH-dependent oxidases (Nox) was a surprise. This is because the sole function of this enzyme family members is the production of ROS [6,22,44], or to be precise, superoxide (O2−) [21,45,46,47,48,49], which was associated only with detrimental effects on cellular structures at the time. The first described family member was Nox2 [50,51,52,53,54], also named gp91phox (after its main subunit) or phagocyte NADPH oxidase [55,56,57] (after the most representative cell type, where it is expressed). Nox2 is responsible for the respiratory burst in phagocytes [55,58,59,60,61,62], and the ROS produced inside the phagosome of macrophages, neutrophils and monocytes fell back into the same functional role as before, i.e., a destructive or at least inactivating factor, but inside the phagosome at least directed to a specific target, which is the engulfed pathogen [45,63,64,65,66,67,68,69,70,71]. It is noteworthy that pathogen engulfment as process is not the sole pre-requisite for the respiratory burst. Phagocytosis itself triggers ROS production into the phagosome, nevertheless with varying intensities in dependency of the cargo [72,73,74]. Furthermore, Nox2 is not restricted to phagocytes, but also found in other cells and tissues [75,76,77,78]. In a short time, other Nox family members were discovered, and the enzyme family consists of seven members to date [6,13,79,80], namely Nox1 [81,82], Nox2 [83], Nox3 [84], Nox4 [52,85,86], Nox5 [52,87] and the two Dual oxidases (Duox), Duox1 and Duox2 [88,89,90,91,92]. It became quickly evident that the other Nox family members were either not only present or even absent in phagocytes, but likewise expressed in cells of the adaptive immune system [93,94] as well as in non-immune cells [22,48,66,86,89,95,96,97,98,99,100,101,102,103,104]. ROS production in non-phagocytes has more subtle functions [89,105,106,107,108] in contrast to the vast amounts of ROS (~2 nmol/min per 106 human cells) [55,62,109,110] that are produced in the phagosome during the respiratory burst [22,25,70,111]. These effects of ROS, which strongly diverge from their destructive power in the phagosome, regulate many important processes, such as cell growth and transformation [36,81,92,112,113,114,115,116,117,118,119,120,121,122], angiogenesis [123,124], vasodilatation [125,126,127,128], hormone synthesis [129], tissue remodeling [130], signaling transduction [31,131,132,133,134,135,136,137,138] neuronal development [139,140,141,142], and the list is continuously expanding [80,89,143,144,145,146]. Notably, in addition to their various functions, all isoforms are involved in immune responses during pathogenic invasion [35,147,148]. While oxidative stress describes an imbalance of the cellular redox status in general, the beneficial effects of an oxidative milieu, as listed before, are summarized under the term “oxidative eustress” [7,8]. Of course, when ROS production occurs in an uncontrolled manner or in the wrong subcellular location [149,150,151], a phenomenon termed oxidative distress [7,8], it leads, independently of the ROS source, to cell-, tissue- and organ damage or death [152,153,154,155,156,157,158,159]. Oxidative distress can finally contribute to the development of diseases, such as atherosclerosis [160,161]; cardiovascular diseases [100,108,162,163,164,165,166,167,168,169,170], such as stroke [130,171,172,173,174,175,176] or diabetes [177,178]; cancer [113,122,179,180,181,182,183,184] and neurodegenerative diseases [98,185,186,187,188,189].
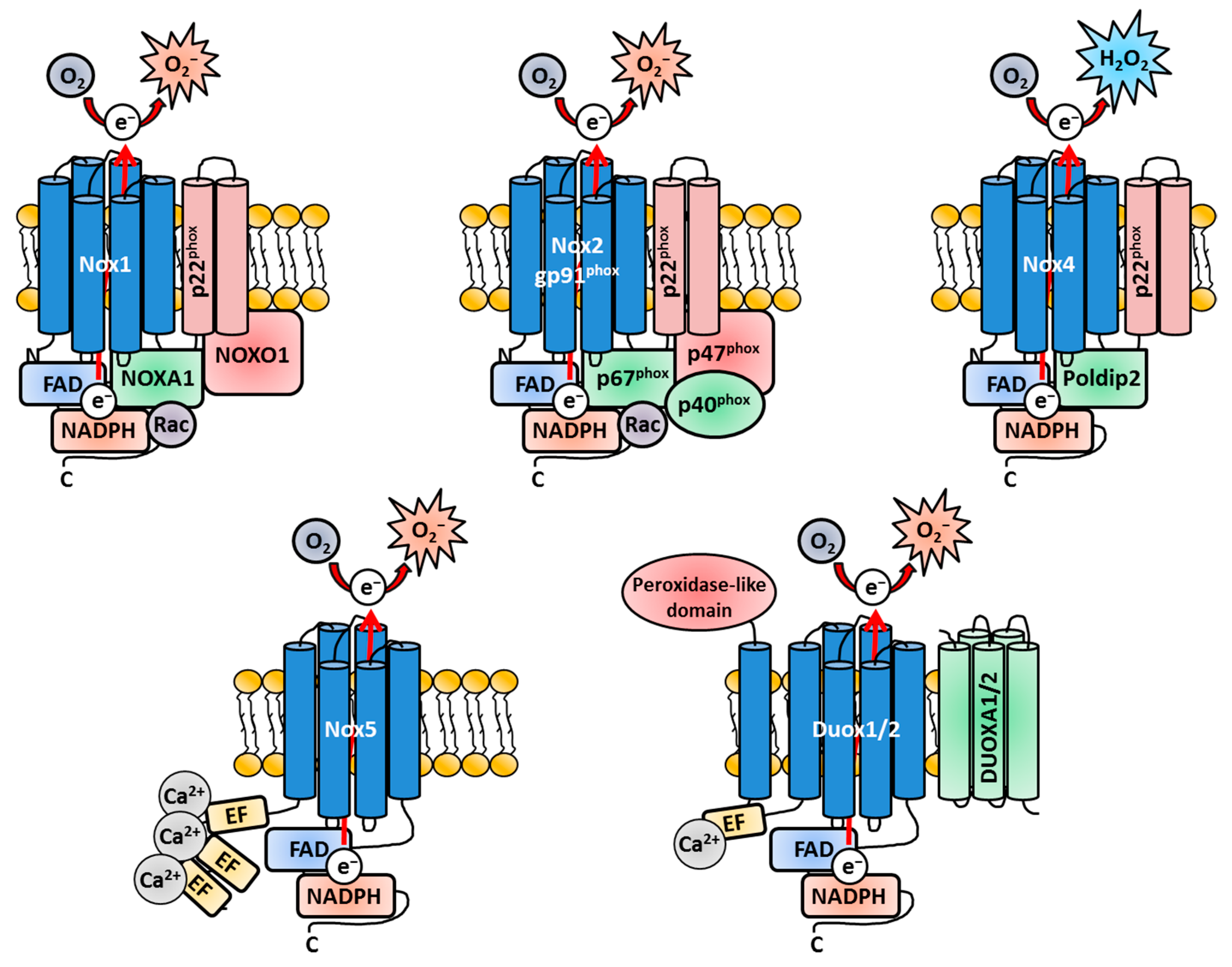
Structural Components of Nox Enzymes
Since Nox3 is a remarkable exception concerning the usage of Nox-related subunits, a general overview, which covers the similarities and differences of the Nox isoforms, is necessary and will support a better understanding of the latter parts of this review (Figure 1). All Nox family members share a membrane-bound catalytic core structure, a glycoprotein consisting of six trans-membrane α-helical domains (the actual gp91phox in Nox2), which contains two conserved heme groups near the N-terminus [109,190,191,192]. This core component is synthetized as a 65-kilo Dalton (kDa) precursor protein in the endoplasmatic reticulum (ER) [193] and gains its name-giving molecular weight of 91 kDa after heavy glycosylation during the transport through the Golgi network [194,195,196,197]. All Nox core structures end in a long cytosolic C-terminal tail, where the FAD- and NADPH-binding regions are located [198,199]. The gp91phox core unit forms a heterodimer with the membrane-bound protein p22phox [50,51,194,195,197,200,201] called b558 when fully assembled [51,53,54,202,203,204]. The heterodimer was named after the characteristic spectrum peak at 558 nanometers (nm) [53,202,205,206]. p22phox is an integral part of the Nox family members Nox1-4 [16,96,207,208] but is absent in Nox5, Duox1 and Duox2 [16]. Structurally, p22phox consists of four trans-membrane α-helices [209,210] and a proline-rich cytosolic region, which functions as docking site for other cytosolic adaptor subunits for the Nox enzymes [16,211]. The core subunit p22phox does not only serve as docking site for the cytosolic adaptor subunits of the Nox enzymes [207,212,213] but also has crucial functions for the flavocytochrome b558 core complex of Nox1-4 itself [27,207,214]. It has an important effect on gp91phox stabilization and loss of p22phox leads to retention of gp91phox in the ER [193,194,195,215,216,217]. p22phox further mediates the localization of gp91phox to cellular membranes in general [217,218,219] and the localization to the plasma membrane in particular [16,217,220]. While p22phox is not essential for all Nox isoforms, the subunit gp91phox represents the obligatory core component for all Nox enzymes [221], which contains the electron-shuttling apparatus. Electrons are transported from NADPH to FAD through the heme-containing domains and react with molecular oxygen to O2− [13,222] (Figure 1).
If and what adaptor proteins are necessary for enhanced activation or basal enzymatic activity greatly vary between the Nox isoforms [55,83,172,223,224,225]. Nox2 is activated by the cytosolic regulatory proteins p47phox [215,226,227,228], p67phox [227,228,229,230,231] and p40phox [232,233,234,235]. Nox1 utilizes NADPH oxidase organizer 1 (NOXO1) [25,236] and NADPH oxidase activator 1 (NOXA1) [25,236]. Furthermore, Nox1 and Nox2 both need the Ras-related C3 botulinum toxin substrate (Rac) enzymes, small guanosine triphosphate phosphohydrolases (GTPases), for enzymatic activity [96,237,238,239,240] (Figure 1).
p47phox is not active in unstimulated cells due to its auto-inhibitory region [212,241,242]. However, after stimulation (e.g., by pathogenic or chemical molecules), p47phox is phosphorylated on several serine residues [212,243,244,245,246,247] and binds to p22phox [211,213,246,248,249,250,251,252]. The kinase enzyme responsible for p47phox phosphorylation can be one of the various isoforms of the Protein kinase C (PKC) family. Which PKC isoform is activated depends on the stimulus, but p47phox-dependent Nox activation was discovered for PKCβ [253,254] and PKCζ [255,256]. p47phox phosphorylation leads to a conformational change, which allows its binding to p22phox [213,248,249,250,257]. In the cytosol, p47phox and p67phox are already tethered together via tail-to-tail interactions [252,258,259,260,261] and recruited to the plasma membrane/phagosomal membrane-residing flavocytochrome b558 complex [257,260,262,263,264]. p67phox is critical for the oxidase activity itself, since it regulates the electron flow from FAD to the two heme groups [232]. p67phox also binds directly to Rac, therefore facilitating its transport to the plasma membrane [255,265,266]. Similar to p47phox, p67phox is phosphorylated on many sites by various agonists, e.g., by members of the mitogen-activated protein kinases (MAPK) like p38 and extracellular signal-regulated kinase 1/2 (ERK1/2) [267,268,269,270]. While the direct activation of Nox2 strictly depends on p67phox, the recruitment of p67phox to the p22phox-gp91phox heterodimer is completely dependent on p47phox. In the absence of p47phox, p67phox will not translocate to the heterodimer and Nox2 is not activated [271,272]. Therefore, p47phox serves as important recruitment unit for the other subunits (Figure 1).
p40phox [258,273] and Rac [274,275], which beforehand transits into its active GTP-bound state [96,237,239,240,276,277,278,279,280], are both likewise recruited to the forming active Nox complex together with p47phox. While p67phox and NOXA1 are termed activator subunits [222,281,282] and p47phox and NOXO1 are regarded as organizer subunits (for Nox2 and Nox1/Nox3, respectively) [79,222,245,248,252,283,284,285], p40phox serves as scaffold-like platform, facilitating the translocation of the other subunits at least in the case of Nox2 [236,284]. While p40phox-mediated scaffolding is not essential for Nox2 enzyme activation per se [286,287], the involvement of p40phox leads to a two-fold increased ROS production [236] by facilitating the recruitment of p67phox to the plasma membrane [236] and phagosomal membrane [274,288,289].
The Rac proteins, specifically Rac1 (in human monocytes and macrophages) [290,291,292] and Rac2 (in human neutrophils) [293,294,295,296,297,298,299], are crucial for the activation of Nox1 and Nox2 [238,265,293,296,300,301,302], while they are completely dispensable for Nox4, Nox5 and the Duox enzymes [218,303,304]. The Rac enzymes serve two purposes in the context of Nox activation [83,290,305,306]. Firstly, they physically tether p67phox [265,302,307,308,309] to the plasma membrane [280] and the cytochrome b558; [310,311], and secondly, they induce a conformational shift of p67phox, thereby inducing its activation [231,232,265,302,312,313,314,315,316]. Initial experiments in cell-free systems have suggested a complete subunit-independent translocation of Rac enzymes to gp91phox [96,273,274,290,311]; however, it was later shown that Rac enzymes at least interact with and support the translocation of the subunits p67phox [266,307,312] 2004) and NOXA1 [274,300]. For further reading about the complex topic of the different Rac isoforms and their specific roles during Nox activation, I redirect the interested readers to other excellent reviews [275,317].
The accepted most current model for the activation of Nox2 as most representative isoform depicts as follows: The adaptor proteins for Nox2, namely p67phox and p47phox, exist in the cytosol as preformed complexes together with p40phox [234,318], which functions as linchpin between these two subunits [234,319,320,321]. After phosphorylation, the SH3 region of p47phox is exposed to the cytosol and binds to p22phox [246,249,252,271,283]. Since p67phox is tethered to p47phox, the heterodimer translocates together to p22phox [236,259,260,261,322,323], where p67phox also associates with gp91phox [232,255,312,314,315,324]. Hence, it is reasonable that no localization of this complex near the plasma membrane is observed when either gp91phox or p22phox is missing [234,273,319,320,321,325]. After dissociation of its inhibitory factor Rho GDP-dissociation inhibitor (RhoGDI) [238,311,326], Rac translocates to the membrane, where it binds to p67phox [265,266,307] and to the flavocytochrome b556 core complex [327]. p47phox as well as Rac are not absolutely essential for Nox2 activation [310,328], but play the role of important support units. They bind and orientate p67phox for optimal electron flow and activation of the Nox2 complex [24,109,232,312,314]. Nox1 is activated in a similar manner, however by utilizing its unique organizer and adaptor subunit NOXO1 and NOXA1, respectively (Figure 1). It is noteworthy that Nox1, while also being dependent on Rac for activation, cannot utilize the Nox2-related subunits p47phox and p67phox for functioning (Figure 1).
After full assembly, the Nox complex transfers electrons as hydride ions (H−) from NADPH to FAD. This step is mediated by the recruitment the p67phox subunit [231,232,255,329,330]. From FAD, the electrons are shuttled by the two heme molecules through the membrane-spanning part of the complex [109,331]. On the other site of the membrane, the electrons are transferred to molecular oxygen and form O2− [24,200,207,218]. So far, only the release of H2O2 instead of O2− has been clearly proven for Nox4 [332,333], while O2− is still the first-generated ROS subspecies at the Nox4 enzyme [334,335,336]. Nox4 contains a special E-loop on the extracellular site, which slows down the diffusion of O2− until it is dismutated to H2O2 [220,333]. Nox4 is a unique isoform in terms of regulation since no stimuli or regulatory subunits are necessary to directly induce Nox4 activity [218,337]. Nox4 is defined as being permanently active, as long as p22phox for the complete core structure is present [84,218,220,334,338] (Figure 1). The major adaptation for Nox4-derived ROS production is achieved by degradation- or new expression-induced various stimuli or stress conditions [339,340,341,342,343,344,345]. Nevertheless, some stimuli, like insulin [346] or LPS [347], can quickly trigger Nox4-mediated ROS production, which cannot be explained by expression of the protein itself. Accordingly, a few years after its discovery, polymerase (DNA-directed) delta-interacting protein 2 (Poldip2) [348,349] was identified as positive regulator, which directly binds to p22phox and increases Nox4-mediated ROS production [350,351] (Figure 1). Some other regulating proteins, e.g. Toll-like receptor (TLR) 4 [347,352] or protein disulfate isomerase [353], were identified, slowly revising the view of Nox4 as not being regulated by other factors except its expression [85]. Nox5, Duox1 and Duox2 have EF-hand domain-containing extensions on the cytosolic N-terminus, which bind Ca2+ [87,88,354] (Figure 1). Indeed, Ca2+ is the main activating factor for ROS production of these three Nox family members [87,88,355]. Additional adaptor proteins, Dual Oxidase Maturation Factor 1/2 (DuoxA1/2), were identified as factors necessary for maturation of Duox1/2 [356,357]. Duox1/2 also contains an additional peroxidase-like domain that extrudes to the extracellular site [88,109,358]. However, so far, only the Duox isoform of Drosophila melanogaster has shown an active peroxidase function of this domain, similar to the myeloperoxidase reaction The Duox isoform processes the produced H2O2 to generate hypochlorous acid (HOCl) [35]. Since this review summarizes new and old findings of Nox3, the reader is directed to other excellent reviews about Nox enzymes in general and in detail [13,21,22,23,24,25,26,27,28,80,359,360,361].
1.2. Nox3: Structure and Subunits
Nox3 combines many features of Nox1, Nox2 and Nox4 in terms of basal activation and regulatory subunit involvement, which is unique among the Nox enzymes. However, it took some years of intensive research to shed light on this most flexible Nox isoform. Nox3 was discovered together with Nox4 and Nox5 during a genetic screen in search for homologs of the firstly discovered Nox2, or more precisely, for proteins similar to the membrane-bound subunit gp91phox [362,363]. This research field was intensively investigated after the discovery that non-phagocytic cells also produce ROS and that phagocytes are not the only cells capable of this process. After Nox3, Nox4 and Nox5 were identified, Nox1 was cloned and described by Suh and colleagues [81], followed by Duox1 and Duox2 [88,109,364], therefore completing the enzyme family. The NOX3 gene is located on chromosome 6 in humans (gene locus 6q25.3), and suggestions were made that this Nox isoform appeared after the emergence of fish and amphibians [16]. The protein structure of Nox3 is very similar to Nox1, Nox2 and Nox4. Indeed, Nox3, which consists of 568 amino acids (aa), shows the strongest sequence similarity with gp91phox (58%) [109,365]. Initially, Nox3 was only weakly detected in human fetal kidney and the placenta [52,363], and further research related to Nox3 was dampened afterwards. New insights, like the predominant tissue locations or exact protein structures of Nox4 [84,334], Nox5 [87,354,366] and Duox1/2 [43,367,368], were unraveled shortly after their identification [81,82]. In contrast, it took 3 years until Nox3-focused research achieved a new momentum, mainly by three studies from the labs of Prof. Lambeth and Prof. Krause [355,369,370]. Nox3 shares many similarities with Nox2 concerning their protein structure (like the dependency on p22phox) and regulatory subunits (i.e., p47phox/NOXA1 and p67phox/NOXO1). Initial experiments of several research groups, which exclusively focused on Nox research, delivered the first observations where, if and how Nox3 is located and activated. All of these studies did not investigate ex vivo cells but instead used human cancer cell lines and co-expression approaches to combine the Nox3 core protein and various Nox subunits. Most of the findings are consistent between these initial studies, but some differences emerged back then, probably due to differences in the used culture cells. These differences were quite at the awareness of the researchers and discussed in the community back then [371]. Nevertheless, these first studies gained impactful insights into the Nox3 protein and its regulation, which will be discussed now.
1.2.1. Adaptor Subunits of Nox3
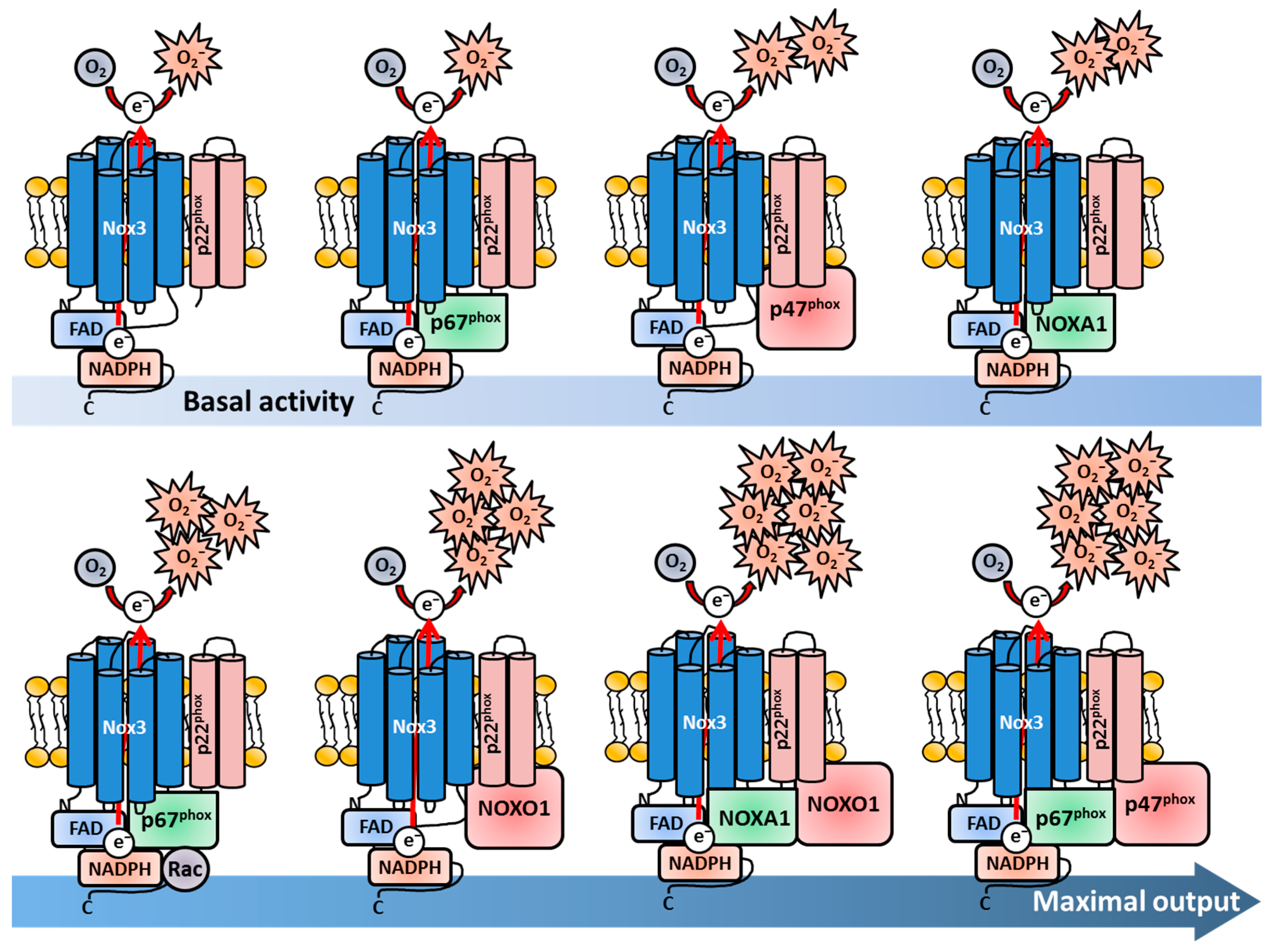
In 2004, two research groups investigated and published findings regarding the regulation of Nox3 nearly simultaneously [355,372]. Cheng and colleagues from the Lambeth lab used in vitro experiments, in which different combinations of human Nox3 (and Nox2/gp91phox and Nox1) and different Nox adaptor proteins were expressed in HEK293-H cells and COS-7 cells as “experimental vessels” [369]. They investigated the subunits associated with Nox2 and Nox1 at basal conditions and after stimulation with Phorbol 12-myristate 13-acetate (PMA). This chemical stimulates association of the Nox subunits by activation of the PKC, which results in robust ROS production [73,74,373,374]. Cheng et al. found that p67phox alone was not sufficient for basal or PMA-stimulated ROS production, while the expression of p47phox was sufficient for moderate ROS production by Nox3. Notably, this ROS production could not be further increased by PMA treatment. The combined presence of p47phox and p67phox led to the highest ROS production, which could be further increased by PMA stimulation. The presence or absence of Rac did not change the activation rate of Nox3. Interestingly, the expression of the Nox1 subunit NOXO1 also led to a strong activation of Nox3, which could not be further increased with PMA. In contrast, NOXA1 only slightly induced ROS production. Combinatory expression of adaptor proteins either for Nox2 (p47phox, p67phox) or Nox1 (NOXO1, NOXA1) led to maximal ROS output of Nox3. NOXO1 in combination with p67phox showed only minimally increased ROS production in comparison to sole NOXO1 presence. The combined expression of NOXA1 and p47phox in this system only led to PMA-dependent activation of Nox3. These data nicely showed that Nox3 is much more flexible than Nox1 or Nox2. While Nox2 strictly needs both p47phox and p67phox for activation, p47phox alone leads to moderate Nox3-derivedderived ROS production. Nox1, on the other hand, needs NOXO1 and NOXA1 for activation, while NOXO1 alone induced strong ROS production together with Nox3. Combinations of different adaptor proteins (e.g., NOXO1/p67phox or NOXA1/p47phox) only resulted in the low activation of Nox1 and Nox2, but induced a strong ROS production of Nox3. Taken together, these first experiments revealed the high flexibility of Nox3 in terms of adaptor protein usage (Figure 2).
Banfi and colleagues confirmed the flexibility of Nox3 in terms of adaptor protein utilization. The group analyzed mouse tissue samples via qRT-PCR and used histological staining to investigate the localization of Nox3 and the different subunits for the first time in vivo [355]. They detected mRNA expression of Nox3, NOXA1/p47phox and, to a lesser extent, also NOXO1/p67phox in the inner ear of mice. They followed a similar approach as Cheng and colleagues and analyzed the molecular function and regulation of Nox3 by a co-expression system in HEK293-H cells as “empty vessels”, which are devoid of Nox3. They confirmed ROS production by Nox3 in the complete absence of any adaptor subunit. In contrast to the findings of Cheng and colleagues, the group saw that p47phox or NOXA1 alone are not sufficient to increase Nox3-derived ROS production. Similar to Cheng and colleagues, they measured the strongest increase in ROS production, when either NOXO1 and NOXA1 or when NOXO1 and p67phox were expressed together. This Nox3-mediated ROS production could not be further enhanced by PMA stimulation. The combination of p47phox and p67phox or the combination of p47phox and NOXA1 resulted in a robust PMA-induced ROS increase, while basal ROS production was only minimally increased [355]. The most obvious discrepancy between the two studies from Cheng et al. and Banfi et al. was the Nox3-derived ROS production with NOXO1 as sole subunit. In the study from Banfi et al., only a small increase in ROS production was measured after PMA stimulation [355], while Cheng and colleagues described a strong increase in basal ROS production, which could not further be enhanced by PMA [369]. One has to consider that both studies did not use ex vivo cells of any kind, in which the actual Nox3 is present and active. It was and is common practice to use artificial co-expression systems in cancer cell lines to obtain initial insights into protein function. Of course, with varying cell lines, the experimental outcomes can also differ, which explains the contrasting results of the two groups. Nevertheless, it is impressive that both studies gathered the mostly similar results for Nox3, which delivered important first hints of the regulation (and location) of this Nox family member. The flexibility of adaptor unit utilization by Nox3 is especially distinct from other Nox family members (Figure 2). The Lambeth lab later discussed that the potential or non-potential adaptability of different Nox enzymes reflect their functions and tissue locations [370]. In contrast to phagocytes, for non-immune tissue cells, it may be more biologically relevant to have a redundant subunit protein backup in case of gene mutations or deletions to keep the Nox enzyme functional and suppress disease development in case one or more regulatory Nox subunits are altered. Nox3, with its flexibility, is a shining example for this statement.
Takeya and colleagues analyzed and compared different splicing variants of the NOXO1 gene (termed α, β, Δ, γ) and its subsequent protein products [96,229,367] in the context of Nox3 activation. While the most abundant variant NOXO1β was investigated before [369], Takeya et al. reported that also the splicing variant NOXO1γ was sufficient to induce Nox3-mediated ROS production [371]. They also described a strict dependency on the p22phox subunit for NOXO1γ-mediated activation of Nox3. Furthermore, they showed that binding of either NOXO1β or NOXO1γ to phosphatidylserins in the plasma membrane is mediated by a specific amino acid sequence, called the PX motif. This PX motif is required for membrane binding of Nox subunits in general [237,369,375,376,377]. However, a basal ROS production by Nox3 was detected even after destruction of the PX domains in the NOXO1 subunits [371]. Following studies intensified the details of the interplay between Nox3 and its various subunits. Miyana and colleagues observed slightly enhanced Nox3 activation by p67phox alone in HeLa, CHO and COS-7 cells and detected enhancement with NOXA1 in CHO, COS-7, but not in HeLa cells [378]. Maehara et al. showed that the SH3 domain of p67phox, which is necessary for the full activation of Nox2 [379], is interestingly not needed for Nox3 activation [380]. In a follow-up study, the group further identified a highly conserved activation domain of p67phox (in the aa 190–210), which is crucial for activation of Nox1, Nox2 and Nox3. The identified crucial residues were Tyrosine 198, Leucin 199, Valin 204, Leucin 193 and Asparagin 197 [222]. However, this domain did not have any influence when Nox3 was activated by p67phox together with p47phox, explaining the previous observation that p67phox and its SH3 domain alone are of no significance for Nox3 activation [380]. Taura et al. investigated the SH3 domain of p47phox and NOXO1 and observed that the domain is important for full Nox3 activation after PMA-stimulation [381]. This occurred even in the absence of p67phox, therefore arranging NOXO1 and p47phox above p67phox in the hierarchy of Nox3 activation. The group further identified the amino residue Interleukin 152 in a short N-terminal tandem region in the SH3 region of p47phox. This residue was found to be crucial for the activation of p47phox-mediated activation of Nox3, even in the absence of p67phox. Notably, the residue was also found in NOXO1 [381].
1.2.2. The Nox3-p22phox Complex
While the interactions of Nox3 and the diverse Nox subunits obtained some new and fascinating insights during that time, Ueno and colleagues investigated the interplay of the other membrane-bound subunit of the Nox1/2/3/4 complex namely p22phox [382]. Ueno et al. also used co-expressing systems in COS-7, CHO and HEK293-H cells to investigate this topic. Most importantly, they demonstrated, for the first time, that Nox3 physically interacts with p22phox and that both the presence and the interaction of ph22phox are essential for Nox3 activity. They detected minimal, but constitutive ROS production in the absence of any adaptor subunit, confirming previous findings [355,372] (Figure 2). They also measured enhanced ROS production after co-expression of Nox3 with p47phox, which could be further enhanced by combined expression with p67phox. Like Cheng et al., but in contrast to Banfi et al., the group found that the sole expression of NOXO1 leads to a strongly enhanced ROS production independently of PMA stimulation. Interestingly, they could show that NOXO1 is constitutively bound to p22phox, which explains the PMA-independent increase in ROS production. NOXA1 alone only slightly increased ROS production like seen before [355,372]. In contrast, the p47phox-mediated Nox3 activation could be further increased by PMA stimulation. The group also described no necessity for Rac proteins in Nox3 activation as reported previously [370].
Kawahara et al. not only characterized the role of p22phox during activation of Nox1-5 in co-transfected HEK293 cells, but also investigated the interaction of p22phox with various adaptor subunits [337]. They saw that Nox3 activation was strongly diminished in p22phox-silenced HEK293 cells despite the co-transfection of any subunit confirming the crucial role for p22phox in Nox3 basal activation. Notably, they also saw a strong activation of Nox3 with NOXO1 or p67phox alone, a minor but detectable activation with NOXA1 alone and the strongest activation after combined transfection with either NOXO1/NOXA1 or p67phox/p47phox. In addition, Kawahara and colleagues generated a C-terminally-truncated p22phox protein, which still formed a complex with gp91phox, but could not bind to the organizer subunits NOXO1/p47phox. HEK293 cells, which co-expressed this truncated p22phox protein together with NOXO1, showed a strongly diminished activity, suggesting that p22phox has not only a direct stabilizing effect for Nox3, but is also important for binding of organizer subunits and subsequent Nox3 activation. As shown by two groups before [370,372], Nox3, in contrast to Nox1 and Nox2, has the remarkable ability to be activated only by the presence of an activator subunit (p67phox, NOXA1) without any organizer subunit (p47phox, NOXO1). This phenomenon is also unique among the Nox enzyme family members (Figure 2) Interestingly, during additional co-expression of NOXA1 together with Nox3, NOXO1 and the truncated p22phox protein, the group observed a nearly restored activity of Nox3. These findings indicated the possibility that Nox3, as exclusive exception in the Nox enzyme family, might be able to bypass the p22phox-mediated binding of the organizer subunits NOXO1/p47phox by the direct binding of an activator or organizer protein to the Nox3 core structure itself. This again demonstrates the fascinating flexibility of Nox3 [337]. Nakano et al. characterized the role of p22phox for the actual biosynthesis of Nox3 in co-expression systems with HEK238 and CHO cells [217]. They firstly described the characteristic spectrum peak at 558 nm for Nox3, which suggested that the structure of Nox3 probably resembles Nox2. Chemical inhibition of heme synthesis in HEK293 cells transfected with Nox3 and p22phox resulted in a completely blunted ROS production demonstrating that heme is crucial for Nox3 functioning. In vitro translation with cDNA in a rabbit reticulocyte lysate system resulted in a 53 kDa-sized protein product, which was the first size description of Nox3. This product further underwent N-linked glycosylation as previously discovered for gp91phox [197] and knock-down of p22phox via small interfering (si)RNA resulted in reduced ROS production, as described previously [337]. The group also observed that p22phox was crucial for plasma membrane targeting of Nox3, which remained diffusely distributed in the cytosol in the absence of p22phox, which was also in line with previous observations [274,337,382]. In addition, the study confirmed other previous results concerning NOXO1 and p22phox, i.e., that NOXO1 interacts with p22phox at the plasma membrane [274,337,382] but does not necessarily directly bind to it [372,383]. Miyano and colleagues confirmed the N-glycosylation and p22phox-dependent maturation of Nox3 in CHO cells [384], hence completing the picture of the interplay between Nox3 and p22phox.
1.2.3. Nox3 and Rac
Previous observations of Rac-independent activation of Nox3 [370,382] were revised and challenged by a study from Ueyama et al., which investigated this topic by a co-expression system in HEK293-H and CHO-K1 cells [274]. The group could confirm many of the previous findings, like constitutive Nox3 activity without any subunit and maximal activity enhancement with NOXO1 alone or with p47phox/p67phox combined [96,355,372,382]. Interestingly, the authors described a strong increase of ROS production after co-expression of Nox3 and Rac1 alone, which is in contrast to previous results. Additional expression of p67phox further enhanced the activity of Nox3. This study confirmed again the flexible use of Nox activator or organizer proteins and underlines the observation that Nox3 does not strictly need organizer and adaptor units for moderate ROS production. Accordingly, Miyano and colleagues revised their findings concerning Rac dependency for Nox3 activation of their previous study in 2005 [382]. In co-expression experiments with various cancer cell lines (HeLa, CHO, COS-7), they observed a small p67phox/NOXA1-dependent enhancement of Nox3 activation by Rac. They further showed that p47phox, either in combination with p67phox or NOXA1 was necessary for maximal Nox3 activation. In contrast, the combined expression of NOXO1 and either p67phox or NOXA1 showed no dependency on Rac. Moreover, Nox3 was activated even more strongly when Rac binding was inhibited by site mutation of p67phox or NOXA1 [378]. The group next focused on the role of all Rac isoforms (Rac1-3) in respect to Nox1 and Nox3 activation [275]. For that again a co-expression systems with HeLa and HEK293 cells was used. In addition, human neutrophil fractions and the macrophage-like cancer cell line RAW246.7 were analyzed to investigate this topic. The group could confirm previous results of the Rac1-dependent enhancement of ROS production mediated by p67phox or NOXA1 [274,378] and showed that Rac2 and Rac3 can function redundantly in this process. For this reason, Rac is not a crucial component for Nox3-derived ROS production but can enhance Nox3 enzyme activity in combination with a defined set of subunits (Figure 2).
A complete new set of organizer subunits, not only for Nox3, but also for Nox1, was identified by Gianni and coworkers [385]. They investigated a possible role of the two Tyrosine kinases (tyrosine kinase substrate with five SH3 domains), Tsk4 and Tsk5 [386,387,388], for Nox enzyme activation. Co-expression of Nox3 and Tsk4/5 alone increased the ROS production by Nox3 in HEK293 cells, similar to NOXO1 or p47phox. Unfortunately, while also a role in Nox4 activation was discovered [353,389], to date, no further investigation of these new interesting subunits and Nox3 was conducted.
Taken together, the heterodimer consisting of Nox3 and p22phox is the basic minimal structure independent of the investigated species and alone is sufficient to produce substantial amounts of ROS without any adaptor subunit. Both p22phox and Nox3 depend on each other for proper maturation in the ER and for plasma membrane translocation, and either one is degraded when the other part is missing. Similar to the flavocytochrome b558 of Nox2, the Nox3-p22phox heterodimer shows the characteristic spectrum band at 558 nm, contains the two essential heme groups and undergoes heavy glycosylation during maturation. However, in contrast to Nox2 or Nox1, if and to what extent the basal ROS production of Nox3 can be enhanced highly depends on the regulatory subunit set and the investigated species [172,390]. In human cells, the basic ROS-producing activity of Nox3 can be enhanced by p47phox, NOXA1 or NOXO1 alone, with NOXO1 showing the strongest effect [391]. The ROS production can be maximized by presence of both NOXO1 and NOXA1 or p47phox and p67phox. p67phox alone is not sufficient for ROS production enhancement of Nox3 and either needs Rac1 or p47phox in addition. In mice, NOXA1, but not NOXO1 alone, is sufficient to slightly enhance the basal ROS production, however in dependency on Rac. A combination of NOXA1 and NOXO1 leads to maximal ROS production. p67phox-mediated enhancement of ROS production can be further increased by Rac, however not when both p67phox and p47phox are present. In summary, Nox3 is unique among all Nox isoforms since it can utilize all Nox organizer or activator subunits or a combination of them, while also showing basal activity like Nox4 (Figure 2). As a critical side note, all described findings and molecular interactions of Nox3 and its adaptor subunits have not been investigated to date in any ex vivo cell type that naturally expresses these proteins.
2. Location of Nox3
2.1. Location of Nox3 in Organs and Tissues
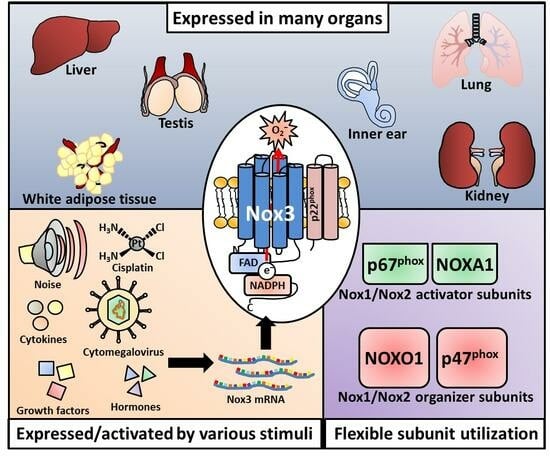
While often cited in many articles [12,13,172,285,361,392], including previous articles from our own lab [393,394], as “only/exclusively expressed in/restricted to the inner ear”, Nox3 was detected in various organs and cell types over time. This restriction of Nox3 presence to the inner ear, the continuous transmission through the literature and the subsequent underestimation of influence on its cellular processes were due to the first in vivo investigation in animals that lacked Nox3. The animals showed a remarkable head-tilting phenotype [370], and the inner ear as a localization of Nox3 was swiftly discovered [355]. However, several years of excellent research on Nox3 revealed other important locations of Nox3, for example the lung [395] and the liver [396]. Nevertheless, this review will firstly focus on the inner ear as the first location where Nox3 was discovered in vivo and then continues with a broad overview of organs and cell types where Nox3 could or could not be detected.
2.1.1. Nox3 in the Inner Ear
The inner ear of mammals provides two crucial functions for the orientation of the organism, namely the sensation of sound and the sensation of balance and orientation [397,398]. While the cochlea is responsible for sound processing [399,400], the vestibular system maintains balance and orientation [401,402,403]. Paffenholz and colleagues discovered the probably most intriguing phenotype for Nox3, since Nox3-deficient mice showed a strong head-tilting behavior, targeting the inner ear as only research focus for Nox3 for some years [370]. Since this review focuses on Nox3, descriptions of the vestibular system and cochlea are mandatory at this point. It should be mentioned that as sure as Nox3 is not only expressed in the inner ear, Nox3 is also not the only Nox isoform expressed in the inner ear [404]. Cheng and Lambeth detected the expression of Nox2 and Nox4 besides Nox3 in the murine inner ear, while Nox1, Duox1 and Duox2 were absent [369]. Nox2 is expressed in the microglia, which reside in the spiral ganglion [405,406], while Nox4 is expressed in the vascular endothelium, which also supports the stria vascularis [407]. More importantly, studies that investigate the expression profile of Nox enzymes must carefully distinguish between the vestibular system and the cochlea and not generalize their findings to the whole inner ear.
Nox3 in the Vestibular System
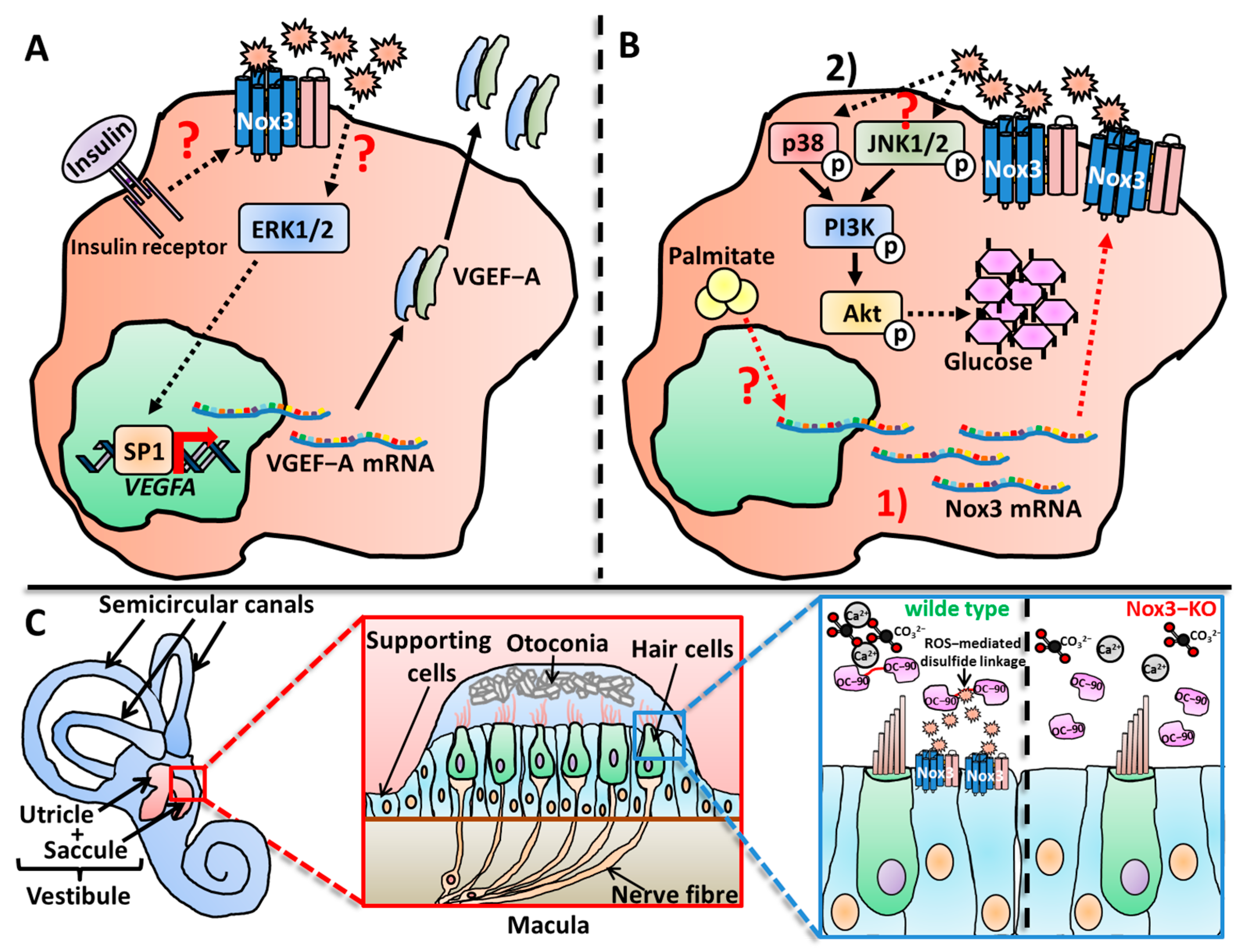
In the vestibular system, three semicircular canals and the cristae ampullas form a functional unit to detect and coordinate angular (rotational) acceleration [408]. In the otolith organs (consisting of the saccule and the utricle) the neuroepithelial maculae, a layer of sensory epithelial cells, detect gravity and linear acceleration [409,410]. An extracellular gelatinous matrix is located on top of the maculae and embedded in this matrix layer are crystalline, polymorphic structures called otoconia [411]. The otoconia are formed directly above the sensory hair cells, which are mechanoreceptors that transfer the sensory information to the ganglion cells via chemical synaptic activation. Ganglion cells show discharge patterns in the absence of any stimulation [412,413,414], which are mediated by a steady neurotransmitter release from the pre-synaptic hair cells in a calcium ion (Ca2+)-dependent manner [415,416,417,418,419,420]. Otoconia function as solid masses, which are affected by change of gravity or linear acceleration [401,408,410,421]. Calcium carbonate (CaCO3) is the main inorganic compound forming the crystalline structure of the otoconia [401,422]. Indeed, the protein Pendrin, a HCO3−/Cl− exchanger channel, as well as Otopetrin1, a proton channel [423,424], are crucial for proper otoconial formation [425,426,427,428,429]. The otoconia are not completely inorganic, since a number of proteins were identified as organizing or structuring components, and the list of otoconia-relevant genes is expanding [430,431,432,433]. The major proteinaceous component found in the otoconia is otoconin 90/95 (OC-90/95) [434,435,436]. OC-90/95 is a 90–95 kDa-sized glycoprotein that belongs to the family of secretory phospholipases A2 [435,436]. OC90/95 is produced by the non-sensory epithelial cells of the inner ear from where it is secreted into the endolymph [433,434,436,437]. It is necessary for proper formation of the inorganic CaCO3 crystallites into the otoconial organic mass [438,439], and OC-90-deficient mice lose nearly 50% of their otoconial structures, leading to imbalance. Importantly, the hearing capabilities remain intact in these animals [438,440]. Moreover, a disturbed longitudinal flow of OC-90 from the vestibule to the endolymphatic sac also leads to otoconial malformation, meaning that not only the presence, but also the location of OC-90/95, is of importance for otoconial formation [441,442]. OC-90 also recruits other proteins, such as Otolin-1 [438,443], a component of the gelatinous matrix. Other examples are Otogelin, which is found in the surrounding extracellular layer [444,445], and Otoancorin, which is located between the sensory hair cells and the overlaying extracellular matrix [446]. The concerted action and coordination of the various inorganic and organic components are necessary for the proper formation of functional otoconia [409,410,431,447,448].
For the investigation of the vestibular system, mice (or other model organisms), which harbor mutations in gene loci that affect the otoconial formation are obviously the most useful [421,449,450,451,452]. Several altered gene loci that led to loss, disturbed size or dislocation of otoconia and, subsequently, to a malfunctioning vestibular system were identified and phenotypically described [408,410,425,447,452,453]. The first-described gene locus associated with the head-tilting phenotype in mice was logically named “Tilted-head” (thd) [454]. Unfortunately, besides the phenotypical description of the mice, this locus was not further investigated. In the second detected locus “tilted” (tlt), the gene that encodes otopetrin 1 is localized. Otopetrin 1 is also crucial for otoconia development [425,426,427,428]. The analyzed third locus “head tilt” (het) containing two mutated alleles, het [455] and het2J, was characterized and both mutated alleles were associated with loss of otoconia [456]. After further characterization [370], this locus was logically renamed Nox3het [457]. The Nox3het−3J allele was generated during a mutagenesis project [458] and later investigated and associated with Nox3 by Paffenholz and colleagues [370]. The Nox3het−4J allele was also generated during a mutagenesis program in C57BL/6J mice [459] and the Nox3het−5J allele spontaneously appeared at a Jackson Laboratory in the CBySmn.CB17-Prkdcscid/J mouse strain [459].
Paffenholz et al. analyzed some other natural occurring and mutagenesis-induced mutated alleles in the het locus, which were named hetR96, hetR542 and het3J [370]. Several affected genes were identified, one of them with a high homology to the previously described human NADPH oxidase 3 gene NOX3 [52,363]. The hetR96 mutant allele resulted in a Nox3 protein, which lacked three of the trans-membrane α-helices, a complete catalytic domain and the binding sites for NADPH and FAD (see Section 1.2 and Figure 1). Also, a region responsible for heme binding was disturbed. The homologous deletion of Nox3 manifested itself by an obvious heat-tilting phenotype and lowered motor coordination (i.e., disturbance during balancing and swimming). Notably, while the vestibular system was clearly disturbed in Nox3-deficient mice, the hearing capacity was unaffected, at least in these investigated animals. Histological analysis of the vestibular system in Nox3-deficient mice revealed that the observed phenotype was based on the complete lack of otoconia in homozygous (but not heterozygous) mice throughout the complete lifespan (embryonic stage to adult) [370]. Paffenholz and colleagues described Nox3 as a ROS-producing enzyme in the inner ear that is crucial for the morphogenesis of the otoconia and subsequently for a properly functioning vestibular system. However, at that time, the molecular mechanism of the Nox3-derived ROS, which is responsible for otoconia formation, was pure speculation [370].
A parallel study of Banfi and colleagues also reported Nox3 presence in the inner ear of mice and rats by cloning experiments with cDNA [355]. The group also detected Nox3 expression at low protein levels in the brain, the skull and the fetal kidney. Nox3 expression in the fetal rat kidney was later confirmed by Reinehr and colleagues [460]. The predicted murine amino acid structure showed 81% sequence similarity with the human sequence. The group could also confirm the vestibular system as Nox3-expressing tissue [355,370] and further specified, for the first time, the sub-tissue location, i.e., the non-sensory epithelial cell layer of the saccule, by in situ staining [355].
All of the so far described mutant alleles of Nox3 (namely Nox3het, Nox3het−2J, Nox3het−3J, Nox3het−4J, Nox3het−5J, Nox3hetR96 and Nox3hetR542) lead to otoconial and/or vestibular-evoked potential responses, which can be measured by a non-invasive method developed by Jones et al. as reliable tool to identify loss-of-function mutations for Nox3 [408,461]. The results of these measurements were comparatively analyzed and summarized in the work of Flaherty and colleagues [457] and recommended for further interested readers.
A few years later, Mohri and colleagues generated mice that expressed Nox3 coupled to the red fluorescence tag dtTomato to re-investigate the precise locations of Nox3 in the inner ear in a ground-breaking study for the field [462]. They reported the “tilted head” phenotype and otoconial defects in Nox3-deficient animals as described before [355,370]. Additionally, they observed strong Nox3 protein expression in the endolymphatic sac and duct at early embryonic stages (at day 18.5). However, right after birth and 3 days after birth, only weak Nox3 expression was detected in the semicircular canals and the vestibule. Importantly, the group further showed that Nox3-derived ROS are majorly produced by non-sensor epithelial cells [355], which face the lumen of the endolymphotic sac and duct, as well as the semicircular canals and vestibule. A mechanism of Nox3-derived ROS for otoconial development was not made during this investigation. Together, these studies clearly showed that Nox3 is located in the vestibular system and is crucial for the proper development of the otoconia and, accordingly, for balancing (see Section 4.3.1).
Nox3 in the Cochlea
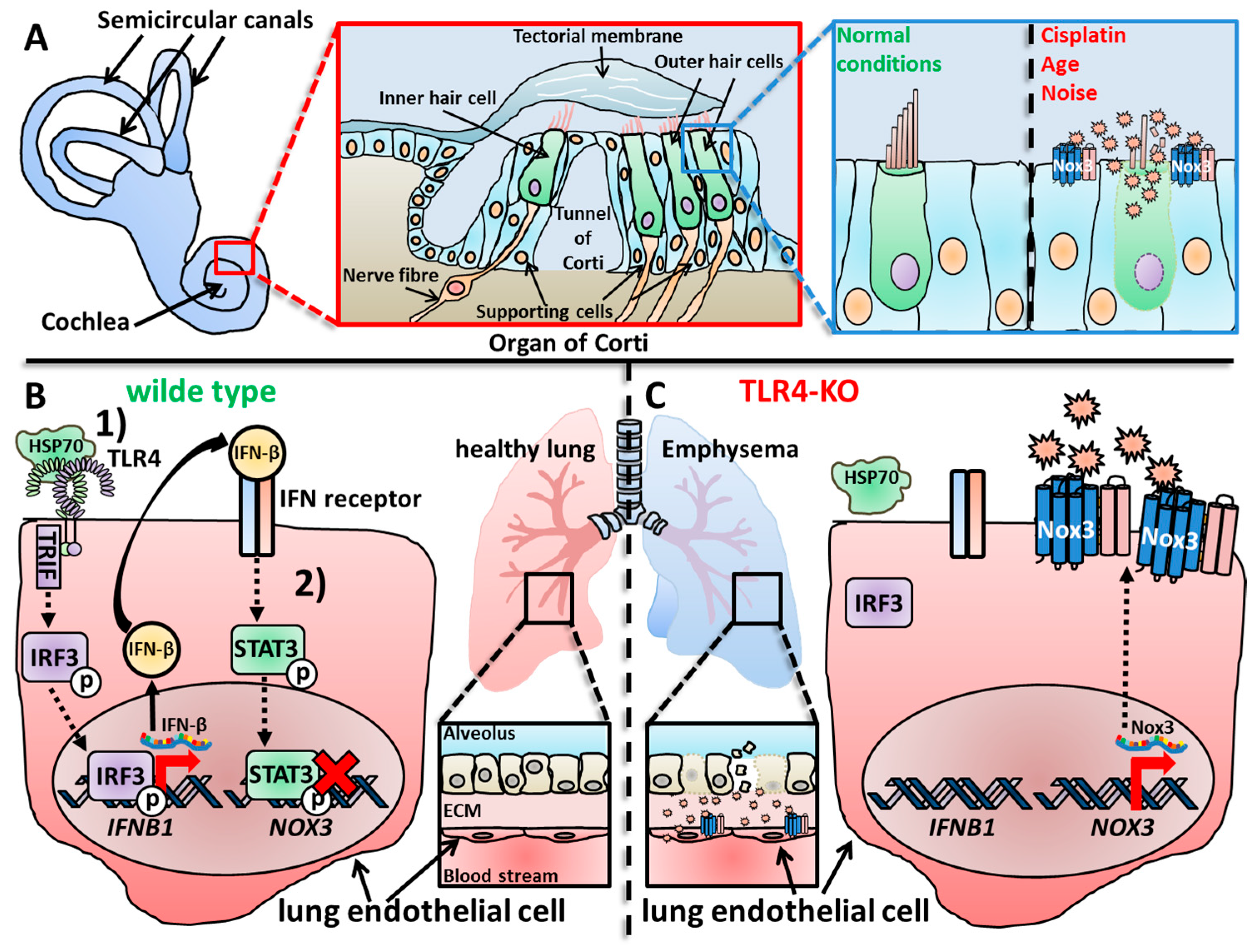
The cochlea is the organ responsible for hearing [399,400,463], and several studies have described Nox3 expression in this area of the inner ear [355,404,462]. Banfi et al. detected expression of Nox3 mRNA in parts of the adult mouse cochlea, precisely the organ of Corti and the spiral ganglia, while Nox3 was not expressed in dorsal root ganglia [355]. However, in contrast to Banfi and colleagues, who analyzed mouse samples, Nox3 was not detected in the spiral ganglion neurons of the rat cochlea [370]. However, while the loss of Nox3 and the correlative deficiency of otoconia is detrimental for balance, head positioning and gravity sensing [370,408,464,465,466], the loss of Nox3 in the cochlea leads to a rather protective outcome for the tissue and the hearing capacity (see Section 5.1). Overproduction or production of ROS in the wrong location can lead to irreversible cell and tissue damage, called oxidative distress [7,8]. This phenomenon was also described in previous studies, which showed that excessive ROS production in the cochlea in general has a great impact on age-, noise- and drug-induced hearing loss (see Section 5.1.1, Section 5.1.2, Section 5.1.3 and Section 5.1.4) [467,468,469,470,471,472,473,474]. Since Nox3 was firstly discovered in the inner ear, it was only reasonable during the time of early Nox3-related research to assume that Nox3 is most probably responsible for the destructive ROS production in the cochlea [355,370]. However, it took several years until this correlation was proven true [462,475,476,477,478]. Similar to Nox isoform expression in the vestibular system, Nox3 is not the only Nox isoform expressed in the cochlea. Vlajkovic and colleagues detected all seven isoforms, Nox1-5 and Duox1-2 in the rat cochlea [479]. The group further investigated the specific cellular expression of the Nox isoforms, which will be discussed later in this review (Section 2.2.1). Mohri and colleagues used their well-established mouse strain, in which Nox3 is coupled to the red fluorescence tag dtTomato [462,480,481]. They detected no Nox3 expression in the cochlea after 1 and 2 months after birth. Nox3 expressions started, at the earliest, after 6 months accompanied by outer hair cell (OHC) loss. Further analysis revealed an increasing Nox3 expression in supporting cells between 1 and 6 months, while OHCs showed no Nox3 expression. This is a ground-breaking study for Nox3-related research, since Mohri and colleagues not only investigated the exact location of Nox3 in the cochlea, but also described its role for different forms of hearing loss, which will be discussed in Section 5.1. This was further completed by Rousset and colleagues who detected expression of Nox2 and Nox3 mRNA in the mouse cochlea, but more importantly, in the human cochlea [404].
2.1.2. Nox3 in Other Organs
Many studies have investigated the topic of Nox3 expression in various organs and tissues. Surprisingly, during years of intensive research, it became clear that Nox3 is present in many organs and cell types with a plethora of different functions, which will be discussed later in Section 4. Unfortunately, most expression data available for Nox3 are restricted to mouse or rat tissue, and information of Nox3 expression patterns in human tissues is scarce.
In addition to the inner ear, Nox3 was detected either as protein or, mostly, as mRNA in mouse lung tissue [482,483], in mouse testes [484], in mouse white adipose tissue [485] and in the mouse upper circumvallate papillary epithelium of the tongue [486]. Nox3 mRNA could not be detected in the naïve mouse fetal or adult liver [487].
In the rat, Nox3 mRNA expression was detected in rat skeletal muscle, testis, lung, prostate, colon [488], brain [488,489,490], spinal cord neurons [491] and the adult rat kidney [492].
In contrast to murine or rat tissue, Nox3 is expressed in the avian liver [493].
The few studies which investigated Nox3 expression in ex vivo human tissue samples have described Nox3 expression in human placental tissue [494], as well as in non-tumor and tumor pancreatic tissue (with no significant differences in dependency of these two settings) [495]; Nox3 expression was detected in the human fetal, but not in the adult kidney [487]. Juhasz et al. investigated the expression of Nox enzymes in various human cancer cell lines and, importantly, in ex vivo tumor tissues [113]. Nox3 mRNA was absent in all isolated tumor tissues derived from the colon, liver, lung, kidney, prostate, stomach, ovary, breast, testis and brain.
2.2. Expression of Nox3 in Cell Types
While detection in tissues or whole organs was and is a challenging task, the investigation of Nox3 protein expression in specific cell types, especially in cell lines, was extensively performed and delivered a broad catalogue of data addressing the topic where Nox3 is expressed and where it is absent. I should note that I do not share the opinion of cell lines of cancerous origin as “normal cells” for in vitro investigations as a sole line of evidence. Primary isolated ex vivo cells should be preferred; however, their isolation and cultivation remain difficult. Notably, most of the in vitro studies which addressed Nox3 have used cancer-derived cell lines like HepG2 (as hepatocyte model) or HEI-OI (as an inner ear hair cell model). Therefore, I listed only cancer cells under Section 2.2.7, which were clearly addressed as cancer cells in a context of tumor-associated research.
2.2.1. Nox3 in Cells of the Inner Ear
It is not surprising that the most detailed knowledge of cellular Nox3 expression accumulated around the cells of the inner ear and, as mentioned before, Nox3 is not the only Nox isoform expressed in the inner ear. Vlajkovis and colleagues first described a detailed overview of Nox isoform expression in the rat cochlea [479]. In detail, Nox1 mRNA was found in OHCs and Deiters’ cells; and Nox2 mRNA was expressed in OHCs and Claudius’ cells, Deiters’ cells and inner border cells, but was strongest in inner sulcus cells. Nox3 mRNA was strongly expressed in the inner sulcus cells but only weakly expressed in cells of the organ of Corti. Neither Nox2 nor Nox3 were detected in the lateral wall tissues or spiral ganglion neurons, which was confirmed for Nox3 protein expression by Zuhang et al. [496]. Nox4 was expressed in Hensen’s cells and inner sulcus cells but strongest in the blood vessels of the cochlear lateral wall and the Rosenthal’s canal. Duox1 was only weakly detected in sensory inner hair cells (IHCs) and supporting cells of the organ of Corti. Duox2 was strongly expressed in the inner sulcus cells and weakly expressed in the organ of Corti. The location of Nox3 in inner sulcus cells is especially notable, since these epithelial cells line the endolymphatic compartment where they clear the endolymph from cell debris, which occurs, for example, after severe acoustic trauma [497]. Accordingly, these cells play a pivotal role for cochlear repair and ion homeostasis [498].
Mohri and colleagues analyzed Nox3 expression in vivo using their Nox3-coupled dtTomato fluorescence system [462]. They described Nox3 expression in non-sensory epithelial cells of the endolymphatic sac and duct, of the vestibule and of the semicircular canals, but no Nox3 expression in the hair cells of maculae or ampullae. They also saw Nox3 expression after 7 days of birth in the root cells of the lateral cochlea wall. After 2 months, Deiters’ cells, Claudius’ cells and OHCs started to express Nox3. After 12 months, Nox3 expression further increased in Deiters’ cells, Claudius’ cells and outer and inner phalangeal border cells. IHCs showed Nox3 expression for the first time after 12 months. While these studies delivered excellent detailed information of Nox3 expression in rats and mice, so far, no detailed description of the cellular expression patterns of Nox3 has been conducted in the human inner ear.
2.2.2. Nox3 in Lung Cells
2.2.3. Nox3 in Liver Cells
The human liver cell line HepG2 naturally expresses Nox3 mRNA and protein [363,487,503,504], which is of critical importance, since this cell line serves as cellular model for most of the Nox3-related research on liver diseases (see Section 5.4.1 and Section 5.4.2). This is in notable contrast to the absence of Nox3 in the naïve murine fetal or adult liver [487].
2.2.4. Nox3 in Fibroblasts, Endothelial and Epithelial Cells in General
Ahmarani and colleagues expanded the list of cells in which Nox3 is naturally expressed [102]. They detected Nox3 in human endocardial endothelial cells (hEECs), human vaginal endothelial cells (hVECs) and vascular smooth muscle cells (hVSMCs). Interestingly, they reported a heterogeneous distribution in dependence of the cell type. In hEECs, Nox3 was found in clusters at the intracellular cell membranes, while in hVEVs and hVSMCs, it was equally distributed in intracellular membranes, including the nuclear membranes. Moreover, in all cell types, Nox3 was more abundant at the nuclear membranes compared to all intracellular membranes. Among the cell types, hVECS showed the strongest density of Nox3. Nox3 mRNA was further detected in late endothelial progenitor cells (EPC) together with Nox1, Nox2, Nox4 and Nox5 [505], in human nasal polyp-derived fibroblasts [506] and expressed as protein in the fibroblast-like cell line 3T3-l1 [485]. Notably, Zhang et al. found that Nox2 is the main ROS source in primary human dermal fibroblasts. All other Nox isoforms were at least expressed at the mRNA level, while Nox3 was not detectable at all [507]. Nox3 was also not detected in human umbilical endothelial cells (HUVECS) [508].
2.2.5. Nox3 in Cells of the Eye
Not many studies have investigated Nox3 as a possible ROS source in the eye. Brown et al. analyzed Nox enzymes in rabbit conjunctival fibroblast in the context of the fibrotic response [509]. The group found that Nox2, Nox4 and Nox5 and Nox3 mRNA were strongly expressed in this cell type, while Nox1 or the Duox enzymes were not detectable. Transforming growth factor (TGF)-β treatment, which was used as a stimulating factor in this study, did not stimulate the expression of Nox3; therefore, the role of Nox3 in this context was not further investigated. Furthermore, O Brian and colleagues could not detect Nox3 mRNA or protein expression in human corneal stromal cells [510]. As a result, if and how Nox3 might play a role during human eye diseases is completely unknown.
2.2.6. Nox3 in Cells of the Nervous System
Olguin-Alberne et al. investigated the involvement of Nox-derived ROS during the cell death of murine astrocytes induced by Staurosporin [511]. They could not detect Nox3 mRNA in astrocytes cultured for 2 weeks, while Nox1, Nox2 and Nox4 were detected. Nox3 absence in astrocytes was later confirmed by Reinehr et al. [460]. Oddly enough, Olguin-Alberne et al. further investigated Nox3-deficient mice and, not surprisingly, there was no difference between WT astrocytes and Nox3-deficient astrocytes. Notably, Acette et al. detected Nox3 mRNA expression in the oligodendrocyte cell line MO3-13 [512]. herefore, Nox3 should not be fully excluded from neuronal research.
2.2.7. Nox3 in Cancer Cells
During a previous analysis of ex vivo human cancer tissues, Nox3 was not detected. Further screening of various cancer cell lines, however, showed strong Nox3 mRNA and protein expression in the cell lines H28 (mesothelioma), H358 (bronchoalveolar) and A549 (adenocarcinoma). Nox3 was weakly expressed in H157 (squamous), H727 (carcinoid) and H838 (adenocarcinoma) [513]; in the cervix cancer cell line HeLa; in the lung cancer cell line GLC-82 [503]; in the human pancreatic cancer cell line Panc-1 [514], as well as in the human adenocarcinoma cancer cell lines MDA-MB-231, MDA-MB-468 and Hs578T [515]. Nox3 mRNA was also detected in the murine breast cancer line 4T1 [516].
In addition to these cancer cell lines, in which Nox3 was readily detectable, the majority of studies have described the absence of Nox3 in cancer cells, i.e., in the cancer cell lines H322 (bronchoalveolar), H520 (squamous), H1299 (large cell carcinoma), H2122 (adenocarcinoma) and HT29 (colon cancer) [513]; in the squamous carcinoma cell lines HSC-2, HSC-3, HSC-4, SAS and OSC-19 [517]; in the osteosarcoma cell lines HOS, MOS, MG-63, NOS-1 and HuO 9N2 [518]; in the malignant pleural mesothelioma cell lines ACC-MESO-1, ACC-MESO4, Y-MESO-8A, MSTO-2211H, NCI-H28, NCI-H290 and NCI-H2052 and the untransformed mesothelial cell line (Met-5A) [519]. Furthermore, no Nox3 expression was detected in the myeloid leukemia cell line K-562 [508] and, finally, in several other cancer cell lines (LS180, Caco2, LS174T, HT-29, PC-3, LNCap, DU145, MCF-7, BT474, ZR-75, MB-468, K562, HL-60, OVCAR-3, Skov-3, SK-Mel 5, A2058, HepG2, HEK293, TC-71) investigated in a broad screening study by Juhasz and colleagues [113].
2.2.8. Nox3 in Immune Cells
The first description of Nox3 expression in an immune cell type was made by van Buul et al., which detected Nox3 in the T-cell cancer line Jurkat [508]. Miyano and colleagues firstly showed that Nox3 is expressed and active in innate immune cells, namely the macrophage-like cancer cell line RAW 246.7 [275], which was confirmed in later studies [520,521]. In contrast, Nox3 mRNA was not detected in ex vivo Kupffer macrophages [460], and since no other ex vivo cell analysis was performed until now, it remains unclear if Nox3 belongs to the basic Nox repertoire of macrophages or if it is more part of the cancerous phenotype of RAW cells.
Feng and colleagues reported, for the first time, Nox3 expression on the mRNA and protein level in murine spleen B cells and in the human B cell line BAL17 [522], while Nox3 was not detected in the human B cell line Ramos [508]. Therefore, these findings remain somewhat contradictory.
Gaurav et al. investigated the role of eosinophils during allergic asthma [523] and detected high amounts of Nox2, Duox1 and Doux2 mRNA in human peripheral blood eosinophils, but only minor mRNA levels of Nox3 and Nox5.
Li et al. investigated the role of Nox enzymes in murine mast cells after UVA-induced Ca2+ fluctuations [524]. They detected strong mRNA expression of Nox2 and of its subunits p22phox, p47phox, p67phox, p40phox and Rac 1/2, as well as moderate expression Duox1 in the rat mast cell line RBL-2H3. All other Nox isoforms, including Nox3, were not detected.
The rarity of studies which have investigated Nox3 in immune cells in general and the partially contradicting findings of the already conducted studies clearly demonstrate that this topic represents a vast empty field for future research.
2.2.9. Nox3 in Other Cell Types
Nox enzymes were reported to be expressed in placental tissue before [525,526,527], but Polettini and colleagues dug deeper into this topic and analyzed human amniochorions, i.e., fetal membranes [494]. Expression of Nox2, Nox3 and Nox4 mRNA were detectable in healthy patients and in patients with either preterm premature rupture of membranes or preterm birth with intact membranes. Patients with chorioamnionitis were excluded from this investigation, since infiltrating immune cells would have confounded the obtained data. Nox1 and Nox5 mRNA was not detectable in the samples. Notably, the localization of Nox3 protein expression was present in both amnion and chorion cells.
Morimoto et al. described, in stably proliferating germline stem cells, strong expression of Nox1, while Nox3 and Nox4 were only weakly expressed [484,528]. However, dependent on the presence or absence of growth factors, the germline stem cells displayed a strongly fluctuating Nox isoform expression, with Nox3 as majorly expressed protein (see also Section 4.2). Issa et al. detected Nox3 mRNA and protein in the adipocyte cell line 3T3-1L [529]. Nox3 could not be detected in human induced pluripotent stem cell (iPSC)-derived CD34+ hematopoietic precursor cells [530], in immortalized primary human myometrial or in fibroid uterine cells [531].
2.3. Subcellular Locations of Nox3
While the expression either on the mRNA or the protein level was extensively described for Nox3 in tissues and cells in general, only a few studies have investigated the exact location of Nox3 in cells. For other Nox isoforms cellular locations were extensively investigated. Nox2 shows a rather restricted placement at the plasma membrane and at the membrane of phagosomes/endosomes, while Nox4 is broadly distributed over many intracellular structures [84,207], such as the nucleus [338] or the ER [218].
Uemaya and colleagues first described Nox3 localization at the plasma membrane, together with p22phox, p67phox and, as described before [369], NOXO1 in co-transfected HEK-293 cells [274]. The authors also suggested a mainly extracellular ROS production based on this observation. Nakano and colleagues also reported p22phox-dependent localization of Nox3 at the plasma membrane in co-expression systems with HEK-293 and CHO cell lines [217]. During their analysis of the general Nox3 expression in cells, Ahmarani and colleagues reported a heterogeneous distribution of Nox3 in dependence of the cell type [102]. In hEECs, Nox3 was found in clusters at intracellular cell membranes, while in hVEVs and hVSMCs, Nox3 was equally distributed in intracellular membranes including the nuclear membranes. Moreover, in all cell types, Nox3 was more abundant at the nuclear membranes compared to all intracellular membranes. The exact location of Nox3 for most of the cell types is still unclear and represents a highly interesting research field.
Taken together, a plethora of studies have investigated and reported Nox3 expression (some on the protein level, but most of them only on the mRNA expression level), in many organs, tissues (in vivo or ex vivo as explants) and cell types (as primary cells or cell lines). These findings revise the often-cited statement of Nox3 as “only expressed in the inner ear”. Sadly, studies which have investigated the exact subcellular location that obviously is dependent on the cell type, are scarce. Nevertheless, it seems that Nox3 might also exploit an interesting variability in terms of the subcellular location. Considering the vast amount of research, which was conducted so far to determine the structure (Section 1.2), induction/regulation (Section 3) and functions (Section 4) of Nox3, as well as possible therapeutically treatment options (Section 5) that target Nox3, it is highly surprising that nearly nothing is known about Nox3 in humans except for the expression in some organs [363,372,404,487,494]. No human material from organs, where Nox3 was clearly involved in pivotal functions in other species, such as rats and mice (e.g., from the inner ear, lung or liver, Section 4 and Section 5) was investigated, let alone that any treatment option, which targets Nox3 in a mouse or rat model went into a clinical trial so far. Thus, in the nearly complete lack of information for Nox3 in ex vivo human tissue lies a huge potential for new and fruitful research.
3. Activation and Regulation of Nox3
Considering the expression of Nox3 in various cell types and tissues, logically, each cell type of a specific organ or body compartment reacts differently to external and internal stimuli. These factors can be of endogenous origin, e.g., growth factors, cytokines and hypoxia or enter from the exterior, like pathogenic infection and physical or chemical hazards. When, how and if Nox3 is activated by these stimuli will be discussed in this section. A strict separation was made between the actual activation of the Nox3 enzyme, i.e., induced ROS production, and the regulatory processes, which also include modifications of Nox3 mRNA expression in any way [13,390]. Nox3 resembles Nox4 in terms of basal ROS production. Accordingly, an increase of Nox3 protein expression can correlate with higher ROS production and might influence the subsequent cellular events. However, this is not actually an induction of the enzymatic activity.
3.1. Activation of Nox3
Undoubtedly, the reader will swiftly notice that only a few studies have investigated and experimentally showed Nox3 activation, which is ROS production after cdefined stimuli. Most of the studies only analyzed mRNA or protein expression in this context, which both do not necessarily correlate with actual enzyme presence [530,532,533,534], activation and directed production of ROS. Therefore, when studies only performed expression analysis without providing clear evidence of Nox3 being the actual ROS source (e.g., via knock-out or knock-down) and/or without any ROS measurements at all, these studies will be discussed in Section 3.2, which summarizes the regulation of Nox3.
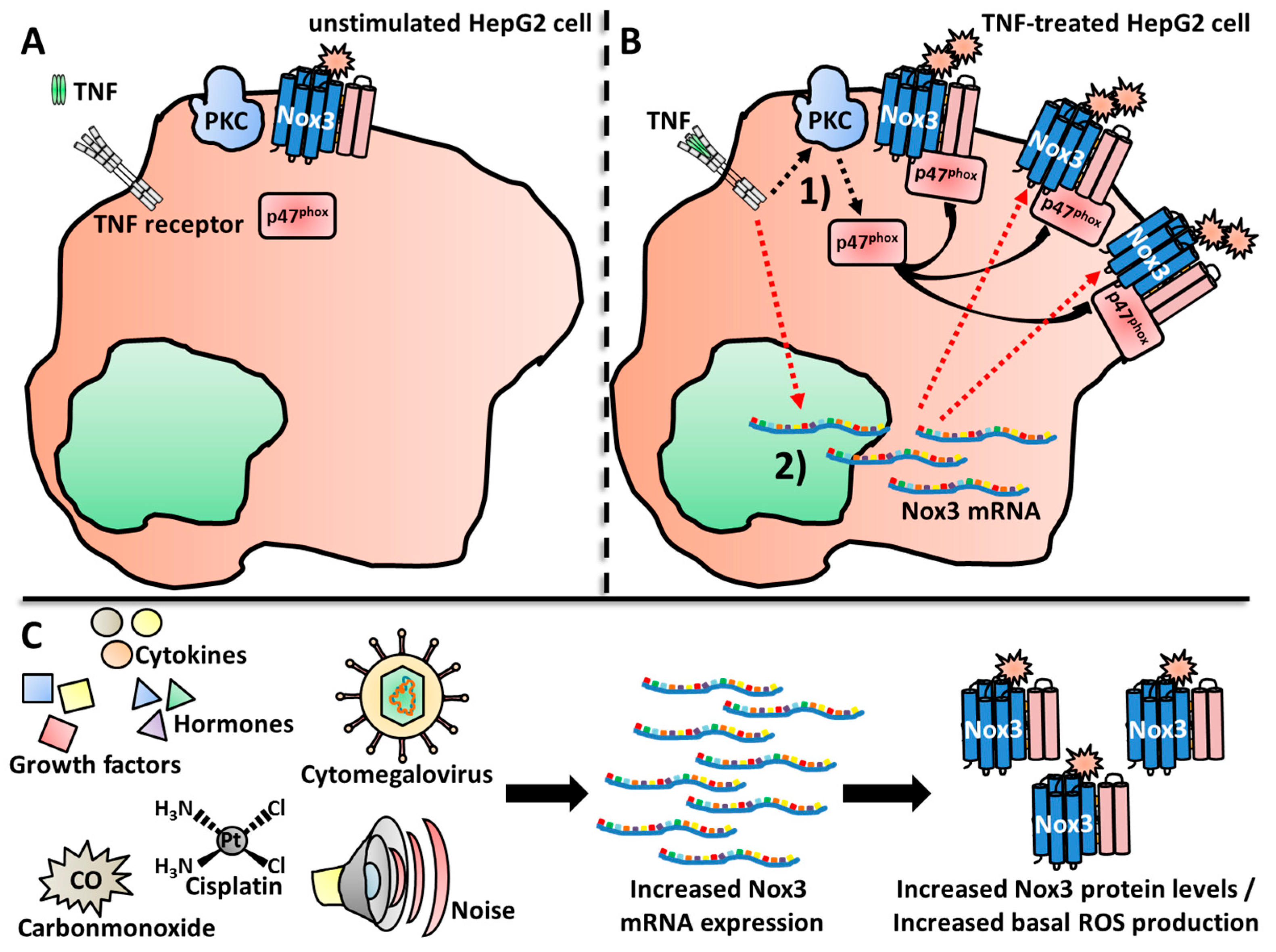
Nox3 was found to be activated by various stimuli involved in diseases progression, such as insulin in HepG2 cells [487], cisplatin treatment in the organ of Corti and the associated cells [355] and, for the first and only time so far, in B cells, via BCR-ligand triggering [522]. Li and colleagues described a direct activation of Nox3 after TNF treatment, which was mediated by PKC activation and subsequent p47phox translocation to Nox3 at the plasma membrane [396] (Figure 3A,B).
Similar to the knowledge about subcellular Nox3 location, also a clear scientific picture of Nox3 activation and ROS production, which does not always correlate with increased expression, is sadly very low. Considering the many discovered organs, tissues and cells in which Nox3 is expressed aside from the inner ear, a lot of interesting research potential lies in the question by which stimuli Nox3-derived ROS production is activated, especially in ex vivo cells.
3.2. Regulation of Nox3
3.2.1. Nox3 Regulation on the Expression Level
As mentioned before, most of the studies that investigated Nox3, especially in the context of in vitro or in vivo functions, only analyzed mRNA expression of Nox3. First of all, mRNA content does not necessarily reflect the presence of the build protein [530,532,533,534], making the few studies that took the extra work of depicting the Nox3 protein expression much more conclusive. Secondly, many studies did not confirm Nox3 as precisely responsible for the observed effects, since no genetic evidence, i.e., by knock-out or knock-down, was performed. Nevertheless, regulation of mRNA and protein expression is an important factor of Nox3-mediated ROS production, which will be summarized in the following sections.
Up-Regulation Nox3 on the Expression Level
A number of endogenous factors such as cytokines, growth factors, hormones or altered body homeostasis lead to the up-regulation Nox3 expression (Figure 3C). In germline stem (GS) cells, Nox3 protein expression was up-regulated after stimulation with the cytokines glial cell line-derived neurotrophic factor (GDNF) and fibroblast growth factor 2 (FGF2) [484]. Issa et al. described an increase of Nox3 protein after three hours of TGF-β treatment in the adipocyte line 3T3-1L [529]. Similarly, Yasuoka and colleagues detected an increase in Nox3 mRNA after TGF-β or integrin beta-5 (IGBT-5) treatment in primary human lung fibroblasts [502]. Nox3 mRNA expression was increased in the murine breast cancer line 4T1 after isolation from an established tumor setting in mice [516]. These animals were additionally treated with TWS119, a substance that leads to glycogen synthase kinase-3 β (GSK-3β) phosphorylation. GSK-3β is a protein kinase with a high correlation to cancer transformation [535,536]. TWS119 treatment led to a further up-regulation of Nox3 mRNA in the isolated 4T1 tumor cells.
Insulin treatment increased Nox3 protein levels in HepG2 cells, a commonly used cell line for investigation of liver diseases. This phenomenon was also observed 3T3-L1 cells and white adipose tissue in mice [485]. Palmitate treatment also increases Nox3 protein levels in an adipose animal model [537]. Michihara et al. also found that Nox3 mRNA and protein levels were increased in the brain of hypertensive rats [489]. Adipositas, as well as hypertension, can contribute to cardiovascular diseases and the role of Nox3 in this context will be discussed in Section 5.4.
Li and colleagues reported Nox3 mRNA up-regulation after treatment of HepG2 cells with the pro-inflammatory cytokine TNF [396] (Figure 3A,B). Kathanal et al. observed Nox3 mRNA up-regulation after treatment with the Gram-negative bacterial cell wall component lipopolysaccharide (LPS) [521]. Both findings suggest a possible role for Nox3 during infection and inflammation.
Many exogenous factors, most of them physical or chemical inducers of inflammation, were described to increase Nox3 mRNA and protein levels. The most prominent substance is probably the anti-cancer drug cisplatin, which induces toxic damage by many correlative events that all increase the inflammatory profile of the inner ear, especially in the cochlea [538,539]. Accordingly, several studies have described an increase of Nox3 mRNA [477,540] or protein [462,476,541,542] after cisplatin treatment (Figure 3C).
Exposure to physical hazards also influences Nox3 expression. Carbone monoxide (CO) exposure (3000 parts per million [ppm]) induced Nox3 mRNA expression in the rat striatum [543], and Wang et al. saw a strong increase of Nox3 protein after 1 hour of heavy ion irradiation (1–4 gray) of HeLa, HepG2 and GLC-82 cells [503]. Habashy and colleagues investigated the oxidant and antioxidant responses in chicken livers after mild heat stress (35 °C) [493]. The group detected a basal mRNA expression of Nox3 in liver tissue, which was up-regulated after 1 and 12 days of applied heat stress. Finally, as reported by various studies [462,476,478,544], noise exposure leads to an increase in Nox3 mRNA and protein levels in the cochlea (Figure 3C).
Chemical exposure can also lead to altered Nox3 expression. Kim et al. described an up-regulation of Nox3 mRNA after treatment with endosulfan [545], a widely used pesticide that is associated with immune response dysregulation [546,547]. Ye and colleagues investigated the interplay between oxidative and anti-oxidative responses in rat kidney after phenol-induced kidney injury [492] and detected an increase of Nox2, Nox3, p22phox and p47phox mRNA in isolated brain nuclei. Kim et al. detected a protein up-regulation of Nox3 after mono sodium urate crystal treatment in RAW cells [520].
Some bioactive, substances isolated from medical plants, such as Brevilin A [548] or Genipin [521], also induced Nox3 mRNA and/or protein up-regulation.
Down-Regulation or No Effect on Nox3 Expression
Owens and colleagues noted a correlation of Nox3 mRNA levels and the Rieske-Iron-Sulfur protein (RISP) in the Complex III of the mitochondrial respiratory chain. After RISP knock-down in various breast cancer cell lines they detected a decrease in Nox3 mRNA [550].
In contrast to other studies [462,476,478,544], Vlajkovic et al. observed that Nox3 expression is down-regulated in the rat cochlea after noise exposure (100–110 decibels [dB]). More precisely, they showed that Nox3, but not Nox2, is down-regulated in the inner sulcus cell region [479]. Li and colleagues detected Nox3 mRNA in late EPCs together with Nox1, Nox2, Nox4 and Nox5. Angiotensin-II treatment resulted in a strong increase in the mRNA expression of Nox2, Nox4 and Nox5, but no expression changes were detected for Nox3 [505]. Finally, the antioxidative substances Simvastatin and curcumin reduced Nox3 mRNA levels [504].
Nox3 Regulation via Other Factors
Qian et al. showed a regulatory role of nitric oxide on the direct enzymatic activity of Nox3 [551]. In COS-7 cells, which were co-transfected with Nox3, as well as NOXO1 and NOXA1, the addition of the NO donator DETA-NONOate inhibited Nox3-mediated superoxide production in a dose-dependent manner. The group of Kiss et al. reported dependency of PKC during p47phox-mediated activation of Nox3 [464] (Figure 3A,B), confirming the findings of Li and colleagues [396].
4. Functions of Nox3
It is not surprising that Nox3-derived ROS, in regard to Nox3 expression in many different tissues and cell types, fulfill various functions in the body. In this section, the beneficial functions of Nox3-derived ROS will be discussed, while the causes of ROS overproduction or ROS production in the wrong locations, which lead do various diseases, will be summarized in Section 5.
4.1. Signaling Functions of Nox3
Remarkably, three very convincing and nicely conducted studies, which investigated Nox3-derived ROS in cellular signaling processes, all investigated the signaling functions of ROS in the context of diabetic liver diseases. The fourth study investigated several cancer cell lines, and these four studies are, so far, the only research conducted for Nox3-derived ROS in the context of signaling pathway modifications.
Previous studies have reported a swift increase of H2O2 production after insulin treatment [552,553,554] in liver cells and Carnesecchi et al. investigated possible ROS sources involved in this context in the hepatocyte-like cancer cell line HepG2 [487]. The group measured a basal H2O2 production without any stimulus and a robust increase (28–40%) of H2O2 production after treatment with 100 nM insulin. Down-regulation of Nox3 by siRNA nicely solidified Nox3 as the source of ROS, since Nox3 knock-down led to the abolishment of H2O2 production. After insulin treatment, HepG2 cells showed increased phosphorylation of the signaling kinases ERK1/2 and Akt. While Akt phosphorylation was not altered after Nox3 knock-down, phosphorylation of ERK1/2 was decreased through the whole time course of insulin treatment. Insulin-induced ERK1/2 activation leads to Vascular Endothelial Growth Factor (VEGF)-A mRNA and protein expression in HepG2 cells and keratinocytes [555,556,557]. The group further investigated this topic in the context of Nox3-derived ROS production. Indeed, an increase in VEGF-A mRNA and protein expression after insulin treatment was detected, which was strongly decreased after Nox3 knock-down. Notably, the exogenous addition of H2O2 rescued this effect, thus connecting Nox3, H2O2 and VEGF-A expression. Finally, the group observed a decreased binding activity of the transcription factor Specific protein 1 (Sp1) [558], which plays a central role in VEGF-A expression [559,560]. This study is one of the few examples during the time course of Nox3-focused research, which clearly shows a consistent line of evidence for Nox3-derived ROS involvement. All critical parameters for Nox-related research were investigated, i.e., the ROS production-inducing stimulus (insulin), confirmation of the ROS source by genetic evidence (via siRNA-mediated knock-down); furthermore, an actual decrease in ROS production confirmed by ROS measurements (same stimulus, same cell type), a connection of the produced ROS and the regulated signaling pathways (ERK and Akt signaling) and finally the influenced cellular outcomes (transcription factor regulation) (Figure 4A).
A study from Li and colleagues investigated the effect of Nox3-derived ROS on the glycogen levels in HepG2 cells [396]. Insulin resistance is a key feature of type 2 diabetes and several studies have documented the involvement of elevated ROS production in insulin resistant cells and tissues [561], which lead to disturbed signaling pathways that regulate the intracellular glycogen levels [562,563]. The group focused on TNF-induced signaling as an inhibiting factor of insulin signaling [564,565]. Wistar rats were fed with a high-fat diet (HFD) for 12 weeks to induce insulin resistance. This was accompanied by increased TNF plasma levels, decreased hepatic glycogen levels and enhanced hepatic ROS production. To link these correlative data sets, the researchers switched to an in vitro model. HepG2 cells were treated with TNF (4–6 ng/mL for 4 days) and showed decreased intracellular glycogen levels and enhanced total cellular ROS production. qRT-PCR analysis revealed Nox3 as the only expressed Nox isoform on the mRNA level in HepG2 cells, together with p22phox, p47phox, p67phox and Rac1. TNF treatment increased the mRNA expression of Nox3, which was also noticed in the liver in vivo after a HFD. Since not NOXO1, but and only p47phox as possible regulatory subunit of Nox3 was detected in HepG2 cells, the group tested the previously suggested involvement of PKC during Nox3 activation [464]. Indeed, the PKC inhibitor hypericin abolished TNF-induced ROS production. Since PKC signaling induces p47phox translocation from the cytosol to the plasma membrane, this was also investigated. Fluorescence microscopic and Western blot analysis of membrane protein extractions confirmed the translocation of p47phox to the plasma membrane after TNF stimulation. These data show that Nox3-mediated ROS-production is increased by two independent mechanisms in HepG2 after TNF treatment: Firstly, the mRNA expression of Nox3 is increased after TNF treatment, and secondly Nox3, is activated via TNF-mediated PKC activation and p47phox translocation (Figure 3B). The group next confirmed via siRNA-mediated knock-down Nox3 as the sole ROS source after TNF stimulation in these cells. Glycogen levels also remained stable after knock-down of Nox3 in contrast to not transfected cells, showing the involvement of Nox3-meditated ROS in this process. They further investigated the C-Jun-N-terminal Kinase 1/2 (JNK1/2) signaling pathway as the link between ROS and the observed glycogen decrease, since this pathway is not only modulated by ROS [566] but is also involved in insulin sensitivity in mice [567,568]. TNF treatment resulted in phosphorylation of JNK1/2, which could be reversed via Nox3 knock-down. This nice publication identified Nox3 as sole ROS source in TNF–stimulated HepG2 cells, the JNK-pathway as ROS-mediated target, the involvement of ROS in cellular insulin resistance and a possible interplay of TNF, PKC and p47phox-mediated activation of Nox3. Furthermore, these findings reveal a clear contrast to the regulation of Nox3 in the inner ear, where Nox3 is only activated via NOXO1 [355,372]. Finally, the group unraveled two very distinct possibilities to regulate Nox3-derived ROS production, i.e., on the expression level or by direct signaling-mediated activation.
A follow-up study from the same lab further focused on the role of free fatty acids (FFA) during insulin resistance and the role of Nox3 in this context [537]. It was previously shown that elevated ROS levels in general are correlated to insulin resistance [569,570] and an involvement of FFA was suggested [571,572,573,574]. The group saw elevated insulin, glycohemoglobin and FFA levels in plasma, as well as decreased hepatic glycogen levels and increased hepatic ROS levels in leptin-deficient mice (db/db mice). This mouse strain is a commonly used model for type 2 diabetes investigations [575]. In vitro studies with HepG2 cells revealed an increase of gluconeogenesis and an impaired cellular glycogen content after palmitate treatment, which mimics insulin resistance in vivo. In this context also increased total cellular ROS levels were observed. A previous study documented the expression of Nox3, p22phox, p67phox, p47phox and Rac1 in HepG2 cells, but not of other Nox isoforms or subunits [396]. Indeed, expression of Nox3 was up-regulated after palmitate treatment in HepG2 cells and in livers of db/db mice, while Nox1, Nox2, Nox4 or Nox5 were not expressed. Knock-down of Nox3 via siRNA in HepG2 cells reduced Nox3 mRNA expression and ROS production in untreated and palmitate-treated cells, nicely establishing Nox3 as the ROS-producing enzyme in this context. Previous studies have discovered critical roles of the MAPKs, JNK1/2 [396,576,577] and p38 [578] during insulin resistance. ROS-mediated modifications of these pathways [566,579], especially during insulin resistance [580] were also suggested. Indeed, palmitate treatment led to increased JNK1/2 and p38 phosphorylation in HepG2 cells. The activation of these two kinases subsequently led to phosphorylation of the kinases Akt, glycogen synthase kinase-3 (GSK3) and the transcription factor Forkhead box protein O1 (FoxO1), finally resulting in increased gluconeogenesis and reduced glycogen levels. Knock-down of Nox3 reduced phosphorylation of JNK1/2 and p38 as well as suppressed gluconeogenesis (Figure 4B). Again, this is an example of a nice and convincing study in which both, the exact ROS source and the mode of action were clearly described in a cellular system. However, in contrast to the lab’s previous study [396], a direct link of Nox3-derived ROS in vivo was unfortunately not found.
Maletter et al. investigated a completely different topic, i.e., the role of Nox3-derived ROS during cell death signaling. The group focused on the effects of the CD95/Fas ligand CD95L on the human adenocarcinoma cancer cell lines MDA-MB-231, MDA-MB-468 and Hs578T [515]. CD95L treatment, previously cleaved by a metalloproteinase [581,582,583], resulted in a switch from an apoptotic [584] to a pro-motile metastatic phenotype [585]. Binding of cleaved CD95L to the Fas receptor led to subsequent Ca2+ release mediated by the transcription factor c-yes [586,587]. The elevated Ca2+ levels activated PI3K [585,588] and induced total cellular ROS production. Although the used cancer cell lines expressed Nox2, Nox3 and Nox4, only Nox3 was recruited to the membrane-located signaling platform, which formed after CD95L treatment. Silencing of Nox3 by siRNA abrogated Ca2+ release and cell migration in CD95L-treated cells. Unfortunately, no ROS measurements were performed in Nox3-silenced cells. Therefore, no evidence in this otherwise convincing study for a direct link between Nox3-derived ROS and the observed signaling effects in this context could be made.
4.2. Functions of Nox3 in Cell Differentiation
Sasaki and colleagues firstly investigated the involvement of Nox3-derived ROS during cell differentiation [589]. They used RAW246.7 cells to investigate a possible role of Nox-derived ROS during osteoclast differentiation. Previous studies for this topic were contradictory. Osteoclasts express Nox2 [590]. However, Nox2-defcient osteoclasts still produce O2−, and Nox2-deficient mice show no abnormalities in their bone structure [86]. As redundant ROS sources, Nox4 in differentiated osteoclasts [86,591] and Nox1 in osteoclast precursors [592] were suggested. Notably, in Nox1-deficient [125,593] and Nox3-deficient mice [370], no bone abnormalities occur. Sasaki et al. detected small amounts of Nox3 mRNA in RAW246.7 cells (0.001% in comparison to the highly expressed Nox2), while NOXA1 mRNA could not be detected. After treatment with Receptor Activator of NF-κB Ligand (RANKL), which is an osteoclast differentiation factor, Nox2 mRNA expression was strongly down-regulated, while Nox3 expression only slightly decreased. Nox1 expression on the other hand was strongly increased. Notably, expression of NOXO1, an important enhancer of ROS production of both Nox1 and Nox3, decreased. Accordingly, the O2− production was reduced but did not vanish completely. This suggests a flexible adaptive switch of Nox enzymes for ROS production during differentiation of osteoclasts. Unfortunately, there was no direct evidence of Nox3-derived ROS during this process, since only p22phox or p67phox were down-regulated via siRNA.
Several lines of evidence suggest that ROS in general are necessary for the differentiation of cells of the nervous system [594,595,596], which was shown in detail for the PC12 cell line [597], glia cells [598], neuroblastoma cells [599] and oligodendrocytes [600]. Previous studies, which investigated a possible role for Nox-derived ROS during oligodendrocyte differentiation [598], only used the very unspecific inhibitors apocyanin [601,602,603] or DPI [7,13,604,605,606]. No genetic evidence (knock-out or knock-down) was provided for Nox enzyme involvement [79], so this issue has remained unresolved. Acette and colleagues further investigated this issue in the oligodendrocyte-like human cancer cell line MO3-13 [512]. They found that MO3-13 cells express Duox1, Duox2, Nox5 and Nox3. Nox3 knock-down reduced the expression of Myelin Basic Protein and the nuclear factor Olig-2, which are two important markers of oligodendrocyte differentiation [607,608,609]. Unfortunately, no other cell responses, especially the ROS production, were analyzed after knock-down of Nox3. Hence, again, a direct link of Nox3-derived ROS and the expression of differentiation markers in oligodendrocytes could not be made.
Morimoto and colleagues investigated in a nicely conducted study a putative function for Nox3-derived ROS during proliferation of murine GS cells in vitro and in vivo [528]. In a previous study, the group identified Nox1 as the majorly expressed ROS source in stably-growing GS cells. Cellular knock-down of Nox1 in cells or a full body knock-out of Nox1 in mice led to a reduced proliferation activity of GS cells [484]. In this follow-up study, the group reported a strongly dynamic expression pattern of Nox enzymes in dependency on their proliferation status. Stably proliferating GS cells strongly expressed Nox1, while Nox3 and Nox4 were only weakly expressed, as shown previously [484]. Without any proliferation-stimulating factors, i.e., the growth factors FGF2 and GDNF, Nox1, Nox2 and Nox3 expression was strongly up-regulated. Notably, when FGF2 and GDNF were added to actively proliferating GS cells, Nox1 and Nox2 mRNA levels were down-regulated and only Nox3 mRNA was up-regulated. The expression of Nox3 was modulated by the MAPK and PI3K signaling pathways, since chemical inhibition of both pathways led to a strong down-regulation of Nox3 mRNA expression. Knock-down of Nox3 resulted in decreased ROS production and reduced gene expression of ld4, etv5, Nanos3, Neurig3, Blc6b, Ztb16, Cdkn1a, CCnd2 and Ccnd3. An increase of gene expression was detected for Ccnd1, Sohlh1 and CDkn1b. Nox3 knock-down finally led to apoptotic cell death and a defect in active proliferation. The group furthermore expanded their findings by analyzing the testes from 7- to 10-day-old mice. Isolated testicular cells were treated with small hairpin (sh)RNA against Nox3 and subsequently showed increased self-renewal activity in comparison to the control cells. This study firstly described the presence of Nox3 in testesand analyzed its contribution to the self-renewal capability after cytokine stimulation as well as during GS cell maintenance under non-stimulated conditions. Sadly, a direct mechanistic link between Nox3- or Nox1-derived ROS and the proliferation capacities was not investigated.
Mazzonetto and colleagues investigated of the interplay of Nox3 and Sonic Hedgehog (SHH)-mediated signaling during the development of granule cell precursor differentiation [610]. Purkinje cells produce and secrete SHH [611], a protein that is the major proliferatory stimulus for granule cell precursors [611,612,613,614]. After binding to the SHH receptor called Patched, the intracellular signaling pathway is activated and leads to induction of proliferation [615]. Dysfunction of this pathway and the subsequent disturbance of cerebellar neurons during development can cause ataxia, which manifests in neurological malfunction and motor discoordination [616]. Mazzonetto et al. characterized a BALB/c mutant mouse line that showed a phenotype which resembled ataxia and was established in a previous mutagenesis screening [617]. The most obvious phenotype of the mutant animals was a lack of motor coordination. This phenotype did not get worse with age indicating a developmental defect. Linkage analysis revealed the location of the mutation in chromosome 17, which is the syntenic region of the human chromosome 6. Several other studies have located mutations linked to Nox3 in chromosome 6 in patients with developmental disturbances and hearing loss [618]. The candidate genes present in the mutated regions were Tiam2, Tfb1m, Cldn20 and NOX3. After applying a singular nucleotide variant filtering and further data analysis, only one singular nucleotide variant remained in exon 3 of the NOX3 gene. The mouse line was subsequently named NOX3eqlb after the newly discovered allele. NOX3eqlb mice showed an unaltered NOX3 gene expression, while the NOX1 gene was strongly expressed in comparison to WT mice. The group isolated cerebellar and neural stem (NS) cells and observed a slight increase in total cellular ROS in NOX3eqlb-derived cells after 7 days and much less ROS production after 12 days in cell culture. The cerebellum of NOX3eqlb mice showed no abnormalities, but the group reported a thicker external granular layer, a disorganized Purkinje cell monolayer and more Bromodeoxyuridine (BrdU)-positive cells indicating increased proliferation. Organotypic in vitro cultures of cerebella and granular precursor proliferator cells isolated from in NOX3eqlb mutant mice showed a higher proliferation rate in comparison to WT mice. Cultured neurospheres from isolated NS increased in size much earlier when derived from NOX3eqlb mice but normalized at day 10 to a similar degree as in WT mice. Other organs such as the heart, liver, muscles, kidney and other brain regions showed no increased proliferative activity. Microarray analysis detected 116 up-regulated and 40 down-regulated genes at day 6 after birth. At day 15, 64 genes were up- and 5 were down-regulated. All of these genes were involved in proliferation and cell growth, e.g., Cdkn2a, Cd133, CCnb1, Cdk1, Rb1, Cdc25, Akt1 and Sox2 [619]. Increased levels of SHH protein, the main mitogenic driver in this context, was detected in the cerebella of NOX3eqlb mice. Additionally, increased expression of genes down-stream of the SHH pathway (Ccnd1 and Gli1, 2, 3 [620,621,622]) was detected. Since SHH-mediated signaling is activated via ROS [623], a connective mechanism was suggested. Unfortunately, in this otherwise excellent study, only the unspecific inhibitor apocyanin was used, and no direct evidence was given for the involvement of Nox3-derived ROS in this context.
Feng and colleagues investigated, for the first time, the presence and function of Nox3 in B cells [522]. Upon exposure to antigens, B cells undergo proliferation and activation mediated by a complex signaling cascade [624,625,626,627]. The involvement of ROS during cellular signaling was established in various immune cells before [4,80,145,146], including B cells [628,629,630,631,632]. While mitochondria [632] and Nox2 [629] were identified as activated ROS source in B cells, other Nox enzymes were not investigated. Notably Nox2-deficient B cells normally proliferate, which suggests that ROS produced by other Nox isoforms might be more important in this process [629]. Feng et al. analyzed this topic and found that in ex vivo murine splenic B cells and in the B cell line BAL-17 the mRNA expressions of the NOX1, NOX3, DUOX2, NOXA1 and NOXO1 genes were up-regulated after B cell receptor activation. Nox4 and Duox1 mRNA could not be detected. The group measured no total cellular ROS production in Nox2- or DuoxA1/2-deficient B cells in the early phase (1 h), but prolonged ROS production at later time points (4–6 h). Additionally, no disturbance in proliferation was reported, suggesting no role for the early ROS production mediated by Nox2 and Duox2. Interestingly, prolonged ROS production and proliferation in B cells was abolished in p22phox- or Nox3-deficient BAL-17 cells, but not in Nox1-deficient cells. Via the CRISPR/Cas9 knock-out system, the group nicely identified Nox3 as responsible ROS source. However, instead of using the nicely established knock-out cell lines, the group only used the globally working ROS scavenger NAC to investigate the role of ROS in the signaling cascade, which mediates B cell activation. Therefore, a direct mechanistic link between Nox3-derived ROS and the signaling cascade necessary for B cell activation was not demonstrated.
Park et al. investigated the role of 8-hydroxy-2′-deoxyguanosine (8-OHdG), an oxidatively modified DNA base and biomarker of oxidative distress [179,633,634,635], and its paradoxical role as exogenous anti-inflammatory and anti-oxidative component [636]. Treatment of human pancreas cancer cell line (Panc-1) cells with 8-OhdG resulted in decreased total cellular ROS production and a reduction of Nox1, Nox2 and Nox3 mRNA expression. [514]. However, none of these Nox isoforms were confirmed as involved ROS sources via knock-down or knock-out experiments.
Al-Sabbagh et al. investigated the functions of Nox enzymes during decidualization, a process which summarizes the cellular changes for pregnancy preparation of human endometrial stromal cells [637]. After 8-bromo-cAMP stimulation, which induces signaling events that lead to decidualization, the group observed a p22phox-dependent and Rac1-independent response of stromal cells. Despite the fact that Rac1 is not completely necessary for full Nox3-mediated ROS production, they excluded Nox3 as ROS source and focused on Nox4, which was confirmed as major ROS source via siRNA knock-down experiments. Unfortunately, since no broad experimental screening via qRT-PCR or Western blot experiments were conducted, it remains elusive if and how Nox3 is expressed or if Nox3-derived ROS fulfill other important functions in human endometrial stromal cells.
4.3. Functions of Nox3 in the Inner Ear
Before Nox3 can be put in the context of ROS-associated benefits or malfunctions for the inner ear, it should be mentioned that ROS and their subsequent effects on the inner ear were described before the discovery of Nox3 [469,472]. Moreover, as mentioned earlier (Section 2), Nox3 is also not the only Nox isoform expressed in the inner ear [479]. It is also noteworthy that the striking overlap of NOXO1 and Nox3 mRNA expression patterns, as well as the observed similar phenotypes of NOXO1- or Nox3-deficient animals led to a synonymous use for NOXO1 and Nox3 deficiency in some occasions [355,370,464].
4.3.1. Functions of Nox3 in the Vestibular System
While the effects of Nox3 deficiency on otoconial development were described in vivo by Paffenholz and colleges [370], it took 2 years until an indirect hint for the precise mechanism of Nox3-derived ROS in this process was discovered by Kiss et al. [464]. The group analyzed a spontaneously emerged mouse mutant line with severe balance deficits, named “head slant” (hslt), described by Gagnon and colleagues in 2013. (short report available as PDF on the Jackson Laboratory Website, https://www.informatics.jax.org/downloads/Reference_texts/J86035.pdf, accessed on 2 February 2024). Kiss et al. further characterized this mutant mouse line and confirmed strong balance and orientation deficits, while the hearing capacities were not altered. All of the hslt mutant mice were homozygous for a mutant NOXO1 gene allele (therefore named NOXO1hslt). The group elegantly showed via a transgenic rescue with a functional NOXO1 gene allele that the dysfunctional NOXO1 subunit is indeed responsible for this severe phenotype, since all transgenically rescued animals showed normal gravity and balance perception similar to wild-type (WT) animals. Strikingly, in all NOXO1hslt animals, a complete absence of CaCO3 and otoconia in the inner ear was reported. Instead of functional otoconia, otoconia-like unstructured conglomerates were spotted directly above the sensory hair cells. Other compartments of the inner ear, like the sensory epithelia, the tectorial membrane of the organ of Corti and the ampullae of semicircular canals, were all intact. The group also reported a broad expression of NOXO1 mRNA in the sensory and nonsensory epithelial cell layers of the saccule, in the ampullae of semicircular canals, in the epithelium lining of the scala media and in spiral ganglion neurons. In vitro expression of the NOXO1hslt protein in HEK293 cells resulted in an abolished Nox3-mediated ROS production in comparison to the cells, which expressed the NOXO1wt protein. This was also observed after co-transfection of NOXA1 and NOXO1hslt, while the co-expression of NOXA1 and NOXO1wt showed maximal ROS production via Nox3. This study firstly showed a direct and not a correlative connection between NOXO1, Nox3-derived ROS and otoconia formation. The group suggested changes in the OC-90/95 protein itself or during the delivery of OC-90/95 to the forming otoconia. These suggestions supported the observations of Paffenholz et al., which showed that H2O2 leads to disulfide linkage and conformational changes in the secreted OC-90/95 protein [370]. During the period in which they conducted their study, Paffenholz and colleagues hypothesized that Nox3-derived ROS might lead to peroxidation of the lipid vesicles in which the globular substance for otoconia formation is stored. The lipid vesicle peroxidation then could lead to Ca2+ release on the one hand and accessibility of OC-90/95 to the globular substances on the other hand. Also, the involvement of Nox3-derived ROS and their influence on CaCO3 concentrations at the otoconia-forming regions of the vestibular system was suggested as mechanism by a later study [462]. Although the impressive phenotype of Nox3-deficient animals regarding otoconia was intensively investigated and described in vivo, the study by Kiss and colleagues firstly provided a deepened mechanistic explanation how the Nox3-derived ROS might contribute to otoconia formation. The suggested theory of Paffenholz et al. and Kiss et al. of ROS-mediated disulfide-linkage of OC-90/95 and its important effect on otoconial formation should be proven right, however 15 years later [466].
This follow-up study after nearly 15 years was conducted by Xu and colleagues, who firstly investigated the suggested direct mechanistic interplay of Nox3-derived ROS and OC-90/95 in vitro and in vivo [466]. Considering the vast amount of time which passed from the first discovery of the correlative presence of Nox3 and otoconia [355,370], it is notable and laudable that at least one study has investigated this research issue directly. The group therefore generated OC-90/Nox3 double knock-out mice via cross breeding of previously described mouse strains [370,438] and compared WT, single knock-out and double knock-out animals with various experimental approaches. OC-90 and Nox3-deficient animals stayed for a shorter period of time on the rotarod, a testing device for balancing [638], in comparison to WT animals. OC-90/Nox3-deficient animals endured for an even shorter period. Notably, while WT mice adapted from experiment to experiment, all other mouse strains did not adapt, indicating a permanent deficit of balance functions. Measurements of the vestibular evoked potentials, which reflect the activity of the vestibular nerve, were completely absent in OC-90/Nox3-deficient mice. The vestibular nerve and its information relay to the subsequent neuronal network depend on the proper function of the utricle and saccule, where otoconia are located. The absence of any vestibular-evoked potential in the double-deficient animals suggested a severe impairment of gravitational reception [409,461]. Accordingly, in Nox3- and OC-90/Nox3-deficient animals, otoconial structures were completely absent as depicted via scanning electron microscopy. Double-deficient animals also displayed loss of hair cell bundles. This loss was not present from birth but appeared after 3 months. The group used a co-expression system in the NIH/3T3 cell line cultured under extracellular calcification conditions in vitro. Strikingly, transfection with an empty vector or expression of Nox3 alone induced no or minor calcification, respectively. Sole expression of OC-90 induced a stronger formation of calcified nodules on the cell surfaces. Finally, the co-expression of OC-90 and Nox3 together resulted in the strongest calcification process. This simple but nicely conducted study is the one and only research performed so far, which clearly showed the importance of Nox3 for the process of otoconia formation directly and not as a correlative observation. The only experiments, which would have added important information to these findings, are (i) ROS measurements, to prove that the transfected Nox3 indeed produces ROS into the extracellular milieu, and (ii) the addition of a ROS scavenger into the medium, e.g., N-acetyl cysteine (NAC), to prove that the Nox3-derived ROS are the potentiating factor of OC-90-mediated otoconial formation. OC-90 has a remarkable number of cysteine residues [435,639,640,641], and Xu et al. suggested a mechanism in which disulfide bond-dependent multimer formation of OC-90 in the endolymph, which is otherwise a soluble monomer, then serves as scaffolding platform for otoconial growth (Figure 4C). So far, this is the most reasonable mechanism of Nox3-derived ROS for otoconial formation, and future studies will hopefully further investigate this important topic that is still not fully resolved.
Jones et al. characterized a number of mouse strains, which all lack otoconia, namely head tlt, het-Nox3, tilted and tlt-Otop1 [642]. Otoconia-deficient mice failed to swim and orientate like described before [370,464,466]. The group measured spontaneous activity of the vestibular primary afferents, which innervate the maculae, even in the absence of otoconia [643]. The vestibular primary afferents further displayed higher discharge rates in comparison to WT animals. These data suggest that in absence of stimulation due to otoconia loss the resting activity in macular primary afferents and the ribbon synapses present in hair cells of otoconia-deficient mice are still functional. Basaldella and colleagues further investigated the interplay of the vestibular and proprioceptive system and body balance in an impressive study [644]. For this purpose, the group used Nox3-deficient mice as in vivo model system. As mentioned above, these mice are devoid of otoconia in the utricle and the saccule of the inner ear [370], which leads to defects in perception of gravity and linear acceleration, while the auditory system remains intact. The research group analyzed the communication with the lateral vestibular nucleus and other motor neuron pools. The lateral vestibular nucleus is one of the four major nuclei that form the vestibular complex. This complex is essential for maintaining the head position and clear vision during movement [645,646]. In this context, the group reported that the lateral vestibular nuclei (LVe) neurons maintain the synaptic input to motor neurons even in Nox3-deficient mice, in which otoconia are absent and vestibular signaling is non-functional. They also reported a higher synaptic input density, but no differences in the synaptic inputs mediated from the LVe neurons to other motor neuron pools. This study demonstrated that genetic distortion via Nox3 deficiency of vestibular input channels or silencing of the synaptic output of vestibular neurons leads to comparable connectivity defects between LVe neurons and flexor motor neurons. Ward and colleagues conducted a comparative study with WT mice and Nox3-deficient (head tilt, Nox3het−3J) mice as model for otoconial-deficient inner ear lesions [465]. The group described no eye movement in response to static body tilts about the earth-horizontal axis in Nox3-deficient mice. Through application of a magnetic field, nystagmus occurrence in mice can be studied [647,648,649]. Using this technique, the group saw that WT mice showed different variants of nystagmus. Nose-first entry into the magnetic field induced a left-beating nystagmus, tail-first entry resulted in a right-beating nystagmus. Nox3-deficient mice showed no nystagmus in any of these tested positions. The group nicely showed that the nystagmus occurrence, usually observed in mice with intact vestibular functions, was absent in Nox3-deficient mice and concluded that a functional otoconial structure is critical for the development of a nystagmus in magnetic fields.
4.3.2. Functions of Nox3 in the Cochlea
Interestingly, while Nox3-derived ROS are crucial for otoconia formation and a functional vestibular system [355,370,408,464,466] (Section 4.3.1), in the cochlea, no physiological functions of the Nox3-derived ROS have been described since its discovery [355,392]. On the contrary, Nox3-deficient mice showed normal hearing capacities [217,355,650]. Instead, non-physiological ROS overproduction by Nox3 results in cochlear damage with severe outcomes [404,476,651]. Nox3-mediated ROS overproduction can be easily triggered, e.g., by cisplatin-treatment [652,653,654], by noise exposure [462,470,655] or when blood flow [656,657] or oxygen tension decrease [656]. The correlative involvement of ROS in the cochlea and destruction of hair cells as cause for hearing loss was shown in many studies before [470,658,659]. Of course, the most obvious way to treat this ROS-induced damage is to counter-act with anti-oxidants [469,660], such as methionine [661,662], lipoic acid [659] or NAC [663,664,665] that reduce the global oxidative distress [659,666,667,668,669,670]. No beneficial role for Nox3-derived ROS in the cochlea was discovered so far. Moreover, all studies, which investigated this research topic always reported overproduction of ROS via Nox3 and subsequent cochlea damage and hearing loss. Therefore, this field will be completely discussed in the next section, which summarizes Nox3 involvement in diseases. Remarkably, this topic, i.e. investigation of a possible beneficial effect of Nox3-derived ROS in the cochlea is one of the most intriguing areas for future studies in the Nox3 research field.
5. Roles of Nox3 in Diseases
Since research on Nox3 mainly focused on the most prominent expression region, namely the inner ear, most of the research of Nox3-associated diseases satellite around ear-associated illnesses [462,476,478,651]. Therefore, this section will start with this topic. Nevertheless, tremendous research exploited important roles of Nox3 during lung and cardiovascular diseases, again revising the view of Nox3 as “restricted to the inner ear”.
5.1. Role of Nox3 in Hearing Loss
Hearing loss affects one out of six people and it is one of the major common sensory impairments of humans worldwide [656,671,672]. Hearing loss can be caused by various extrinsic and intrinsic factors, i.e., noise exposure, drug application (including cisplatin), infections and age-related degeneration [672,673,674,675]. The hearing loss in general results from compromised functioning of the organ of Corti in the cochlea and/or the nerve pathways connected to the auditory part of the brain [676]. Several research studies have reported that the nerve connection from the auditory system of the brain to the sound detecting cells (i.e., the hair cells) of the organ of Corti are the most vulnerable parts damaged by endogenous or exogenous sources [404,677,678,679]. The organ of Corti is built up from IHCs and OHCs surrounded by inner and outer phalangeal cells (or Deiters’ cells), inner and outer pillar cells, Hensens cells and Claudius’ cells, all summed up under the term “supporting cells” [398] (Figure 5A). The hair cells detect low- or high-frequency sounds in dependency on their position [680]. Sensory hair cells, in general, do not regenerate in mammals [681,682] and continuous damage results in the permanent loss of hair cells [683]. Oxidative distress is a major driver of hair cell death and subsequent cochlear damage [470,539,651,673,684,685,686,687], which can be induced alongside noise [688,689,690,691], antibiotics [653,692,693], ototoxic anticancer drugs [653,694], infection [695,696] and aging [686,687,697,698,699]. Theses exogenous or endogenous stress factors all result in increased metabolic activity of the cochlea and increased ROS production [667,690,700]. In some cases, like low blood pressure and/or oxygen deprivation, ROS production waves were measured, which started at the luminal surface of the marginal cells in the stria vascularis [657] and re-occurred after reperfusion of the cochlea. The increased ROS levels can last for a long period of time, for example, up to 10 days after noise exposure [469,667,700,701]. This continuous oxidative distress ultimately contributes to death of OHCs and spiral ganglion cells [702,703,704], irreversible cochlea damage and, tragically, permanent hearing loss [404,705,706]. There are many ROS sources in cells with mitochondria [707,708,709] and Nox enzymes as the most prominent ones [6,13,361]. Importantly, mitochondria of OHCs increase their respiratory activity after noise exposure and generate increased amounts of ROS as byproduct [470,710,711], which also contribute to the harmful oxidative damage besides Nox enzymes in general and Nox3 in particular. I point to various excellent reviews about ROS in the inner ear [712,713] or Nox enzymes in this context [686,714] and focus on Nox3-derived ROS. Notably, many studies have used in vivo Wistar rat models, whose hearing ranges are from around 200 Hertz (Hz) to 90 kHz [715] and measured auditory brainstem responses (ABR) for determining the hearing capacity as major experimental output [462,716].
Nagamani et al. first reported a correlation of four patients with interstitial deletion in the 6q region of the long arm of chromosome 6 and Nox3 expression [618]. Deletions of the 6q region were reported before to be associated with ear anomalies [717,718,719,720], but hearing loss was rarely reported [721]. The study suggested that hearing loss occurred because of interstitial or terminal deletions in the 6q25 region, precisely between the regions 6q25.2 and 6q25.3. This area harbors 12 protein-coding genes, with the NOX3 gene among them. The study described for the first time a possible involvement of Nox3-related inner ear diseases in humans, which started the investigation of Nox3 as harmful ROS source and possible therapeutic target (Section 6) for patients.
5.1.1. Noise-Induced Ototoxicity
Prolonged exposure to noise is the most common cause of hearing loss worldwide [722,723,724,725,726,727] responsible for 20% of all cases of hearing loss [675]. Exposure to sound pressure levels that exceed 85 dB or immediate exposure to noise impulses lead to irreversible cochlea damage. Exposure to moderate sound levels over a prolonged time period can also harm the spiral ganglion neurons [728,729]. Noise-induced hearing loss is a result from the combined damaging effects of synaptic damage and cochlear hair cell death [730,731]. The noise-induced hearing loss can be temporary or permanent in dependency of the duration, severity and combination of the damaging factors [404]. A number of additional factors can worsen the progress of hearing loss, e.g., other diseases [164,732], social [470,473] and work behavior [733,734] or working conditions [735]. The frequency ranges of the impairment lie between 3.4 and 6 kHz [735]. Previous studies have also suggested genetic components, which might influence the outcome and severity of noise-induced hearing loss [736,737]. For example, mice that already showed age-induced hearing loss were more susceptible to additional noise-induced hearing loss [738]. Furthermore, several mouse lines, which were deficient for antioxidant components, such as superoxide dismutase 1 (SOD1) [700], glutathione peroxidase 1(GPX1) [700], plasma membrane calcium ATPase 2(PMCA2) [739] or Cadherin Related 23 (CDH23) [740] showed also increased sensitivity to noise-induced hearing loss. These findings suggest an important role for ROS in this context in general. Accordingly, a previous study from Ramkumar et al. reported that noise exposure resulted in an increase of ROS levels, oxidative distress and increased pro-inflammatory responses in the chinchilla cochlea [741]. The pro-inflammatory status in the cochlea is mainly attributed to infiltrating immune cells, mainly monocytes [742,743,744], which respond to the cochlear tissue damage and the previously released chemokines from cochlear cells. Together with the already increased ROS production by Nox3 and mitochondria, the pro-inflammatory environment induces a vicious cycle that further increases the cochlear damage instead of dampening it [688,744,745,746]. Importantly, this pro-inflammatory, pro-oxidative setting is not restricted to noise-induced ototoxicity but can be applied to any effect that leads to increased ROS production and cochlear tissue damage. This scenario represents a complex network of cellular mechanisms and communication in the cochlea that still is incompletely understood and needs further investigation [747].
A number of studies performed genetic screens to identify possible factors that might contribute to noise-induced hearing loss. Lavinsky et al. used a well-established Hybrid Mouse Diversity Panel [748,749,750] to investigate possible loci for susceptibility towards noise-induced hearing loss [650]. The Nox3het allele on the murine chromosome 17 was identified as candidate factor. Nox3het mice were exposed to noise and ABR threshold shifts (4, 8, 12, 16, 24 and 32 kHz) were analyzed. The group measured a reduction in the ABR threshold shifts of WT mice in comparison to Nox3het mice at 8 kHz suggesting a role for Nox3 during noise-induced hearing loss in the lower frequency spectrum. Zhao and colleagues performed a genome wide association study (GWAS) in 614 patients of a case-control study to investigate the interplay of noise kurtosis and lifestyle factors with noise-induced hearing loss [751]. Complex noise induces greater damage to the auditory system than steady noise in both animals and humans [752,753]. A complex noise is defined as continuous background noise with temporal appearance of randomly occurring high-level noises [754]. By transforming time-domain variables, like pulse interval distribution or duration, into simple variables by kurtosis [755,756,757], this experimental approach allows to assess the biological effects of complex noise in animal models [752,753,756]. The group reported that the risk of acquiring noise-induced hearing loss was 0.806-times higher for people, which were exposed to complex noise, as shown previously [753,756]. They detected an increased Guanine-to-Tyrosine polymorphism (single nucleotide polymorphism [SNP] rs12195525, GG phenotype) in the locus, which is located in the coding region of the NOX3 gene. They also observed an increased risk for noise-induced hearing loss in GG phenotype patient groups in which further risk factors, such as smoking or high-volume outputs of technical devices, occurred.
The first study that connected the several correlative dots, i.e., Nox3 expression in the cochlea, per se [355], genetic correlations of noise-induced hearing loss with Nox3 [650,751], increased ROS levels in the cochlea as damaging factors [470,539,651,673,686,687], induction of ROS production by noise exposure [667,690,700] and the subsequent hearing loss, was conducted by Mohri and colleagues [462]. The group investigated the role of Nox3 during noise-induced hearing loss in their dtTomato-Cre reporter system for Nox3 detection in mice [462]. The group exposed 2-month-old WT and Nox3-deficient mice (Nox3 marked with the dtTomato fluorescence tag) to harmful noise at 120 dB for three hours and analyzed the ABR thresholds. At day 7, a lower ABR threshold shift at a high frequency (32 kHz) was measured in Nox3-deficient mice in comparison to WT animals. This was accompanied by a reduced OHC loss in Nox3-deficent animals directly linking Nox3 as damaging factor to hearing loss during noise exposure. A recent study from Rousset and colleagues revised previous findings [462,650] concerning the role of Nox3-derived ROS during hearing loss after white noise exposure [478]. Rousset et al. used the previously described C57BL/6J-NOX3het−4J mouse strain [457], which carries a loss-of-function allele of Nox3. They applied RNAscope in situ hybridization on murine cochlea explants and detected strong Nox3 mRNA expression in the spiral ganglion, while Nox3 was only weakly expressed in the stria vascularis and not detectable in the organ of Corti. The latter is contradictory to several previous studies [355,462,475,479]. Additionally, they detected Nox3 mRNA in the peripheral auditory neurons in Rosenthal’s canal. After noise exposure, Nox3 mRNA expression was increased in cochlear explants, precisely in the medial and the apical cochlea turns. The group also analyzed the hearing capacities of Nox3-deficient mice and observed no difference in the audiograms in comparison to WT animals after 6 weeks of age confirming not a general deficit of hearing in Nox3-deficient animals. Deafening noise exposure (116 dB) led to an elevation of hearing thresholds at frequencies between 16 and 32 kHz after 24 h in WT mice. A protective effect in Nox3-deficient animals was only observed for 32 kHz. After 7 days of noise exposure, ABR measurements showed a better recovery of hearing in Nox3-deficient mice, while WT animals showed no recovery. Histological examinations of cochlear explants further showed that Nox3-deficient animals had reduced hair cells loss, conserved auditory synapses and intact neuron integrity, which all were deceased in WT animals. This study nicely confirmed previous results [404,462,476,655,758], showing that Nox3 has no direct role for cochlear development and structures in sharp contrast to the otoconia formation in the vestibular system [217,370,464]. Even worse, after noise exposure, Nox3-mediated ROS overproduction results in increased oxidative distress and damage of cochlear structures (Figure 5A).
Goodarzi and colleagues investigated the combined effects of noise exposure and silver nanoparticles (Ag-NPs) on the cochlear function in rats [544]. The influence, either beneficial or detrimental, of nanoparticles, in general, on biological functions of the organism is a swiftly expanding research topic [747,759,760,761]. However, metallic nanoparticles, in particular, exploit toxic effects on cells by increasing the ROS production and pro-inflammatory cytokine release [762]. Ag-NPs can enter the body in various ways, e.g., via ingestion, inhalation or even skin contact [763]. Previous studies have reported toxic effects of Ag-NPs to the cochlea [764,765,766]. Goodarzi et al. compared completely untreated Wistar rats with rats exposed to loud noise (104 dB) for different time intervals. The animals either received not further treatment or were intra-peritoneally injected with Ag-NPs (100 mg/kg body weight). The group measured distortion product otoacoustic emissions (DPOAEs) for screening the inner ear function [767,768]. Animals showed a higher rate of hearing loss when exposed to both noise and Ag-NP at frequencies of 7.26, 8.47 and 9.86 kHz. Oppositely, malondialdehyde (MDA) and SOD levels in the serum were either increased by noise exposure or Ag-NP treatment alone but were not further increased by the combined treatment. qRT-PCR analysis further showed that TNFSF2, IL6 and NOX3 gene expressions in the cochlea were increased by one of the treatments alone but were not further increased by the combinatory treatment. Further investigations concerning Nox3-derived ROS were not made. A similar research topic was investigated by Shahtaheri et al. The group investigated the effects of white noise in combination with aluminum oxide (Al2-O3) nanoparticles (AO-NPs) on the cochlear structure in rats [769]. AO-NPs are widely used as thermal insulation material [770], and the exposure to workers that are involved in AO-NPs manufacturing [771,772,773] is correlated with many harmful effects on workers’ health [774,775,776]. Additionally, workers are often exposed to extreme noise levels. Regarding this harmful work environment, Shahtaheri and colleagues analyzed the combinatory harmful effects of AO-NPs and noise exposure (95 dB/20 Hz–20 kHz, 8 h per day) on the cochlea of Wistar rats. The group detected reduced auditory capacities analyzed by DOPAE measurements [727,777] and cochlear damage by histochemical analysis in rats exposed to noise. The damage was further increased by treatment with AO-NPs. AO-NP treatment alone did not alter the investigated parameters. Notably, Nox3 mRNA levels also increased after noise exposure in the cochlea, while AO-NP treatment alone did not change the mRNA expression levels of Nox3. The combinatory effect of both increased Nox3 mRNA expression significantly in comparison to noise exposure alone. This was accompanied by OHC and a supporting cell decrease, while IHC numbers showed no alterations. The authors suggested an enhanced damaging effect of white noise exposure and AO-NP treatment on the cochlea due to increased Nox3-mediated oxidative distress. Critically, neither Nox3 knock-down experiments nor ROS measurements were performed in this context. Hence, again direct evidence for a Nox3 involvement is missing in this study.
5.1.2. Cisplatin-Induced Ototoxicity
Cisplatin is a commonly used chemotherapeutical agent against solid tumors [778,779,780,781,782]. Similar to most chemotherapeutical applications, cisplatin treatment results in strong side effects for the patients like nephrotoxicity and ototoxicity [473,783,784,785,786,787]. Cisplatin-induced nephrotoxicity can be treated with diuretics [788,789], while cisplatin-induced ototoxicity is a much more severe, cumulative and untreatable problem [539,786,790]. It manifests as sensorineural, irreversible hearing loss [791,792,793,794,795] due to damage of the organ of Corti in the cochlea [666,673,796,797]. Specifically, cell death of IHCs and OHCs [473,654,659,798], of spiral ganglion cells [652,799,800,801] and of marginal cells of the stria vascularis [802,803] is increased after cisplatin treatment. Inflammation after cisplatin treatment is another driving factor, which further progresses the cochlear damage [804,805,806,807,808,809]. On the sub-cellular level, cisplatin-mediated cytotoxicity induces DNA damage [810,811], mitochondrial dysfunction [812,813] and increased ROS production by various ROS sources [355,469,813,814,815,816,817]. The accumulating damage due to the oxidative distress further progresses the dysfunction of cochlea [653,654] and vestibular system [817,818,819].
Banfi and colleagues first reported cisplatin-induced Nox3-mediated ROS production by using a co-expression system in HEK293 cells [355]. Mukrerhajea et al. provided further evidence in vivo in the rat cochlea and in vitro in the OHC line UB-OC-1 [820]. Cisplatin treatment induced in both systems increased Nox3 expression and ROS production [541]. Kim and colleagues investigated the role of Nox enzymes during cisplatin-induced ototoxicity in general [540]. They used the mouse auditory cancer cell line HEI-OC1 and in vivo experiments for this approach. Cisplatin treatment induced Nox1 and Nox4 mRNA expression starting after 1 hour. Unfortunately, they claimed that Nox3 mRNA was not detectable; however, the data were not shown in the publication. Notably, in vivo injection of cisplatin for 4 days showed a strong induction of the already basally expressed Nox3 mRNA in the cochlea. However, the group focused on Nox1 and Nox4, and Nox3 as ROS source was not further analyzed. Mohri and colleagues investigated, besides several other important Nox3-related topics (see Section 2, Section 3 and Section 4), also the role of Nox3 during cisplatin-induced hearing loss [476,477]. The group used their well-established reporter system with the dtTomato-coupled Nox3 protein [462]. Tone-burst stimuli (8, 16, 24 and 32 kHz) were applied on 2-month-old WT and Nox3-deficient mice either treated with cisplatin or left untreated. ABR threshold shifts were measured, and WT animals showed deteriorated ABR thresholds at frequencies of 24 and 32 kHz after cisplatin treatment compared to Nox3-deficient animals, which showed no deterioration. WT mice also showed OHC loss, which was lower in Nox3-deficient mice. TdT-mediated dUTP-biotin nick end labeling (TUNEL) assays confirmed increased apoptosis of OHC in WT animals in this context as reported before [476,821]. In Nox3-deficient animals fewer TUNEL-positive OHC were detected. In the lateral wall of the cochlea and the stria vascularis no TUNEL-positive cells were seen in both WT and Nox3-deficient mice. The group furthermore showed that cisplatin treatment increased Nox3 expression in the cochlea predominantly at the basal turn and in the supporting cells. In detail, no Nox3-expressing OHC either with or without cisplatin treatment could be detected in WT animals, while weak Nox3-expression in IHCs and strong Nox3 expression in supporting cells could be observed at least after cisplatin treatment. Together, these studies provide solid evidence that cisplatin treatment increases the presence of Nox3 in the cochlea, which leads to a harmful elevation of ROS production and finally to ototoxicity. Interestingly, in vivo Nox3 is mainly present in the supporting cells and not the OHCs, which nevertheless suffer the greatest damage through the increased ROS production (Figure 5A).
Several studies have provided evidence for a protective role of the activating adenosine A1 receptors (A1ARs) [822] and its agonist adenosine during cochlea-related diseases [479,823,824,825,826,827,828,829,830]. In this context, Kaur and colleagues investigated the role of ROS for the A1AR signaling during cisplatin-induced ototoxicity [758]. They reported that activation of the A1AR signaling pathway by N6-R-phenylisopropyladenosine (R-PIA) prevents hearing loss induced by cisplatin and OHC damage in the rat in vivo. They used the OHC line UB-OC-1 to investigate a role for Nox3-derived ROS in vitro, since ROS have a pro-inflammatory effect during cisplatin-induced ototoxicity [355,541]. Cisplatin treatment for 24 h induced A1AR mRNA and protein expression and increased Nox3 mRNA as well as the total cellular ROS levels. Treatment with R-PIA reduced ROS generation and Nox3 mRNA expression in UB-OC1 cells and in the rat cochlea. Cisplatin treatment of UB-OC-1 cells also induced phosphorylation and nuclear translocation of Signal transducer and activator of transcription 1 (STAT1), which could be inhibited by additional R-PIA treatment. STAT1 signaling contributes to the pro-inflammatory response during cisplatin-induced ototoxicity [831]. Accordingly, treatment with R-PIA reduced cisplatin-induced expression of TNF in the rat cochlea. However, no experiments after Nox3 knock-down or knock-out were performed. Therefore, evidence for the identification of Nox3 as relevant ROS source in this context is missing.
5.1.3. Cytomegalovirus-Induced Hearing Loss
Congenital Cytomegalovirus (CMV) infection often leads to sensorineural hearing loss accompanied by neurological and developmental disabilities [832,833,834,835]. Several studies have monitored apoptotic cell death in the murine cochlea [836] in neonatal mice after CMV infection, subsequently leading to sensorineural hearing loss [549,837]. A correlative increase in total cellular ROS levels was also described in this setting [549]. Due to these previous observations, Zhuang and colleagues picked this topic up and investigated the possible ROS sources and the effect of the anti-inflammatory substance Berberine [838] during CMV-induced ototoxicity [496]. The group detected an increase in apoptosis and total cellular ROS in neonatal murine ex vivo cultured spiral ganglion cells. An increase in Nox3 protein expression was also observed. Additional treatment of Berberine reduced apoptosis, ROS levels and Nox3 expression. However, no genetic evidence was given to validate Nox3 as ROS source. Most critically, the authors claimed that Nox3 was connected to mitochondrial ROS production. No specific mitochondrial ROS measurements were performed, and no co-localization studies of Nox3 with mitochondria, e.g., by immunolabeling and fluorescence microscopy, were conducted. Nevertheless, this is so far the one and only study that has described an induction of Nox3 protein expression as response to infection.
5.1.4. Age-Induced Hearing Loss
Age-induced hearing loss (presbycusis) [839] affects, as the name implies, elderly people. This disease is associated with tremendous social consequences [840,841,842,843]. Similar to other causes for hearing loss, age-induced hearing loss can further progress due to prolonged noise exposure or ototoxic drugs [844]. On the cellular level, the loss of hair cells, spiral ganglion cells and cells of the stria vascularis leads to hearing loss majorly at higher frequencies [841,845]. An increase of age is also accompanied with a disturbance of redox homeostasis not only in the cochlea [686,697], but also in other organs, since gene expression of anti-oxidant systems decrease with age [687,698,699].
Du and colleagues investigated the effects of a HFD in combination with a D-galactosidase-induced rat animal model of aging [846,847] to investigate the cumulative effects on hearing loss [848]. In this animal model, the continuous administration of D-galactose leads to numerous detrimental effects based on metabolic disturbance that mimic the aging process [846,849,850,851]. These effects include dysfunctional mitochondria [850,852,853], increased apoptosis [437,854], neurotoxicity [850,855,856], a shortened lifespan [857] and, after 8 weeks of treatment, symptoms that mimic aging of the cochlea due to increased ROS production [858,859,860,861]. Furthermore, after 8 weeks of D-galactose treatment deletions in the mitochondrial DNA (mtDNA) in the cochlea increase and mitochondria show an oval round shape indicating massive damage. The isolated mtDNA from rat cochlea cells showed increased oxidative damage and subsequent common deletion, which are both biomarkers for oxidative distress, aging and age-related hearing loss [846,862,863,864,865,866]. Du et al. analyzed ABR thresholds and detected the highest ABR threshold shifts for four tested frequencies (4, 8, 16, 32 kHz) in groups treated with both HFD and D-galactose after 12 months. After sole D-galactose treatment Nox3 protein levels increased in the stria vascularis and the spiral ganglion. HFD treatment alone increased Nox3 protein levels only in the stria vascularis. The combined treatment of D-galactose and HFD led to the highest Nox3 expression not only in the stria vascularis and the spiral ganglion, but also in the organ of Corti. Apoptotic cell death in the inner ear was observed for all three conditions, but again the highest cell death rate was reported after the combined treatments. Additionally, all three treatments increased the accumulation of mitochondrial common deletion [867,868], which accompanies mitochondrial damage due to aging [869,870]. Du and colleagues deepened their findings from this previous study [848] with the same D-galactose-induced aging model via RT-PCR and Western blot analysis and reported an increase in Nox3 and p22phox mRNA and protein expression in D-galactose-treated rats in the cochlea [475]. Additional Western blot analysis and TUNEL staining showed that apoptosis increased in the cochlea after D-galactose treatment. These two studies by Du and colleagues gave the first correlative insights of increased Nox3 expression during aging, an associated damaging effect to cochlear structures and the subsequent hearing loss. However, since mitochondria are heavily damaged during this aging model and neither in vivo experiments with Nox3-deficient animals nor in vitro experiments with Nox3 knock-down in cells were conducted, the explicit role and the contribution of Nox3-derived ROS in comparison to ROS produced by the damaged mitochondria remained elusive.
Rousset and colleagues used the A/J mouse strain nmf333, which carries a missense mutation in the p22phox subunit [871], to characterize the role of Nox enzymes in the cochlea during age-induced hearing loss [404]. The group firstly defined age-induced hearing loss in WT animals in their experimental setting. They analyzed ABR threshold levels over an age range from 4 to 26 weeks and observed threshold shifts close to 45 dB after 4 weeks, which progressed up to 75 dB with age. They also detected a progressive hearing loss 32 kHz (in 4-week-old mice) and 5.7 kHz (in 26-week-old mice). In accordance with these data sets, a progressive degeneration of the sensory epithelium from the base to the apical turn was described with a more pronounced cellular degeneration in the basal region. Further analysis of IHC innervation revealed a dramatic decrease in the number of synaptic ribbons per IHC, as well as a decrease in the total neuronal density in the spiral ganglion, which also progressed with age. Since a deficiency of p22phox affects Nox1-4, the group analyzed the presence and distribution of Nox mRNA expression in both the mouse and, highly notably, in the human cochlea. qRT-PCR and in RNAscope in situ hybridization measurements showed high mRNA expression of Nox2, Nox3 and Nox4 in mouse and human cochlea tissue. While Nox2 and Nox4 mRNA was evenly distributed throughout the whole cochlea, Nox3 mRNA was concentrated in the spiral ganglion and moderately expressed in the stria vascularis. Most interestingly, Nox3 mRNA was not detected in hair cells, which is in line with the study from Mohri et al. [462]. p22phox-deficient animals showed no disturbance in hearing at young age in comparison to WT mice. However, the loss of the hearing capacities at high frequencies observed in aged WT mice, was nearly absent in p22phox-deficient animals together with an intact sensory epithelium and preserved synaptic ribbons. The group further performed a transcriptome analysis of 6-week-old cochlea tissue and detected a down-regulation of ryanodine receptors (Ryr) 1, 2 and 3, which are important for Ca2+ homeostasis and accordingly for proper neuronal signaling. Several other genes, all revolving around Ca2+ homoeostasis, such as Otoferlin, Vamp1, and Snap25 or the glutamate transport, such as Slc17a6, Slc17a8 and Gria2 were down-regulated in absence of p22phox. The group narrowed down the auditory neurons as main cell type where the down-regulation was observed. This remarkable study firstly analyzed the mRNA expression of Nox3 in the human cochlea and clearly solidified a rather detrimental effect of Nox presence on cochlear structures, precisely the neuronal part. Unfortunately, like in other previous studies of the Nox3 research field, the group did not clarify the exact interplay of Nox-derived ROS during Ca2+ signaling and the subsequent age-related hearing loss. Moreover, while nicely showing that also Nox2 and Nox4 mRNA is present in the cochlea, the analysis of p22phox-deficient animals only enabled suggestions considering the general role of Nox enzymes in the cochlea and not specifically the role of Nox3, especially since Nox2 and Nox4 might also play important roles in this organ [405,406,407]. Protein expression, for example, in cochlea tissue lysates, was not analyzed, Instead, the research group solely relied on mRNA-detecting techniques. Since the opinion that mRNA always correlates with protein presence or even activity of the protein is outdated [529,530,532,533,534], protein level analysis of the cochlea, especially from human samples would have been a ground-breaking contribution to the field of Nox3-related research. Human-related data sets of this topic are still largely missing to date. In their favor, the group mentioned and discussed these critical points already in their paper. In summary, the studies of Rousset and colleagues [478,651,872], together with Mohri et al. [462] represent milestone research articles considering Nox3 investigations in the inner ear. Continuing in this sense, Mohri and colleagues also investigated the topic of age-induced hearing loss with their generated mouse line, which expresses the fluorescent reporter dtTomato in cells that display Nox3 expression [462] (Section 2, Section 3 and Section 4). The group compared the ABR threshold shifts in WT and Nox3-deficient animals after 1, 2 and 6 months after birth. An increase of Nox3 protein in the cochlea as well as increased ABR threshold shifts at frequencies of 8, 24, and 32 kHz occurred in WT mice over time. Nox3-deficient mice showed no ABR threshold shift increase at all. Especially at high frequencies (24 and 32 kHz), the ABR thresholds were higher in WT mice in comparison to Nox3-deficient animals at 6 months from birth. In addition, histologic analysis of the organ of Corti showed that WT mice at 6 months after birth exhibited OHC loss, while hair cell loss in Nox3-deficient mice was significantly lower. These findings suggest that increased Nox3 expression in the organ of Cori leads to OHC destruction and subsequently contributes to age-related hearing loss (Figure 5A).
5.2. Role of Nox3 during Vertigo
The only study which investigated a rather harmful effect of Nox3-derived ROS on the vestibular system (in contrast to the crucial function of otoconia formation), was conducted by Zhang et al., who investigated factors that influence benign paroxysmal positional vertigo (BPPV) [873]. BPPV is the most common peripheral vertigo-related disease [874,875] occurring in 2.4% of people [876], which increases with age [877]. BPPV is characterized by the detachment of otolith particles, particle movement into the semicircular canal and subsequent loss of otoconial function [441,878]. BPPV is therefore also termed otolithiasis. In dependency how the proper function of the otoconia is impaired, BPPV can be classified in primary BBPV and secondary BPPV. Primary BBPV is induced by factors that directly damage the otoliths or their surroundings, e.g. hair cell damage or loss, endolymph ion changes, decreased otolith protein secretion and defects in otolith-anchoring proteins [441,879]. Secondary BBPV is defined as damage, which is induced as side effect of other harmful events, such as ear surgery, trauma, ototoxic drugs, Meniere’s disease [880] or vestibular neuronitis [881]. Systemic factors like osteoporosis [882], vitamin D deficiency, hypertension, diabetes or cerebrovascular diseases [883] can also contribute to the severity of this disease. Zhang and colleagues focused on vitamin D deficiency during BPPV, since vitamin D is important for proper Ca2+ homoeostasis in general [884,885] and for proper otolith formation and function in particular [881]. Overall, 48 patients with diagnosed BPPV and 48 control patients from the Affiliated Hospital of Inner Mongolia Medical University [886] were analyzed in this study. While no difference in age, body mass index, sex, occurrence of diabetes or hypertension was observed between the groups, BBPV-diagnosed patients showed a decreased bone density and plasma vitamin D levels. Notably, mRNA and protein levels of both OC-90 and Nox3 in the serum were decreased in patients with BPPV. To further analyze the role of vitamin D in this context, vitamin D receptor (VDR)-deficient mice were analyzed. In whole-tissue lysates of the inner ear, mRNA and protein levels of OC-90 and Nox3 were decreased in VDR-deficient mice suggesting a regulatory role of vitamin D in this context. A direct mechanism for VDR-mediated signaling for Nox3-derived ROS production and OC-90 assembly was not investigated.
5.3. Role of Nox3 during Lung Diseases
For a long time, Nox3-related research only focused on either the inner ear or studies focused on broad expression studies to improve the catalogue, which lists if, when and where Nox isoforms are expressed. Most of the latter studies have not focused explicitly on Nox3, but rather described its expression as additional finding. Zhang and colleagues investigated, for the first time, a possible connection between Nox3 and pulmonary emphysema, which is a major contributor to chronic pulmonary diseases [887,888] in a mouse model [483]. They described developing emphysemas in naive TLR4- and MyD88-deficient mice beginning at 3 months after birth and peaking between 6 months and 1 year. This was reflected by increased lung volumes, enlarged air spaces distal to the terminal bronchioles and by destruction of the normal alveolar architecture. These factors are typical for emphysema [887] and occurred in both knock-out animal strains. Notably, all mice strains did not show any significant differences in any pro-inflammatory parameter that was analyzed. However, TLR4-deficient animals showed a decreased elastase inhibitory capacity and increased elastolytic activity in the lung tissue. Since increased oxidative distress is an important correlative factor of emphysema [889] and lung injuries [890,891,892,893,894,895,896,897], the group analyzed the total antioxidant capacity, namely levels of glutathione (GSH) and other antioxidant components in the branchio-alveolar fluid. A strong decrease of GSH levels was detected in the fluid of knock-out animals. Moreover, isolated lungs and isolated lung cells from TLR4-deficient animals showed increased O2− production in comparison to WT animals. The increased ROS levels further led to more oxidative DNA damage, which is also correlated with emphysema [898]. Interestingly, while Nox3 mRNA was only weakly expressed in WT animal lungs and isolated endothelial lung cells, TLR4-deficient lung samples and lung cells showed an increased Nox3 mRNA expression. Additionally, isolated lung cells from TLR4-deficient animals showed an increased elastolytic activity similar to the lung tissue. Knock-down of Nox3 via siRNA in TLR4-deficient lung cells led to a rescue effect of elastolytic activity, nicely confirming the involvement of Nox3. These results clearly demonstrated the connection of TLR4 deficiency, increased Nox3 expression, Nox3 as cause for the increased elastolytic activity and therefore the developed emphysema. A direct mechanism for Nox3-derived ROS was not investigated at that time. Nevertheless, the study of Zhang and colleagues broke the ”inner ear” stigma of Nox3 in terms of disease developement.
In a follow-up study from Zhang and colleagues, a role of Nox enzymes during hyperoxia was investigated. Hyperoxia can occur during sustained oxygen supply in critically ill patients, which can result in respiratory failure [899,900]. Hyperoxia is also an established model for oxidant-induced lung injury [901,902]. Previous reports of the group demonstrated that TLR4-deficient mice showed increased oxidant production in lung tissue and subsequent lung destruction [483], as well as enhanced susceptibility to hyperoxia-induced acute lung injury [903]. An increase in Nox3 mRNA was also reported in TLR4-deficient animals, and siRNA-mediated knock-down partially rescued the phenotype related to TLR4-deficiency [483]. WT mice exposed to hyperoxia showed increased TLR4 mRNA and protein levels in mouse lung endothelial cells and lung lysates. TLR4-deficient mice were more susceptible to hyperoxia, as reported before [903], but interestingly, Nox3-deficient animals showed an increased survival rate. Additional knock-out of Nox3 in TLR4-deficient animals (TLR4/Nox3 double-deficient mice) nearly rescued the animals comparable to WT controls. Hyperoxia conditions increased macrophage, lymphocyte and neutrophil infiltration into the lungs of WT animals, which was further enhanced in TLR4-deficient animals. Nox3-deficient animals, however, showed no differences compared to WT animals. Notably, TLR4/Nox3 double-deficient animals showed a partial rescue from this phenotype. In WT mice, increased lactate dehydrogenase release as well as increased H2O2 and lipid peroxidation levels were detected in lungs after hyperoxia exposure. TLR4 deficiency further increased these parameters, while Nox3-defcient animals showed reduced levels in comparison to WT animals. These data nicely show that TLR4 signaling somehow inhibits Nox3-mediated ROS production in lungs, which is uncoupled when TLR4 as regulating factor is missing. The Nox3-mediated uncontrolled ROS production then leads to lung destruction. When Nox3, as an ROS source, is removed, it either protects the mice in general from lung injury during hyperoxia, or it leaves the TLR4-dependent inhibition as the terminal factor without any effect. The group also discovered that the Heat Shock Protein 70 (Hsp70) [904,905] is necessary for the TLR4-mediated Nox3 inhibition, since mice and endothelial lung cells deficient for Hsp70 showed increased Nox3 mRNA and protein levels. Notably, mitochondrial matrix O2− levels were decreased in TLR4-deficient lung cells and were not altered in Nox3-deficient cells, excluding mitochondria as a potential ROS source in this setting. In addition, this study firstly investigated possible transcription factors that might influence Nox3 mRNA expression. Chromatin immune-precipitation assays identified regions between −2534/−2360 and −1792/−1498 base pairs upstream of the Nox3 promoter as critical binding sites for STAT3 during Nox3 inhibition. In lungs and endothelial lung cells from endothelial STAT3-deficient mice, more Nox3 expression during both basal and hyperoxia conditions was detected. Electrophoretic Mobility Shift Assay (EMSA) analysis showed that Hsp70 induced the STAT3 binding to the Nox3 promoter region only in WT or Myd88-deficient endothelial lung cells, but not in TLR4- or TRIF-deficient cells. Taken together, this study by Zhang and colleagues is probably the most detailed report about Nox3 activation, regulation and function in a specific context so far. The results were solidified by genetic models and ROS measurements not only in vitro, but also in vivo and no cell type or tissue switching during the study was performed. This is a remarkable example of how to perform a scientific analysis about a Nox enzyme and its functions (Figure 5A,B).
Ruwanpura and colleagues further investigated the role of TLR4 and its adaptors MyD88 adapter-like/Toll/interleukin-1 receptor domain-containing adaptor protein (MAL/TIRAP) [906,907] for normal lung architecture and function in mice [908]. They confirmed the findings from Zhang and colleagues [483], i.e., enlargement of the distal air spaces and destruction of normal alveolar architecture without any inflammation in 6-month-old TLR4-deficient mice. Functionally, they found that the static compliance (pulmonary compliance during the inspiratory pause) was significantly increased in TLR4-deficient mice, which was determined by forced oscillatory technique [909,910,911]. The group further described increased oxidative distress in lung tissue, increased Nox3 mRNA and increased apoptosis of alveolar septal cells. Notably, TLR2 deficiency did not alter any of the observed parameters suggesting a TLR4-specific mechanism in this context.
Yasuoka et al. focused on the influence of ROS during the development of lung fibrosis [502]. During lung diseases, fibrosis is a common side effect, which poses a significant increase in morbidity and mortality in patients [912,913,914]. ROS have been implicated as drivers of fibrosis-related pathophysiology [915,916,917] and lung dysfunction [918,919,920]. Fibrosis is accompanied with tissue remodeling and tissue growth as well as development and is regulated by a plethora of growth factors. Yasukoa et al. focused on the insulin-like growth factor binding protein-5 (IGFBP-5), a prominent factor in this context [921,922], and its connection to lung fibrosis and ROS production. They found that primary human lung fibroblasts increased Nox3 mRNA levels and total cellular ROS production after IGFBP-5 or TGF-β treatment. siRNA-mediated knock-down of Nox3 reduced the ROS production in these cells to baseline levels. However, a role for Nox3-derived ROS in the investigated in vivo setting was not conducted.
The discovery of Nox3 as important player for the progression of lung diseases was furthermore confirmed by a series of genetic screens, which delivered correlative data between the NOX3 gene and different lung diseases. Tremblay et al. conducted a GWAS to identify candidate genes as predisposing factors for genetic asthma association studies [923]. The scan, in combination with the Genes-to-Diseases computational analysis tool [924,925], analyzed 609 subjects from the Saguenay-Lac-St-Jean founder population in Quebec, Canada [926,927]. Amongst several other genes, the NOX3 gene was identified as the only NADPH oxidase-related gene. Yin et al. investigated genetic etiology in the context of non-idiopathic pulmonary hypertension (PH) [928,929,930]. Overall, 208 patients were included, 109 patients were diagnosed with non-idiopathic PH and 99 healthy volunteers were included as controls. A total of 143 SNPs were detected in the 109 PH patients with the top hits located in the chromosome 6, precisely in the locus of the NOX3 gene (SNP termed rs6557421). Notably, PH patients with the detected SNP rs6557421 genotype had a 10-fold-higher risk to develop PH in comparison to healthy control samples. Cantu et al. searched for genetic variations that might increase the risk of primary graft dysfunction (PGD) after lung transplantation by a SNP set analysis [931,932]. Rejection of the grafted lung and subsequent organ dysfunction is a major cause of death during the early transplantational period, affecting up to 30% of all patients [933,934,935]. One of the major pathophysiological aspects associated with PGD is increased oxidative distress occurring during ischemia/reperfusion events [936,937,938,939]. In total, 1039 lung transplant recipients and 392 donors were included in this study, and 314 of the 1038 recipients developed PDG and four genes were identified encoding glutathione peroxidase 1 (GPX1), nuclear factor (erythroid-derived 2)-like 2 (NFE2L2), nitric oxide synthase 3 (NOS3) and glutathione S-transferase mu 2 (GSTM2), which all are involved in antioxidant responses [940,941,942]. In the donor group, the genes for Nox3 (NOX3), nitric oxide synthase 1 adaptor protein (NOS1AP) and paraoxonase 1 (PON1) were associated with the development of PGD. Within the NOX3 gene, the SNP rs3749930 had the strongest association with PGD. The detected SNP marks a nucleotide conversion, which resulted in a threonine to lysine aa substitution in a trans-membrane portion of the Nox3 protein. In addition, several intronic SNPs within the NOX3 gene were associated with increased risk of PGD.
All of these studies clearly demonstrate a critical involvement for Nox3-mediated ROS production as rather destructive factor during lung diseases.
5.4. Role of Nox3 during Cardiovasclar Diseases
The term “cardiovascular diseases“ summarizes a broad catalogue of diseases that affect one or many components of the cardiovascular system directly. This includes the heart or the blood circulation system, but also simply all other organs and parts of the body as well, since oxygen and nutrient supply, mediated by the blood stream, are crucial for proper functioning of the organism. Thus, this topic intervenes with many other diseases, which are affected by the cardiovascular system. Similar to nearly any other disease outcome, as well as during any kind of cardiovascular disease, increased ROS production is a major contributing factor that worsens diseases progression [943,944,945,946,947,948]. Of course, the involvement of Nox enzymes as ROS sources was intensively investigated, including Nox3 [162].
5.4.1. Nox3 and Type 2 Diabetes
While fatty acids are crucial components of cellular membranes, chronically increased levels of FFA, consumed with a HFD (Section 5.1.4) lead to obesity due to excessive depositing in non-adipose tissues, e.g. the liver [949,950,951,952]. Subsequently, the development of insulin resistance [537,571], type 2 diabetes [953] and other hepatic diseases [954] dramatically increases. Diabetes mellitus affects more than 300 million people worldwide and represents a disease with high morbidity [955,956]. Type 2 diabetes is associated with various chronic and acute toxic side effects, leading to dyslipidemia, hyperglycemia [957,958,959], diabetic retinopathy [960,961] and chronic hyperinsulinemia. All of these conditions can further induce or enhance adipositas, which is closely related to insulin resistance [561,567,962,963]. Type 2 diabetes and insulin resistance often correlate with increased oxidative distress and an increased systemic pro-inflammatory profile [964] in the according tissues and cells, especially in the liver. Of course, the roles of the Nox isoforms, as primary ROS producers, were investigated in this context [162]. Since Nox3 was identified as an important ROS source in association with diabetic diseases in vitro for HepG2 cells [396,487] (Section 5.1.4) and in a mouse model in vivo [537], further research mostly focused on treatment options. Cremonini et al. investigated the role of the flavanol (-)-Epicatechin [965,966] during HFD-induced insulin-resistance in mice [967]. The group detected a strong up-regulation of Nox3 (60%), Nox4 (274%) and p22phox (237%) protein levels in the liver of mice, which received a HFD in comparison to normally fed mice. Supplemental Epicatechin in the diet prevented this up-regulation. On the in vitro level, similar results were observed in HepG2 cells treated with palmitate and Epicatechin, with exception of p22phox, which remained unaltered. The increased expression of Nox3 and Nox4 resulted in an increased total cellular ROS production. No genetic evidence was provided, and only inhibitors for Nox enzymes were used. Therefore, the specific role of Nox3 or Nox4 could not be determined.
Gupta et al. investigated the effects of Pancreastatin (PST) on adipocyte cells in vitro and in vivo [485]. PST is a peptide secreted by neuroendocrine cells [968], which exploits diabetogenic effects, such as glucose uptake inhibition in liver cells [969,970] or the pancreatic β cell response to insulin [971,972]. Accordingly, treatment with PST is associated with insulin resistance, type 2 diabetes and adipositas [973,974,975]. Since increased ROS levels are involved in lipolysis of adipocytes [529,553] and often correlate with type 2 diabetes progression in patients [162,976,977], the effects of PST on the oxidative distress and chronic insulin induced lipogenesis were also investigated in this study. Neither insulin treatment nor PST treatment alone were sufficient for induction of total cellular ROS production in the adipocyte-like cancer cell line 3T3-L1. Combined treatment induced a slight increase of ROS levels. This corresponded with increased Nox3 protein expression and JNK1/2 phosphorylation. An increase of Nox3 protein expression and JNK1/2 phosphorylation was also detected in white adipose tissue of mice with artificially induced insulin-resistance [978]. While these results nicely contributed to previous findings [396,487,537], no siRNA-mediated knock-down of Nox3 or Nox3-deficient animals were used to clearly confirm Nox3 as the responsible ROS source. Building up from their previous study, the group around Gupta and colleagues researched on possible treatment options with the Pancreastatin inhibitor PSTi8 against insulin resistance [979]. Palmitate treatment of HepG2 cells resulted in lipid accumulation, increased Nox3 mRNA expression, total cellular ROS production and decreased glycogen synthesis. All of these effects were reversed by additional treatment of PSTi. PA also induced phosphorylation of JNK1/2 and p38, which was again prevented by PSTi8 treatment. These findings mark PSTi8 as a potential candidate for diabetic treatment. However, since in both studies, Nox3 was not confirmed as a responsible ROS source, especially since Nox4 is also a prominent ROS source in adipocytes [346,980,981], a clear involvement for Nox3-derived ROS remains elusive.
Malik et al. investigated a previously described therapeutic role of Pterostilbene against insulin resistance [982]. Several studies already described anti-cancer and anti-oxidant effects of Pterostilbene [983], which is a methoxylated Reservatrol analogue [984]. An anti-diabetic effect was also described [985,986,987]. A mechanism of action was not investigated yet. Malik et al. treated HepG2 cells with palmitate, which induced cell death, lipid accumulation, Nox3 mRNA expression, total cellular ROS production and lipid oxidation. Additionally, PA treatment increased expression of genes for proteins involved in fatty acid metabolism, i.e., Sterol regulatory element–binding protein (SREBP1c), Carnitine palmitoyl transferase1 (CPT1), a mitochondrial PA transporter and its transcription factor Peroxisome proliferator-activated receptor alpha (PPARα). All of these effects were strongly reduced after additional treatment with Pterostilbene. While anti-oxidant effects were previously described for Pterostilbene, contradictory, the group observed down-regulation of anti-oxidative enzymes after additional Pterostilbene treatment, therefore outruling an anti-oxidative effect in this context. Since no siRNA knock-down of Nox3 was performed a direct effect of Pterostilbene on Nox3-derived ROS production was not investigated.
Type 2 diabetes negatively affects the outcome of wound healing [988,989] and increased ROS levels correlate with chronic open wounds in patients suffering from Diabetes mellitus [990]. Kim et al. investigated a possible treatment option for improved wound healing [991] by testing the anti-oxidative substance Edaravone. Edaravone was already in use for treatment of acute cerebrovascular diseases [992]. The group used primary human dermal fibroblasts from patients or healthy controls and used the human keratinocyte cell line HaCaT. Furthermore, they conducted a murine in vivo wound healing experiment [993]. Using this model, the group could analyze the expression of Nox3 in tissue flaps near the wound healing area and observed no differences between normo- and hyperglycemic mice after 5 days of operative wound creation. The addition of fibrin for wound healing stimulation or the application of Edaravone did not change Nox3 protein expression. Since no ROS measurements with siRNA knock-down of Nox3 or Nox3 deficient cells were performed, the role of Nox3-derived ROS during the wound healing process remains elusive.
As in the case of lung diseases [923,928,931], also for cardiovascular diseases, GWAS studies were conducted to identify possible risk factors which might influence the disease outcome [994,995,996]. Radowski et al. performed a GWAS to identify genes related to hypertension in 340 patients with type 2 diabetes [997]. Among the six identified genes, the NOX3 gene was also detected, which was previously associated with hypertension [998]. Kwak et al. conducted a GWAS of people with type 2 diabetes to broaden the spectrum of factors, which could help identifying risk factors for cardiovascular diseases in general and type 2 diabetes in particular before the disease outbreak occurs [999]. In their pre-print, they described three variants in genetic loci associated with cardiovascular diseases, especially with type 2 diabetes. Among them, on chromosome 6, there was an intergenic variant between the genes TFB1N and NOX3 (SNP termed rs335407).
5.4.2. Nox3 and Adipositas
Similar to type 2 diabetes and insulin resistance, ROS also play a role during the inflammatory settings associated with adipositas [561,1000,1001,1002,1003]. In adipocytes, the presence of Nox3 was reported before [485]. Issa et al. investigated the influence of cytokines on ROS production and lipolysis in the adipocyte-like cell line 3T3-L1 [529]. Treatment with various pro-inflammatory cytokines (TNF, IL-1β, IFN-γ) induced a slight increase in cellular O2− production after 8 h in differentiated 3T3-L1 cells. It was previously shown that Nox4-derived ROS play an important role for adipocyte differentiation [346,980,981]. Undifferentiated and differentiated 3T3-L1 cells expressed Nox3 as well as Nox4 mRNA. However, only differentiated cells contained the produced Nox3 and Nox4 proteins. While Nox4 expression remained unaltered after cytokine treatment, Nox3 protein levels strongly increased after 8 h. This study nicely showed a decrease in ROS production via Nox3-knockdown after cytokine treatment. Nox-derived ROS were associated with lipolysis in adipocytes before [553] and, indeed, Nox3 knock-down led to an increased lipolysis in 3T3-L1 cells. On the mechanistic level, the group identified an increased phosphorylation of the hormone-sensitive lipase, an enzyme which mediates lipolysis in adipocytes, at the serine residue 536.
5.4.3. Nox3 and Stroke
Stroke is a major consequence of hypertension [1004,1005], and elevated ROS levels have been associated with cerebral hemorrhage [1006,1007,1008]. Michihara and colleagues therefore investigated the role of Nox enzymes during stroke development [489]. The group analyzed the cerebrum in a spontaneously hypertensive rat (stroke-prone) model (SHRSP) [1009]. These SHRSP animals show lower serum cholesterol levels [1010] and increased levels of oxidized proteins in the aorta, heart, kidney [1011] and brain [1012]. Furthermore, increased 8-OHdG levels in the urine and increased ROS levels in the brain of 16-week-old SHRSP animals were reported [1013]. Increased O2− levels, enhanced general Nox activity and increased SOD protein levels were also detected in the brains of SHRSP animals [1014]. Michihara et al. analyzed the mRNA levels of Nox enzymes in the cerebrum of SHRSP animals and found increased mRNA levels of Nox2 and Nox3, while Nox1 and Nox4 were not altered and Nox5 was not detected. Notably, Nox3 protein levels were also increased, while Nox2 levels did not change in comparison to the control animals. This is a nice example that both mRNA and protein levels should always be investigated when suggesting changes in protein presence. However, again, no siRNA-mediated knock-down or Nox3 knock-out model was used to provide evidence that Nox3 is the responsible ROS source for the observed effects in SHRSP animals.
5.4.4. Nox3 and Heart Failure
Several studies have investigated Nox enzymes and their roles for the cardiovascular system in general [102,108,1015] and during human [75,1016,1017,1018], mouse [149,1019,1020,1021] and rat heart failure in particular [186,1022]. While Nox1, Nox2, Nox4 and Nox5 were detected and investigated in this context, the role of Nox3 remained elusive until its detection in murine embryonic stem cell-derived cardiomyocytes by Li and colleagues [149]. The group mainly detected Nox4 mRNA expression, while Nox3 was only weakly expressed and accordingly focused on Nox4. Bkaily and colleagues further analyzed the role of Nox3 in this setting [1023]. For this purpose, they used the hereditary cardiomyopathy hamster model [1024,1025,1026], which is well established for cardiovascular disease studies [1027]. They detected Nox1, Nox2 and Nox4, but no Nox3 protein in the ventricular heart muscles of normal hamsters. In the ventricular heart muscles of cardiomyopathic hamsters, they observed a reduction of Nox1 and Nox4 protein levels and an increase of Nox3 protein, while Nox2 levels remained unchanged. These findings nicely demonstrate that Nox isoforms can show a dynamic expression in dependency of the tissue status. The fluctuation of Nox enzyme expression also demonstrates again that siRNA-mediated knock-down or knock-out experiments are strictly needed when claiming a specific role for a certain Nox enzyme as ROS source. Unfortunately, this was also not conducted in this study.
ROS production is also associated with the pathogenesis of ischemia/reperfusion (I/R)-induced heart injuries [1028,1029] occurring during a myocardial infarction. These injuries include myocardial cell damage and death, arrhythmias or microvascular dysfunction [1030,1031,1032]. Morimoto et al. investigated a putative interplay of ROS and the chemokine monocyte chemoattractant protein-1 (MCP-1) [1033,1034,1035] during I/R [1036]. In vitro experiments with neonatal cardiomyocytes showed that under normoxic conditions MCP-1 had no protective effect. However, after I/R induction, apoptotic cell death increased after 6 h and was reduced by treatment with MCP-1. They used Langendorff-perfused mouse hearts from MHC/MCP-1 mice, which overexpress MCP-1 in the heart for further in vivo investigations [1037]. The group reported an increase of MCP-1 mRNA and ROS production in WT mice after I/R, which was abolished in hearts from MHC/MCP-1 mice. Notably, the group observed mRNA expression of Nox1, Nox2 and Nox3 in the hearts of WT mice, which decreased after I/R. In the hearts of MHC/MCP-1 mice mRNA levels were lower at basal conditions and rose after I/R, again suggesting a dynamic interplay of Nox-derived ROS production. Unfortunately, no Nox silencing or knock-out neither in vitro nor in vivo was performed. Furthermore, no protein expression was analyzed for Nox3. Hence, if and how Nox3-derived ROS production is activated and if these ROS are involved in the context of MCP-1-mediated cardioprotection could not be clarified.
Vats et al. performed a retrospective cohort study [1038] from a population-based Malmö Diet and Cancer Study [1039] with 30,446 subjects over 24.3 years. The group analyzed SNPs to detect genetic variations in genes related to oxidative distress and vitamin intake. The study focused on abdominal aortic aneurysm (AAA) [1040] and unpredictable ruptured AAA [1041,1042], both manifesting in an irreversible and life-threatening dilation of the abdominal aorta [1040,1043,1044]. Accordingly, the study only included participants with occurrence of AAA (25,252 patients in total) [1039]. Oxidative distress has been suggested as a possible link between various factors that contribute to AAA, such as chronic inflammation and cell death [1045,1046], with Nox enzymes as correlated endogenous ROS sources [1045,1047]. During this study, 399 (1.6%) participants were diagnosed with AAA, and 71 (0.2%) were diagnosed with rAAA in general. Furthermore, an amazing effort was made in terms of sub-analytic parameter analysis such as sex, smoking status and physical activity by integrating patient information [1048]. The genetic loci were identified by GWAS and altered SNPs for the NOX5 gene (rs150003957), and the NOX3 gene (rs3749930) were detected. The according male patients showed elevated hazard ratios for AAA, while female patients showed no alterations. Furthermore, participants with the dominant NOX3 gene SNP rs3749930 showed an increased risk for rAAA in the overall study. The group additionally performed subgroup analysis to investigate if the detected oxidative distress-related genotypes had an influence on the effect of antioxidant vitamin intake. They reported that men with the NOX3 gene variant rs3749930 showed an inverse association between higher riboflavin vitamin uptake and a hazard risk for intact AAA, which was also confirmed for the overall study population after sex covariate adjusting.
5.5. Role of Nox3 during Renal Diseases
Chen et al. conducted a GWAS for three phenotypes associated with risk of nephropathy, i.e., serum creatinine levels, creatinine clearance and the glomerular infiltration rate [1049,1050,1051] in 691 type 2 diabetes patients from West Africa to analyze potential factors for reduced renal functions as major consequence of diabetic diseases [998]. The screen detected linkage regions that contain genes, which might influence these three phenotypes. The most prominent candidate genes in these regions that have been implicated in diabetes-induced nephropathy and renal damage were the genes encoding p22phox, (linker region 16q24), Nox1 (linker region 10q22) and Nox3 (linker region 6q25.1–6q26). Together with the study from Ye et al. [492], only two studies investigated Nox3 during kidney-related diseases.
5.6. Role of Nox3 during Gastrointetinal Disaeses
The most dominant Nox isoform in the gastrointestinal tract is Nox1, which was long termed the “colon NADPH oxidase” [13,81,1052]. Nox1 was also detected in the stomach under normal and disease conditions [82,118,1053,1054]. In addition to Nox1, Duox2 is expressed in the rectum, cecum and ascending colon [92,364,1055], and Nox2 and Nox5 were detected in human gastric samples [1053]. However, so far, Nox3 has not been detected nor associated with the gastrointestinal tract.
5.7. Role of Nox3 in Other Diseases
Plantinga et al. investigated genetic variants associated with susceptibility to agranulocytosis [1056]. Agranulocytosis is defined as a reduced concentration of granulocytes in peripheral blood (<500 granulocytes/mL blood) [1057,1058]. Agranulocytosis can be induced by various factors, such as anti-psychotic drugs [1059] or antibiotics [1060,1061], but is also observed in rare events (0.1–0.35%) in patients during treatment with thionamides to medicate hyperthyroidism [1062,1063,1064]. This anti-thyroid drug-induced agranulocytosis (ATDAC) can be a life-threatening condition [1058,1065], especially after the usage of higher doses of anti-thyroid drugs [1066]. During the conducted GWAS of Plantinga et al., two independent families and six patients with Graves’ disease (GD) that developed ATDAC during treatment were analyzed. In 7 out of 11 GD-positive ATDAC patients, a variant of the NOX3 gene were identified. The group reported that the NOX3 gene variants p.Asn8Ser, p.Ala198Thr and p.Arg100Ile were absent in ATDAC-negative GD patients and were not detected in previous genetic screens for predisposition to GD [1067,1068]. Notably, all variants were located in regions of the membrane-spanning α-helices of the Nox3 protein.
CO poisoning is a consequence of malfunctioning oxygen supply due to carboxyhemoglobin forming in red blood cells [1069,1070]. The subsequent hypoxia leads to damage in various brain regions, such as the hippocampus or the striatum [1071]. However, several research groups have suggested that hypoxia alone cannot be addressed as solely responsible for the brain damage. The involvement of various ROS subspecies has been discussed by Hara et al. and others as possible damage-inducing molecules in this context [490,1072,1073,1074,1075]. A previous study already detected increased Duox2 mRNA after CO exposure (3000 ppm, 40 min) in the rat striatum [1076], but no mRNA of other Nox isoforms was detected. Hara et al. revised their findings [543] and used their well-established rat model in which CO exposition (1000 ppm or 3000 ppm) [1072,1077,1078] simulates CO poisoning and brain damage [1073,1079]. The group found a small increase in Nox3 mRNA, while Nox1, Nox2 and Nox4 remained unchanged.
Mikkola et al. performed a GWAS for identification of new gene loci associated with canine hip dysplasia [1080]. This canine skeletal disease is a hereditary disorder [1081,1082] of which the severity varies based on genetic variations [1083,1084,1085] and the dog breed [1084,1086,1087]. The group analyzed 750 German shepherd dogs and identified three new genetic loci associated with this disease. One of these newly identified loci is located on chromosome 1 in an intergenic position between the NOX3 gene and the ARDI1B gene. The group identified the SNP BICF2P468585, which showed the strongest association with the disease and which was located approximately 196 kilobases upstream from the NOX3 gene. Another detected SNP, BICF2S23248027 (also termed rs21911799), was located in the intron between the exons 9 and 10 of the NOX3 gene.
During a study which investigated the therapeutic effects of Dimethyl fumarate (DMF) on relapsing-remitting multiple sclerosis (RRMS) in 564 participants, Carlströem et al. detected a SNP in the NOX3 gene associated with a better DMF treatment outcome [1088]. RRMS is an autoimmune disease characterized by the entry of immune cells into the central nervous system (CNS), which leads to pro-inflammatory tissue damage accompanied by neurological dysfunction [1089,1090]. Like in many other autoimmune pathological settings [1091,1092], oxidative distress was reported to be a modulating factor in RRMS [1093,1094,1095]. DMF (Tecfidera®) is one of the most prescribed substances for patients that suffer from RRMS [1089,1096]. The identified SNP rs6919626 in the NOX3 gene allele was associated with a probability of an insufficient DMF treatment response. The group stimulated CD14+ monocytes isolated from patients with the identified NOX3 SNP rs6919626 with Escherichia coli in vitro and detected a reduced total cellular ROS production. This study suggested for the first time a possible link between Nox3-derived ROS and MS disease outcome and treatment.
Li et al. analyzed thyroid tissue samples from 11 patients who suffered from tertiary hyperparathyroidism (THPT) [1097]. Hyperparathyroidism manifests itself by an enlargement of the parathyroid gland, increased levels of circulating parathyroid hormone, as well as disturbed bone and mineral metabolism [1098,1099]. THPT develops during chronic kidney diseases and differs from hyperparathyroidism in an uncontrolled hypercalcemia, i.e., excessive Ca2+ levels in the blood [1100]. Since the molecular mechanisms of this process remain largely unknown, Li and colleagues investigated this topic by analyzing blood and thyroid tissue samples from 16 Chinese THPT patients. The group used whole-exome sequencing for the detection of SNPs and insertions or deletions variants. During the screen, 17,401 mutations (6690 missense variants, 3078 frameshift variants, 2005 stop-gained variants and 1630 synonymous variants) were detected in THPT patient samples. From this data set, a further driver mutation analysis identified 179 mutated genes, one of them being the NOX3 gene. Expression quantification by qRT-PCR additionally revealed decreased levels of NOX3 gene mRNA in thyroid gland samples from THPT patients.
6. Nox3 as Therapeutic Target
Although Nox3 and Nox3-derived ROS are associated with many different diseases (see Section 5), of course, due to the initial discovery in and research focus on the inner ear, most therapeutical approaches targeted Nox3 in this organ [651,1101]. Nevertheless, research was also conducted to develop therapeutic approaches for Nox3-related involvement during diabetes, cancer and MS.
6.1. Therapeutic Nox3 Targeting in the Inner Ear
6.1.1. Therapeutic Treatment of Cisplatin-Induced Hearing Loss
The first study which investigated Nox3 as a therapeutical target was conducted by Mukherjea and colleagues [541]. They focused on treatment of cisplatin-induced ototoxicity but did not target Nox3 directly. Instead, the group focused on the protein Transient Receptor Potential Vanilloid 1 (TRPV1), which is expressed in the organ of Corti [1102] and can be activated by ROS [1103]. The group reported cell death of OHCs, IHCs, supporting cells, spiral ganglion cells and the stria vascularis in the rat cochlea after 72 h of cisplatin treatment. Cisplatin treatment increased mRNA levels of both TRPV1 (starting at 24 h and increasing over 72 h) and Nox3 (maximum at 24 h). Further in vitro studies with the UB-OC-1 hair cell line confirmed the in vivo observations. Cisplatin treatment induced TRPV1 protein expression and resulted in a higher channel activity, i.e. an increased Ca2+ influx. Nox3 mRNA and protein levels as well as total cellular ROS production were also increased after cisplatin treatment. siRNA-mediated knock-down of Nox3 reduced the cisplatin-induced ROS production, nicely confirming Nox3 as ROS source in this in vitro setting. Moreover, Nox3 down-regulation also decreased TRPV1 mRNA expression. Vice versa, down-regulation of TRPV1 via siRNA reduced the cisplatin-mediated increase of Nox3, suggesting a complex cross-talk between these two proteins. This is not very surprising, since ROS and Ca2+ are major factors, which influence various cellular signaling pathways. TRPV1 as a target for siRNA-mediated therapy was subsequently investigated in vivo. For the analysis of hearing loss, ABR threshold measurements were performed in Wistar rats, which were injected with scrambled or TRPV1-specific siRNA application into the cochlea. Animals were then either left untreated or injected with cisplatin (13 mg/kg, i.p.). In control animals, cisplatin treatment increased ABR thresholds at various frequencies (8, 16, 32 kHz) within 72 h, which indicates progressing hearing loss. In rats pre-treated with TRPV1 siRNA the ABR thresholds at 8 and 16 kHz were moderately reduced, while the cisplatin-induced ABR shifts at 32 kHz showed the strongest reduction after TRPV1 siRNA injection. Histochemical analysis of the cochlea showed damage to and/or loss of hair cells in rats after cisplatin treatment in the basal and middle turns of the cochlea, which was reduced after TRPV1 knock-down. While this study nicely establishes TRPV1 as potential target for therapeutic treatment against cisplatin-induced hearing loss, no in vivo experiments for analysis of Nox3 in this context were performed. Nevertheless, the study clearly showed that cisplatin treatment induces Nox3 expression in the inner ear and confirmed Nox3 as ROS source in the hair cell line UB-OC-1 in this context. Encouraged by their previous findings [541], Mukherjea and colleagues made the next logical step and focused on Nox3 as siRNA target [476]. Transfection of siRNA against Nox3 led to a strong reduction of Nox3 mRNA and protein expression in the cochlea of Wistar rats. As previously discovered [541], cisplatin treatment induced mRNA and protein expression of Nox3 in the cochlea, which was reduced after siRNA treatment. Similar observations were made for spiral ganglion cells and the stria vascularis. Furthermore, cisplatin-induced cell death of OHCs, spiral ganglion cells and the stria vascularis, which occurred after 3 days, was reduced after siRNA-mediated knock-down of Nox3. Finally, by analyzing ABR thresholds, the group observed that the cisplatin-induced ABR threshold shift to 35 dB was reduced to 23 dB after additional treatment with siRNA against Nox3. These two studies from Mukherjea and colleagues [476,541] not only confirm Nox3 expression in the rat cochlea, the rather harmful ROS production during ototoxic drug applications and cisplatin-mediated increase of Nox3 expression, but also nicely demonstrate that targeting of Nox3 via siRNA shows promising potentials for a therapeutic treatment of hearing loss.
The group among Mukherjea, Rakumar and colleagues further investigated possible treatment options for cisplatin-induced hearing loss and focused on the increased pro-inflammatory profile in the cochlea reported after cisplatin treatment [804,805,806,807,809]. The group investigated in this context a possible involvement of the cytosolic transcription factor STAT1 [831]. STAT1 is phosphorylated after the detection of various cellular stress factors, including pro-inflammatory cytokines, translocates to the nucleus and regulates expression of iNOS, TNF [1104,1105] and various factors involved in cell death [1106,1107]. The group observed STAT1 phosphorylation and STAT1 translocation into the nucleus after cisplatin treatment in UB-OC-1 cells and in rat cochlea explants. Increased phosphorylation of STAT1 was specifically detected in OHCs, the stria vascularis and spiral ganglion cells. The group nicely identified Nox3-derived ROS as important factor, since Nox3 knock-down via siRNA reduced the phosphorylation of STAT1 in UB-OC-1 cells and in rat cochlea explants. Moreover, the increased pro-inflammatory profile and cell death of UB-OC-1 cells and rat cochlea OHCs, and the induced hearing loss measured by ABR thresholds after cisplatin treatment all could be attenuated by STAT1 knock-down. Unfortunately, a direct protein target for the Nox3-derived ROS on the mechanistic level, which regulates STAT1, was not identified. The latest study from the Ramkumar lab investigated the role of the chemokine C-X-C motif chemokine ligand 1 (CXCL1) in a similar context [1108]. Cisplatin treatment led to CXCL1-mediated signaling in vitro and in vivo finally resulting in increased pro-inflammatory factors and hearing loss, as described before [1109,1110,1111]. Inhibition of CXCL1 signaling showed a protective effect against hearing loss. Among the up-regulated factors induced by CXCL1 signaling, the NOX3 gene was also described. This study focused on the therapeutic possibilities by targeting the chemokine signaling during cisplatin-induced hearing loss. Therefore, the role of Nox3 in this setting was not further investigated.
Shin et al. designed and investigated a new synthetic compound, named KR-22332, as a treatment for cisplatin-induced hearing loss [542] 2013. Interestingly, the group analyzed zebra fish larvae for their evaluations as addition to the commonly used Wistar rat model. Zebra fish possess hair cells in their lateral line system [1112], which remarkably resembles mammalian inner ear hair cells [1112,1113,1114]. Ototoxic hair cell death can therefore be easily analyzed with this approach [807,1113,1115]. Cisplatin treatment induced significant hair cell loss in zebra fish, which could be reduced by additional treatment with KR-22332. The same results were obtained in vitro with the hair cell line HEI-OC1. Further in vivo experiments with Wistar rats were conducted. Similar to previous findings [355,476,541], the group reported increased cochlear damage and an increase of Nox3 protein levels after cisplatin exposure, which both were reduced by additional K-22332 treatment. Cisplatin-induced hearing loss, determined by an ABR threshold shift measurements at 67 dB, was also reduced to 38.5 dB after K-22332 treatment, altogether suggesting a promising compound for hearing loss treatment after cisplatin exposure. Among steroids, which showed some promising treatment options against cisplatin-induced ototoxicity [1116,1117,1118], the glucocorticoid dexamethasone exploited minimal side effects and protective effects against cisplatin-inflicted damage in the inner ear [1119,1120,1121,1122,1123,1124,1125]. Dinh et al. analyzed the effects of dexamethasone in vitro with rat explants of the organ of Corti [1126]. Cisplatin treatment induced Nox3 mRNA expression, total cellular ROS production and, subsequently, OHC death starting from the basal turn of the organ of Corti after 72 h, while IHCs were not affected. Dexamethasone treatment showed a dose-dependent protective effect in this context.
Kim and colleagues investigated a possible treatment option, which reverses peroxisomal and mitochondrial dysfunction during cisplatin-induced ototoxicity [1127]. They focused on fenofibrate, a pharmaceutical substance usually used to treat unbalanced lipid blood levels [1128]. Fenofibrate already displayed protection against gentamycin-induced ototoxicity [1129] and against cisplatin-induced nephrotoxicity [1130]. Fibrates, such as fenofibrate work mechanistically by binding to the peroxisome proliferation-associated receptor (PPAR), which regulates various cellular functions, mainly the cellular lipid and energy metabolism [1131,1132]. The group investigated hearing loss by ABR threshold shift measurements after cisplatin and fenofibrate treatment and observed, like before, that cisplatin induces higher ABR shifts at all frequency (4, 8, 16, 32 kHz) in mice. Additional fenofibrate treatment alone did not change ABR thresholds. Further analysis of cochlear rat explants showed destruction of OHCs and IHCs, which could be prevented by additional treatment with fenofibrate. These results were confirmed in vitro in the hair cell line HEI-OC1. The group also measured a strong increase in Nox3 and Nox4 protein levels and total cellular ROS production in murine cochlea explants after cisplatin treatment, which was reduced after fenofibrate treatment. They also saw a correlative increase in NF-κB p65 protein levels, which suggests a possible regulatory role for Nox3 expression.
6.1.2. Therapeutic Treatment of Noise-Induced Hearing Loss
Building up on their previous findings, which showed a connection between TVRP1-mediated Ca2+ influx and Nox3-mediated ROS [541], as well as noise-induced Nox3-derived ROS production in rat cochleae [741], the group of Mukerjeah and colleagues further investigated the complex interplay of Ca2+, ROS and pro-inflammatory cytokines during noise-induced hearing loss [655]. ROS as critical drivers of cochlea damage in general have been described several times before [657,667,690,700]. Noise exposure results in increased Ca2+ levels in the cochlea, which, in turn, leads to chronically increased ROS levels [668,1133,1134,1135,1136]. Both factors, permanently increased Ca2+ and ROS levels, subsequently lead to an increased pro-inflammatory status of the cochlea [804,805,806], which attracts immune cells that also further progress the inflammation [807,1137,1138,1139]. Dhukhwa et al. focused on the pro-inflammatory cytokine TNF, which was associated with noise exposure in the rat cochlea before [745,1140,1141] as a possible therapeutic target. After noise exposure, the group measured ABR threshold shifts from 25 to 50 dB at frequencies of 8, 16 and 32 kHz in Wistar rats. This was associated with increased mRNA and protein expression of TRPV1, Nox3, TNF, COX2 and iNOS in the rat cochlea. Additional treatment of animals with capsaicin, the typical agonist of TRPV1 [1142], strongly reduced the mRNA and protein expression of mentioned proteins. Sequestration of TNF by treatment of animals with Etanercept, an IgG1 receptor covalently linked to two TNF receptors [806,1143], reduced TNF and Nox3 protein levels as well as ABR threshold shifts. Early administration (first 2 h of noise application) showed an even stronger otoprotection. However, neither in the in vivo model, nor in the in vitro cell culture experiments, Nox3 knock-out or knock-down animals or cells were used to provide evidence of Nox3 as activated ROS source. Moreover, the possibility that the noise application itself might activate the ROS production of Nox3 alone was not investigated, since no ROS measurements were performed.
After Oishi et al. successfully down-regulated Nox3 expression in the cochlea via direct injection into the murine inner ear [1144], Nacher-Soler and colleagues targeted Nox3 via siRNA in vivo as therapeutical option against sensorineural hearing loss [716]. The group developed a screening method for detecting especially effective Nox3-directed siRNA by establishing a co-expression system. This system resembles early research studies during discovery of the enzyme (Section 1), in which Nox3, p22phox, NOXO1 and NOXA1 of either mouse or human origin, were expressed in the cell lines HEK239, HeLa and CHO. ROS production measured by a water soluble triazonium salt (WTS) reduction assay was used as Nox activity output. Using this biomarker assay, the group identified two potent siRNAs out of ten tested in total, which showed strong down-regulation of Nox3 at very low concentration ranging between 0.1 and 1.13 nM. In mouse cochlear explants, a concentration of 80 nM was necessary to induce a reduction of Nox3 expression to 50% after 48 h. The siRNA #248 showed the most potent effect of the two selected siRNAs and fully matched with the human Nox3 sequence. In vivo delivery of siRNA via intracochlear injection resulted in 60% down-regulation of Nox3 siRNA in the mouse and might therefore bear a relevant human therapeutic approach.
Rousset et al. focused on the new therapeutical possibilities of in vitro designed microRNAs [872]. Like shRNA- or siRNA, microRNA-mediated knock-down of the targeted mRNA is commonly used techniques to investigate cellular processes [1145,1146,1147,1148] and depicts new opportunities for therapeutically uses in patients [1149,1150,1151]. Rousset and colleagues addressed the optimization of miRNAs for a better therapeutical use in general [1152,1153,1154] and chose the subunit p22phox of the Nox isoforms Nox1-4 as one of the therapeutic targets. Indeed, they reported a decrease in p22phox mRNA expression in hair cells, after transduction with an optimized miRNA. However, by only targeting p22phox and not Nox3 specifically, this therapeutic approach will target all Nox isoforms, which can show a highly fluctuating expression profile in dependency on the biological context [6,361,505].
6.2. Therapeutic Nox3 Targeting as Diabetic Treatment
Type 2 diabetes is accompanied by vision loss due to diabetic retinopathy, which is a major complication in diabetic patients [960,961,1155,1156]. Vision loss during diabetic retinopathy is caused by a loss of pericytes and vascular endothelial cells, which leads to vascular dysfunction and neurological inflammation [1157]. There are several treatment options available already, such as the application of anti-VEGF or PKC inhibitors [1158,1159], which are, however, not fully satisfactory due to the complex processes involved in diabetes, such as hyperglycemia and increased oxidative distress [1160,1161,1162]. Cai et al. investigated the glucagon-like peptide 1 (GLP-1), an insulin tropic peptide, which showed potential for diabetes treatment [1163,1164] due to its anti-oxidative properties [1165,1166]. They induced type 2 diabetes in Wistar rats by applying a HFD to investigate this topic. The group described high glucose levels, reduced thickness of retinal cellular structures, namely the columnar and cone photoreceptors, the outer and inner nuclear layer, the inner plexiform layer and the retinal ganglion cell layer and an increased apoptotic cell death of the according retinal cells. All of these parameters were reduced after treatment with GLP-1. The authors also investigated a possible role of Nox3, however, only immunohistological staining of WT retinal explants was performed. A proper negative control staining for Nox3 in Nox3-deficient samples was not conducted. Furthermore, no mRNA or protein level expression was analyzed from retinal lysates to further validate Nox3 involvement and no ROS production was performed. As a result, neither the involvement nor the role of Nox3-derived ROS could be made in this study.
6.3. Therapeutic Nox3 Targeting during Cancer
Saleem et al. investigated a possible therapeutic role of Brevilin A, a plant-derived sesquiterpene lactone [1167,1168,1169], against breast cancer cells [548]. The group used the human breast cancer cell line MCF-7 in this context and reported a dose-dependent reduction of migratory abilities, induction of cell cycle arrest and subsequent cell death after Brevilin A treatment. Further supplementation of the globally working ROS scavenger NAC led to reversed effects suggesting the general involvement of ROS. Indeed, the group measured an increase in total cellular ROS levels after Brevilin A treatment starting at 1 hour and reaching its peak after 2 h. Notably, the group measured an increase of Nox2 and Nox3 protein levels after Brevilin A treatment and therefore suggested the involvement of these two Nox isoforms. However, in comparison to the ROS production, which peaks after two hours, the increase of Nox2 and Nox3 was reported earliest after 24 h, which is too late to explain the described early and quick ROS burst. Moreover, no Nox3-deficient cells or siRNA-mediated knock-down of Nox enzymes was conducted. Therefore, the role of Nox3 in this setting remains elusive.
6.4. Therapeutic Nox3 Targeting during Multiple Sclerosis
Choi et al. investigated an agonist of the lysophosphatidic acid (LPA) receptors as possible treatment option in multiple sclerosis (MS) [1170]. ROS overproduction plays a critical role for the pathological development of MS [1171,1172,1173,1174], e.g., disruption of the blood–brain barrier or acceleration of trans-endothelial migration of peripheral immune cells into the CNS, which further lead to tissue damage. Therefore, ROS indirectly contribute to lesion persistence and deterioration in MS. Several reports have mentioned an induction of ROS production after LPA treatment and subsequent signaling after binding to plasma membrane-located receptors [1175,1176,1177]. LPA functions as both a plasma membrane component and an extracellular signaling mediator in various tissues [1178]. As signaling molecule, LPA induces various processes, such as cell survival, angiogenesis, neurogenesis, and neuroplasticity in the nervous system [1178,1179,1180]. Choi et al. tested the LAP receptor antagonist Ki16425 on MS development and the role of ROS in this context. The group therefore used an established MS model in mice [1181]. Treatment with Ki16425 deteriorated the motor disability, spinal demyelination, enhanced the infiltration of immune cells, such as microglia and Th1 or Th17 helper cells into the spinal cord [1182] and progressed blood–brain barrier disruption [1183]. These events massively worsened the MS symptoms in treated mice. The group also detected increased levels of pro-inflammatory cytokines and of 4-Hydroxynonenal, a common marker of oxidative distress [1184], in the spinal cord of Ki16425-treated animals. They also reported increased mRNA levels of Nox2 and Nox3 after high doses Ki6425-treatment. On the contrary, treatment with the LPAR agonist 1-oleoyl-LPA alleviated the described parameters including Nox3 and Nox2 mRNA expression in the spinal cord of treated animals. While the effects of LAP-mediated signaling on the MS disease outcome were clearly demonstrated in this study, no evidence was given that Nox3-derived ROS were responsible for the observed enhanced oxidative distress in the spinal cord.
7. Concluding Remarks
According to PubMed there are 192 articles, which mention Nox3. The detailed knowledge about this Nox isoform, however, is surprisingly low. With exception of a few ground-breaking milestone articles, most of the studies have only described correlative increase or decrease of Nox3 mRNA in their research context. Confirmation of Nox3-derived ROS as an involved physiological factor on the genetic level or ROS measurements as representative enzymatic output were rare events, on average, of all conducted studies. Despite the fact that Nox3 is expressed not only in the inner ear but also in various cell types and organs, the “inner ear stigma” remains until today. Because of that, Nox3 might be the most underrated Nox isoform to date. Therefore, this review should not only be a helpful compendium of Nox3-associated research but should also function as an encouraging call to all researchers interested in Nox enzymes and Nox-dependent ROS production to focus more on this Nox isoform. The roles and functions of Nox3 are not just limited to the inner ear but extend far beyond it.
Author Contributions
The author has read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the state of North-Rhine-Westphalia, Germany, (AZ: 323-8.04.10.02-141905), German Center for Infection Research, DZIF (TTU 08.927 and TTU 08.928) and the Deutsche Forschungsgemeinschaft (DFG), SFB 670 to M.H procured by Prof. Dr. Martin Krönke. Publication costs were salaried by the author via the thankful utilization of the MDPI reviewer voucher system.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I want to thank Martin Krönke for funding my position. This article was prepared without any use of AI and AI-assisted technology in line with the Committee on Publication Ethics (COPE).
Conflicts of Interest
The author declares no conflict of interest.
References
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The evolution of reactive oxygen species metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best practice & research. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar]
- Holmstrom, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Neish, A.S. Nox enzymes and new thinking on reactive oxygen: A double-edged sword revisited. Annu. Rev. Pathol. 2014, 9, 119–145. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Biol. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. Beobachtungen über die oxydationsprozesse im seeigelei. Biol. Chem. 1908, 57, 1–16. [Google Scholar] [CrossRef]
- Shapiro, B.M. The control of oxidant stress at fertilization. Science 1991, 252, 533–536. [Google Scholar] [CrossRef]
- Nasr-Esfahani, M.M.; Johnson, M.H. The origin of reactive oxygen species in mouse embryos cultured in vitro. Development 1991, 113, 551–560. [Google Scholar] [CrossRef]
- Sirokmany, G.; Donko, A.; Geiszt, M. Nox/duox family of NADPH oxidases: Lessons from knockout mouse models. Trends Pharmacol. Sci. 2016, 37, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schroder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Krause, K.H. The Nox family of Ros-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef]
- Kawahara, T.; Quinn, M.T.; Lambeth, J.D. Molecular evolution of the reactive oxygen-generating NADPH oxidase (Nox/Duox) family of enzymes. BMC Evol. Biol. 2007, 7, 109. [Google Scholar] [CrossRef]
- Karplus, P.A.; Daniels, M.J.; Herriott, J.R. Atomic structure of ferredoxin-NADP+ reductase: Prototype for a structurally novel flavoenzyme family. Science 1991, 251, 60–66. [Google Scholar] [CrossRef]
- Carrillo, N.; Ceccarelli, E.A. Open questions in ferredoxin-NADP+ reductase catalytic mechanism. Eur. J. Biochem. 2003, 270, 1900–1915. [Google Scholar] [CrossRef]
- Wang, M.; Roberts, D.L.; Paschke, R.; Shea, T.M.; Masters, B.S.; Kim, J.J. Three-dimensional structure of NADPH-cytochrome p450 reductase: Prototype for fmn- and fad-containing enzymes. Proc. Natl. Acad. Sci. USA 1997, 94, 8411–8416. [Google Scholar] [CrossRef]
- Leclerc, D.; Wilson, A.; Dumas, R.; Gafuik, C.; Song, D.; Watkins, D.; Heng, H.H.; Rommens, J.M.; Scherer, S.W.; Rosenblatt, D.S.; et al. Cloning and mapping of a cdna for methionine synthase reductase, a flavoprotein defective in patients with homocystinuria. Proc. Natl. Acad. Sci. USA 1998, 95, 3059–3064. [Google Scholar] [CrossRef]
- Vignais, P.V. The superoxide-generating NADPH oxidase: Structural aspects and activation mechanism. Cell. Mol. Life Sci. CMLS 2002, 59, 1428–1459. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, J.D. Nox enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Quinn, M.T. Assembly of the phagocyte NADPH oxidase: Molecular interaction of oxidase proteins. J. Leukoc. Biol. 1996, 60, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. Assembly of the phagocyte NADPH oxidase. Histochem. Cell Biol. 2004, 122, 277–291. [Google Scholar] [CrossRef]
- Geiszt, M.; Leto, T.L. The Nox family of NAD(P)H oxidases: Host defense and beyond. J. Biol. Chem. 2004, 279, 51715–51718. [Google Scholar] [CrossRef]
- Petry, A.; Weitnauer, M.; Gorlach, A. Receptor activation of NADPH oxidases. Antioxid. Redox Signal. 2010, 13, 467–487. [Google Scholar] [CrossRef]
- Stasia, M.J. Cyba encoding p22(phox), the cytochrome b558 alpha polypeptide: Gene structure, expression, role and physiopathology. Gene 2016, 586, 27–35. [Google Scholar] [CrossRef]
- Buvelot, H.; Jaquet, V.; Krause, K.-H. Mammalian NADPH oxidases. In NADPH Oxidases: Methods and Protocols; Knaus, U.G., Leto, T.L., Eds.; Springer: New York, NY, USA, 2019; pp. 17–36. [Google Scholar]
- Davis, A.R.; Mascolo, P.L.; Bunger, P.L.; Sipes, K.M.; Quinn, M.T. Cloning and sequencing of the bovine flavocytochrome b subunit proteins, gp91-phox and p22-phox: Comparison with other known flavocytochrome b sequences. J. Leukoc. Biol. 1998, 64, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Onouchi, H.; Hamada, S.; Machida, C.; Hammond-Kosack, K.E.; Jones, J.D. Six arabidopsis thaliana homologues of the human respiratory burst oxidase (gp91phox). Plant J. Cell Mol. Biol. 1998, 14, 365–370. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species in non-phagocytic cells. J. Leukoc. Biol. 1999, 65, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Teshima, S.; Rokutan, K.; Nikawa, T.; Kishi, K. Guinea pig gastric mucosal cells produce abundant superoxide anion through an NADPH oxidase-like system. Gastroenterology 1998, 115, 1186–1196. [Google Scholar] [CrossRef]
- Linnerz, T.; Hall, C.J. The diverse roles of phagocytes during bacterial and fungal infections and sterile inflammation: Lessons from zebrafish. Front. Immunol. 2020, 11, 1094. [Google Scholar] [CrossRef]
- Inoue, Y.; Suenaga, Y.; Yoshiura, Y.; Moritomo, T.; Ototake, M.; Nakanishi, T. Molecular cloning and sequencing of Japanese pufferfish (Takifugu rubripes) NADPH oxidase cdnas. Dev. Comp. Immunol. 2004, 28, 911–925. [Google Scholar] [CrossRef]
- Ha, E.M.; Oh, C.T.; Bae, Y.S.; Lee, W.J. A direct role for dual oxidase in drosophila gut immunity. Science 2005, 310, 847–850. [Google Scholar] [CrossRef]
- Carol, R.J.; Dolan, L. The role of reactive oxygen species in cell growth: Lessons from root hairs. J. Exp. Bot. 2006, 57, 1829–1834. [Google Scholar] [CrossRef] [PubMed]
- Keller, T.; Damude, H.G.; Werner, D.; Doerner, P.; Dixon, R.A.; Lamb, C. A plant homolog of the neutrophil NADPH oxidase gp91phox subunit gene encodes a plasma membrane protein with Ca2+ binding motifs. Plant Cell 1998, 10, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Pathogen-induced, NADPH oxidase-derived reactive oxygen intermediates suppress spread of cell death in arabidopsis thaliana. Nat. Genet. 2005, 37, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J. Exploitation of reactive oxygen species by fungi: Roles in host-fungus interaction and fungal development. J. Microbiol. Biotechnol. 2014, 24, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Lara-Ortiz, T.; Riveros-Rosas, H.; Aguirre, J. Reactive oxygen species generated by microbial NADPH oxidase noxa regulate sexual development in aspergillus nidulans. Mol. Microbiol. 2003, 50, 1241–1255. [Google Scholar] [CrossRef] [PubMed]
- Lardy, B.; Bof, M.; Aubry, L.; Paclet, M.H.; Morel, F.; Satre, M.; Klein, G. NADPH oxidase homologs are required for normal cell differentiation and morphogenesis in dictyostelium discoideum. Biochim. Biophys. Acta 2005, 1744, 199–212. [Google Scholar] [CrossRef]
- Eichinger, L.; Noegel, A.A. Comparative genomics of dictyostelium discoideum and entamoeba histolytica. Curr. Opin. Microbiol. 2005, 8, 606–611. [Google Scholar] [CrossRef]
- Edens, W.A.; Sharling, L.; Cheng, G.; Shapira, R.; Kinkade, J.M.; Lee, T.; Edens, H.A.; Tang, X.; Sullards, C.; Flaherty, D.B.; et al. Tyrosine cross-linking of extracellular matrix is catalyzed by duox, a multidomain oxidase/peroxidase with homology to the phagocyte oxidase subunit gp91phox. J. Cell Biol. 2001, 154, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Wingler, K.; Hermans, J.J.; Schiffers, P.; Moens, A.; Paul, M.; Schmidt, H.H. Nox1, 2, 4, 5: Counting out oxidative stress. Br. J. Pharmacol. 2011, 164, 866–883. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological defense mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef]
- Jesaitis, A.J. Structure of human phagocyte cytochrome b and its relationship to microbicidal superoxide production. J. Immunol. 1995, 155, 3286–3288. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A. Activation of the neutrophil respiratory burst oxidase. J. Infect. Dis. 1999, 179 (Suppl. 2), S309–S317. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.H. Tissue distribution and putative physiological function of nox family NADPH oxidases. Jpn. J. Infect. Dis. 2004, 57, S28–S29. [Google Scholar] [PubMed]
- Green, T.R.; Shangguan, X. Stoichiometry of O2 metabolism and NADPH oxidation of the cell-free latent oxidase reconstituted from cytosol and solubilized membrane from resting human neutrophils. J. Biol. Chem. 1993, 268, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Parkos, C.A.; Allen, R.A.; Cochrane, C.G.; Jesaitis, A.J. Purified cytochrome b from human granulocyte plasma membrane is comprised of two polypeptides with relative molecular weights of 91,000 and 22,000. J. Clin. Investig. 1987, 80, 732–742. [Google Scholar] [CrossRef]
- Parkos, C.A.; Dinauer, M.C.; Walker, L.E.; Allen, R.A.; Jesaitis, A.J.; Orkin, S.H. Primary structure and unique expression of the 22-kilodalton light chain of human neutrophil cytochrome b. Proc. Natl. Acad. Sci. USA 1988, 85, 3319–3323. [Google Scholar] [CrossRef]
- Cheng, G.; Cao, Z.; Xu, X.; van Meir, E.G.; Lambeth, J.D. Homologs of gp91phox: Cloning and tissue expression of Nox3, Nox4, and Nox5. Gene 2001, 269, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.M.; Dunne, M.J.; Segal, A.W. Purification of cytochrome b-245 from human neutrophils. Biochem. J. 1984, 219, 519–527. [Google Scholar] [CrossRef]
- Dinauer, M.C.; Orkin, S.H.; Brown, R.; Jesaitis, A.J.; Parkos, C.A. The glycoprotein encoded by the x-linked chronic granulomatous disease locus is a component of the neutrophil cytochrome b complex. Nature 1987, 327, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Lambeth, J.D.; Nauseef, W. The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 2002, 397, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Baehner, R.L.; Nathan, D.G. Leukocyte oxidase: Defective activity in chronic granulomatous disease. Science 1967, 155, 835–836. [Google Scholar] [CrossRef] [PubMed]
- Baehner, R.L.; Nathan, D.G. Quantitative nitroblue tetrazolium test in chronic granulomatous disease. N. Engl. J. Med. 1968, 278, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Iyer, G.Y.; Islam, M.F.; Quastel, J.H. Biochemical aspects of phagocytosis. Nature 1961, 192, 535–541. [Google Scholar] [CrossRef]
- Baldridge, C.W.; Gerard, R.W. The extra respiration of phagocytosis. Am. J. Physiol.-Leg. Content 1932, 103, 235–236. [Google Scholar] [CrossRef]
- Rossi, F.; Zatti, M. Biochemical aspects of phagocytosis in polymorphonuclear leucocytes. Nadh and NADPH oxidation by the granules of resting and phagocytizing cells. Experientia 1964, 20, 21–23. [Google Scholar] [CrossRef]
- Morel, F.; Doussiere, J.; Vignais, P.V. The superoxide-generating oxidase of phagocytic cells. Physiological, molecular and pathological aspects. Eur. J. Biochem. 1991, 201, 523–546. [Google Scholar] [CrossRef]
- Babior, B.M. NADPH oxidase: An update. Blood 1999, 93, 1464–1476. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. The phagocyte nox2 NADPH oxidase in microbial killing and cell signaling. Curr. Opin. Immunol. 2019, 60, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Sbarra, A.J.; Karnovsky, M.L. The biochemical basis of phagocytosis. I. Metabolic changes during the ingestion of particles by polymorphonuclear leukocytes. J. Biol. Chem. 1959, 234, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Mandell, G.L. Bactericidal activity of aerobic and anaerobic polymorphonuclear neutrophils. Infect. Immun. 1974, 9, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Quinn, M.T.; Gauss, K.A. Structure and regulation of the neutrophil respiratory burst oxidase: Comparison with nonphagocyte oxidases. J. Leukoc. Biol. 2004, 76, 760–781. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef]
- Cross, A.R.; Segal, A.W. The NADPH oxidase of professional phagocytes—Prototype of the NOX electron transport chain systems. Biochim. Biophys. Acta 2004, 1657, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef]
- Nauseef, W.M. How human neutrophils kill and degrade microbes: An integrated view. Immunol. Rev. 2007, 219, 88–102. [Google Scholar] [CrossRef]
- Lam, G.Y.; Huang, J.; Brumell, J.H. The many roles of Nox2 NADPH oxidase-derived ROS in immunity. Semin. Immunopathol. 2010, 32, 415–430. [Google Scholar] [CrossRef]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef]
- Badwey, J.A.; Curnutte, J.T.; Robinson, J.M.; Lazdins, J.K.; Briggs, R.T.; Karnovsky, M.J.; Karnovsky, M.L. Comparative aspects of oxidative metabolism of neutrophils from human blood and guinea pig peritonea: Magnitude of the respiratory burst, dependence upon stimulating agents, and localization of the oxidases. J. Cell. Physiol. 1980, 105, 541–545. [Google Scholar] [CrossRef]
- Ohno, Y.; Hirai, K.; Kanoh, T.; Uchino, H.; Ogawa, K. Subcellular localization of H2O2 production in human neutrophils stimulated with particles and an effect of cytochalasin-b on the cells. Blood 1982, 60, 253–260. [Google Scholar] [CrossRef]
- Heymes, C.; Bendall, J.K.; Ratajczak, P.; Cave, A.C.; Samuel, J.L.; Hasenfuss, G.; Shah, A.M. Increased myocardial NADPH oxidase activity in human heart failure. J. Am. Coll. Cardiol. 2003, 41, 2164–2171. [Google Scholar] [CrossRef]
- Javeshghani, D.; Magder, S.A.; Barreiro, E.; Quinn, M.T.; Hussain, S.N. Molecular characterization of a superoxide-generating NAD(P)H oxidase in the ventilatory muscles. Am. J. Respir. Crit. Care Med. 2002, 165, 412–418. [Google Scholar] [CrossRef]
- Reinehr, R.; Becker, S.; Eberle, A.; Grether-Beck, S.; Haussinger, D. Involvement of NADPH oxidase isoforms and src family kinases in cd95-dependent hepatocyte apoptosis. J. Biol. Chem. 2005, 280, 27179–27194. [Google Scholar] [CrossRef]
- Piccoli, C.; Ria, R.; Scrima, R.; Cela, O.; D’Aprile, A.; Boffoli, D.; Falzetti, F.; Tabilio, A.; Capitanio, N. Characterization of mitochondrial and extra-mitochondrial oxygen consuming reactions in human hematopoietic stem cells. Novel evidence of the occurrence of NAD(P)H oxidase activity. J. Biol. Chem. 2005, 280, 26467–26476. [Google Scholar] [CrossRef]
- Altenhofer, S.; Radermacher, K.A.; Kleikers, P.W.; Wingler, K.; Schmidt, H.H. Evolution of NADPH oxidase inhibitors: Selectivity and mechanisms for target engagement. Antioxid. Redox Signal. 2015, 23, 406–427. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, Z.M.; Henneke, P.; Kolter, J. From flies to men: Ros and the NADPH oxidase in phagocytes. Front. Cell Dev. Biol. 2021, 9, 628991. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.A.; Arnold, R.S.; Lassegue, B.; Shi, J.; Xu, X.; Sorescu, D.; Chung, A.B.; Griendling, K.K.; Lambeth, J.D. Cell transformation by the superoxide-generating oxidase Mox1. Nature 1999, 401, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Banfi, B.; Maturana, A.; Jaconi, S.; Arnaudeau, S.; Laforge, T.; Sinha, B.; Ligeti, E.; Demaurex, N.; Krause, K.H. A mammalian h+ channel generated through alternative splicing of the NADPH oxidase homolog noh-1. Science 2000, 287, 138–142. [Google Scholar] [CrossRef]
- Bokoch, G.M.; Diebold, B.A. Current molecular models for NADPH oxidase regulation by Rac gtpase. Blood 2002, 100, 2692–2696. [Google Scholar] [CrossRef] [PubMed]
- Geiszt, M.; Kopp, J.B.; Varnai, P.; Leto, T.L. Identification of renox, an NAD(P)H oxidase in kidney. Proc. Natl. Acad. Sci. USA 2000, 97, 8010–8014. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Chen, X. The human Nox4: Gene, structure, physiological function and pathological significance. J. Drug Target. 2015, 23, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Madyastha, P.; Bingel, S.; Ries, W.; Key, L. A new superoxide-generating oxidase in murine osteoclasts. J. Biol. Chem. 2001, 276, 5452–5458. [Google Scholar] [CrossRef]
- Banfi, B.; Molnar, G.; Maturana, A.; Steger, K.; Hegedus, B.; Demaurex, N.; Krause, K.H. A Ca2+-activated NADPH oxidase in testis, spleen, and lymph nodes. J. Biol. Chem. 2001, 276, 37594–37601. [Google Scholar] [CrossRef] [PubMed]
- De Deken, X.; Wang, D.; Many, M.C.; Costagliola, S.; Libert, F.; Vassart, G.; Dumont, J.E.; Miot, F. Cloning of two human thyroid cDNAs encoding new members of the NADPH oxidase family. J. Biol. Chem. 2000, 275, 23227–23233. [Google Scholar] [CrossRef]
- Bokoch, G.M.; Knaus, U.G. NADPH oxidases: Not just for leukocytes anymore! Trends Biochem. Sci. 2003, 28, 502–508. [Google Scholar] [CrossRef]
- Sumimoto, H.; Miyano, K.; Takeya, R. Molecular composition and regulation of the Nox family NAD(P)H oxidases. Biochem. Biophys. Res. Commun. 2005, 338, 677–686. [Google Scholar] [CrossRef]
- Donko, A.; Peterfi, Z.; Sum, A.; Leto, T.; Geiszt, M. Dual oxidases. Philosophical transactions of the Royal Society of London. Ser. B Biol. Sci. 2005, 360, 2301–2308. [Google Scholar]
- Geiszt, M.; Witta, J.; Baffi, J.; Lekstrom, K.; Leto, T.L. Dual oxidases represent novel hydrogen peroxide sources supporting mucosal surface host defense. FASEB J. 2003, 17, 1502–1504. [Google Scholar] [CrossRef]
- Belikov, A.V.; Schraven, B.; Simeoni, L. T cells and reactive oxygen species. J. Biomed. Sci. 2015, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Holmdahl, R.; Sareila, O.; Pizzolla, A.; Winter, S.; Hagert, C.; Jaakkola, N.; Kelkka, T.; Olsson, L.M.; Wing, K.; Backdahl, L. Hydrogen peroxide as an immunological transmitter regulating autoreactive t cells. Antioxid. Redox Signal. 2013, 18, 1463–1474. [Google Scholar] [CrossRef]
- Meier, B.; Cross, A.R.; Hancock, J.T.; Kaup, F.J.; Jones, O.T. Identification of a superoxide-generating NADPH oxidase system in human fibroblasts. Biochem. J. 1991, 275 Pt 1, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Takeya, R.; Sumimoto, H. Molecular mechanism for activation of superoxide-producing NADPH oxidases. Mol. Cells 2003, 16, 271–277. [Google Scholar] [CrossRef]
- Schroder, K.; Zhang, M.; Benkhoff, S.; Mieth, A.; Pliquett, R.; Kosowski, J.; Kruse, C.; Luedike, P.; Michaelis, U.R.; Weissmann, N.; et al. Nox4 is a protective reactive oxygen species generating vascular NADPH oxidase. Circ. Res. 2012, 110, 1217–1225. [Google Scholar] [CrossRef]
- Nayernia, Z.; Jaquet, V.; Krause, K.H. New insights on Nox enzymes in the central nervous system. Antioxid. Redox Signal. 2014, 20, 2815–2837. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Sorescu, D.; Ushio-Fukai, M. NAD(P)H oxidase: Role in cardiovascular biology and disease. Circ. Res. 2000, 86, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Zalba, G.; San Jose, G.; Moreno, M.U.; Fortuno, M.A.; Fortuno, A.; Beaumont, F.J.; Diez, J. Oxidative stress in arterial hypertension: Role of NAD(P)H oxidase. Hypertension 2001, 38, 1395–1399. [Google Scholar] [CrossRef]
- Jones, S.A.; Hancock, J.T.; Jones, O.T.; Neubauer, A.; Topley, N. The expression of NADPH oxidase components in human glomerular mesangial cells: Detection of protein and mRNA for p47phox, p67phox, and p22phox. J. Am. Soc. Nephrol. JASN 1995, 5, 1483–1491. [Google Scholar] [CrossRef]
- Ahmarani, L.; Avedanian, L.; Al-Khoury, J.; Perreault, C.; Jacques, D.; Bkaily, G. Whole-cell and nuclear NADPH oxidases levels and distribution in human endocardial endothelial, vascular smooth muscle, and vascular endothelial cells. Can. J. Physiol. Pharmacol. 2013, 91, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Valencia, A.; Rajadurai, A.; Carle, A.B.; Kochevar, I.E. 7-dehydrocholesterol enhances ultraviolet a-induced oxidative stress in keratinocytes: Roles of NADPH oxidase, mitochondria, and lipid rafts. Free Radic. Biol. Med. 2006, 41, 1704–1718. [Google Scholar] [CrossRef] [PubMed]
- Sambo, P.; Baroni, S.S.; Luchetti, M.; Paroncini, P.; Dusi, S.; Orlandini, G.; Gabrielli, A. Oxidative stress in scleroderma: Maintenance of scleroderma fibroblast phenotype by the constitutive up-regulation of reactive oxygen species generation through the NADPH oxidase complex pathway. Arthritis Rheum. 2001, 44, 2653–2664. [Google Scholar] [CrossRef]
- Quinn, M.T.; Ammons, M.C.; Deleo, F.R. The expanding role of NADPH oxidases in health and disease: No longer just agents of death and destruction. Clin. Sci. 2006, 111, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.R.; Kachra, Z.; Spokes, K.C.; Aird, W.C. NADPH oxidase activity is required for endothelial cell proliferation and migration. FEBS Lett. 2000, 486, 252–256. [Google Scholar] [CrossRef]
- D’Autreaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Lassegue, B.; San Martin, A.; Griendling, K.K. Biochemistry, physiology, and pathophysiology of NADPH oxidases in the cardiovascular system. Circ. Res. 2012, 110, 1364–1390. [Google Scholar] [CrossRef]
- Lambeth, J.D. Regulation of the phagocyte respiratory burst oxidase by protein interactions. J. Biochem. Mol. Biol. 2000, 33, 427–439. [Google Scholar]
- Reeves, E.P.; Lu, H.; Jacobs, H.L.; Messina, C.G.; Bolsover, S.; Gabella, G.; Potma, E.O.; Warley, A.; Roes, J.; Segal, A.W. Killing activity of neutrophils is mediated through activation of proteases by k+ flux. Nature 2002, 416, 291–297. [Google Scholar] [CrossRef]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Inside the neutrophil phagosome: Oxidants, myeloperoxidase, and bacterial killing. Blood 1998, 92, 3007–3017. [Google Scholar] [CrossRef]
- Skonieczna, M.; Hejmo, T.; Poterala-Hejmo, A.; Cieslar-Pobuda, A.; Buldak, R.J. NADPH oxidases: Insights into selected functions and mechanisms of action in cancer and stem cells. Oxidative Med. Cell. Longev. 2017, 2017, 9420539. [Google Scholar] [CrossRef]
- Juhasz, A.; Ge, Y.; Markel, S.; Chiu, A.; Matsumoto, L.; van Balgooy, J.; Roy, K.; Doroshow, J.H. Expression of NADPH oxidase homologues and accessory genes in human cancer cell lines, tumours and adjacent normal tissues. Free Radic. Res. 2009, 43, 523–532. [Google Scholar] [CrossRef]
- Le Belle, J.E.; Orozco, N.M.; Paucar, A.A.; Saxe, J.P.; Mottahedeh, J.; Pyle, A.D.; Wu, H.; Kornblum, H.I. Proliferative neural stem cells have high endogenous ros levels that regulate self-renewal and neurogenesis in a pi3k/akt-dependant manner. Cell Stem Cell 2011, 8, 59–71. [Google Scholar] [CrossRef]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell. Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, J.; Post, J.A. Molecular events associated with reactive oxygen species and cell cycle progression in mammalian cells. Gene 2004, 337, 1–13. [Google Scholar] [CrossRef]
- Arnold, R.S.; Shi, J.; Murad, E.; Whalen, A.M.; Sun, C.Q.; Polavarapu, R.; Parthasarathy, S.; Petros, J.A.; Lambeth, J.D. Hydrogen peroxide mediates the cell growth and transformation caused by the mitogenic oxidase Nox1. Proc. Natl. Acad. Sci. USA 2001, 98, 5550–5555. [Google Scholar] [CrossRef] [PubMed]
- Teshima, S.; Kutsumi, H.; Kawahara, T.; Kishi, K.; Rokutan, K. Regulation of growth and apoptosis of cultured guinea pig gastric mucosal cells by mitogenic oxidase 1. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G1169–G1176. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Bunderson, M.; Wilham, J.; Black, S.M. Important role for Rac1 in regulating reactive oxygen species generation and pulmonary arterial smooth muscle cell growth. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L1314–L1322. [Google Scholar] [CrossRef] [PubMed]
- Block, K.; Gorin, Y. Aiding and abetting roles of nox oxidases in cellular transformation. Nat. Rev. Cancer 2012, 12, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Weyemi, U.; Redon, C.E.; Parekh, P.R.; Dupuy, C.; Bonner, W.M. NADPH oxidases Noxs and Duoxs as putative targets for cancer therapy. Anti-Cancer Agents Med. Chem. 2013, 13, 502–514. [Google Scholar]
- Brar, S.S.; Kennedy, T.P.; Sturrock, A.B.; Huecksteadt, T.P.; Quinn, M.T.; Whorton, A.R.; Hoidal, J.R. An NAD(P)H oxidase regulates growth and transcription in melanoma cells. Am. J. Physiol. Cell Physiol. 2002, 282, C1212–C1224. [Google Scholar] [CrossRef] [PubMed]
- Seshiah, P.N.; Weber, D.S.; Rocic, P.; Valppu, L.; Taniyama, Y.; Griendling, K.K. Angiotensin ii stimulation of NAD(P)H oxidase activity: Upstream mediators. Circ. Res. 2002, 91, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Ushio-Fukai, M. Redox signaling in angiogenesis: Role of NADPH oxidase. Cardiovasc. Res. 2006, 71, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, K.; Yamada, H.; Iwata, K.; Jin, D.; Katsuyama, M.; Matsuki, M.; Takai, S.; Yamanishi, K.; Miyazaki, M.; Matsubara, H.; et al. Nox1 is involved in angiotensin ii-mediated hypertension: A study in nox1-deficient mice. Circulation 2005, 112, 2677–2685. [Google Scholar] [CrossRef]
- Lassegue, B.; Clempus, R.E. Vascular NAD(P)H oxidases: Specific features, expression, and regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R277–R297. [Google Scholar] [CrossRef] [PubMed]
- Santillo, M.; Colantuoni, A.; Mondola, P.; Guida, B.; Damiano, S. Nox signaling in molecular cardiovascular mechanisms involved in the blood pressure homeostasis. Front. Physiol. 2015, 6, 194. [Google Scholar] [CrossRef]
- Schramm, A.; Matusik, P.; Osmenda, G.; Guzik, T.J. Targeting NADPH oxidases in vascular pharmacology. Vascul. Pharmacol. 2012, 56, 216–231. [Google Scholar] [CrossRef]
- Corvilain, B.; van Sande, J.; Laurent, E.; Dumont, J.E. The H2O2-generating system modulates protein iodination and the activity of the pentose phosphate pathway in dog thyroid. Endocrinology 1991, 128, 779–785. [Google Scholar] [CrossRef]
- Harrison, D.; Griendling, K.K.; Landmesser, U.; Hornig, B.; Drexler, H. Role of oxidative stress in atherosclerosis. Am. J. Cardiol. 2003, 91, 7A–11A. [Google Scholar] [CrossRef] [PubMed]
- Dworakowski, R.; Anilkumar, N.; Zhang, M.; Shah, A.M. Redox signalling involving NADPH oxidase-derived reactive oxygen species. Biochem. Soc. Trans. 2006, 34, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Irani, K.; Xia, Y.; Zweier, J.L.; Sollott, S.J.; Der, C.J.; Fearon, E.R.; Sundaresan, M.; Finkel, T.; Goldschmidt-Clermont, P.J. Mitogenic signaling mediated by oxidants in Ras-transformed fibroblasts. Science 1997, 275, 1649–1652. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, P.; Cirri, P. Redox regulation of protein tyrosine phosphatases during receptor tyrosine kinase signal transduction. Trends Biochem. Sci. 2003, 28, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Ziff, M. Increased superoxide anion release from human endothelial cells in response to cytokines. J. Immunol. 1986, 137, 3295–3298. [Google Scholar] [CrossRef] [PubMed]
- Werner, E. Gtpases and reactive oxygen species: Switches for killing and signaling. J. Cell Sci. 2004, 117, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef]
- Anilkumar, N.; Weber, R.; Zhang, M.; Brewer, A.; Shah, A.M. Nox4 and Nox2 NADPH oxidases mediate distinct cellular redox signaling responses to agonist stimulation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1347–1354. [Google Scholar] [CrossRef]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Cooney, S.J.; Bermudez-Sabogal, S.L.; Byrnes, K.R. Cellular and temporal expression of NADPH oxidase (nox) isotypes after brain injury. J. Neuroinflamm. 2013, 10, 155. [Google Scholar] [CrossRef]
- Coyoy, A.; Olguin-Albuerne, M.; Martinez-Briseno, P.; Moran, J. Role of reactive oxygen species and NADPH-oxidase in the development of rat cerebellum. Neurochem. Int. 2013, 62, 998–1011. [Google Scholar] [CrossRef]
- Olguin-Albuerne, M.; Moran, J. Ros produced by nox2 control in vitro development of cerebellar granule neurons development. ASN Neuro 2015, 7, 1759091415578712. [Google Scholar] [CrossRef]
- Yoneyama, M.; Kawada, K.; Gotoh, Y.; Shiba, T.; Ogita, K. Endogenous reactive oxygen species are essential for proliferation of neural stem/progenitor cells. Neurochem. Int. 2010, 56, 740–746. [Google Scholar] [CrossRef]
- Cook-Mills, J.M. Reactive oxygen species regulation of immune function. Mol. Immunol. 2002, 39, 497–498. [Google Scholar] [CrossRef]
- Irani, K. Oxidant signaling in vascular cell growth, death, and survival: A review of the roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic signaling. Circ. Res. 2000, 87, 179–183. [Google Scholar] [CrossRef]
- Reczek, C.R.; Chandel, N.S. Ros-dependent signal transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. Ros function in redox signaling and oxidative stress. Curr. Biol. CB 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Tsukasaki, Y.; Chan, W.C.; Le, J.P.; Kwok, M.L.; Zhou, J.; Natarajan, V.; Mostafazadeh, N.; Maienschein-Cline, M.; Papautsky, I.; et al. Trans-endothelial neutrophil migration activates bactericidal function via piezo1 mechanosensing. Immunity 2024, 57, 52–67.e10. [Google Scholar] [CrossRef]
- Kawahara, T.; Teshima, S.; Oka, A.; Sugiyama, T.; Kishi, K.; Rokutan, K. Type i Helicobacter pylori lipopolysaccharide stimulates toll-like receptor 4 and activates mitogen oxidase 1 in gastric pit cells. Infect. Immun. 2001, 69, 4382–4389. [Google Scholar] [CrossRef]
- Li, J.; Stouffs, M.; Serrander, L.; Banfi, B.; Bettiol, E.; Charnay, Y.; Steger, K.; Krause, K.H.; Jaconi, M.E. The NADPH oxidase nox4 drives cardiac differentiation: Role in regulating cardiac transcription factors and map kinase activation. Mol. Biol. Cell 2006, 17, 3978–3988. [Google Scholar] [CrossRef] [PubMed]
- Oakley, F.D.; Smith, R.L.; Engelhardt, J.F. Lipid rafts and caveolin-1 coordinate interleukin-1beta (il-1beta)-dependent activation of nfkappab by controlling endocytosis of Nox2 and il-1beta receptor 1 from the plasma membrane. J. Biol. Chem. 2009, 284, 33255–33264. [Google Scholar] [CrossRef] [PubMed]
- Prosser, B.L.; Ward, C.W.; Lederer, W.J. X-ros signaling: Rapid mechano-chemo transduction in heart. Science 2011, 333, 1440–1445. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef]
- Coyoy, A.; Valencia, A.; Guemez-Gamboa, A.; Moran, J. Role of NADPH oxidase in the apoptotic death of cultured cerebellar granule neurons. Free Radic. Biol. Med. 2008, 45, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Carbente, M.R.; Castro-Obregon, S.; Covarrubias, L.; Narvaez, V. Motoneuronal death during spinal cord development is mediated by oxidative stress. Cell Death Differ. 2005, 12, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Jezek, P.; Hlavata, L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. Int. J. Biochem. Cell Biol. 2005, 37, 2478–2503. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Sulik, K.K.; Chen, S.Y. The role of nox enzymes in ethanol-induced oxidative stress and apoptosis in mouse embryos. Toxicol. Lett. 2010, 193, 94–100. [Google Scholar] [CrossRef]
- Valencia, A.; Sapp, E.; Kimm, J.S.; McClory, H.; Reeves, P.B.; Alexander, J.; Ansong, K.A.; Masso, N.; Frosch, M.P.; Kegel, K.B.; et al. Elevated NADPH oxidase activity contributes to oxidative stress and cell death in huntington’s disease. Hum. Mol. Genet. 2013, 22, 1112–1131. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH oxidases: An overview from structure to innate immunity-associated pathologies. Cell. Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative stress in atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Batty, M.; Bennett, M.R.; Yu, E. The role of oxidative stress in atherosclerosis. Cells 2022, 11, 3843. [Google Scholar] [CrossRef]
- Elnakish, M.T.; Hassanain, H.H.; Janssen, P.M.; Angelos, M.G.; Khan, M. Emerging role of oxidative stress in metabolic syndrome and cardiovascular diseases: Important role of rac/NADPH oxidase. J. Pathol. 2013, 231, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Konior, A.; Schramm, A.; Czesnikiewicz-Guzik, M.; Guzik, T.J. NADPH oxidases in vascular pathology. Antioxid. Redox Signal. 2014, 20, 2794–2814. [Google Scholar] [CrossRef]
- Frederiksen, T.W.; Ramlau-Hansen, C.H.; Stokholm, Z.A.; Grynderup, M.B.; Hansen, A.M.; Kristiansen, J.; Vestergaard, J.M.; Bonde, J.P.; Kolstad, H.A. Noise-induced hearing loss—A preventable disease? Results of a 10-year longitudinal study of workers exposed to occupational noise. Noise Health 2017, 19, 103–111. [Google Scholar]
- Kuroda, J.; Sadoshima, J. NADPH oxidase and cardiac failure. J. Cardiovasc. Transl. Res. 2010, 3, 314–320. [Google Scholar] [CrossRef]
- Hahn, N.E.; Meischl, C.; Kawahara, T.; Musters, R.J.; Verhoef, V.M.; van der Velden, J.; Vonk, A.B.; Paulus, W.J.; van Rossum, A.C.; Niessen, H.W.; et al. Nox5 expression is increased in intramyocardial blood vessels and cardiomyocytes after acute myocardial infarction in humans. Am. J. Pathol. 2012, 180, 2222–2229. [Google Scholar] [CrossRef]
- Byrne, J.A.; Grieve, D.J.; Bendall, J.K.; Li, J.M.; Gove, C.; Lambeth, J.D.; Cave, A.C.; Shah, A.M. Contrasting roles of NADPH oxidase isoforms in pressure-overload versus angiotensin ii-induced cardiac hypertrophy. Circ. Res. 2003, 93, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Steinhorn, B.; Sorrentino, A.; Badole, S.; Bogdanova, Y.; Belousov, V.; Michel, T. Chemogenetic generation of hydrogen peroxide in the heart induces severe cardiac dysfunction. Nat. Commun. 2018, 9, 4044. [Google Scholar] [CrossRef]
- Giordano, F.J. Oxygen, oxidative stress, hypoxia, and heart failure. J. Clin. Investig. 2005, 115, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Sirker, A.; Zhang, M.; Shah, A.M. NADPH oxidases in cardiovascular disease: Insights from in vivo models and clinical studies. Basic Res. Cardiol. 2011, 106, 735–747. [Google Scholar] [CrossRef]
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative stress and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef]
- Lambeth, J.D. Nox enzymes, ros, and chronic disease: An example of antagonistic pleiotropy. Free Radic. Biol. Med. 2007, 43, 332–347. [Google Scholar] [CrossRef]
- Allen, C.L.; Bayraktutan, U. Oxidative stress and its role in the pathogenesis of ischaemic stroke. Int. J. Stroke 2009, 4, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Kim, J.H.; Lee, K.H.; Kim, H.Y.; Kim, Y.S.; Choi, W.S.; Lee, J. Role of neuronal NADPH oxidase 1 in the peri-infarct regions after stroke. PLoS ONE 2015, 10, e0116814. [Google Scholar] [CrossRef] [PubMed]
- Infanger, D.W.; Sharma, R.V.; Davisson, R.L. NADPH oxidases of the brain: Distribution, regulation, and function. Antioxid. Redox Signal. 2006, 8, 1583–1596. [Google Scholar] [CrossRef]
- Kahles, T.; Luedike, P.; Endres, M.; Galla, H.J.; Steinmetz, H.; Busse, R.; Neumann-Haefelin, T.; Brandes, R.P. NADPH oxidase plays a central role in blood-brain barrier damage in experimental stroke. Stroke 2007, 38, 3000–3006. [Google Scholar] [CrossRef] [PubMed]
- Ellis, E.A.; Guberski, D.L.; Somogyi-Mann, M.; Grant, M.B. Increased H2O2, vascular endothelial growth factor and receptors in the retina of the bbz/wor diabetic rat. Free Radic. Biol. Med. 2000, 28, 91–101. [Google Scholar] [CrossRef]
- Gardner, C.D.; Eguchi, S.; Reynolds, C.M.; Eguchi, K.; Frank, G.D.; Motley, E.D. Hydrogen peroxide inhibits insulin signaling in vascular smooth muscle cells. Exp. Biol. Med. 2003, 228, 836–842. [Google Scholar] [CrossRef]
- Nakabeppu, Y.; Sakumi, K.; Sakamoto, K.; Tsuchimoto, D.; Tsuzuki, T.; Nakatsu, Y. Mutagenesis and carcinogenesis caused by the oxidation of nucleic acids. Biol. Chem. 2006, 387, 373–379. [Google Scholar] [CrossRef]
- Roy, K.; Wu, Y.; Meitzler, J.L.; Juhasz, A.; Liu, H.; Jiang, G.; Lu, J.; Antony, S.; Doroshow, J.H. NADPH oxidases and cancer. Clin. Sci. 2015, 128, 863–875. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem.-Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Storz, P. Reactive oxygen species in tumor progression. Front. Biosci. A J. Virtual Libr. 2005, 10, 1881–1896. [Google Scholar] [CrossRef]
- Szatrowski, T.P.; Nathan, C.F. Production of large amounts of hydrogen peroxide by human tumor cells. Cancer Res. 1991, 51, 794–798. [Google Scholar] [PubMed]
- Han, M.; Zhang, T.; Yang, L.; Wang, Z.; Ruan, J.; Chang, X. Association between NADPH oxidase (NOX) and lung cancer: A systematic review and meta-analysis. J. Thorac. Dis. 2016, 8, 1704–1711. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Fukui, H.; Moraes, C.T. The mitochondrial impairment, oxidative stress and neurodegeneration connection: Reality or just an attractive hypothesis? Trends Neurosci. 2008, 31, 251–256. [Google Scholar] [CrossRef]
- Sompol, P.; Ittarat, W.; Tangpong, J.; Chen, Y.; Doubinskaia, I.; Batinic-Haberle, I.; Abdul, H.M.; Butterfield, D.A.; St Clair, D.K. A neuronal model of alzheimer’s disease: An insight into the mechanisms of oxidative stress-mediated mitochondrial injury. Neuroscience 2008, 153, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.T.; Sharma, R.; Lim, J.L.; Haider, L.; Frischer, J.M.; Drexhage, J.; Mahad, D.; Bradl, M.; van Horssen, J.; Lassmann, H. NADPH oxidase expression in active multiple sclerosis lesions in relation to oxidative tissue damage and mitochondrial injury. Brain A J. Neurol. 2012, 135, 886–899. [Google Scholar] [CrossRef]
- Tarafdar, A.; Pula, G. The role of NADPH oxidases and oxidative stress in neurodegenerative disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef]
- Yu, L.; Quinn, M.T.; Cross, A.R.; Dinauer, M.C. Gp91(phox) is the heme binding subunit of the superoxide-generating NADPH oxidase. Proc. Natl. Acad. Sci. USA 1998, 95, 7993–7998. [Google Scholar] [CrossRef]
- Quinn, M.T.; Mullen, M.L.; Jesaitis, A.J. Human neutrophil cytochrome-b contains multiple hemes—Evidence for heme associated with both subunits. J. Biol. Chem. 1992, 267, 7303–7309. [Google Scholar] [CrossRef]
- Nisimoto, Y.; Otsuka-Murakami, H.; Lambeth, D.J. Reconstitution of flavin-depleted neutrophil flavocytochrome b558 with 8-mercapto-fad and characterization of the flavin-reconstituted enzyme. J. Biol. Chem. 1995, 270, 16428–16434. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.D.; Parkar, M.H.; Verhoeven, A.J.; Levinsky, R.J.; Collins, M.K.; Kinnon, C. P22-phox-deficient chronic granulomatous disease: Reconstitution by retrovirus-mediated expression and identification of a biosynthetic intermediate of gp91-phox. Blood 1994, 84, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; DeLeo, F.R.; Biberstine-Kinkade, K.J.; Renee, J.; Nauseef, W.M.; Dinauer, M.C. Biosynthesis of flavocytochrome b558. Gp91(phox) is synthesized as a 65-kda precursor (p65) in the endoplasmic reticulum. J. Biol. Chem. 1999, 274, 4364–4369. [Google Scholar] [CrossRef]
- Yu, L.; Zhen, L.; Dinauer, M.C. Biosynthesis of the phagocyte NADPH oxidase cytochrome b558. Role of heme incorporation and heterodimer formation in maturation and stability of gp91phox and p22phox subunits. J. Biol. Chem. 1997, 272, 27288–27294. [Google Scholar] [CrossRef]
- Wallach, T.M.; Segal, A.W. Analysis of glycosylation sites on gp91phox, the flavocytochrome of the NADPH oxidase, by site-directed mutagenesis and translation in vitro. Biochem. J. 1997, 321 Pt 3, 583–585. [Google Scholar] [CrossRef]
- DeLeo, F.R.; Burritt, J.B.; Yu, L.; Jesaitis, A.J.; Dinauer, M.C.; Nauseef, W.M. Processing and maturation of flavocytochrome b558 include incorporation of heme as a prerequisite for heterodimer assembly. J. Biol. Chem. 2000, 275, 13986–13993. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.W.; West, I.; Wientjes, F.; Nugent, J.H.; Chavan, A.J.; Haley, B.; Garcia, R.C.; Rosen, H.; Scrace, G. Cytochrome b-245 is a flavocytochrome containing fad and the NADPH-binding site of the microbicidal oxidase of phagocytes. Biochem. J. 1992, 284 Pt 3, 781–788. [Google Scholar] [CrossRef]
- Rotrosen, D.; Yeung, C.L.; Leto, T.L.; Malech, H.L.; Kwong, C.H. Cytochrome b558: The flavin-binding component of the phagocyte NADPH oxidase. Science 1992, 256, 1459–1462. [Google Scholar] [CrossRef]
- Koshkin, V.; Lotan, O.; Pick, E. Electron transfer in the superoxide-generating NADPH oxidase complex reconstituted in vitro. Biochim. Biophys. Acta 1997, 1319, 139–146. [Google Scholar] [CrossRef]
- Koshkin, V.; Pick, E. Generation of superoxide by purified and relipidated cytochrome b559 in the absence of cytosolic activators. FEBS Lett. 1993, 327, 57–62. [Google Scholar] [CrossRef]
- Segal, A.W.; Jones, O.T. Novel cytochrome b system in phagocytic vacuoles of human granulocytes. Nature 1978, 276, 515–517. [Google Scholar] [CrossRef] [PubMed]
- Pember, S.O.; Heyl, B.L.; Kinkade, J.M., Jr.; Lambeth, J.D. Cytochrome b558 from (bovine) granulocytes. Partial purification from triton x-114 extracts and properties of the isolated cytochrome. J. Biol. Chem. 1984, 259, 10590–10595. [Google Scholar] [CrossRef] [PubMed]
- Knoller, S.; Shpungin, S.; Pick, E. The membrane-associated component of the amphiphile-activated, cytosol-dependent superoxide-forming NADPH oxidase of macrophages is identical to cytochrome b559. J. Biol. Chem. 1991, 266, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.R.; Higson, F.K.; Jones, O.T.; Harper, A.M.; Segal, A.W. The enzymic reduction and kinetics of oxidation of cytochrome b-245 of neutrophils. Biochem. J. 1982, 204, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.R.; Jones, O.T.; Harper, A.M.; Segal, A.W. Oxidation-reduction properties of the cytochrome b found in the plasma-membrane fraction of human neutrophils. A possible oxidase in the respiratory burst. Biochem. J. 1981, 194, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Ambasta, R.K.; Kumar, P.; Griendling, K.K.; Schmidt, H.H.; Busse, R.; Brandes, R.P. Direct interaction of the novel Nox proteins with p22phox is required for the formation of a functionally active NADPH oxidase. J. Biol. Chem. 2004, 279, 45935–45941. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Takeda, Y.; Iwasaki, Y.; Yoshizaki, F. Simultaneous estimation of geniposide and genipin in mouse plasma using high-performance liquid chromatography. Anal. Sci. Int. J. Jpn. Soc. Anal. Chem. 2001, 17, 1237–1239. [Google Scholar] [CrossRef]
- Rae, J.; Noack, D.; Heyworth, P.G.; Ellis, B.A.; Curnutte, J.T.; Cross, A.R. Molecular analysis of 9 new families with chronic granulomatous disease caused by mutations in cyba, the gene encoding p22(phox). Blood 2000, 96, 1106–1112. [Google Scholar] [CrossRef]
- Dahan, I.; Issaeva, I.; Gorzalczany, Y.; Sigal, N.; Hirshberg, M.; Pick, E. Mapping of functional domains in the p22(phox) subunit of flavocytochrome b(559) participating in the assembly of the NADPH oxidase complex by “peptide walking”. J. Biol. Chem. 2002, 277, 8421–8432. [Google Scholar] [CrossRef]
- Zhu, Y.; Marchal, C.C.; Casbon, A.J.; Stull, N.; von Lohneysen, K.; Knaus, U.G.; Jesaitis, A.J.; McCormick, S.; Nauseef, W.M.; Dinauer, M.C. Deletion mutagenesis of p22phox subunit of flavocytochrome b558: Identification of regions critical for gp91phox maturation and NADPH oxidase activity. J. Biol. Chem. 2006, 281, 30336–30346. [Google Scholar] [CrossRef]
- Groemping, Y.; Lapouge, K.; Smerdon, S.J.; Rittinger, K. Molecular basis of phosphorylation-induced activation of the NADPH oxidase. Cell 2003, 113, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Nobuhisa, I.; Takeya, R.; Ogura, K.; Ueno, N.; Kohda, D.; Inagaki, F.; Sumimoto, H. Activation of the superoxide-producing phagocyte NADPH oxidase requires co-operation between the tandem sh3 domains of p47phox in recognition of a polyproline type ii helix and an adjacent alpha-helix of p22phox. Biochem. J. 2006, 396, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Prior, K.K.; Leisegang, M.S.; Josipovic, I.; Lowe, O.; Shah, A.M.; Weissmann, N.; Schroder, K.; Brandes, R.P. Crispr/cas9-mediated knockout of p22phox leads to loss of nox1 and nox4, but not nox5 activity. Redox Biol. 2016, 9, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.W. The NADPH oxidase and chronic granulomatous disease. Mol. Med. Today 1996, 2, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Maly, F.E.; Schuerer-Maly, C.C.; Quilliam, L.; Cochrane, C.G.; Newburger, P.E.; Curnutte, J.T.; Gifford, M.; Dinauer, M.C. Restitution of superoxide generation in autosomal cytochrome-negative chronic granulomatous disease (a22(0) cgd)-derived b lymphocyte cell lines by transfection with p22phax cdna. J. Exp. Med. 1993, 178, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Banfi, B.; Jesaitis, A.J.; Dinauer, M.C.; Allen, L.A.; Nauseef, W.M. Critical roles for p22phox in the structural maturation and subcellular targeting of Nox3. Biochem. J. 2007, 403, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Martyn, K.D.; Frederick, L.M.; von Loehneysen, K.; Dinauer, M.C.; Knaus, U.G. Functional analysis of nox4 reveals unique characteristics compared to other NADPH oxidases. Cell. Signal. 2006, 18, 69–82. [Google Scholar] [CrossRef]
- Serrander, L.; Cartier, L.; Bedard, K.; Banfi, B.; Lardy, B.; Plastre, O.; Sienkiewicz, A.; Forro, L.; Schlegel, W.; Krause, K.H. Nox4 activity is determined by mrna levels and reveals a unique pattern of Ros generation. Biochem. J. 2007, 406, 105–114. [Google Scholar] [CrossRef]
- Von Lohneysen, K.; Noack, D.; Jesaitis, A.J.; Dinauer, M.C.; Knaus, U.G. Mutational analysis reveals distinct features of the nox4-p22 phox complex. J. Biol. Chem. 2008, 283, 35273–35282. [Google Scholar] [CrossRef]
- Magnani, F.; Nenci, S.; Millana Fananas, E.; Ceccon, M.; Romero, E.; Fraaije, M.W.; Mattevi, A. Crystal structures and atomic model of NADPH oxidase. Proc. Natl. Acad. Sci. USA 2017, 114, 6764–6769. [Google Scholar] [CrossRef]
- Maehara, Y.; Miyano, K.; Yuzawa, S.; Akimoto, R.; Takeya, R.; Sumimoto, H. A conserved region between the TPR and activation domains of p67phox participates in activation of the phagocyte NADPH oxidase. J. Biol. Chem. 2010, 285, 31435–31445. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.R. P40(phox) participates in the activation of NADPH oxidase by increasing the affinity of p47(phox) for flavocytochrome b(558). Biochem. J. 2000, 349, 113–117. [Google Scholar] [CrossRef]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef]
- Clark, R.A.; Epperson, T.K.; Valente, A.J. Mechanisms of activation of NADPH oxidases. Jpn. J. Infect. Dis. 2004, 57, S22–S23. [Google Scholar]
- Roos, D.; de Boer, M.; Kuribayashi, F.; Meischl, C.; Weening, R.S.; Segal, A.W.; Ahlin, A.; Nemet, K.; Hossle, J.P.; Bernatowska-Matuszkiewicz, E.; et al. Mutations in the x-linked and autosomal recessive forms of chronic granulomatous disease. Blood 1996, 87, 1663–1681. [Google Scholar] [CrossRef]
- Nunoi, H.; Rotrosen, D.; Gallin, J.I.; Malech, H.L. Two forms of autosomal chronic granulomatous disease lack distinct neutrophil cytosol factors. Science 1988, 242, 1298–1301. [Google Scholar] [CrossRef]
- Volpp, B.D.; Nauseef, W.M.; Clark, R.A. Two cytosolic neutrophil oxidase components absent in autosomal chronic granulomatous disease. Science 1988, 242, 1295–1297. [Google Scholar] [CrossRef]
- Leto, T.L.; Lomax, K.J.; Volpp, B.D.; Nunoi, H.; Sechler, J.M.; Nauseef, W.M.; Clark, R.A.; Gallin, J.I.; Malech, H.L. Cloning of a 67-kd neutrophil oxidase factor with similarity to a noncatalytic region of p60c-src. Science 1990, 248, 727–730. [Google Scholar] [CrossRef]
- Han, C.H.; Freeman, J.L.; Lee, T.; Motalebi, S.A.; Lambeth, J.D. Regulation of the neutrophil respiratory burst oxidase. Identification of an activation domain in p67(phox). J. Biol. Chem. 1998, 273, 16663–16668. [Google Scholar] [CrossRef] [PubMed]
- Nisimoto, Y.; Motalebi, S.; Han, C.H.; Lambeth, J.D. The p67(phox) activation domain regulates electron flow from nadph to flavin in flavocytochrome b(558). J. Biol. Chem. 1999, 274, 22999–23005. [Google Scholar] [CrossRef] [PubMed]
- Someya, A.; Nagaoka, I.; Yamashita, T. Purification of the 260 kda cytosolic complex involved in the superoxide production of guinea pig neutrophils. FEBS Lett. 1993, 330, 215–218. [Google Scholar] [CrossRef]
- Wientjes, F.B.; Hsuan, J.J.; Totty, N.F.; Segal, A.W. P40phox, a third cytosolic component of the activation complex of the nadph oxidase to contain src homology 3 domains. Biochem. J. 1993, 296 Pt 3, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Sathyamoorthy, M.; de Mendez, I.; Adams, A.G.; Leto, T.L. P40(phox) down-regulates nadph oxidase activity through interactions with its sh3 domain. J. Biol. Chem. 1997, 272, 9141–9146. [Google Scholar] [CrossRef] [PubMed]
- Kuribayashi, F.; Nunoi, H.; Wakamatsu, K.; Tsunawaki, S.; Sato, K.; Ito, T.; Sumimoto, H. The adaptor protein p40(phox) as a positive regulator of the superoxide-producing phagocyte oxidase. EMBO J. 2002, 21, 6312–6320. [Google Scholar] [CrossRef] [PubMed]
- Banfi, B.; Clark, R.A.; Steger, K.; Krause, K.H. Two novel proteins activate superoxide generation by the nadph oxidase nox1. J. Biol. Chem. 2003, 278, 3510–3513. [Google Scholar] [CrossRef] [PubMed]
- Ago, T.; Kuribayashi, F.; Hiroaki, H.; Takeya, R.; Ito, T.; Kohda, D.; Sumimoto, H. Phosphorylation of p47phox directs phox homology domain from sh3 domain toward phosphoinositides, leading to phagocyte nadph oxidase activation. Proc. Natl. Acad. Sci. USA 2003, 100, 4474–4479. [Google Scholar] [CrossRef] [PubMed]
- Abo, A.; Pick, E.; Hall, A.; Totty, N.; Teahan, C.G.; Segal, A.W. Activation of the nadph oxidase involves the small gtp-binding protein p21rac1. Nature 1991, 353, 668–670. [Google Scholar] [CrossRef]
- Jaffe, A.B.; Hall, A. Rho gtpases: Biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef]
- Heasman, S.J.; Ridley, A.J. Mammalian rho gtpases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef]
- Ago, T.; Nunoi, H.; Ito, T.; Sumimoto, H. Mechanism for phosphorylation-induced activation of the phagocyte NADPH oxidase protein p47(phox). Triple replacement of serines 303, 304, and 328 with aspartates disrupts the sh3 domain-mediated intramolecular interaction in p47(phox), thereby activating the oxidase. J. Biol. Chem. 1999, 274, 33644–33653. [Google Scholar]
- Yuzawa, S.; Ogura, K.; Horiuchi, M.; Suzuki, N.N.; Fujioka, Y.; Kataoka, M.; Sumimoto, H.; Inagaki, F. Solution structure of the tandem src homology 3 domains of p47phox in an autoinhibited form. J. Biol. Chem. 2004, 279, 29752–29760. [Google Scholar] [CrossRef]
- Inanami, O.; Johnson, J.L.; McAdara, J.K.; Benna, J.E.; Faust, L.R.; Newburger, P.E.; Babior, B.M. Activation of the leukocyte NADPH oxidase by phorbol ester requires the phosphorylation of p47phox on serine 303 or 304. J. Biol. Chem. 1998, 273, 9539–9543. [Google Scholar] [CrossRef]
- Johnson, J.L.; Park, J.W.; Benna, J.E.; Faust, L.P.; Inanami, O.; Babior, B.M. Activation of p47(phox), a cytosolic subunit of the leukocyte NADPH oxidase. Phosphorylation of ser-359 or ser-370 precedes phosphorylation at other sites and is required for activity. J. Biol. Chem. 1998, 273, 35147–35152. [Google Scholar] [CrossRef]
- El-Benna, J.; Dang, P.M.; Gougerot-Pocidalo, M.A.; Marie, J.C.; Braut-Boucher, F. P47phox, the phagocyte NADPH oxidase/nox2 organizer: Structure, phosphorylation and implication in diseases. Exp. Mol. Med. 2009, 41, 217–225. [Google Scholar] [CrossRef]
- Huang, J.; Kleinberg, M.E. Activation of the phagocyte NADPH oxidase protein p47(phox). Phosphorylation controls sh3 domain-dependent binding to p22(phox). J. Biol. Chem. 1999, 274, 19731–19737. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Babior, B.M. Activation of the leukocyte NADPH oxidase subunit p47phox by protein kinase c. A phosphorylation-dependent change in the conformation of the c-terminal end of p47phox. Biochemistry 1997, 36, 7474–7480. [Google Scholar] [CrossRef] [PubMed]
- Leto, T.L.; Adams, A.G.; de Mendez, I. Assembly of the phagocyte NADPH oxidase: Binding of src homology 3 domains to proline-rich targets. Proc. Natl. Acad. Sci. USA 1994, 91, 10650–10654. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, H.; Hata, K.; Mizuki, K.; Ito, T.; Kage, Y.; Sakaki, Y.; Fukumaki, Y.; Nakamura, M.; Takeshige, K. Assembly and activation of the phagocyte NADPH oxidase. Specific interaction of the n-terminal Src homology 3 domain of p47phox with p22phox is required for activation of the NADPH oxidase. J. Biol. Chem. 1996, 271, 22152–22158. [Google Scholar] [CrossRef] [PubMed]
- Shiose, A.; Sumimoto, H. Arachidonic acid and phosphorylation synergistically induce a conformational change of p47phox to activate the phagocyte NADPH oxidase. J. Biol. Chem. 2000, 275, 13793–13801. [Google Scholar] [CrossRef]
- Ogura, K.; Nobuhisa, I.; Yuzawa, S.; Takeya, R.; Torikai, S.; Saikawa, K.; Sumimoto, H.; Inagaki, F. Nmr solution structure of the tandem src homology 3 domains of p47phox complexed with a p22phox-derived proline-rich peptide. J. Biol. Chem. 2006, 281, 3660–3668. [Google Scholar] [CrossRef] [PubMed]
- De Mendez, I.; Homayounpour, N.; Leto, T.L. Specificity of p47phox sh3 domain interactions in NADPH oxidase assembly and activation. Mol. Cell. Biol. 1997, 17, 2177–2185. [Google Scholar] [CrossRef] [PubMed]
- Abramov, A.Y.; Jacobson, J.; Wientjes, F.; Hothersall, J.; Canevari, L.; Duchen, M.R. Expression and modulation of an NADPH oxidase in mammalian astrocytes. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 9176–9184. [Google Scholar] [CrossRef]
- Dekker, L.V.; Leitges, M.; Altschuler, G.; Mistry, N.; McDermott, A.; Roes, J.; Segal, A.W. Protein kinase c-beta contributes to NADPH oxidase activation in neutrophils. Biochem. J. 2000, 347 Pt 1, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Dang, P.M.; Cross, A.R.; Babior, B.M. Assembly of the neutrophil respiratory burst oxidase: A direct interaction between p67phox and cytochrome b558. Proc. Natl. Acad. Sci. USA 2001, 98, 3001–3005. [Google Scholar] [CrossRef]
- Frey, R.S.; Rahman, A.; Kefer, J.C.; Minshall, R.D.; Malik, A.B. Pkczeta regulates tnf-alpha-induced activation of NADPH oxidase in endothelial cells. Circ. Res. 2002, 90, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Leusen, J.H.; de Boer, M.; Bolscher, B.G.; Hilarius, P.M.; Weening, R.S.; Ochs, H.D.; Roos, D.; Verhoeven, A.J. A point mutation in gp91-phox of cytochrome b558 of the human NADPH oxidase leading to defective translocation of the cytosolic proteins p47-phox and p67-phox. J. Clin. Investig. 1994, 93, 2120–2126. [Google Scholar] [CrossRef]
- Massenet, C.; Chenavas, S.; Cohen-Addad, C.; Dagher, M.C.; Brandolin, G.; Pebay-Peyroula, E.; Fieschi, F. Effects of p47phox c terminus phosphorylations on binding interactions with p40phox and p67phox. Structural and functional comparison of p40phox and p67phox sh3 domains. J. Biol. Chem. 2005, 280, 13752–13761. [Google Scholar] [CrossRef]
- Mizuki, K.; Takeya, R.; Kuribayashi, F.; Nobuhisa, I.; Kohda, D.; Nunoi, H.; Takeshige, K.; Sumimoto, H. A region c-terminal to the proline-rich core of p47phox regulates activation of the phagocyte NADPH oxidase by interacting with the c-terminal sh3 domain of p67phox. Arch. Biochem. Biophys. 2005, 444, 185–194. [Google Scholar] [CrossRef]
- Finan, P.; Shimizu, Y.; Gout, I.; Hsuan, J.; Truong, O.; Butcher, C.; Bennett, P.; Waterfield, M.D.; Kellie, S. An sh3 domain and proline-rich sequence mediate an interaction between two components of the phagocyte NADPH oxidase complex. J. Biol. Chem. 1994, 269, 13752–13755. [Google Scholar] [CrossRef]
- Leusen, J.H.; Fluiter, K.; Hilarius, P.M.; Roos, D.; Verhoeven, A.J.; Bolscher, B.G. Interactions between the cytosolic components p47phox and p67phox of the human neutrophil NADPH oxidase that are not required for activation in the cell-free system. J. Biol. Chem. 1995, 270, 11216–11221. [Google Scholar] [CrossRef]
- Doussiere, J.; Pilloud, M.C.; Vignais, P.V. Cytosolic factors in bovine neutrophil oxidase activation. Partial purification and demonstration of translocation to a membrane fraction. Biochemistry 1990, 29, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.; Volpp, B.D.; Leidal, K.G.; Nauseef, W.M. Two cytosolic components of the human neutrophil respiratory burst oxidase translocate to the plasma membrane during cell activation. J. Clin. Investig. 1990, 85, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Kami, K.; Takeya, R.; Sumimoto, H.; Kohda, D. Diverse recognition of non-pxxp peptide ligands by the sh3 domains from p67(phox), grb2 and pex13p. EMBO J. 2002, 21, 4268–4276. [Google Scholar] [CrossRef]
- Koga, H.; Terasawa, H.; Nunoi, H.; Takeshige, K.; Inagaki, F.; Sumimoto, H. Tetratricopeptide repeat (tpr) motifs of p67(phox) participate in interaction with the small gtpase Rac and activation of the phagocyte NADPH oxidase. J. Biol. Chem. 1999, 274, 25051–25060. [Google Scholar] [CrossRef]
- Nisimoto, Y.; Freeman, J.L.; Motalebi, S.A.; Hirshberg, M.; Lambeth, J.D. Rac binding to p67(phox). Structural basis for interactions of the rac1 effector region and insert region with components of the respiratory burst oxidase. J. Biol. Chem. 1997, 272, 18834–18841. [Google Scholar] [CrossRef]
- Dusi, S.; Rossi, F. Activation of NADPH oxidase of human neutrophils involves the phosphorylation and the translocation of cytosolic p67phox. Biochem. J. 1993, 296 Pt 2, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Forbes, L.V.; Moss, S.J.; Segal, A.W. Phosphorylation of p67phox in the neutrophil occurs in the cytosol and is independent of p47phox. FEBS Lett. 1999, 449, 225–229. [Google Scholar] [CrossRef]
- Forbes, L.V.; Truong, O.; Wientjes, F.B.; Moss, S.J.; Segal, A.W. The major phosphorylation site of the NADPH oxidase component p67phox is thr233. Biochem. J. 1999, 338 Pt 1, 99–105. [Google Scholar] [CrossRef]
- Dang, P.M.; Morel, F.; Gougerot-Pocidalo, M.A.; El Benna, J. Phosphorylation of the NADPH oxidase component p67(phox) by erk2 and p38mapk: Selectivity of phosphorylated sites and existence of an intramolecular regulatory domain in the tetratricopeptide-rich region. Biochemistry 2003, 42, 4520–4526. [Google Scholar] [CrossRef]
- Heyworth, P.G.; Curnutte, J.T.; Nauseef, W.M.; Volpp, B.D.; Pearson, D.W.; Rosen, H.; Clark, R.A. Neutrophil nicotinamide adenine dinucleotide phosphate oxidase assembly. Translocation of p47-phox and p67-phox requires interaction between p47-phox and cytochrome b558. J. Clin. Investig. 1991, 87, 352–356. [Google Scholar] [CrossRef]
- Freeman, J.L.; Lambeth, J.D. NADPH oxidase activity is independent of p47phox in vitro. J. Biol. Chem. 1996, 271, 22578–22582. [Google Scholar] [CrossRef] [PubMed]
- Dusi, S.; Donini, M.; Rossi, F. Mechanisms of NADPH oxidase activation: Translocation of p40phox, rac1 and rac2 from the cytosol to the membranes in human neutrophils lacking p47phox or p67phox. Biochem. J. 1996, 314 Pt 2, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, T.; Geiszt, M.; Leto, T.L. Involvement of rac1 in activation of multicomponent nox1- and nox3-based NADPH oxidases. Mol. Cell. Biol. 2006, 26, 2160–2174. [Google Scholar] [CrossRef] [PubMed]
- Miyano, K.; Koga, H.; Minakami, R.; Sumimoto, H. The insert region of the Rac gtpases is dispensable for activation of superoxide-producing NADPH oxidases. Biochem. J. 2009, 422, 373–382. [Google Scholar] [CrossRef] [PubMed]
- De Leo, F.R.; Ulman, K.V.; Davis, A.R.; Jutila, K.L.; Quinn, M.T. Assembly of the human neutrophil NADPH oxidase involves binding of p67phox and flavocytochrome b to a common functional domain in p47phox. J. Biol. Chem. 1996, 271, 17013–17020. [Google Scholar] [CrossRef]
- Grizot, S.; Faure, J.; Fieschi, F.; Vignais, P.V.; Dagher, M.C.; Pebay-Peyroula, E. Crystal structure of the rac1-rhogdi complex involved in NADPH oxidase activation. Biochemistry 2001, 40, 10007–10013. [Google Scholar] [CrossRef]
- Bokoch, G.M.; Bohl, B.P.; Chuang, T.H. Guanine nucleotide exchange regulates membrane translocation of rac/rho gtp-binding proteins. J. Biol. Chem. 1994, 269, 31674–31679. [Google Scholar] [CrossRef]
- Le Cabec, V.; Mohn, H.; Gacon, G.; Maridonneau-Parini, I. The small gtp-binding protein Rac is not recruited to the plasma membrane upon NADPH oxidase activation in human neutrophils. Biochem. Biophys. Res. Commun. 1994, 198, 1216–1224. [Google Scholar] [CrossRef]
- Kreck, M.L.; Freeman, J.L.; Abo, A.; Lambeth, J.D. Membrane association of Rac is required for high activity of the respiratory burst oxidase. Biochemistry 1996, 35, 15683–15692. [Google Scholar] [CrossRef]
- Vergnaud, S.; Paclet, M.H.; El Benna, J.; Pocidalo, M.A.; Morel, F. Complementation of NADPH oxidase in p67-phox-deficient cgd patients p67-phox/p40-phox interaction. Eur. J. Biochem. 2000, 267, 1059–1067. [Google Scholar] [CrossRef]
- Paclet, M.H.; Coleman, A.W.; Vergnaud, S.; Morel, F. P67-phox-mediated NADPH oxidase assembly: Imaging of cytochrome b558 liposomes by atomic force microscopy. Biochemistry 2000, 39, 9302–9310. [Google Scholar] [CrossRef]
- Sumimoto, H.; Kage, Y.; Nunoi, H.; Sasaki, H.; Nose, T.; Fukumaki, Y.; Ohno, M.; Minakami, S.; Takeshige, K. Role of src homology 3 domains in assembly and activation of the phagocyte NADPH oxidase. Proc. Natl. Acad. Sci. USA 1994, 91, 5345–5349. [Google Scholar] [CrossRef]
- Sumimoto, H.; Kamakura, S.; Ito, T. Structure and function of the pb1 domain, a protein interaction module conserved in animals, fungi, amoebas, and plants. Sci. STKE Signal Transduct. Knowl. Environ. 2007, 2007, re6. [Google Scholar] [CrossRef]
- Katsuyama, M.; Matsuno, K.; Yabe-Nishimura, C. Physiological roles of nox/NADPH oxidase, the superoxide-generating enzyme. J. Clin. Biochem. Nutr. 2012, 50, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Bagaitkar, J.; Matute, J.D.; Austin, A.; Arias, A.A.; Dinauer, M.C. Activation of neutrophil respiratory burst by fungal particles requires phosphatidylinositol 3-phosphate binding to p40(phox) in humans but not in mice. Blood 2012, 120, 3385–3387. [Google Scholar] [CrossRef] [PubMed]
- Van de Geer, A.; Nieto-Patlan, A.; Kuhns, D.B.; Tool, A.T.; Arias, A.A.; Bouaziz, M.; de Boer, M.; Franco, J.L.; Gazendam, R.P.; van Hamme, J.L.; et al. Inherited p40phox deficiency differs from classic chronic granulomatous disease. J. Clin. Investig. 2018, 128, 3957–3975. [Google Scholar] [CrossRef]
- Ellson, C.D.; Davidson, K.; Ferguson, G.J.; O’Connor, R.; Stephens, L.R.; Hawkins, P.T. Neutrophils from p40phox-/- mice exhibit severe defects in NADPH oxidase regulation and oxidant-dependent bacterial killing. J. Exp. Med. 2006, 203, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- Bissonnette, S.A.; Glazier, C.M.; Stewart, M.Q.; Brown, G.E.; Ellson, C.D.; Yaffe, M.B. Phosphatidylinositol 3-phosphate-dependent and -independent functions of p40phox in activation of the neutrophil NADPH oxidase. J. Biol. Chem. 2008, 283, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Benard, V.; Bohl, B.P.; Bokoch, G.M. The molecular basis for adhesion-mediated suppression of reactive oxygen species generation by human neutrophils. J. Clin. Investig. 2003, 112, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- De Curtis, I. Functions of Rac gtpases during neuronal development. Dev. Neurosci. 2008, 30, 47–58. [Google Scholar] [CrossRef]
- Abo, A.; Webb, M.R.; Grogan, A.; Segal, A.W. Activation of NADPH oxidase involves the dissociation of p21rac from its inhibitory gdp/gtp exchange protein (rhogdi) followed by its translocation to the plasma membrane. Biochem. J. 1994, 298 Pt 3, 585–591. [Google Scholar] [CrossRef]
- Roberts, A.W.; Kim, C.; Zhen, L.; Lowe, J.B.; Kapur, R.; Petryniak, B.; Spaetti, A.; Pollock, J.D.; Borneo, J.B.; Bradford, G.B.; et al. Deficiency of the hematopoietic cell-specific rho family gtpase rac2 is characterized by abnormalities in neutrophil function and host defense. Immunity 1999, 10, 183–196. [Google Scholar] [CrossRef]
- Kim, C.; Dinauer, M.C. Rac2 is an essential regulator of neutrophil nicotinamide adenine dinucleotide phosphate oxidase activation in response to specific signaling pathways. J. Immunol. 2001, 166, 1223–1232. [Google Scholar] [CrossRef]
- Knaus, U.G.; Heyworth, P.G.; Kinsella, B.T.; Curnutte, J.T.; Bokoch, G.M. Purification and characterization of Rac 2. A cytosolic gtp-binding protein that regulates human neutrophil NADPH oxidase. J. Biol. Chem. 1992, 267, 23575–23582. [Google Scholar] [CrossRef]
- Knaus, U.G.; Heyworth, P.G.; Evans, T.; Curnutte, J.T.; Bokoch, G.M. Regulation of phagocyte oxygen radical production by the gtp-binding protein Rac 2. Science 1991, 254, 1512–1515. [Google Scholar] [CrossRef]
- Mizuno, T.; Kaibuchi, K.; Ando, S.; Musha, T.; Hiraoka, K.; Takaishi, K.; Asada, M.; Nunoi, H.; Matsuda, I.; Takai, Y. Regulation of the superoxide-generating NADPH oxidase by a small gtp-binding protein and its stimulatory and inhibitory gdp/gtp exchange proteins. J. Biol. Chem. 1992, 267, 10215–10218. [Google Scholar] [CrossRef] [PubMed]
- Heyworth, P.G.; Knaus, U.G.; Settleman, J.; Curnutte, J.T.; Bokoch, G.M. Regulation of NADPH oxidase activity by Rac gtpase activating protein(s). Mol. Biol. Cell 1993, 4, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.A.; Zheng, Y.; Cancelas, J.A. Rho gtpases and regulation of hematopoietic stem cell localization. Methods Enzymol. 2008, 439, 365–393. [Google Scholar] [PubMed]
- Miyano, K.; Ueno, N.; Takeya, R.; Sumimoto, H. Direct involvement of the small gtpase Rac in activation of the superoxide-producing NADPH oxidase Nox1. J. Biol. Chem. 2006, 281, 21857–21868. [Google Scholar] [CrossRef] [PubMed]
- Ambruso, D.R.; Knall, C.; Abell, A.N.; Panepinto, J.; Kurkchubasche, A.; Thurman, G.; Gonzalez-Aller, C.; Hiester, A.; deBoer, M.; Harbeck, R.J.; et al. Human neutrophil immunodeficiency syndrome is associated with an inhibitory Rac2 mutation. Proc. Natl. Acad. Sci. USA 2000, 97, 4654–4659. [Google Scholar] [CrossRef]
- Lapouge, K.; Smith, S.J.; Walker, P.A.; Gamblin, S.J.; Smerdon, S.J.; Rittinger, K. Structure of the tpr domain of p67phox in complex with rac.Gtp. Mol. Cell 2000, 6, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Kamiguti, A.S.; Serrander, L.; Lin, K.; Harris, R.J.; Cawley, J.C.; Allsup, D.J.; Slupsky, J.R.; Krause, K.H.; Zuzel, M. Expression and activity of nox5 in the circulating malignant b cells of hairy cell leukemia. J. Immunol. 2005, 175, 8424–8430. [Google Scholar] [CrossRef]
- Fortemaison, N.; Miot, F.; Dumont, J.E.; Dremier, S. Regulation of h2o2 generation in thyroid cells does not involve rac1 activation. Eur. J. Endocrinol. 2005, 152, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Glogauer, M.; Marchal, C.C.; Zhu, F.; Worku, A.; Clausen, B.E.; Foerster, I.; Marks, P.; Downey, G.P.; Dinauer, M.; Kwiatkowski, D.J. Rac1 deletion in mouse neutrophils has selective effects on neutrophil functions. J. Immunol. 2003, 170, 5652–5657. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Filippi, M.D.; Cancelas, J.A.; Siefring, J.E.; Williams, E.P.; Jasti, A.C.; Harris, C.E.; Lee, A.W.; Prabhakar, R.; Atkinson, S.J.; et al. Hematopoietic cell regulation by Rac1 and Rac2 guanosine triphosphatases. Science 2003, 302, 445–449. [Google Scholar] [CrossRef]
- Diekmann, D.; Abo, A.; Johnston, C.; Segal, A.W.; Hall, A. Interaction of Rac with p67phox and regulation of phagocytic NADPH oxidase activity. Science 1994, 265, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Diekmann, D.; Nobes, C.D.; Burbelo, P.D.; Abo, A.; Hall, A. Rac gtpase interacts with gaps and target proteins through multiple effector sites. EMBO J. 1995, 14, 5297–5305. [Google Scholar] [CrossRef] [PubMed]
- Kwong, C.H.; Adams, A.G.; Leto, T.L. Characterization of the effector-specifying domain of Rac involved in NADPH oxidase activation. J. Biol. Chem. 1995, 270, 19868–19872. [Google Scholar] [CrossRef]
- Freeman, J.L.; Abo, A.; Lambeth, J.D. Rac “insert region” is a novel effector region that is implicated in the activation of NADPH oxidase, but not pak65. J. Biol. Chem. 1996, 271, 19794–19801. [Google Scholar] [CrossRef]
- Heyworth, P.G.; Bohl, B.P.; Bokoch, G.M.; Curnutte, J.T. Rac translocates independently of the neutrophil NADPH oxidase components p47phox and p67phox. Evidence for its interaction with flavocytochrome b558. J. Biol. Chem. 1994, 269, 30749–30752. [Google Scholar] [CrossRef]
- Sarfstein, R.; Gorzalczany, Y.; Mizrahi, A.; Berdichevsky, Y.; Molshanski-Mor, S.; Weinbaum, C.; Hirshberg, M.; Dagher, M.C.; Pick, E. Dual role of Rac in the assembly of NADPH oxidase, tethering to the membrane and activation of p67phox: A study based on mutagenesis of p67phox-rac1 chimeras. J. Biol. Chem. 2004, 279, 16007–16016. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Takeshige, K.; Sumimoto, H. Roles for proline-rich regions of p47phox and p67phox in the phagocyte NADPH oxidase activation in vitro. Biochem. Biophys. Res. Commun. 1997, 241, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Diebold, B.A.; Bokoch, G.M. Molecular basis for Rac2 regulation of phagocyte NADPH oxidase. Nat. Immunol. 2001, 2, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Berdichevsky, Y.; Mizrahi, A.; Ugolev, Y.; Molshanski-Mor, S.; Pick, E. Tripartite chimeras comprising functional domains derived from the cytosolic NADPH oxidase components p47phox, p67phox, and rac1 elicit activator-independent superoxide production by phagocyte membranes: An essential role for anionic membrane phospholipids. J. Biol. Chem. 2007, 282, 22122–22139. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.Y.; Gianni, D.; Bohl, B.; Taylor, R.M.; Bokoch, G.M. Identification of a conserved Rac-binding site on NADPH oxidases supports a direct gtpase regulatory mechanism. J. Biol. Chem. 2008, 283, 12736–12746. [Google Scholar] [CrossRef] [PubMed]
- Hordijk, P.L. Regulation of NADPH oxidases: The role of Rac proteins. Circ. Res. 2006, 98, 453–462. [Google Scholar] [CrossRef]
- Iyer, S.S.; Pearson, D.W.; Nauseef, W.M.; Clark, R.A. Evidence for a readily dissociable complex of p47phox and p67phox in cytosol of unstimulated human neutrophils. J. Biol. Chem. 1994, 269, 22405–22411. [Google Scholar] [CrossRef]
- Wientjes, F.B.; Panayotou, G.; Reeves, E.; Segal, A.W. Interactions between cytosolic components of the NADPH oxidase: P40phox interacts with both p67phox and p47phox. Biochem. J. 1996, 317 Pt 3, 919–924. [Google Scholar] [CrossRef]
- Zhan, S.; Vazquez, N.; Zhan, S.; Wientjes, F.B.; Budarf, M.L.; Schrock, E.; Ried, T.; Green, E.D.; Chanock, S.J. Genomic structure, chromosomal localization, start of transcription, and tissue expression of the human p40-phox, a new component of the nicotinamide adenine dinucleotide phosphate-oxidase complex. Blood 1996, 88, 2714–2721. [Google Scholar] [CrossRef]
- Lapouge, K.; Smith, S.J.; Groemping, Y.; Rittinger, K. Architecture of the p40-p47-p67phox complex in the resting state of the NADPH oxidase. A central role for p67phox. J. Biol. Chem. 2002, 277, 10121–10128. [Google Scholar] [CrossRef]
- Quinn, M.T.; Evans, T.; Loetterle, L.R.; Jesaitis, A.J.; Bokoch, G.M. Translocation of Rac correlates with NADPH oxidase activation. Evidence for equimolar translocation of oxidase components. J. Biol. Chem. 1993, 268, 20983–20987. [Google Scholar] [CrossRef]
- Park, J.W.; Babior, B.M. The translocation of respiratory burst oxidase components from cytosol to plasma-membrane is regulated by guanine-nucleotides and diacylglycerol. J. Biol. Chem. 1992, 267, 19901–19906. [Google Scholar] [CrossRef]
- Dang, P.M.; Johnson, J.L.; Babior, B.M. Binding of nicotinamide adenine dinucleotide phosphate to the tetratricopeptide repeat domains at the n-terminus of p67phox, a subunit of the leukocyte nicotinamide adenine dinucleotide phosphate oxidase. Biochemistry 2000, 39, 3069–3075. [Google Scholar] [CrossRef]
- Uhlinger, D.J.; Tyagi, S.R.; Inge, K.L.; Lambeth, J.D. The respiratory burst oxidase of human neutrophils. Guanine nucleotides and arachidonate regulate the assembly of a multicomponent complex in a semirecombinant cell-free system. J. Biol. Chem. 1993, 268, 8624–8631. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Kaibuchi, K.; Sasaki, T.; Hiraoka, K.; Nishiyama, T.; Mizuno, T.; Asada, M.; Nunoi, H.; Matsuda, I.; Matsuura, Y. Post-translational processing of Rac p21s is important both for their interaction with the gdp/gtp exchange proteins and for their activation of NADPH oxidase. J. Biol. Chem. 1992, 267, 25709–25713. [Google Scholar] [CrossRef]
- Freeman, J.L.; Kreck, M.L.; Uhlinger, D.J.; Lambeth, J.D. Ras effector-homologue region on Rac regulates protein associations in the neutrophil respiratory burst oxidase complex. Biochemistry 1994, 33, 13431–13435. [Google Scholar] [CrossRef]
- Koshkin, V.; Lotan, O.; Pick, E. The cytosolic component p47(phox) is not a sine qua non participant in the activation of NADPH oxidase but is required for optimal superoxide production. J. Biol. Chem. 1996, 271, 30326–30329. [Google Scholar] [CrossRef]
- Nisimoto, Y.; Ogawa, H.; Miyano, K.; Tamura, M. Activation of the flavoprotein domain of gp91phox upon interaction with n-terminal p67phox (1–210) and the Rac complex. Biochemistry 2004, 43, 9567–9575. [Google Scholar] [CrossRef] [PubMed]
- Wallach, T.M.; Segal, A.W. Stoichiometry of the subunits of flavocytochrome b558 of the NADPH oxidase of phagocytes. Biochem. J. 1996, 320 Pt 1, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Hashida, S.; Yuzawa, S.; Suzuki, N.N.; Fujioka, Y.; Takikawa, T.; Sumimoto, H.; Inagaki, F.; Fujii, H. Binding of fad to cytochrome b558 is facilitated during activation of the phagocyte NADPH oxidase, leading to superoxide production. J. Biol. Chem. 2004, 279, 26378–26386. [Google Scholar] [CrossRef]
- Nisimoto, Y.; Diebold, B.A.; Cosentino-Gomes, D.; Lambeth, J.D. Nox4: A hydrogen peroxide-generating oxygen sensor. Biochemistry 2014, 53, 5111–5120. [Google Scholar] [CrossRef]
- Takac, I.; Schroder, K.; Zhang, L.; Lardy, B.; Anilkumar, N.; Lambeth, J.D.; Shah, A.M.; Morel, F.; Brandes, R.P. The e-loop is involved in hydrogen peroxide formation by the NADPH oxidase nox4. J. Biol. Chem. 2011, 286, 13304–13313. [Google Scholar] [CrossRef] [PubMed]
- Shiose, A.; Kuroda, J.; Tsuruya, K.; Hirai, M.; Hirakata, H.; Naito, S.; Hattori, M.; Sakaki, Y.; Sumimoto, H. A novel superoxide-producing NAD(P)H oxidase in kidney. J. Biol. Chem. 2001, 276, 1417–1423. [Google Scholar] [CrossRef]
- Ago, T.; Kitazono, T.; Ooboshi, H.; Iyama, T.; Han, Y.H.; Takada, J.; Wakisaka, M.; Ibayashi, S.; Utsumi, H.; Iida, M. Nox4 as the major catalytic component of an endothelial NAD(P)H oxidase. Circulation 2004, 109, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Ellmark, S.H.; Dusting, G.J.; Fui, M.N.; Guzzo-Pernell, N.; Drummond, G.R. The contribution of nox4 to NADPH oxidase activity in mouse vascular smooth muscle. Cardiovasc. Res. 2005, 65, 495–504. [Google Scholar] [CrossRef]
- Kawahara, T.; Ritsick, D.; Cheng, G.; Lambeth, J.D. Point mutations in the proline-rich region of p22phox are dominant inhibitors of nox1- and nox2-dependent reactive oxygen generation. J. Biol. Chem. 2005, 280, 31859–31869. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, J.; Nakagawa, K.; Yamasaki, T.; Nakamura, K.; Takeya, R.; Kuribayashi, F.; Imajoh-Ohmi, S.; Igarashi, K.; Shibata, Y.; Sueishi, K.; et al. The superoxide-producing NAD(P)H oxidase nox4 in the nucleus of human vascular endothelial cells. Genes Cells Devoted Mol. Cell. Mech. 2005, 10, 1139–1151. [Google Scholar] [CrossRef]
- Amanso, A.M.; Debbas, V.; Laurindo, F.R. Proteasome inhibition represses unfolded protein response and nox4, sensitizing vascular cells to endoplasmic reticulum stress-induced death. PLoS ONE 2011, 6, e14591. [Google Scholar] [CrossRef]
- Lu, X.; Guo, X.; Wassall, C.D.; Kemple, M.D.; Unthank, J.L.; Kassab, G.S. Reactive oxygen species cause endothelial dysfunction in chronic flow overload. J. Appl. Physiol. 2011, 110, 520–527. [Google Scholar] [CrossRef]
- Ahmad, M.; Kelly, M.R.; Zhao, X.; Kandhi, S.; Wolin, M.S. Roles for nox4 in the contractile response of bovine pulmonary arteries to hypoxia. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1879–H1888. [Google Scholar] [CrossRef]
- Pendyala, S.; Moitra, J.; Kalari, S.; Kleeberger, S.R.; Zhao, Y.; Reddy, S.P.; Garcia, J.G.; Natarajan, V. Nrf2 regulates hyperoxia-induced nox4 expression in human lung endothelium: Identification of functional antioxidant response elements on the nox4 promoter. Free Radic. Biol. Med. 2011, 50, 1749–1759. [Google Scholar] [CrossRef]
- Ben Mkaddem, S.; Pedruzzi, E.; Werts, C.; Coant, N.; Bens, M.; Cluzeaud, F.; Goujon, J.M.; Ogier-Denis, E.; Vandewalle, A. Heat shock protein gp96 and NAD(P)H oxidase 4 play key roles in toll-like receptor 4-activated apoptosis during renal ischemia/reperfusion injury. Cell Death Differ. 2010, 17, 1474–1485. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Hou, X.; Jourd’heuil, D.; Weisbrod, R.M.; Cohen, R.A. Upregulation of nox4 by tgfbeta1 oxidizes serca and inhibits no in arterial smooth muscle of the prediabetic zucker rat. Circ. Res. 2010, 107, 975–983. [Google Scholar] [CrossRef]
- St Hilaire, C.; Koupenova, M.; Carroll, S.H.; Smith, B.D.; Ravid, K. Tnf-alpha upregulates the a2b adenosine receptor gene: The role of NAD(P)H oxidase 4. Biochem. Biophys. Res. Commun. 2008, 375, 292–296. [Google Scholar] [CrossRef]
- Mahadev, K.; Motoshima, H.; Wu, X.; Ruddy, J.M.; Arnold, R.S.; Cheng, G.; Lambeth, J.D.; Goldstein, B.J. The NAD(P)H oxidase homolog nox4 modulates insulin-stimulated generation of H2O2 and plays an integral role in insulin signal transduction. Mol. Cell. Biol. 2004, 24, 1844–1854. [Google Scholar] [CrossRef]
- Park, H.S.; Jung, H.Y.; Park, E.Y.; Kim, J.; Lee, W.J.; Bae, Y.S. Cutting edge: Direct interaction of tlr4 with NAD(P)H oxidase 4 isozyme is essential for lipopolysaccharide-induced production of reactive oxygen species and activation of nf-kappa b. J. Immunol. 2004, 173, 3589–3593. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Rodriguez-Belmonte, E.M.; Mazloum, N.; Xie, B.; Lee, M.Y. Identification of a novel protein, pdip38, that interacts with the p50 subunit of DNA polymerase delta and proliferating cell nuclear antigen. J. Biol. Chem. 2003, 278, 10041–10047. [Google Scholar] [CrossRef] [PubMed]
- Xie, B.; Li, H.; Wang, Q.; Xie, S.; Rahmeh, A.; Dai, W.; Lee, M.Y. Further characterization of human DNA polymerase delta interacting protein 38. J. Biol. Chem. 2005, 280, 22375–22384. [Google Scholar] [CrossRef]
- Miller, F.J., Jr. NADPH oxidase 4: Walking the walk with poldip2. Circ. Res. 2009, 105, 209–210. [Google Scholar] [CrossRef]
- Lyle, A.N.; Deshpande, N.N.; Taniyama, Y.; Seidel-Rogol, B.; Pounkova, L.; Du, P.; Papaharalambus, C.; Lassegue, B.; Griendling, K.K. Poldip2, a novel regulator of nox4 and cytoskeletal integrity in vascular smooth muscle cells. Circ. Res. 2009, 105, 249–259. [Google Scholar] [CrossRef]
- Suzuki, Y.; Hattori, K.; Hamanaka, J.; Murase, T.; Egashira, Y.; Mishiro, K.; Ishiguro, M.; Tsuruma, K.; Hirose, Y.; Tanaka, H.; et al. Pharmacological inhibition of tlr4-nox4 signal protects against neuronal death in transient focal ischemia. Sci. Rep. 2012, 2, 896. [Google Scholar] [CrossRef] [PubMed]
- Janiszewski, M.; Lopes, L.R.; Carmo, A.O.; Pedro, M.A.; Brandes, R.P.; Santos, C.X.; Laurindo, F.R. Regulation of NAD(P)H oxidase by associated protein disulfide isomerase in vascular smooth muscle cells. J. Biol. Chem. 2005, 280, 40813–40819. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Anagnostopoulou, A.; Rios, F.; Montezano, A.C.; Camargo, L.L. Nox5: Molecular biology and pathophysiology. Exp. Physiol. 2019, 104, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Banfi, B.; Malgrange, B.; Knisz, J.; Steger, K.; Dubois-Dauphin, M.; Krause, K.H. Nox3, a superoxide-generating NADPH oxidase of the inner ear. J. Biol. Chem. 2004, 279, 46065–46072. [Google Scholar] [CrossRef] [PubMed]
- Luxen, S.; Belinsky, S.A.; Knaus, U.G. Silencing of duox NADPH oxidases by promoter hypermethylation in lung cancer. Cancer Res. 2008, 68, 1037–1045. [Google Scholar] [CrossRef]
- Grasberger, H.; Refetoff, S. Identification of the maturation factor for dual oxidase. Evolution of an eukaryotic operon equivalent. J. Biol. Chem. 2006, 281, 18269–18272. [Google Scholar] [CrossRef] [PubMed]
- De Deken, X.; Wang, D.; Dumont, J.E.; Miot, F. Characterization of thox proteins as components of the thyroid h(2)o(2)-generating system. Exp. Cell Res. 2002, 273, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the roscue: Mechanisms of NADPH oxidase activation and bacterial resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Belambri, S.A.; Rolas, L.; Raad, H.; Hurtado-Nedelec, M.; Dang, P.M.; El-Benna, J. NADPH oxidase activation in neutrophils: Role of the phosphorylation of its subunits. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12951. [Google Scholar] [CrossRef]
- Vermot, A.; Petit-Hartlein, I.; Smith, S.M.E.; Fieschi, F. NADPH oxidases (nox): An overview from discovery, molecular mechanisms to physiology and pathology. Antioxidants 2021, 10, 890. [Google Scholar] [CrossRef]
- Lambeth, J.D.; Cheng, G.; Arnold, R.S.; Edens, W.A. Novel homologs of gp91phox. Trends Biochem. Sci. 2000, 25, 459–461. [Google Scholar] [CrossRef]
- Kikuchi, H.; Hikage, M.; Miyashita, H.; Fukumoto, M. NADPH oxidase subunit, gp91(phox) homologue, preferentially expressed in human colon epithelial cells. Gene 2000, 254, 237–243. [Google Scholar] [CrossRef]
- Dupuy, C.; Pomerance, M.; Ohayon, R.; Noel-Hudson, M.S.; Deme, D.; Chaaraoui, M.; Francon, J.; Virion, A. Thyroid oxidase (thox2) gene expression in the rat thyroid cell line frtl-5. Biochem. Biophys. Res. Commun. 2000, 277, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Lalucque, H.; Silar, P. NADPH oxidase: An enzyme for multicellularity? Trends Microbiol. 2003, 11, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.G.; Ansorena, E.; Izal, I.; Zalba, G.; de Miguel, C.; Milagro, F.I. Structure, regulation, and physiological functions of NADPH oxidase 5 (nox5). J. Physiol. Biochem. 2023, 79, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Geiszt, M.; Lekstrom, K.; Witta, J.; Leto, T.L. Proteins homologous to p47phox and p67phox support superoxide production by NAD(P)H oxidase 1 in colon epithelial cells. J. Biol. Chem. 2003, 278, 20006–20012. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.C.; Bikker, H.; Kempers, M.J.; van Trotsenburg, A.S.; Baas, F.; de Vijlder, J.J.; Vulsma, T.; Ris-Stalpers, C. Inactivating mutations in the gene for thyroid oxidase 2 (thox2) and congenital hypothyroidism. N. Engl. J. Med. 2002, 347, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Lambeth, J.D. Noxo1, regulation of lipid binding, localization, and activation of nox1 by the phox homology (px) domain. J. Biol. Chem. 2004, 279, 4737–4742. [Google Scholar] [CrossRef] [PubMed]
- Paffenholz, R.; Bergstrom, R.A.; Pasutto, F.; Wabnitz, P.; Munroe, R.J.; Jagla, W.; Heinzmann, U.; Marquardt, A.; Bareiss, A.; Laufs, J.; et al. Vestibular defects in head-tilt mice result from mutations in Nox3, encoding an NADPH oxidase. Genes Dev. 2004, 18, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Takeya, R.; Sumimoto, H. Regulation of novel superoxide-producing NAD(P)H oxidases. Antioxid. Redox Signal. 2006, 8, 1523–1532. [Google Scholar] [CrossRef]
- Cheng, G.; Ritsick, D.; Lambeth, J.D. Nox3 regulation by noxo1, p47phox, and p67phox. J. Biol. Chem. 2004, 279, 34250–34255. [Google Scholar] [CrossRef]
- Babior, B.M.; Curnutte, J.T.; McMurrich, B.J. The particulate superoxide-forming system from human neutrophils. Properties of the system and further evidence supporting its participation in the respiratory burst. J. Clin. Investig. 1976, 58, 989–996. [Google Scholar] [CrossRef]
- Dewald, B.; Baggiolini, M.; Curnutte, J.T.; Babior, B.M. Subcellular localization of the superoxide-forming enzyme in human neutrophils. J. Clin. Investig. 1979, 63, 21–29. [Google Scholar] [CrossRef]
- Hiroaki, H.; Ago, T.; Ito, T.; Sumimoto, H.; Kohda, D. Solution structure of the px domain, a target of the sh3 domain. Nat. Struct. Biol. 2001, 8, 526–530. [Google Scholar] [CrossRef]
- Bravo, J.; Karathanassis, D.; Pacold, C.M.; Pacold, M.E.; Ellson, C.D.; Anderson, K.E.; Butler, P.J.; Lavenir, I.; Perisic, O.; Hawkins, P.T.; et al. The crystal structure of the px domain from p40(phox) bound to phosphatidylinositol 3-phosphate. Mol. Cell 2001, 8, 829–839. [Google Scholar] [CrossRef]
- Ponting, C.P. Novel domains in NADPH oxidase subunits, sorting nexins, and Ptdins 3-kinases: Binding partners of SH3 domains? Protein Sci. A Publ. Protein Soc. 1996, 5, 2353–2357. [Google Scholar] [CrossRef] [PubMed]
- Miyano, K.; Sumimoto, H. Role of the small gtpase Rac in p22phox-dependent NADPH oxidases. Biochimie 2007, 89, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- De Mendez, I.; Garrett, M.C.; Adams, A.G.; Leto, T.L. Role of p67-phox sh3 domains in assembly of the NADPH oxidase system. J. Biol. Chem. 1994, 269, 16326–16332. [Google Scholar] [CrossRef] [PubMed]
- Maehara, Y.; Miyano, K.; Sumimoto, H. Role for the first sh3 domain of p67phox in activation of superoxide-producing NADPH oxidases. Biochem. Biophys. Res. Commun. 2009, 379, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Taura, M.; Miyano, K.; Minakami, R.; Kamakura, S.; Takeya, R.; Sumimoto, H. A region n-terminal to the tandem sh3 domain of p47phox plays a crucial role in the activation of the phagocyte NADPH oxidase. Biochem. J. 2009, 419, 329–338. [Google Scholar] [CrossRef]
- Ueno, N.; Takeya, R.; Miyano, K.; Kikuchi, H.; Sumimoto, H. The NADPH oxidase nox3 constitutively produces superoxide in a p22phox-dependent manner: Its regulation by oxidase organizers and activators. J. Biol. Chem. 2005, 280, 23328–23339. [Google Scholar] [CrossRef]
- Ueyama, T.; Lekstrom, K.; Tsujibe, S.; Saito, N.; Leto, T.L. Subcellular localization and function of alternatively spliced noxo1 isoforms. Free Radic. Biol. Med. 2007, 42, 180–190. [Google Scholar] [CrossRef]
- Miyano, K.; Sumimoto, H. N-linked glycosylation of the superoxide-producing NADPH oxidase Nox1. Biochem. Biophys. Res. Commun. 2014, 443, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Gianni, D.; Diaz, B.; Taulet, N.; Fowler, B.; Courtneidge, S.A.; Bokoch, G.M. Novel p47(phox)-related organizers regulate localized NADPH oxidase 1 (Nox1) activity. Sci. Signal. 2009, 2, ra54. [Google Scholar] [CrossRef] [PubMed]
- Lock, P.; Abram, C.L.; Gibson, T.; Courtneidge, S.A. A new method for isolating tyrosine kinase substrates used to identify fish, an sh3 and px domain-containing protein, and src substrate. EMBO J. 1998, 17, 4346–4357. [Google Scholar] [CrossRef]
- Courtneidge, S.A. Isolation of novel src substrates. Biochem. Soc. Trans. 2003, 31, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Courtneidge, S.A.; Azucena, E.F.; Pass, I.; Seals, D.F.; Tesfay, L. The Src substrate tks5, podosomes (invadopodia), and cancer cell invasion. Cold Spring Harb. Symp. Quant. Biol. 2005, 70, 167–171. [Google Scholar] [CrossRef]
- Diaz, B.; Shani, G.; Pass, I.; Anderson, D.; Quintavalle, M.; Courtneidge, S.A. Tks5-dependent, nox-mediated generation of reactive oxygen species is necessary for invadopodia formation. Sci. Signal. 2009, 2, ra53. [Google Scholar] [CrossRef]
- Leto, T.L.; Morand, S.; Hurt, D.; Ueyama, T. Targeting and regulation of reactive oxygen species generation by Nox family NADPH oxidases. Antioxid. Redox Signal. 2009, 11, 2607–2619. [Google Scholar] [CrossRef]
- Geiszt, M. NADPH oxidases: New kids on the block. Cardiovasc. Res. 2006, 71, 289–299. [Google Scholar] [CrossRef]
- Rousset, F.; Kokje, V.B.C.; Sipione, R.; Schmidbauer, D.; Nacher-Soler, G.; Ilmjarv, S.; Coelho, M.; Fink, S.; Voruz, F.; El Chemaly, A.; et al. Intrinsically self-renewing neuroprogenitors from the a/j mouse spiral ganglion as virtually unlimited source of mature auditory neurons. Front. Cell. Neurosci. 2020, 14, 395. [Google Scholar] [CrossRef]
- Herb, M.; Gluschko, A.; Wiegmann, K.; Farid, A.; Wolf, A.; Utermohlen, O.; Krut, O.; Kronke, M.; Schramm, M. Mitochondrial reactive oxygen species enable proinflammatory signaling through disulfide linkage of nemo. Sci. Signal. 2019, 12, eaar5926. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ros in macrophages and antimicrobial immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Zhang, Y.; Shan, P.; Srivastava, A.; Jiang, G.; Zhang, X.; Lee, P.J. An endothelial hsp70-tlr4 axis limits Nox3 expression and protects against oxidant injury in lungs. Antioxid. Redox Signal. 2016, 24, 991–1012. [Google Scholar] [CrossRef]
- Li, L.; He, Q.; Huang, X.; Man, Y.; Zhou, Y.; Wang, S.; Wang, J.; Li, J. Nox3-derived reactive oxygen species promote tnf-alpha-induced reductions in hepatocyte glycogen levels via a JNK pathway. FEBS Lett. 2010, 584, 995–1000. [Google Scholar] [CrossRef]
- Delmaghani, S.; El-Amraoui, A. Inner ear gene therapies take off: Current promises and future challenges. J. Clin. Med. 2020, 9, 2309. [Google Scholar] [CrossRef]
- Frolenkov, G.I.; Belyantseva, I.A.; Friedman, T.B.; Griffith, A.J. Genetic insights into the morphogenesis of inner ear hair cells. Nat. Rev. Genet. 2004, 5, 489–498. [Google Scholar] [CrossRef] [PubMed]
- LeMasurier, M.; Gillespie, P.G. Hair-cell mechanotransduction and cochlear amplification. Neuron 2005, 48, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Driver, E.C.; Kelley, M.W. Development of the cochlea. Development 2020, 147, dev162263. [Google Scholar] [CrossRef] [PubMed]
- Anniko, M. Development of otoconia. Am. J. Otolaryngol. 1980, 1, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chang, R. Anatomy of the vestibular system: A review. NeuroRehabilitation 2013, 32, 437–443. [Google Scholar] [CrossRef]
- Fife, T.D. Chapter 2—overview of anatomy and physiology of the vestibular system. In Handbook of Clinical Neurophysiology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 9, pp. 5–17. [Google Scholar]
- Rousset, F.; Nacher-Soler, G.; Coelho, M.; Ilmjarv, S.; Kokje, V.B.C.; Marteyn, A.; Cambet, Y.; Perny, M.; Roccio, M.; Jaquet, V.; et al. Redox activation of excitatory pathways in auditory neurons as mechanism of age-related hearing loss. Redox Biol. 2020, 30, 101434. [Google Scholar] [CrossRef]
- Chen, P.; Chai, Y.; Liu, H.; Li, G.; Wang, L.; Yang, T.; Wu, H. Postnatal development of microglia-like cells in mouse cochlea. Neural Plast. 2018, 2018, 1970150. [Google Scholar] [CrossRef]
- Sun, S.; Yu, H.; Yu, H.; Honglin, M.; Ni, W.; Zhang, Y.; Guo, L.; He, Y.; Xue, Z.; Ni, Y.; et al. Inhibition of the activation and recruitment of microglia-like cells protects against neomycin-induced ototoxicity. Mol. Neurobiol. 2015, 51, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Morioka, S.; Sakaguchi, H.; Yamaguchi, T.; Ninoyu, Y.; Mohri, H.; Nakamura, T.; Hisa, Y.; Ogita, K.; Saito, N.; Ueyama, T. Hearing vulnerability after noise exposure in a mouse model of reactive oxygen species overproduction. J. Neurochem. 2018, 146, 459–473. [Google Scholar] [CrossRef]
- Jones, S.M.; Erway, L.C.; Bergstrom, R.A.; Schimenti, J.C.; Jones, T.A. Vestibular responses to linear acceleration are absent in otoconia-deficient c57bl/6jei-het mice. Hear. Res. 1999, 135, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.A.; Jones, S.M. Short latency compound action potentials from mammalian gravity receptor organs. Hear. Res. 1999, 136, 75–85. [Google Scholar] [CrossRef]
- Jones, S.M.; Erway, L.C.; Johnson, K.R.; Yu, H.; Jones, T.A. Gravity receptor function in mice with graded otoconial deficiencies. Hear. Res. 2004, 191, 34–40. [Google Scholar] [CrossRef]
- Thalmann, R.; Ignatova, E.; Kachar, B.; Ornitz, D.M.; Thalmann, I. Development and maintenance of otoconia: Biochemical considerations. Ann. N. Y. Acad. Sci. 2001, 942, 162–178. [Google Scholar] [CrossRef]
- Fernandez, C.; Goldberg, J.M. Physiology of peripheral neurons innervating otolith organs of the squirrel monkey. I. Response to static tilts and to long-duration centrifugal force. J. Neurophysiol. 1976, 39, 970–984. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M.; Fernandez, C. Physiology of peripheral neurons innervating semicircular canals of the squirrel monkey. I. Resting discharge and response to constant angular accelerations. J. Neurophysiol. 1971, 34, 635–660. [Google Scholar] [CrossRef]
- Jones, T.A.; Jones, S.M. Spontaneous activity in the statoacoustic ganglion of the chicken embryo. J. Neurophysiol. 2000, 83, 1452–1468. [Google Scholar] [CrossRef]
- Adrian, E.D. Discharges from vestibular receptors in the cat. J. Physiol. 1943, 101, 389–407. [Google Scholar] [CrossRef] [PubMed]
- Annoni, J.M.; Cochran, S.L.; Precht, W. Pharmacology of the vestibular hair cell-afferent fiber synapse in the frog. J. Neurosci. Off. J. Soc. Neurosci. 1984, 4, 2106–2116. [Google Scholar] [CrossRef] [PubMed]
- Flock, A.; Russell, I. Inhibition by efferent nerve fibres: Action on hair cells and afferent synaptic transmission in the lateral line canal organ of the burbot lota lota. J. Physiol. 1976, 257, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Ishii, Y. Neurophysiological studies on hearing in goldfish. J. Neurophysiol. 1967, 30, 1377–1403. [Google Scholar] [CrossRef]
- Furukawa, T.; Ishii, Y.; Matsuura, S. Synaptic delay and time course of postsynaptic potentials at the junction between hair cells and eighth nerve fibers in the goldfish. Jpn. J. Physiol. 1972, 22, 617–635. [Google Scholar] [CrossRef]
- Fuchs, P.A.; Glowatzki, E. Synaptic studies inform the functional diversity of cochlear afferents. Hear. Res. 2015, 330, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Sondag, H.N.; de Jong, H.A.; van Marle, J.; Oosterveld, W.J. Effects of sustained acceleration on the morphological properties of otoconia in hamsters. Acta Oto-Laryngol. 1995, 115, 227–230. [Google Scholar] [CrossRef]
- Suzuki, H.; Ikeda, K.; Takasaka, T. Biological characteristics of the globular substance in the otoconial membrane of the guinea pig. Hear. Res. 1995, 90, 212–218. [Google Scholar] [CrossRef]
- Tu, Y.H.; Cooper, A.J.; Teng, B.; Chang, R.B.; Artiga, D.J.; Turner, H.N.; Mulhall, E.M.; Ye, W.; Smith, A.D.; Liman, E.R. An evolutionarily conserved gene family encodes proton-selective ion channels. Science 2018, 359, 1047–1050. [Google Scholar] [CrossRef]
- Ramsey, I.S.; DeSimone, J.A. Otopetrin-1: A sour-tasting proton channel. J. Gen. Physiol. 2018, 150, 379–382. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Bohne, B.A.; Thalmann, I.; Harding, G.W.; Thalmann, R. Otoconial agenesis in tilted mutant mice. Hear. Res. 1998, 122, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Ying, H.C.; Hurle, B.; Wang, Y.; Bohne, B.A.; Wuerffel, M.K.; Ornitz, D.M. High-resolution mapping of tlt, a mouse mutant lacking otoconia. Mamm. Genome 1999, 10, 544–548. [Google Scholar] [CrossRef]
- Hurle, B.; Ignatova, E.; Massironi, S.M.; Mashimo, T.; Rios, X.; Thalmann, I.; Thalmann, R.; Ornitz, D.M. Non-syndromic vestibular disorder with otoconial agenesis in tilted/mergulhador mice caused by mutations in otopetrin 1. Hum. Mol. Genet. 2003, 12, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Hurle, B.; Lane, K.; Kenney, J.; Tarantino, L.M.; Bucan, M.; Brownstein, B.H.; Ornitz, D.M. Physical mapping of the mouse tilted locus identifies an association between human deafness loci dfna6/14 and vestibular system development. Genomics 2001, 77, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Everett, L.A.; Belyantseva, I.A.; Noben-Trauth, K.; Cantos, R.; Chen, A.; Thakkar, S.I.; Hoogstraten-Miller, S.L.; Kachar, B.; Wu, D.K.; Green, E.D. Targeted disruption of mouse pds provides insight about the inner-ear defects encountered in pendred syndrome. Hum. Mol. Genet. 2001, 10, 153–161. [Google Scholar] [CrossRef]
- Huang, S.; Qian, S. Advances in otolith-related protein research. Front. Neurosci. 2022, 16, 956200. [Google Scholar] [CrossRef]
- Lundberg, Y.W.; Xu, Y. Proteins involved in otoconia formation and maintenance. Otolaryngology 2012, 1, 3–22. [Google Scholar]
- Simmler, M.C.; Cohen-Salmon, M.; El-Amraoui, A.; Guillaud, L.; Benichou, J.C.; Petit, C.; Panthier, J.J. Targeted disruption of otog results in deafness and severe imbalance. Nat. Genet. 2000, 24, 139–143. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, H.; Yang, H.; Zhao, X.; Lovas, S.; Lundberg, Y.W. Expression, functional, and structural analysis of proteins critical for otoconia development. Dev. Dyn. 2010, 239, 2659–2673. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Thalmann, I.; Thalmann, R.; Ornitz, D.M. Mapping the mouse otoconin-90 (oc90) gene to chromosome 15. Genomics 1999, 58, 214–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kowalski, P.E.; Thalmann, I.; Ornitz, D.M.; Mager, D.L.; Thalmann, R. Otoconin-90, the mammalian otoconial matrix protein, contains two domains of homology to secretory phospholipase a2. Proc. Natl. Acad. Sci. USA 1998, 95, 15345–15350. [Google Scholar] [CrossRef] [PubMed]
- Verpy, E.; Leibovici, M.; Petit, C. Characterization of otoconin-95, the major protein of murine otoconia, provides insights into the formation of these inner ear biominerals. Proc. Natl. Acad. Sci. USA 1999, 96, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, D.M.; Zheng, Y.L.; Hu, B.; Zhang, Z.F. Purple sweet potato color alleviates d-galactose-induced brain aging in old mice by promoting survival of neurons via pi3k pathway and inhibiting cytochrome c-mediated apoptosis. Brain Pathol. 2010, 20, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, H.; Yamoah, E.N.; Lundberg, Y.W. Gene targeting reveals the role of oc90 as the essential organizer of the otoconial organic matrix. Dev. Biol. 2007, 304, 508–524. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zhou, D.; Freeman, J.J.; Thalmann, I.; Ornitz, D.M.; Thalmann, R. In vitro effects of recombinant otoconin 90 upon calcite crystal growth. Significance of tertiary structure. Hear. Res. 2010, 268, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jones, S.M.; Yamoah, E.N.; Lundberg, Y.W. Otoconin-90 deletion leads to imbalance but normal hearing: A comparison with other otoconia mutants. Neuroscience 2008, 153, 289–299. [Google Scholar] [CrossRef]
- Lundberg, Y.W.; Xu, Y.; Thiessen, K.D.; Kramer, K.L. Mechanisms of otoconia and otolith development. Dev. Dyn. 2015, 244, 239–253. [Google Scholar] [CrossRef]
- Ignatova, E.G.; Thalmann, I.; Xu, B.; Ornitz, D.M.; Thalmann, R. Molecular mechanisms underlying ectopic otoconia-like particles in the endolymphatic sac of embryonic mice. Hear. Res. 2004, 194, 65–72. [Google Scholar] [CrossRef]
- Deans, M.R.; Peterson, J.M.; Wong, G.W. Mammalian otolin: A multimeric glycoprotein specific to the inner ear that interacts with otoconial matrix protein otoconin-90 and cerebellin-1. PLoS ONE 2010, 5, e12765. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Salmon, M.; El-Amraoui, A.; Leibovici, M.; Petit, C. Otogelin: A glycoprotein specific to the acellular membranes of the inner ear. Proc. Natl. Acad. Sci. USA 1997, 94, 14450–14455. [Google Scholar] [CrossRef]
- El-Amraoui, A.; Cohen-Salmon, M.; Petit, C.; Simmler, M.C. Spatiotemporal expression of otogelin in the developing and adult mouse inner ear. Hear. Res. 2001, 158, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Zwaenepoel, I.; Mustapha, M.; Leibovici, M.; Verpy, E.; Goodyear, R.; Liu, X.Z.; Nouaille, S.; Nance, W.E.; Kanaan, M.; Avraham, K.B.; et al. Otoancorin, an inner ear protein restricted to the interface between the apical surface of sensory epithelia and their overlying acellular gels, is defective in autosomal recessive deafness dfnb22. Proc. Natl. Acad. Sci. USA 2002, 99, 6240–6245. [Google Scholar] [CrossRef]
- Anniko, M.; Wenngren, B.I.; Wroblewski, R. Aberrant elemental composition of otoconia in the dancer mouse mutant with a semidominant gene causing a morphogenetic type of inner ear defect. Acta Oto-Laryngol. 1988, 106, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhao, X.; Xu, Y.; Wang, L.; He, Q.; Lundberg, Y.W. Matrix recruitment and calcium sequestration for spatial specific otoconia development. PLoS ONE 2011, 6, e20498. [Google Scholar] [CrossRef]
- Wenngren, B.I.; Anniko, M. Vestibular hair cell pathology in the dancer mouse mutant. Acta Oto-Laryngol. 1989, 107, 182–190. [Google Scholar] [CrossRef]
- Kozel, P.J.; Friedman, R.A.; Erway, L.C.; Yamoah, E.N.; Liu, L.H.; Riddle, T.; Duffy, J.J.; Doetschman, T.; Miller, M.L.; Cardell, E.L.; et al. Balance and hearing deficits in mice with a null mutation in the gene encoding plasma membrane Ca2+-atpase isoform 2. J. Biol. Chem. 1998, 273, 18693–18696. [Google Scholar] [CrossRef] [PubMed]
- Sondag, H.N.; de Jong, H.A.; van Marle, J.; Oosterveld, W.J. Behavioural changes in hamsters with otoconial malformations. Acta Oto-Laryngol. 1998, 118, 86–89. [Google Scholar]
- Trune, D.R.; Lim, D.J. The behavior and vestibular nuclear morphology of otoconia-deficient pallid mutant mice. J. Neurogenet. 1983, 1, 53–69. [Google Scholar] [CrossRef]
- de Caprona, M.D.; Beisel, K.W.; Nichols, D.H.; Fritzsch, B. Partial behavioral compensation is revealed in balance tasked mutant mice lacking otoconia. Brain Res. Bull. 2004, 64, 289–301. [Google Scholar] [CrossRef]
- Graf, E.; Kelly, R.; Jedin, H. “A history of the council of trent” 1 (book review). Theol. Stud. 1958, 19, 293. [Google Scholar]
- Sweet, H. Head tilt. Mouse News Lett. 1980, 63, 19. [Google Scholar]
- Bergstrom, R.A.; You, Y.; Erway, L.C.; Lyon, M.F.; Schimenti, J.C. Deletion mapping of the head tilt (het) gene in mice: A vestibular mutation causing specific absence of otoliths. Genetics 1998, 150, 815–822. [Google Scholar] [CrossRef]
- Flaherty, J.P.; Fairfield, H.E.; Spruce, C.A.; McCarty, C.M.; Bergstrom, D.E. Molecular characterization of an allelic series of mutations in the mouse Nox3 gene. Mamm. Genome 2011, 22, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Munroe, R.J.; Bergstrom, R.A.; Zheng, Q.Y.; Libby, B.; Smith, R.; John, S.W.; Schimenti, K.J.; Browning, V.L.; Schimenti, J.C. Mouse mutants from chemically mutagenized embryonic stem cells. Nat. Genet. 2000, 24, 318–321. [Google Scholar] [CrossRef]
- Goldowitz, D.; Frankel, W.N.; Takahashi, J.S.; Holtz-Vitaterna, M.; Bult, C.; Kibbe, W.A.; Snoddy, J.; Li, Y.; Pretel, S.; Yates, J.; et al. Large-scale mutagenesis of the mouse to understand the genetic bases of nervous system structure and function. Brain Res. Mol. Brain Res. 2004, 132, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, R.; Gorg, B.; Becker, S.; Qvartskhava, N.; Bidmon, H.J.; Selbach, O.; Haas, H.L.; Schliess, F.; Haussinger, D. Hypoosmotic swelling and ammonia increase oxidative stress by NADPH oxidase in cultured astrocytes and vital brain slices. Glia 2007, 55, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Jones, T.A.; Johnson, K.R.; Yu, H.; Erway, L.C.; Zheng, Q.Y. A comparison of vestibular and auditory phenotypes in inbred mouse strains. Brain Res. 2006, 1091, 40–46. [Google Scholar] [CrossRef]
- Mohri, H.; Ninoyu, Y.; Sakaguchi, H.; Hirano, S.; Saito, N.; Ueyama, T. Nox3-derived superoxide in cochleae induces sensorineural hearing loss. J. Neurosci. 2021, 41, 4716–4731. [Google Scholar] [CrossRef]
- Hudspeth, A.J. Integrating the active process of hair cells with cochlear function. Nat. Rev. Neurosci. 2014, 15, 600–614. [Google Scholar] [CrossRef]
- Kiss, P.J.; Knisz, J.; Zhang, Y.; Baltrusaitis, J.; Sigmund, C.D.; Thalmann, R.; Smith, R.J.; Verpy, E.; Banfi, B. Inactivation of NADPH oxidase organizer 1 results in severe imbalance. Curr. Biol. CB 2006, 16, 208–213. [Google Scholar] [CrossRef]
- Ward, B.K.; Lee, Y.H.; Roberts, D.C.; Naylor, E.; Migliaccio, A.A.; Della Santina, C.C. Mouse magnetic-field nystagmus in strong static magnetic fields is dependent on the presence of Nox3. Otol. Neurotol. 2018, 39, e1150–e1159. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yang, L.; Zhao, X.; Zhang, Y.; Jones, T.A.; Jones, S.M.; Lundberg, Y.W. Functional cooperation between two otoconial proteins oc90 and Nox3. J. Vestib. Res. Equilib. Orientat. 2021, 31, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Darrat, I.; Ahmad, N.; Seidman, K.; Seidman, M.D. Auditory research involving antioxidants. Curr. Opin. Otolaryngol. Head Neck Surg. 2007, 15, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, O.W. Aminoglycoside induced ototoxicity. Toxicology 2008, 249, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.; McFadden, S.L.; Liu, C.C.; Hight, N.; Zheng, X.Y. The role of antioxidants in protection from impulse noise. Ann. N. Y. Acad. Sci. 1999, 884, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear Hear. 2006, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M. A higher oxidative status accelerates senescence and aggravates age-dependent disorders in samp strains of mice. Mech. Ageing Dev. 2002, 123, 1553–1561. [Google Scholar] [CrossRef] [PubMed]
- Kopke, R.; Allen, K.A.; Henderson, D.; Hoffer, M.; Frenz, D.; Van de Water, T. A radical demise. Toxins and trauma share common pathways in hair cell death. Ann. N. Y. Acad. Sci. 1999, 884, 171–191. [Google Scholar] [CrossRef]
- Rybak, L.P.; Ramkumar, V. Ototoxicity. Kidney Int. 2007, 72, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J.H.; Beijnen, J.H.; Balm, A.J.; Schellens, J.H. Future opportunities in preventing cisplatin induced ototoxicity. Cancer Treat. Rev. 2006, 32, 390–397. [Google Scholar] [CrossRef]
- Du, Z.; Li, S.; Liu, L.; Yang, Q.; Zhang, H.; Gao, C. NADPH oxidase 3-associated oxidative stress and caspase 3-dependent apoptosis in the cochleae of d-galactose-induced aged rats. Mol. Med. Rep. 2015, 12, 7883–7890. [Google Scholar] [CrossRef] [PubMed]
- Mukherjea, D.; Jajoo, S.; Kaur, T.; Sheehan, K.E.; Ramkumar, V.; Rybak, L.P. Transtympanic administration of short interfering (si)rna for the nox3 isoform of NADPH oxidase protects against cisplatin-induced hearing loss in the rat. Antioxid. Redox Signal. 2010, 13, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Mukherjea, D.; Whitworth, C.A.; Nandish, S.; Dunaway, G.A.; Rybak, L.P.; Ramkumar, V. Expression of the kidney injury molecule 1 in the rat cochlea and induction by cisplatin. Neuroscience 2006, 139, 733–740. [Google Scholar] [CrossRef]
- Rousset, F.; Nacher-Soler, G.; Kokje, V.B.C.; Sgroi, S.; Coelho, M.; Krause, K.H.; Senn, P. NADPH oxidase 3 deficiency protects from noise-induced sensorineural hearing loss. Front. Cell Dev. Biol. 2022, 10, 832314. [Google Scholar] [CrossRef]
- Vlajkovic, S.M.; Lin, S.C.; Wong, A.C.; Wackrow, B.; Thorne, P.R. Noise-induced changes in expression levels of NADPH oxidases in the cochlea. Hear. Res. 2013, 304, 145–152. [Google Scholar] [CrossRef]
- Besser, S.; Sicker, M.; Marx, G.; Winkler, U.; Eulenburg, V.; Hulsmann, S.; Hirrlinger, J. A transgenic mouse line expressing the red fluorescent protein tdtomato in gabaergic neurons. PLoS ONE 2015, 10, e0129934. [Google Scholar] [CrossRef] [PubMed]
- Yeom, C.J.; Chung, T.; Youn, H.; Kang, K.W.; Lee, D.S.; Chung, J.K. A novel hnis/tdtomato fusion reporter for visualizing the relationship between the cellular localization of sodium iodide symporter and its iodine uptake function under heat shock treatment. Mol. Imaging 2015, 14, 1–10. [Google Scholar]
- Fu, P.; Mohan, V.; Mansoor, S.; Tiruppathi, C.; Sadikot, R.T.; Natarajan, V. Role of nicotinamide adenine dinucleotide phosphate-reduced oxidase proteins in pseudomonas aeruginosa-induced lung inflammation and permeability. Am. J. Respir. Cell Mol. Biol. 2013, 48, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shan, P.; Jiang, G.; Cohn, L.; Lee, P.J. Toll-like receptor 4 deficiency causes pulmonary emphysema. J. Clin. Investig. 2006, 116, 3050–3059. [Google Scholar] [CrossRef]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. Ros are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef]
- Gupta, A.P.; Syed, A.A.; Garg, R.; Goand, U.K.; Singh, P.; Riyazuddin, M.; Valicherla, G.R.; Husain, A.; Gayen, J.R. Pancreastatin inhibitor psti8 attenuates hyperinsulinemia induced obesity and inflammation mediated insulin resistance via mapk/nox3-jnk pathway. Eur. J. Pharmacol. 2019, 864, 172723. [Google Scholar] [CrossRef]
- Kashiwabara, Y.; Ambe, K.; Nakagawa, T.; Watanabe, H. Immunohistochemical localization of nox in mouse circumvallate papillae. Tissue Cell 2015, 47, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Carnesecchi, S.; Carpentier, J.L.; Foti, M.; Szanto, I. Insulin-induced vascular endothelial growth factor expression is mediated by the NADPH oxidase Nox3. Exp. Cell Res. 2006, 312, 3413–3424. [Google Scholar] [CrossRef] [PubMed]
- Maru, Y.; Nishino, T.; Kakinuma, K. Expression of nox genes in rat organs, mouse oocytes, and sea urchin eggs. DNA Seq. 2005, 16, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Michihara, A.; Oda, A.; Mido, M. High expression levels of NADPH oxidase 3 in the cerebrum of ten-week-old stroke-prone spontaneously hypertensive rats. Biol. Pharm. Bull. 2016, 39, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Mizukami, H.; Kurosaki, K.; Kuriiwa, F.; Mukai, T. Existence of a threshold for hydroxyl radical generation independent of hypoxia in rat striatum during carbon monoxide poisoning. Arch. Toxicol. 2011, 85, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Cooney, S.J.; Zhao, Y.; Byrnes, K.R. Characterization of the expression and inflammatory activity of NADPH oxidase after spinal cord injury. Free Radic. Res. 2014, 48, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Zhong, H.; Yanamadala, S.; Campese, V.M. Oxidative stress mediates the stimulation of sympathetic nerve activity in the phenol renal injury model of hypertension. Hypertension 2006, 48, 309–315. [Google Scholar] [CrossRef]
- Habashy, W.S.; Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Expression of genes that encode cellular oxidant/antioxidant systems are affected by heat stress. Mol. Biol. Rep. 2018, 45, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Polettini, J.; Silva, M.G.; Kacerovsky, M.; Syed, T.A.; Saade, G.; Menon, R. Expression profiles of fetal membrane nicotinamide adenine dinucleotide phosphate oxidases (nox) 2 and 3 differentiates spontaneous preterm birth and pprom pathophysiologies. Placenta 2014, 35, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Lyu, P.W.; Xu, X.D.; Zong, K.; Qiu, X.G. Overexpression of duox2 mediates doxorubicin resistance and predicts prognosis of pancreatic cancer. Gland. Surg. 2022, 11, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Li, T.; Wang, C.; Shi, X.; Li, Y.; Zhang, S.; Zhao, Z.; Dong, H.; Qiao, Y. Berberine exerts antioxidant effects via protection of spiral ganglion cells against cytomegalovirus-induced apoptosis. Free Radic. Biol. Med. 2018, 121, 127–135. [Google Scholar] [CrossRef]
- Hamernik, R.P.; Turrentine, G.; Roberto, M.; Salvi, R.; Henderson, D. Anatomical correlates of impulse noise-induced mechanical damage in the cochlea. Hear. Res. 1984, 13, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Wangemann, P. Supporting sensory transduction: Cochlear fluid homeostasis and the endocochlear potential. J. Physiol. 2006, 576, 11–21. [Google Scholar] [CrossRef]
- Kasahara, Y.; Tuder, R.M.; Cool, C.D.; Lynch, D.A.; Flores, S.C.; Voelkel, N.F. Endothelial cell death and decreased expression of vascular endothelial growth factor and vascular endothelial growth factor receptor 2 in emphysema. Am. J. Respir. Crit. Care Med. 2001, 163, 737–744. [Google Scholar] [CrossRef]
- Lu, Q.; Harrington, E.O.; Rounds, S. Apoptosis and lung injury. Keio J. Med. 2005, 54, 184–189. [Google Scholar] [CrossRef]
- Toborek, M.; Hennig, B. Vitamin e attenuates induction of elastase-like activity by tumor necrosis factor-alpha, cholestan-3 beta,5 alpha,6 beta-triol and linoleic acid in cultured endothelial cells. Clin. Chim. Acta Int. J. Clin. Chem. 1993, 215, 201–211. [Google Scholar] [CrossRef]
- Yasuoka, H.; Garrett, S.M.; Nguyen, X.X.; Artlett, C.M.; Feghali-Bostwick, C.A. NADPH oxidase-mediated induction of reactive oxygen species and extracellular matrix deposition by insulin-like growth factor binding protein-5. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L644–L655. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Zhao, W.; Zhou, X.; Miao, G.; Sun, C.; Zhang, H. NADPH oxidase activation contributes to heavy ion irradiation-induced cell death. Dose-Response A Publ. Int. Hormesis Soc. 2017, 15, 1559325817699697. [Google Scholar] [CrossRef]
- Nakayama, N.; Nakamura, T.; Okada, H.; Iwaki, S.; Sobel, B.E.; Fujii, S. Modulators of induction of plasminogen activator inhibitor type-1 in hepg2 cells by transforming growth factor-beta. Coron. Artery Dis. 2011, 22, 468–478. [Google Scholar] [CrossRef]
- Li, H.; Liu, Q.; Wang, N.; Xu, J. Correlation of different NADPH oxidase homologues with late endothelial progenitor cell senescence induced by angiotensin ii: Effect of telmisartan. Intern. Med. 2011, 50, 1631–1642. [Google Scholar] [CrossRef]
- Moon, Y.M.; Kang, H.J.; Cho, J.S.; Park, I.H.; Lee, H.M. Nox4 mediates hypoxia-stimulated myofibroblast differentiation in nasal polyp-derived fibroblasts. Int. Arch. Allergy Immunol. 2012, 159, 399–409. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Wu, L.C.; Dai, T.; Chen, S.Y.; Wang, A.Y.; Lin, K.; Lin, D.M.; Yang, J.Q.; Cheng, B.; Zhang, L.; et al. NADPH oxidase-2 is a key regulator of human dermal fibroblasts: A potential therapeutic strategy for the treatment of skin fibrosis. Exp. Dermatol. 2014, 23, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Van Buul, J.D.; Fernandez-Borja, M.; Anthony, E.C.; Hordijk, P.L. Expression and localization of nox2 and nox4 in primary human endothelial cells. Antioxid. Redox Signal. 2005, 7, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.D.; Shah, M.H.; Liu, G.S.; Chan, E.C.; Crowston, J.G.; Peshavariya, H.M. Transforming growth factor beta1-induced NADPH oxidase-4 expression and fibrotic response in conjunctival fibroblasts. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3011–3017. [Google Scholar] [CrossRef]
- O’Brien, W.J.; Heimann, T.; Rizvi, F. NADPH oxidase expression and production of superoxide by human corneal stromal cells. Mol. Vis. 2009, 15, 2535–2543. [Google Scholar] [PubMed]
- Olguin-Albuerne, M.; Dominguez, G.; Moran, J. Effect of staurosporine in the morphology and viability of cerebellar astrocytes: Role of reactive oxygen species and NADPH oxidase. Oxidative Med. Cell. Longev. 2014, 2014, 678371. [Google Scholar] [CrossRef] [PubMed]
- Accetta, R.; Damiano, S.; Morano, A.; Mondola, P.; Paterno, R.; Avvedimento, E.V.; Santillo, M. Reactive oxygen species derived from nox3 and nox5 drive differentiation of human oligodendrocytes. Front. Cell. Neurosci. 2016, 10, 146. [Google Scholar] [CrossRef] [PubMed]
- Moody, T.W.; Lee, L.; Iordanskaia, T.; Ramos-Alvarez, I.; Moreno, P.; Boudreau, H.E.; Leto, T.L.; Jensen, R.T. Pac1 regulates receptor tyrosine kinase transactivation in a reactive oxygen species-dependent manner. Peptides 2019, 120, 170017. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Han, Y.M.; Jeong, M.; Chung, M.H.; Kwon, C.I.; Ko, K.H.; Hahm, K.B. Synthetic 8-hydroxydeoxyguanosine inhibited metastasis of pancreatic cancer through concerted inhibitions of erm and rho-gtpase. Free Radic. Biol. Med. 2017, 110, 151–161. [Google Scholar] [CrossRef]
- Malleter, M.; Tauzin, S.; Bessede, A.; Castellano, R.; Goubard, A.; Godey, F.; Leveque, J.; Jezequel, P.; Campion, L.; Campone, M.; et al. Cd95l cell surface cleavage triggers a prometastatic signaling pathway in triple-negative breast cancer. Cancer Res. 2013, 73, 6711–6721. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Wu, Z.; Hu, X.; Zhang, J.; Gao, Z.; Han, X.; Qin, J.; Li, C.; Wang, Y. The pi3k/akt/gsk-3beta/ros/eif2b pathway promotes breast cancer growth and metastasis via suppression of nk cell cytotoxicity and tumor cell susceptibility. Cancer Biol. Med. 2019, 16, 38–54. [Google Scholar] [PubMed]
- Ito, K.; Ota, A.; Ono, T.; Nakaoka, T.; Wahiduzzaman, M.; Karnan, S.; Konishi, H.; Furuhashi, A.; Hayashi, T.; Yamada, Y.; et al. Inhibition of nox1 induces apoptosis by attenuating the akt signaling pathway in oral squamous cell carcinoma cell lines. Oncol. Rep. 2016, 36, 2991–2998. [Google Scholar] [CrossRef]
- Kitamoto, K.; Miura, Y.; Karnan, S.; Ota, A.; Konishi, H.; Hosokawa, Y.; Sato, K. Inhibition of NADPH oxidase 2 induces apoptosis in osteosarcoma: The role of reactive oxygen species in cell proliferation. Oncol. Lett. 2018, 15, 7955–7962. [Google Scholar] [CrossRef]
- Tanaka, M.; Miura, Y.; Numanami, H.; Karnan, S.; Ota, A.; Konishi, H.; Hosokawa, Y.; Hanyuda, M. Inhibition of NADPH oxidase 4 induces apoptosis in malignant mesothelioma: Role of reactive oxygen species. Oncol. Rep. 2015, 34, 1726–1732. [Google Scholar] [CrossRef]
- Kim, M.Y.; Bang, E.; Hwangbo, H.; Ji, S.Y.; Kim, D.H.; Lee, H.; Park, C.; Hong, S.H.; Kim, G.Y.; Choi, Y.H. Diallyl trisulfide inhibits monosodium urate-induced nlrp3 inflammasome activation via nox3/4-dependent mitochondrial oxidative stress in raw 264.7 and bone marrow-derived macrophages. Phytomed. Int. J. Phytother. Phytopharm. 2023, 112, 154705. [Google Scholar] [CrossRef] [PubMed]
- Khanal, T.; Kim, H.G.; Do, M.T.; Choi, J.H.; Chung, Y.C.; Kim, H.S.; Park, Y.J.; Jeong, T.C.; Jeong, H.G. Genipin induces cyclooxygenase-2 expression via NADPH oxidase, mapks, ap-1, and nf-kappab in raw 264.7 cells. Food Chem. Toxicol. Int. 2014, 64, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Y.; Tang, M.; Suzuki, M.; Gunasekara, C.; Anbe, Y.; Hiraoka, Y.; Liu, J.; Grasberger, H.; Ohkita, M.; Matsumura, Y.; et al. Essential role of NADPH oxidase-dependent production of reactive oxygen species in maintenance of sustained b cell receptor signaling and b cell proliferation. J. Immunol. 2019, 202, 2546–2557. [Google Scholar] [CrossRef] [PubMed]
- Gaurav, R.; Bewtra, A.K.; Agrawal, D.K. Chloride channel 3 channels in the activation and migration of human blood eosinophils in allergic asthma. Am. J. Respir. Cell Mol. Biol. 2015, 53, 235–245. [Google Scholar] [CrossRef]
- Li, Z.Y.; Jiang, W.Y.; Cui, Z.J. An essential role of NAD(P)H oxidase 2 in uva-induced calcium oscillations in mast cells. Photochem. Photobiol. Sci. 2015, 14, 414–428. [Google Scholar] [CrossRef]
- Cui, X.L.; Brockman, D.; Campos, B.; Myatt, L. Expression of NADPH oxidase isoform 1 (nox1) in human placenta: Involvement in preeclampsia. Placenta 2006, 27, 422–431. [Google Scholar] [CrossRef]
- Matsubara, S.; Sato, I. Enzyme histochemically detectable NAD(P)H oxidase in human placental trophoblasts: Normal, preeclamptic, and fetal growth restriction-complicated pregnancy. Histochem. Cell Biol. 2001, 116, 1–7. [Google Scholar] [CrossRef]
- Manes, C. Human placental NAD(P)H oxidase: Solubilization and properties. Placenta 2001, 22, 58–63. [Google Scholar] [CrossRef]
- Morimoto, H.; Kanatsu-Shinohara, M.; Shinohara, T. Ros-generating oxidase Nox3 regulates the self-renewal of mouse spermatogonial stem cells. Biol. Reprod. 2015, 92, 147. [Google Scholar] [CrossRef] [PubMed]
- Issa, N.; Lachance, G.; Bellmann, K.; Laplante, M.; Stadler, K.; Marette, A. Cytokines promote lipolysis in 3t3-l1 adipocytes through induction of NADPH oxidase 3 expression and superoxide production. J. Lipid Res. 2018, 59, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Brault, J.; Vigne, B.; Meunier, M.; Beaumel, S.; Mollin, M.; Park, S.; Stasia, M.J. Nox4 is the main NADPH oxidase involved in the early stages of hematopoietic differentiation from human induced pluripotent stem cells. Free Radic. Biol. Med. 2020, 146, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, N.M.; Saed, M.G.; Abuanzeh, S.; Abu-Soud, H.M.; Al-Hendy, A.; Diamond, M.P.; Saed, G.M. Nicotinamide adenine dinucleotide phosphate oxidase is differentially regulated in normal myometrium versus leiomyoma. Reprod. Sci. 2014, 21, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Gharib, T.G.; Huang, C.C.; Taylor, J.M.; Misek, D.E.; Kardia, S.L.; Giordano, T.J.; Iannettoni, M.D.; Orringer, M.B.; Hanash, S.M.; et al. Discordant protein and mrna expression in lung adenocarcinomas. Mol. Cell Proteom. 2002, 1, 304–313. [Google Scholar] [CrossRef]
- Pascal, L.E.; True, L.D.; Campbell, D.S.; Deutsch, E.W.; Risk, M.; Coleman, I.M.; Eichner, L.J.; Nelson, P.S.; Liu, A.Y. Correlation of mrna and protein levels: Cell type-specific gene expression of cluster designation antigens in the prostate. BMC Genom. 2008, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Gluschko, A.; Herb, M.; Wiegmann, K.; Krut, O.; Neiss, W.F.; Utermohlen, O.; Kronke, M.; Schramm, M. The β2 integrin mac-1 induces protective lc3-associated phagocytosis of listeria monocytogenes. Cell Host Microbe 2018, 23, 324–337.e325. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, C. What are the bona fide gsk3 substrates? Int. J. Alzheimer’s Dis. 2011, 2011, 505607. [Google Scholar]
- McCubrey, J.A.; Steelman, L.S.; Bertrand, F.E.; Davis, N.M.; Sokolosky, M.; Abrams, S.L.; Montalto, G.; D’Assoro, A.B.; Libra, M.; Nicoletti, F.; et al. Gsk-3 as potential target for therapeutic intervention in cancer. Oncotarget 2014, 5, 2881–2911. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Nong, S.; Huang, X.; Lu, Y.; Zhao, H.; Lin, Y.; Man, Y.; Wang, S.; Yang, J.; Li, J. The effects of palmitate on hepatic insulin resistance are mediated by NADPH oxidase 3-derived reactive oxygen species through jnk and p38mapk pathways. J. Biol. Chem. 2010, 285, 29965–29973. [Google Scholar] [CrossRef]
- Rybak, L.P.; Mukherjea, D.; Ramkumar, V. Mechanisms of cisplatin-induced ototoxicity and prevention. Semin. Hear. 2019, 40, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Mukherjea, D.; Dhukhwa, A.; Rybak, L.P. Oxidative stress and inflammation caused by cisplatin ototoxicity. Antioxidants 2021, 10, 1919. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, J.H.; Kim, S.J.; Oh, G.S.; Moon, H.D.; Kwon, K.B.; Park, C.; Park, B.H.; Lee, H.K.; Chung, S.Y.; et al. Roles of NADPH oxidases in cisplatin-induced reactive oxygen species generation and ototoxicity. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 3933–3946. [Google Scholar] [CrossRef]
- Mukherjea, D.; Jajoo, S.; Whitworth, C.; Bunch, J.R.; Turner, J.G.; Rybak, L.P.; Ramkumar, V. Short interfering rna against transient receptor potential vanilloid 1 attenuates cisplatin-induced hearing loss in the rat. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 13056–13065. [Google Scholar] [CrossRef]
- Shin, Y.S.; Song, S.J.; Kang, S.U.; Hwang, H.S.; Choi, J.W.; Lee, B.H.; Jung, Y.S.; Kim, C.H. A novel synthetic compound, 3-amino-3-(4-fluoro-phenyl)-1h-quinoline-2,4-dione, inhibits cisplatin-induced hearing loss by the suppression of reactive oxygen species: In vitro and in vivo study. Neuroscience 2013, 232, 1–12. [Google Scholar] [CrossRef]
- Hara, S.; Kobayashi, M.; Kuriiwa, F.; Ikematsu, K.; Mizukami, H. Hydroxyl radical production via NADPH oxidase in rat striatum due to carbon monoxide poisoning. Toxicology 2018, 394, 63–71. [Google Scholar] [CrossRef]
- Zahra, G.; Esmaeil, K.; Mohammad, F.; Rashidy-Pour, A.; Mahdi, M.; Mahdi, A.; Ali, K. Combined effects of the exposure to silver nanoparticles and noise on hearing function and cochlea structure of the male rats. Life Sci. 2022, 304, 120724. [Google Scholar] [CrossRef]
- Kim, H.G.; Kim, Y.R.; Park, J.H.; Khanal, T.; Choi, J.H.; Do, M.T.; Jin, S.W.; Han, E.H.; Chung, Y.H.; Jeong, H.G. Endosulfan induces cox-2 expression via NADPH oxidase and the ros, mapk, and akt pathways. Arch. Toxicol. 2015, 89, 2039–2050. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, B.D.; Hussain, Q.Z. Effect of sub-chronic endosulfan exposure on humoral and cell-mediated immune responses in albino rats. Arch. Toxicol. 1986, 59, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Omurtag, G.Z.; Tozan, A.; Sehirli, A.O.; Sener, G. Melatonin protects against endosulfan-induced oxidative tissue damage in rats. J. Pineal Res. 2008, 44, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.Z.; Nisar, M.A.; Alshwmi, M.; Din, S.R.U.; Gamallat, Y.; Khan, M.; Ma, T. Brevilin a inhibits stat3 signaling and induces ros-dependent apoptosis, mitochondrial stress and endoplasmic reticulum stress in mcf-7 breast cancer cells. OncoTargets Ther. 2020, 13, 435–450. [Google Scholar] [CrossRef]
- Zhuang, W.; Wang, C.; Shi, X.; Qiu, S.; Zhang, S.; Xu, B.; Chen, M.; Jiang, W.; Dong, H.; Qiao, Y. Mcmv triggers ros/nlrp3-associated inflammasome activation in the inner ear of mice and cultured spiral ganglion neurons, contributing to sensorineural hearing loss. Int. J. Mol. Med. 2018, 41, 3448–3456. [Google Scholar] [CrossRef] [PubMed]
- Owens, K.M.; Kulawiec, M.; Desouki, M.M.; Vanniarajan, A.; Singh, K.K. Impaired oxphos complex iii in breast cancer. PLoS ONE 2011, 6, e23846. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Chen, F.; Kovalenkov, Y.; Pandey, D.; Moseley, M.A.; Foster, M.W.; Black, S.M.; Venema, R.C.; Stepp, D.W.; Fulton, D.J. Nitric oxide reduces NADPH oxidase 5 (nox5) activity by reversible s-nitrosylation. Free Radic. Biol. Med. 2012, 52, 1806–1819. [Google Scholar] [CrossRef] [PubMed]
- Mahadev, K.; Wu, X.; Zilbering, A.; Zhu, L.; Lawrence, J.T.; Goldstein, B.J. Hydrogen peroxide generated during cellular insulin stimulation is integral to activation of the distal insulin signaling cascade in 3t3-l1 adipocytes. J. Biol. Chem. 2001, 276, 48662–48669. [Google Scholar] [CrossRef]
- Krawczyk, S.A.; Haller, J.F.; Ferrante, T.; Zoeller, R.A.; Corkey, B.E. Reactive oxygen species facilitate translocation of hormone sensitive lipase to the lipid droplet during lipolysis in human differentiated adipocytes. PLoS ONE 2012, 7, e34904. [Google Scholar] [CrossRef]
- Maeda, H.; Rajesh, K.G.; Maeda, H.; Suzuki, R.; Sasaguri, S. Epidermal growth factor and insulin inhibit cell death in pancreatic beta cells by activation of pi3-kinase/akt signaling pathway under oxidative stress. Transplant. Proc. 2004, 36, 1163–1165. [Google Scholar] [CrossRef]
- Stiehl, D.P.; Jelkmann, W.; Wenger, R.H.; Hellwig-Burgel, T. Normoxic induction of the hypoxia-inducible factor 1alpha by insulin and interleukin-1beta involves the phosphatidylinositol 3-kinase pathway. FEBS Lett. 2002, 512, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sen, C.K.; Khanna, S.; Babior, B.M.; Hunt, T.K.; Ellison, E.C.; Roy, S. Oxidant-induced vascular endothelial growth factor expression in human keratinocytes and cutaneous wound healing. J. Biol. Chem. 2002, 277, 33284–33290. [Google Scholar] [CrossRef] [PubMed]
- Berra, E.; Pages, G.; Pouyssegur, J. Map kinases and hypoxia in the control of vegf expression. Cancer Metastasis Rev. 2000, 19, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Li, Y.; Gao, W.; Jiang, Q.; Geng, L.; Ding, J.; Li, S.; Li, J. Transcription factor sp1 transcriptionally enhances gsdme expression for pyroptosis. Cell Death Dis. 2024, 15, 66. [Google Scholar] [CrossRef] [PubMed]
- Pages, G.; Pouyssegur, J. Transcriptional regulation of the vascular endothelial growth factor gene—A concert of activating factors. Cardiovasc. Res. 2005, 65, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Milanini-Mongiat, J.; Pouyssegur, J.; Pages, G. Identification of two sp1 phosphorylation sites for p42/p44 mitogen-activated protein kinases: Their implication in vascular endothelial growth factor gene transcription. J. Biol. Chem. 2002, 277, 20631–20639. [Google Scholar] [CrossRef] [PubMed]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Maddux, B.A.; Goldfine, I.D. The molecular basis for oxidative stress-induced insulin resistance. Antioxid. Redox Signal. 2005, 7, 1040–1052. [Google Scholar] [CrossRef]
- Leclercq, I.A.; Da Silva Morais, A.; Schroyen, B.; Van Hul, N.; Geerts, A. Insulin resistance in hepatocytes and sinusoidal liver cells: Mechanisms and consequences. J. Hepatol. 2007, 47, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Yin, B.; Zhang, H.; Zhang, S.; Zeng, Q.; Wang, J.; Jiang, X.; Yuan, L.; Wang, C.Y.; Li, Z. Blockade of tumor necrosis factor (tnf) receptor type 1-mediated tnf-alpha signaling protected wistar rats from diet-induced obesity and insulin resistance. Endocrinology 2008, 149, 2943–2951. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Kamata, H.; Honda, S.; Maeda, S.; Chang, L.; Hirata, H.; Karin, M. Reactive oxygen species promote tnfalpha-induced death and sustained jnk activation by inhibiting map kinase phosphatases. Cell 2005, 120, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Hirosumi, J.; Tuncman, G.; Chang, L.; Gorgun, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligil, G.S. A central role for jnk in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Nakatani, Y.; Miyatsuka, T.; Kawamori, D.; Matsuoka, T.A.; Matsuhisa, M.; Kajimoto, Y.; Ichijo, H.; Yamasaki, Y.; Hori, M. Possible novel therapy for diabetes with cell-permeable jnk-inhibitory peptide. Nat. Med. 2004, 10, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Fridlyand, L.E.; Philipson, L.H. Reactive species and early manifestation of insulin resistance in type 2 diabetes. Diabetes Obes. Metab. 2006, 8, 136–145. [Google Scholar] [CrossRef]
- Wei, Y.; Sowers, J.R.; Clark, S.E.; Li, W.; Ferrario, C.M.; Stump, C.S. Angiotensin ii-induced skeletal muscle insulin resistance mediated by nf-kappab activation via NADPH oxidase. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E345–E351. [Google Scholar] [CrossRef]
- Nakamura, S.; Takamura, T.; Matsuzawa-Nagata, N.; Takayama, H.; Misu, H.; Noda, H.; Nabemoto, S.; Kurita, S.; Ota, T.; Ando, H.; et al. Palmitate induces insulin resistance in h4iiec3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009, 284, 14809–14818. [Google Scholar] [CrossRef]
- Boden, G. Obesity and free fatty acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646, viii—ix. [Google Scholar] [CrossRef]
- McGarry, J.D. Banting lecture 2001: Dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes 2002, 51, 7–18. [Google Scholar] [CrossRef]
- Azevedo-Martins, A.K.; Monteiro, A.P.; Lima, C.L.; Lenzen, S.; Curi, R. Fatty acid-induced toxicity and neutral lipid accumulation in insulin-producing rinm5f cells. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2006, 20, 1106–1113. [Google Scholar] [CrossRef]
- Mohammed-Ali, Z.; Carlisle, R.E.; Nademi, S.; Dickhout, J.G. Chapter 16—Animal models of kidney disease. In Animal Models for the Study of Human Disease, 2nd ed.; Conn, P.M., Ed.; Academic Press: New York, NY, USA, 2017; pp. 379–417. [Google Scholar]
- Nakatani, Y.; Kaneto, H.; Kawamori, D.; Hatazaki, M.; Miyatsuka, T.; Matsuoka, T.A.; Kajimoto, Y.; Matsuhisa, M.; Yamasaki, Y.; Hori, M. Modulation of the jnk pathway in liver affects insulin resistance status. J. Biol. Chem. 2004, 279, 45803–45809. [Google Scholar] [CrossRef]
- Davis, J.E.; Gabler, N.K.; Walker-Daniels, J.; Spurlock, M.E. The c-jun n-terminal kinase mediates the induction of oxidative stress and insulin resistance by palmitate and toll-like receptor 2 and 4 ligands in 3t3-l1 adipocytes. Horm. Metab. Res. Horm. 2009, 41, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Oxidative stress and stress-activated signaling pathways: A unifying hypothesis of type 2 diabetes. Endocr. Rev. 2002, 23, 599–622. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef]
- Delbosc, S.; Paizanis, E.; Magous, R.; Araiz, C.; Dimo, T.; Cristol, J.P.; Cros, G.; Azay, J. Involvement of oxidative stress and NADPH oxidase activation in the development of cardiovascular complications in a model of insulin resistance, the fructose-fed rat. Atherosclerosis 2005, 179, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, S.; Debure, L.; Moreau, J.F.; Legembre, P. Cd95-mediated cell signaling in cancer: Mutations and post-translational modulations. Cell. Mol. Life Sci. CMLS 2012, 69, 1261–1277. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Holler, N.; Bodmer, J.L.; Hahne, M.; Frei, K.; Fontana, A.; Tschopp, J. Conversion of membrane-bound fas(cd95) ligand to its soluble form is associated with downregulation of its proapoptotic activity and loss of liver toxicity. J. Exp. Med. 1998, 187, 1205–1213. [Google Scholar] [CrossRef]
- Suda, T.; Hashimoto, H.; Tanaka, M.; Ochi, T.; Nagata, S. Membrane fas ligand kills human peripheral blood t lymphocytes, and soluble fas ligand blocks the killing. J. Exp. Med. 1997, 186, 2045–2050. [Google Scholar] [CrossRef]
- Kischkel, F.C.; Hellbardt, S.; Behrmann, I.; Germer, M.; Pawlita, M.; Krammer, P.H.; Peter, M.E. Cytotoxicity-dependent apo-1 (fas/cd95)-associated proteins form a death-inducing signaling complex (disc) with the receptor. EMBO J. 1995, 14, 5579–5588. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, S.; Chaigne-Delalande, B.; Selva, E.; Khadra, N.; Daburon, S.; Contin-Bordes, C.; Blanco, P.; Le Seyec, J.; Ducret, T.; Counillon, L.; et al. The naturally processed cd95l elicits a c-yes/calcium/pi3k-driven cell migration pathway. PLoS Biol. 2011, 9, e1001090. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.H.; Falke, J.J. Ca2+ influx is an essential component of the positive-feedback loop that maintains leading-edge structure and activity in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 16176–16181. [Google Scholar] [CrossRef] [PubMed]
- Cahalan, M.D. Stimulating store-operated Ca2+ entry. Nat. Cell Biol. 2009, 11, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Kleber, S.; Sancho-Martinez, I.; Wiestler, B.; Beisel, A.; Gieffers, C.; Hill, O.; Thiemann, M.; Mueller, W.; Sykora, J.; Kuhn, A.; et al. Yes and pi3k bind cd95 to signal invasion of glioblastoma. Cancer Cell 2008, 13, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Yamamoto, H.; Tominaga, K.; Masuda, K.; Kawai, T.; Teshima-Kondo, S.; Rokutan, K. NADPH oxidase-derived reactive oxygen species are essential for differentiation of a mouse macrophage cell line (raw264.7) into osteoclasts. J. Med. Investig. JMI 2009, 56, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Steinbeck, M.J.; Appel, W.H., Jr.; Verhoeven, A.J.; Karnovsky, M.J. NADPH-oxidase expression and in situ production of superoxide by osteoclasts actively resorbing bone. J. Cell Biol. 1994, 126, 765–772. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, Y.; Ries, W.; Key, L. Expression of nox4 in osteoclasts. J. Cell. Biochem. 2004, 92, 238–248. [Google Scholar] [CrossRef]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in rankl-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef]
- Gavazzi, G.; Banfi, B.; Deffert, C.; Fiette, L.; Schappi, M.; Herrmann, F.; Krause, K.H. Decreased blood pressure in nox1-deficient mice. FEBS Lett. 2006, 580, 497–504. [Google Scholar] [CrossRef]
- Morrison, S.J. Neuronal potential and lineage determination by neural stem cells. Curr. Opin. Cell Biol. 2001, 13, 666–672. [Google Scholar] [CrossRef]
- Kageyama, R.; Ohtsuka, T.; Hatakeyama, J.; Ohsawa, R. Roles of bhlh genes in neural stem cell differentiation. Exp. Cell Res. 2005, 306, 343–348. [Google Scholar] [CrossRef]
- Kennedy, K.A.; Sandiford, S.D.; Skerjanc, I.S.; Li, S.S. Reactive oxygen species and the neuronal fate. Cell. Mol. Life Sci. CMLS 2012, 69, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Katoh, S.; Mitsui, Y.; Kitani, K.; Suzuki, T. Hyperoxia induces the differentiated neuronal phenotype of pc12 cells by producing reactive oxygen species. Biochem. Biophys. Res. Commun. 1997, 241, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, F.; Urra, O.; Alberdi, E.; Matute, C. Oligodendrocyte differentiation from adult multipotent stem cells is modulated by glutamate. Cell Death Dis. 2012, 3, e268. [Google Scholar] [CrossRef] [PubMed]
- Nissen, C.; Ciesielski-Treska, J.; Hertz, L.; Mandel, P. Regulation of oxygen consumption in neuroblastoma cells: Effects of differentiation and of potassium. J. Neurochem. 1973, 20, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Ladi, E.; Mayer-Proschel, M.; Noble, M. Redox state is a central modulator of the balance between self-renewal and differentiation in a dividing glial precursor cell. Proc. Natl. Acad. Sci. USA 2000, 97, 10032–10037. [Google Scholar] [CrossRef] [PubMed]
- Heumuller, S.; Wind, S.; Barbosa-Sicard, E.; Schmidt, H.H.; Busse, R.; Schroder, K.; Brandes, R.P. Apocynin is not an inhibitor of vascular NADPH oxidases but an antioxidant. Hypertension 2008, 51, 211–217. [Google Scholar] [CrossRef]
- Riganti, C.; Costamagna, C.; Bosia, A.; Ghigo, D. The NADPH oxidase inhibitor apocynin (acetovanillone) induces oxidative stress. Toxicol. Appl. Pharmacol. 2006, 212, 179–187. [Google Scholar] [CrossRef]
- Vejrazka, M.; Micek, R.; Stipek, S. Apocynin inhibits NADPH oxidase in phagocytes but stimulates ros production in non-phagocytic cells. Biochim. Biophys. Acta 2005, 1722, 143–147. [Google Scholar] [CrossRef]
- Li, Y.; Trush, M.A. Diphenyleneiodonium, an NAD(P)H oxidase inhibitor, also potently inhibits mitochondrial reactive oxygen species production. Biochem. Biophys. Res. Commun. 1998, 253, 295–299. [Google Scholar] [CrossRef]
- Hutchinson, D.S.; Csikasz, R.I.; Yamamoto, D.L.; Shabalina, I.G.; Wikstrom, P.; Wilcke, M.; Bengtsson, T. Diphenylene iodonium stimulates glucose uptake in skeletal muscle cells through mitochondrial complex i inhibition and activation of amp-activated protein kinase. Cell. Signal. 2007, 19, 1610–1620. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.J.; Anderson, D.J.; McDonald, B.J.; Narayanasami, R.; Bennett, B.M. Inhibition of NADPH-cytochrome p450 reductase and glyceryl trinitrate biotransformation by diphenyleneiodonium sulfate. Biochem. Pharmacol. 1998, 56, 881–893. [Google Scholar] [CrossRef]
- Lu, Q.R.; Sun, T.; Zhu, Z.; Ma, N.; Garcia, M.; Stiles, C.D.; Rowitch, D.H. Common developmental requirement for olig function indicates a motor neuron/oligodendrocyte connection. Cell 2002, 109, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Samanta, J.; Kessler, J.A. Interactions between id and olig proteins mediate the inhibitory effects of bmp4 on oligodendroglial differentiation. Development 2004, 131, 4131–4142. [Google Scholar] [CrossRef]
- Baumann, N.; Pham-Dinh, D. Biology of oligodendrocyte and myelin in the mammalian central nervous system. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef] [PubMed]
- Mazzonetto, P.C.; Ariza, C.B.; Ocanha, S.G.; de Souza, T.A.; Ko, G.M.; Menck, C.F.M.; Massironi, S.M.G.; Porcionatto, M.A. Mutation in NADPH oxidase 3 (nox3) impairs shh signaling and increases cerebellar neural stem/progenitor cell proliferation. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1502–1515. [Google Scholar] [CrossRef]
- Wechsler-Reya, R.J.; Scott, M.P. Control of neuronal precursor proliferation in the cerebellum by sonic hedgehog. Neuron 1999, 22, 103–114. [Google Scholar] [CrossRef]
- Dahmane, N.; Ruiz i Altaba, A. Sonic hedgehog regulates the growth and patterning of the cerebellum. Development 1999, 126, 3089–3100. [Google Scholar] [CrossRef]
- Corrales, J.D.; Rocco, G.L.; Blaess, S.; Guo, Q.; Joyner, A.L. Spatial pattern of sonic hedgehog signaling through gli genes during cerebellum development. Development 2004, 131, 5581–5590. [Google Scholar] [CrossRef]
- Lewis, P.M.; Gritli-Linde, A.; Smeyne, R.; Kottmann, A.; McMahon, A.P. Sonic hedgehog signaling is required for expansion of granule neuron precursors and patterning of the mouse cerebellum. Dev. Biol. 2004, 270, 393–410. [Google Scholar] [CrossRef]
- Vaillant, C.; Monard, D. Shh pathway and cerebellar development. Cerebellum 2009, 8, 291–301. [Google Scholar] [CrossRef]
- Salman, M.S. Epidemiology of cerebellar diseases and therapeutic approaches. Cerebellum 2018, 17, 4–11. [Google Scholar] [CrossRef]
- Massironi, S.M.; Reis, B.L.; Carneiro, J.G.; Barbosa, L.B.; Ariza, C.B.; Santos, G.C.; Guenet, J.L.; Godard, A.L. Inducing mutations in the mouse genome with the chemical mutagen ethylnitrosourea. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas E Biol. 2006, 39, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Nagamani, S.C.; Erez, A.; Eng, C.; Ou, Z.; Chinault, C.; Workman, L.; Coldwell, J.; Stankiewicz, P.; Patel, A.; Lupski, J.R.; et al. Interstitial deletion of 6q25.2-q25.3: A novel microdeletion syndrome associated with microcephaly, developmental delay, dysmorphic features and hearing loss. Eur. J. Hum. Genet. EJHG 2009, 17, 573–581. [Google Scholar] [CrossRef]
- Ahlfeld, J.; Favaro, R.; Pagella, P.; Kretzschmar, H.A.; Nicolis, S.; Schuller, U. Sox2 requirement in sonic hedgehog-associated medulloblastoma. Cancer Res. 2013, 73, 3796–3807. [Google Scholar] [CrossRef] [PubMed]
- Kenney, A.M.; Rowitch, D.H. Sonic hedgehog promotes g(1) cyclin expression and sustained cell cycle progression in mammalian neuronal precursors. Mol. Cell. Biol. 2000, 20, 9055–9067. [Google Scholar] [CrossRef] [PubMed]
- Men, Y.; Zhang, A.; Li, H.; Jin, Y.; Sun, X.; Li, H.; Gao, J. Lkb1 regulates cerebellar development by controlling sonic hedgehog-mediated granule cell precursor proliferation and granule cell migration. Sci. Rep. 2015, 5, 16232. [Google Scholar] [CrossRef] [PubMed]
- Pogoriler, J.; Millen, K.; Utset, M.; Du, W. Loss of cyclin d1 impairs cerebellar development and suppresses medulloblastoma formation. Development 2006, 133, 3929–3937. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.L.; Zhu, S.Y.; Xia, Y.P.; Mao, L.; Mei, Y.W.; Yao, Y.F.; Xue, Y.M.; Hu, B. Sonic hedgehog protects cortical neurons against oxidative stress. Neurochem. Res. 2011, 36, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Kurosaki, T.; Shinohara, H.; Baba, Y. B cell signaling and fate decision. Annu. Rev. Immunol. 2010, 28, 21–55. [Google Scholar] [CrossRef]
- Kurosaki, T.; Hikida, M. Tyrosine kinases and their substrates in b lymphocytes. Immunol. Rev. 2009, 228, 132–148. [Google Scholar] [CrossRef]
- Weber, M.; Treanor, B.; Depoil, D.; Shinohara, H.; Harwood, N.E.; Hikida, M.; Kurosaki, T.; Batista, F.D. Phospholipase c-gamma2 and vav cooperate within signaling microclusters to propagate b cell spreading in response to membrane-bound antigen. J. Exp. Med. 2008, 205, 853–868. [Google Scholar] [CrossRef]
- Choi, M.S.; Brines, R.D.; Holman, M.J.; Klaus, G.G. Induction of nf-at in normal b lymphocytes by anti-immunoglobulin or cd40 ligand in conjunction with il-4. Immunity 1994, 1, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Wienands, J.; Larbolette, O.; Reth, M. Evidence for a preformed transducer complex organized by the b cell antigen receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 7865–7870. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.M.; Clark, E.A. Bcr-induced superoxide negatively regulates b-cell proliferation and t-cell-independent type 2 ab responses. Eur. J. Immunol. 2009, 39, 3395–3403. [Google Scholar] [CrossRef]
- Singh, D.K.; Kumar, D.; Siddiqui, Z.; Basu, S.K.; Kumar, V.; Rao, K.V. The strength of receptor signaling is centrally controlled through a cooperative loop between Ca2+ and an oxidant signal. Cell 2005, 121, 281–293. [Google Scholar] [CrossRef]
- Capasso, M.; Bhamrah, M.K.; Henley, T.; Boyd, R.S.; Langlais, C.; Cain, K.; Dinsdale, D.; Pulford, K.; Khan, M.; Musset, B.; et al. Hvcn1 modulates bcr signal strength via regulation of bcr-dependent generation of reactive oxygen species. Nat. Immunol. 2010, 11, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.L.; Defranco, A.L. Prolonged production of reactive oxygen species in response to b cell receptor stimulation promotes b cell activation and proliferation. J. Immunol. 2012, 189, 4405–4416. [Google Scholar] [CrossRef]
- Musarrat, J.; Arezina-Wilson, J.; Wani, A.A. Prognostic and aetiological relevance of 8-hydroxyguanosine in human breast carcinogenesis. Eur. J. Cancer 1996, 32A, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, P.; Yarlagadda, P.; Vadjunec, N.M.; Proia, A.D.; Harris, R.A.; Portilla, D. Ppar alpha ligand protects during cisplatin-induced acute renal failure by preventing inhibition of renal fao and pdc activity. Am. J. Physiol. Ren. Physiol. 2004, 286, F572–F580. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-ohdg): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health. Part C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed]
- Ock, C.Y.; Hong, K.S.; Choi, K.S.; Chung, M.H.; Kim, Y.; Kim, J.H.; Hahm, K.B. A novel approach for stress-induced gastritis based on paradoxical anti-oxidative and anti-inflammatory action of exogenous 8-hydroxydeoxyguanosine. Biochem. Pharmacol. 2011, 81, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Al-Sabbagh, M.; Fusi, L.; Higham, J.; Lee, Y.; Lei, K.; Hanyaloglu, A.C.; Lam, E.W.; Christian, M.; Brosens, J.J. NADPH oxidase-derived reactive oxygen species mediate decidualization of human endometrial stromal cells in response to cyclic amp signaling. Endocrinology 2011, 152, 730–740. [Google Scholar] [CrossRef]
- Carter, R.J.; Morton, J.; Dunnett, S.B. Motor coordination and balance in rodents. Curr. Protoc. Neurosci. 2001, 15, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Dupureur, C.M.; Zhang, X.; Tsai, M.D. Phospholipase a2 engineering. The roles of disulfide bonds in structure, conformational stability, and catalytic function. Biochemistry 1995, 34, 15307–15314. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A. Diversity of group types, regulation, and function of phospholipase a2. J. Biol. Chem. 1994, 269, 13057–13060. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A. The growing phospholipase a2 superfamily of signal transduction enzymes. Trends Biochem. Sci. 1997, 22, 1–2. [Google Scholar] [CrossRef]
- Jones, T.A.; Jones, S.M.; Hoffman, L.F. Resting discharge patterns of macular primary afferents in otoconia-deficient mice. J. Assoc. Res. Otolaryngol. JARO 2008, 9, 490–505. [Google Scholar] [CrossRef]
- Maklad, A.; Feng, F.; Fritzsch, B. Vestibular primary afferent pathways in mammals. In Encyclopedia of Neuroscience; Binder, M.D., Hirokawa, N., Windhorst, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; p. 4199. [Google Scholar]
- Basaldella, E.; Takeoka, A.; Sigrist, M.; Arber, S. Multisensory signaling shapes vestibulo-motor circuit specificity. Cell 2015, 163, 301–312. [Google Scholar] [CrossRef]
- Hernandez, E.; Das, J.M. Neuroanatomy, nucleus vestibular. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Lim, R.; Brichta, A.M. Chapter 27—Vestibular system. In The Mouse Nervous System; Watson, C., Paxinos, G., Puelles, L., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 661–681. [Google Scholar]
- Houpt, T.A.; Houpt, C.E. Circular swimming in mice after exposure to a high magnetic field. Physiol. Behav. 2010, 100, 284–290. [Google Scholar] [CrossRef]
- Snyder, D.J.; Jahng, J.W.; Smith, J.C.; Houpt, T.A. C-fos induction in visceral and vestibular nuclei of the rat brain stem by a 9.4 t magnetic field. Neuroreport 2000, 11, 2681–2685. [Google Scholar] [CrossRef]
- Cason, A.M.; Kwon, B.; Smith, J.C.; Houpt, T.A. Labyrinthectomy abolishes the behavioral and neural response of rats to a high-strength static magnetic field. Physiol. Behav. 2009, 97, 36–43. [Google Scholar] [CrossRef]
- Lavinsky, J.; Crow, A.L.; Pan, C.; Wang, J.; Aaron, K.A.; Ho, M.K.; Li, Q.; Salehide, P.; Myint, A.; Monges-Hernadez, M.; et al. Genome-wide association study identifies Nox3 as a critical gene for susceptibility to noise-induced hearing loss. PLoS Genet. 2015, 11, e1005094. [Google Scholar]
- Rousset, F.; Carnesecchi, S.; Senn, P.; Krause, K.H. Nox3-targeted therapies for inner ear pathologies. Curr. Pharm. Des. 2015, 21, 5977–5987. [Google Scholar] [CrossRef]
- Rybak, L.P. Cis-platinum associated hearing loss. J. Laryngol. Otol. 1981, 95, 745–747. [Google Scholar] [CrossRef]
- Clerici, W.J.; Hensley, K.; DiMartino, D.L.; Butterfield, D.A. Direct detection of ototoxicant-induced reactive oxygen species generation in cochlear explants. Hear. Res. 1996, 98, 116–124. [Google Scholar] [CrossRef]
- Kopke, R.D.; Liu, W.; Gabaizadeh, R.; Jacono, A.; Feghali, J.; Spray, D.; Garcia, P.; Steinman, H.; Malgrange, B.; Ruben, R.J.; et al. Use of organotypic cultures of corti’s organ to study the protective effects of antioxidant molecules on cisplatin-induced damage of auditory hair cells. Am. J. Otol. 1997, 18, 559–571. [Google Scholar] [PubMed]
- Dhukhwa, A.; Bhatta, P.; Sheth, S.; Korrapati, K.; Tieu, C.; Mamillapalli, C.; Ramkumar, V.; Mukherjea, D. Targeting inflammatory processes mediated by trpvi and tnf-alpha for treating noise-induced hearing loss. Front. Cell. Neurosci. 2019, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Thorne, P.R.; Nuttall, A.L.; Scheibe, F.; Miller, J.M. Sound-induced artifact in cochlear blood flow measurements using the laser doppler flowmeter. Hear. Res. 1987, 31, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Nakai, Y.; Takayama, M.; Konishi, K.; Iguchi, H.; Nakagawa, T.; Shibata, S.; Kato, A.; Sunami, K.; Kawakatsu, C. The emergence of free radicals after acoustic trauma and strial blood flow. Acta Oto-Laryngol. Suppl. 1995, 519, 87–92. [Google Scholar] [CrossRef]
- McAlpine, D.; Johnstone, B.M. The ototoxic mechanism of cisplatin. Hear. Res. 1990, 47, 191–203. [Google Scholar] [CrossRef]
- Rybak, L.P.; Husain, K.; Whitworth, C.; Somani, S.M. Dose dependent protection by lipoic acid against cisplatin-induced ototoxicity in rats: Antioxidant defense system. Toxicol. Sci. Off. J. Soc. Toxicol. 1999, 47, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Understanding mechanisms of antioxidant action in health and disease. Nat. Rev. Mol. Cell Biol. 2024, 25, 13–33. [Google Scholar] [CrossRef]
- Campbell, K.C.; Meech, R.P.; Rybak, L.P.; Hughes, L.F. D-methionine protects against cisplatin damage to the stria vascularis. Hear. Res. 1999, 138, 13–28. [Google Scholar] [CrossRef]
- Korver, K.D.; Rybak, L.P.; Whitworth, C.; Campbell, K.M. Round window application of d-methionine provides complete cisplatin otoprotection. Otolaryngol. Head Neck Surg. 2002, 126, 683–689. [Google Scholar] [CrossRef]
- Choe, W.T.; Chinosornvatana, N.; Chang, K.W. Prevention of cisplatin ototoxicity using transtympanic n-acetylcysteine and lactate. Otol. Neurotol. 2004, 25, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Thomas Dickey, D.; Muldoon, L.L.; Kraemer, D.F.; Neuwelt, E.A. Protection against cisplatin-induced ototoxicity by n-acetylcysteine in a rat model. Hear. Res. 2004, 193, 25–30. [Google Scholar] [CrossRef]
- Feghali, J.G.; Liu, W.; Van De Water, T.R. L-n-acetyl-cysteine protection against cisplatin-induced auditory neuronal and hair cell toxicity. Laryngoscope 2001, 111, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Rybak, L.P.; Kelly, T. Ototoxicity: Bioprotective mechanisms. Curr. Opin. Otolaryngol. Head Neck Surg. 2003, 11, 328–333. [Google Scholar] [CrossRef]
- Yamashita, D.; Jiang, H.Y.; Schacht, J.; Miller, J.M. Delayed production of free radicals following noise exposure. Brain Res. 2004, 1019, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, D.; Jiang, H.Y.; Le Prell, C.G.; Schacht, J.; Miller, J.M. Post-exposure treatment attenuates noise-induced hearing loss. Neuroscience 2005, 134, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Le Prell, C.G.; Yamashita, D.; Minami, S.B.; Yamasoba, T.; Miller, J.M. Mechanisms of noise-induced hearing loss indicate multiple methods of prevention. Hear. Res. 2007, 226, 22–43. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Schacht, J. Emerging treatments for noise-induced hearing loss. Expert Opin. Emerg. Drugs 2011, 16, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.; Barr-Gillespie, P.G. New treatment options for hearing loss. Nat. Rev. Drug Discov. 2015, 14, 346–365. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.; Bonnet, C.; Safieddine, S. Deafness: From genetic architecture to gene therapy. Nat. Rev. Genet. 2023, 24, 665–686. [Google Scholar] [CrossRef] [PubMed]
- Kamogashira, T.; Fujimoto, C.; Yamasoba, T. Reactive oxygen species, apoptosis, and mitochondrial dysfunction in hearing loss. BioMed Res. Int. 2015, 2015, 617207. [Google Scholar] [CrossRef]
- Pittman, C.A.; Ward, B.K.; Nieman, C.L. A review of adult-onset hearing loss: A primer for neurologists. Curr. Treat Options Neurol. 2021, 23, 20. [Google Scholar] [CrossRef]
- Le, T.N.; Straatman, L.V.; Lea, J.; Westerberg, B. Current insights in noise-induced hearing loss: A literature review of the underlying mechanism, pathophysiology, asymmetry, and management options. J. Otolaryngol. Head Neck Surg. Le J. D’oto-Rhino-Laryngol. Chir. Cervico-Faciale 2017, 46, 41. [Google Scholar] [CrossRef]
- Wang, J.; Puel, J.L. Presbycusis: An update on cochlear mechanisms and therapies. J. Clin. Med. 2020, 9, 218. [Google Scholar] [CrossRef]
- Liberman, M.C.; Kujawa, S.G. Cochlear synaptopathy in acquired sensorineural hearing loss: Manifestations and mechanisms. Hear. Res. 2017, 349, 138–147. [Google Scholar] [CrossRef]
- Wu, P.Z.; Liberman, L.D.; Bennett, K.; de Gruttola, V.; O’Malley, J.T.; Liberman, M.C. Primary neural degeneration in the human cochlea: Evidence for hidden hearing loss in the aging ear. Neuroscience 2019, 407, 8–20. [Google Scholar] [CrossRef]
- Peineau, T.; Belleudy, S.; Pietropaolo, S.; Bouleau, Y.; Dulon, D. Synaptic release potentiation at aging auditory ribbon synapses. Front. Aging Neurosci. 2021, 13, 756449. [Google Scholar] [CrossRef]
- Schwander, M.; Kachar, B.; Muller, U. Review series: The cell biology of hearing. J. Cell Biol. 2010, 190, 9–20. [Google Scholar] [CrossRef]
- Atkinson, P.J.; Huarcaya Najarro, E.; Sayyid, Z.N.; Cheng, A.G. Sensory hair cell development and regeneration: Similarities and differences. Development 2015, 142, 1561–1571. [Google Scholar] [CrossRef]
- Furness, D.N. Molecular basis of hair cell loss. Cell Tissue Res. 2015, 361, 387–399. [Google Scholar] [CrossRef]
- Kujawa, S.G.; Liberman, M.C. Translating animal models to human therapeutics in noise-induced and age-related hearing loss. Hear. Res. 2019, 377, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Clerici, W.J.; Yang, L. Direct effects of intraperilymphatic reactive oxygen species generation on cochlear function. Hear. Res. 1996, 101, 14–22. [Google Scholar] [CrossRef]
- Bielefeld, E.C.; Hu, B.H.; Harris, K.C.; Henderson, D. Damage and threshold shift resulting from cochlear exposure to paraquat-generated superoxide. Hear. Res. 2005, 207, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Ryan, A.F. Mechanisms of sensorineural cell damage, death and survival in the cochlea. Front. Aging Neurosci. 2015, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Paludetti, G.; Troiani, D. Targeting dysregulation of redox homeostasis in noise-induced hearing loss: Oxidative stress and ros signaling. Free Radic. Biol. Med. 2019, 135, 46–59. [Google Scholar] [CrossRef]
- Fujioka, M.; Kanzaki, S.; Okano, H.J.; Masuda, M.; Ogawa, K.; Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J. Neurosci. Res. 2006, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Kaygusuz, I.; Ozturk, A.; Ustundag, B.; Yalcin, S. Role of free oxygen radicals in noise-related hearing impairment. Hear. Res. 2001, 162, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Nakai, Y.; Takayama, M.; Iguchi, H.; Nakagawa, T.; Kojima, A. Appearance of free radicals in the guinea pig inner ear after noise-induced acoustic trauma. Eur. Arch. Oto-Rhino-Laryngol. 1995, 252, 504–508. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; Wright, J.S.; Dugan, L.L. Early elevation of cochlear reactive oxygen species following noise exposure. Audiol. Neuro-Otol. 1999, 4, 229–236. [Google Scholar] [CrossRef]
- Priuska, E.M.; Schacht, J. Formation of free radicals by gentamicin and iron and evidence for an iron/gentamicin complex. Biochem. Pharmacol. 1995, 50, 1749–1752. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K.; Hockenbery, D.M.; Rubel, E.W. Reactive oxygen species in chick hair cells after gentamicin exposure in vitro. Hear. Res. 1997, 104, 1–14. [Google Scholar] [CrossRef]
- Dehne, N.; Lautermann, J.; Petrat, F.; Rauen, U.; de Groot, H. Cisplatin ototoxicity: Involvement of iron and enhanced formation of superoxide anion radicals. Toxicol. Appl. Pharmacol. 2001, 174, 27–34. [Google Scholar] [CrossRef]
- Paparella, M.M.; Oda, M.; Hiraide, F.; Brady, D. Pathology of sensorineural hearing loss in otitis media. Ann. Otol. Rhinol. Laryngol. 1972, 81, 632–647. [Google Scholar] [CrossRef]
- Merchant, S.N.; Gopen, Q. A human temporal bone study of acute bacterial meningogenic labyrinthitis. Am. J. Otol. 1996, 17, 375–385. [Google Scholar]
- Fujimoto, C.; Yamasoba, T. Oxidative stresses and mitochondrial dysfunction in age-related hearing loss. Oxidative Med. Cell. Longev. 2014, 2014, 582849. [Google Scholar] [CrossRef]
- Warraich, U.E.; Hussain, F.; Kayani, H.U.R. Aging—Oxidative stress, antioxidants and computational modeling. Heliyon 2020, 6, e04107. [Google Scholar] [CrossRef] [PubMed]
- Kozakiewicz, M.; Kornatowski, M.; Krzywinska, O.; Kedziora-Kornatowska, K. Changes in the blood antioxidant defense of advanced age people. Clin. Interv. Aging 2019, 14, 763–771. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; McFadden, S.L.; Ding, D.L.; Lear, P.M.; Ho, Y.S. Targeted mutation of the gene for cellular glutathione peroxidase (gpx1) increases noise-induced hearing loss in mice. J. Assoc. Res. Otolaryngol. JARO 2000, 1, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.H.; Henderson, D.; Nicotera, T.M. Involvement of apoptosis in progression of cochlear lesion following exposure to intense noise. Hear. Res. 2002, 166, 62–71. [Google Scholar] [CrossRef]
- Bohne, B. Mechanisms of Noise Damage in the Inner Ear; John H. Mills Raven Press: New York, NY, USA, 1976; pp. 41–68. [Google Scholar]
- Bohne, B.A.; Harding, G.W.; Lee, S.C. Death pathways in noise-damaged outer hair cells. Hear. Res. 2007, 223, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Murai, N.; Kirkegaard, M.; Jarlebark, L.; Risling, M.; Suneson, A.; Ulfendahl, M. Activation of jnk in the inner ear following impulse noise exposure. J. Neurotrauma 2008, 25, 72–77. [Google Scholar] [CrossRef]
- Seidman, M.D.; Shivapuja, B.G.; Quirk, W.S. The protective effects of allopurinol and superoxide dismutase on noise-induced cochlear damage. Otolaryngol. Head Neck Surg. 1993, 109, 1052–1056. [Google Scholar] [CrossRef]
- Liu, Z. Experimental study on the mechanism of free radical in blast trauma induced hearing loss. Zhonghua Er Bi Yan Hou Ke Za Zhi 1992, 27, 24–26. [Google Scholar]
- Hamanaka, R.B.; Chandel, N.S. Mitochondrial reactive oxygen species regulate cellular signaling and dictate biological outcomes. Trends Biochem. Sci. 2010, 35, 505–513. [Google Scholar] [CrossRef]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. Tlr signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Myers, A.L.; Harris, C.M.; Choe, K.M.; Brennan, C.A. Inflammatory production of reactive oxygen species by drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun. 2018, 505, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.A.; Romagna, C.D.; Pereira, J.L.; Souza-Pinto, N.C. The role of mitochondrial DNA damage in the citotoxicity of reactive oxygen species. J. Bioenerg. Biomembr. 2011, 43, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta 1998, 1366, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, S.; Wang, L.; Li, H.; Wang, Y.; Liu, H.; Wang, X.; Zhu, X.; Liu, Z.; Ye, F.; et al. Mitochondrial dysfunction in hearing loss: Oxidative stress, autophagy and nlrp3 inflammasome. Front. Cell Dev. Biol. 2023, 11, 1119773. [Google Scholar] [CrossRef] [PubMed]
- Pak, J.H.; Kim, Y.; Yi, J.; Chung, J.W. Antioxidant therapy against oxidative damage of the inner ear: Protection and preconditioning. Antioxidants 2020, 9, 1076. [Google Scholar] [CrossRef]
- Kishimoto-Urata, M.; Urata, S.; Fujimoto, C.; Yamasoba, T. Role of oxidative stress and antioxidants in acquired inner ear disorders. Antioxidants 2022, 11, 1469. [Google Scholar] [CrossRef]
- Fay, R.R. Comparative psychoacoustics. Hear. Res. 1988, 34, 295–305. [Google Scholar] [CrossRef]
- Nacher-Soler, G.; Lenglet, S.; Coelho, M.; Thomas, A.; Voruz, F.; Krause, K.H.; Senn, P.; Rousset, F. Local cisplatin delivery in mouse reliably models sensorineural ototoxicity without systemic adverse effects. Front. Cell. Neurosci. 2021, 15, 701783. [Google Scholar] [CrossRef]
- Milosevic, J.; Kalicanin, P. Long arm deletion of chromosome no. 6 in a mentally retarded boy with multiple physical malformations. J. Ment. Defic. Res. 1975, 19, 139–144. [Google Scholar] [PubMed]
- Hopkin, R.J.; Schorry, E.; Bofinger, M.; Milatovich, A.; Stern, H.J.; Jayne, C.; Saal, H.M. New insights into the phenotypes of 6q deletions. Am. J. Med. Genet. 1997, 70, 377–386. [Google Scholar] [CrossRef]
- Sukumar, S.; Wang, S.; Hoang, K.; Vanchiere, C.M.; England, K.; Fick, R.; Pagon, B.; Reddy, K.S. Subtle overlapping deletions in the terminal region of chromosome 6q24.2–q26: Three cases studied using fish. Am. J. Med. Genet. 1999, 87, 17–22. [Google Scholar] [CrossRef]
- Pandya, A.; Braverman, N.; Pyeritz, R.E.; Ying, K.L.; Kline, A.D.; Falk, R.E. Interstitial deletion of the long arm of chromosome 6 associated with unusual limb anomalies: Report of two new patients and review of the literature. Am. J. Med. Genet. 1995, 59, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Lohscheller, J.; Kummer, P.; Eysholdt, U.; Rosanowski, F. Severe sensory hearing loss in del(6q)-syndrome. Int. J. Pediatr. Otorhinolaryngol. 2003, 67, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Stucken, E.Z.; Hong, R.S. Noise-induced hearing loss: An occupational medicine perspective. Curr. Opin. Otolaryngol. Head Neck Surg. 2014, 22, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Dobie, R.A. The burdens of age-related and occupational noise-induced hearing loss in the united states. Ear Hear. 2008, 29, 565–577. [Google Scholar] [CrossRef]
- Yankaskas, K. Prelude: Noise-induced tinnitus and hearing loss in the military. Hear. Res. 2013, 295, 3–8. [Google Scholar] [CrossRef]
- Guski, R.; Schreckenberg, D.; Schuemer, R. Who environmental noise guidelines for the european region: A systematic review on environmental noise and annoyance. Int. J. Environ. Res. Public Health 2017, 14, 1539. [Google Scholar] [CrossRef]
- Saffree Jeffree, M.; Ismail, N.; Awang Lukman, K. Hearing impairment and contributing factors among fertilizer factory workers. J. Occup. Health 2016, 58, 434–443. [Google Scholar] [CrossRef]
- Habybabady, R.H.; Mohammadi, M.; Mortazavi, S.B.; Khavanin, A.; Mirzaei, R.; Malvajerdi, M.S. The effect of simultaneous exposure to cigarette smoke and noise on distortion product otoacoustic emissions in rats. Toxicol. Ind. Health 2019, 35, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef]
- Kaur, T.; Clayman, A.C.; Nash, A.J.; Schrader, A.D.; Warchol, M.E.; Ohlemiller, K.K. Lack of fractalkine receptor on macrophages impairs spontaneous recovery of ribbon synapses after moderate noise trauma in c57bl/6 mice. Front. Neurosci. 2019, 13, 620. [Google Scholar] [CrossRef] [PubMed]
- Pourbakht, A.; Yamasoba, T. Cochlear damage caused by continuous and intermittent noise exposure. Hear. Res. 2003, 178, 70–78. [Google Scholar] [CrossRef]
- Op de Beeck, K.; Schacht, J.; Van Camp, G. Apoptosis in acquired and genetic hearing impairment: The programmed death of the hair cell. Hear. Res. 2011, 281, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Shargorodsky, J.; Curhan, S.G.; Curhan, G.C.; Eavey, R. Change in prevalence of hearing loss in us adolescents. JAMA 2010, 304, 772–778. [Google Scholar] [CrossRef]
- Wang, X.; Xu, P.; Li, P.; Wang, Z.; Zhao, F.; Gao, Z.; Xu, L.; Luo, Y.J.; Fan, J.; Liu, P. Alterations in gray matter volume due to unilateral hearing loss. Sci. Rep. 2016, 6, 25811. [Google Scholar] [CrossRef]
- Nakajima, K.; Kanda, E.; Hosobuchi, A.; Suwa, K. Subclinical hearing loss, longer sleep duration, and cardiometabolic risk factors in japanese general population. Int. J. Otolaryngol. 2014, 2014, 218218. [Google Scholar] [CrossRef] [PubMed]
- Lie, A.; Skogstad, M.; Johannessen, H.A.; Tynes, T.; Mehlum, I.S.; Nordby, K.C.; Engdahl, B.; Tambs, K. Occupational noise exposure and hearing: A systematic review. Int. Arch. Occup. Environ. Health 2016, 89, 351–372. [Google Scholar] [CrossRef]
- Sliwinska-Kowalska, M.; Pawelczyk, M. Contribution of genetic factors to noise-induced hearing loss: A human studies review. Mutat. Res. 2013, 752, 61–65. [Google Scholar] [CrossRef]
- Heinonen-Guzejev, M.; Vuorinen, H.S.; Mussalo-Rauhamaa, H.; Heikkilä, K.; Koskenvuo, M.; Kaprio, J. Genetic component of noise sensitivity. Twin Res. Hum. Genet. 2012, 8, 245–249. [Google Scholar] [CrossRef]
- Erway, L.C.; Shiau, Y.W.; Davis, R.R.; Krieg, E.F. Genetics of age-related hearing loss in mice. Iii. Susceptibility of inbred and f1 hybrid strains to noise-induced hearing loss. Hear. Res. 1996, 93, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Kozel, P.J.; Davis, R.R.; Krieg, E.F.; Shull, G.E.; Erway, L.C. Deficiency in plasma membrane calcium atpase isoform 2 increases susceptibility to noise-induced hearing loss in mice. Hear. Res. 2002, 164, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Holme, R.H.; Steel, K.P. Progressive hearing loss and increased susceptibility to noise-induced hearing loss in mice carrying a cdh23 but not a myo7a mutation. J. Assoc. Res. Otolaryngol. JARO 2004, 5, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.; Whitworth, C.A.; Pingle, S.C.; Hughes, L.F.; Rybak, L.P. Noise induces a1 adenosine receptor expression in the chinchilla cochlea. Hear. Res. 2004, 188, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Vethanayagam, R.R.; Dong, Y.; Cai, Q.; Hu, B.H. Activation of the antigen presentation function of mononuclear phagocyte populations associated with the basilar membrane of the cochlea after acoustic overstimulation. Neuroscience 2015, 303, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S.; Billings, P.; Harris, J.P.; Firestein, G.S.; Keithley, E.M. Innate immunity contributes to cochlear adaptive immune responses. Audiol. Neuro-Otol. 2005, 10, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, K.; Fujioka, M.; Kanzaki, S.; Okano, H.J.; Shibata, S.; Yamashita, D.; Masuda, M.; Mihara, M.; Ohsugi, Y.; Ogawa, K.; et al. Blockade of interleukin-6 signaling suppressed cochlear inflammatory response and improved hearing impairment in noise-damaged mice cochlea. Neurosci. Res. 2010, 66, 345–352. [Google Scholar] [CrossRef]
- Keithley, E.M.; Wang, X.; Barkdull, G.C. Tumor necrosis factor alpha can induce recruitment of inflammatory cells to the cochlea. Otol. Neurotol. 2008, 29, 854–859. [Google Scholar] [CrossRef]
- Paciello, F.; Di Pino, A.; Rolesi, R.; Troiani, D.; Paludetti, G.; Grassi, C.; Fetoni, A.R. Anti-oxidant and anti-inflammatory effects of caffeic acid: In Vivo evidences in a model of noise-induced hearing loss. Food Chem. Toxicol. 2020, 143, 111555. [Google Scholar] [CrossRef]
- Kurabi, A.; Keithley, E.M.; Housley, G.D.; Ryan, A.F.; Wong, A.C. Cellular mechanisms of noise-induced hearing loss. Hear. Res. 2017, 349, 129–137. [Google Scholar] [CrossRef]
- Bennett, B.J.; Farber, C.R.; Orozco, L.; Kang, H.M.; Ghazalpour, A.; Siemers, N.; Neubauer, M.; Neuhaus, I.; Yordanova, R.; Guan, B.; et al. A high-resolution association mapping panel for the dissection of complex traits in mice. Genome Res. 2010, 20, 281–290. [Google Scholar] [CrossRef]
- Farber, C.R.; Bennett, B.J.; Orozco, L.; Zou, W.; Lira, A.; Kostem, E.; Kang, H.M.; Furlotte, N.; Berberyan, A.; Ghazalpour, A.; et al. Mouse genome-wide association and systems genetics identify asxl2 as a regulator of bone mineral density and osteoclastogenesis. PLoS Genet. 2011, 7, e1002038. [Google Scholar] [CrossRef]
- Park, C.C.; Gale, G.D.; de Jong, S.; Ghazalpour, A.; Bennett, B.J.; Farber, C.R.; Langfelder, P.; Lin, A.; Khan, A.H.; Eskin, E.; et al. Gene networks associated with conditional fear in mice identified using a systems genetics approach. BMC Syst. Biol. 2011, 5, 43. [Google Scholar] [CrossRef]
- Zhao, T.; Wang, Y.; Li, Z.; Xu, X.; Lei, S.; Huang, L.; Xu, L.; Zhang, M.; Yang, L. Associations of noise kurtosis, genetic variations in nox3 and lifestyle factors with noise-induced hearing loss. Environ. Health A Glob. Access Sci. Source 2020, 19, 13. [Google Scholar] [CrossRef]
- Lei, S.F.; Ahroon, W.A.; Hamernik, R.P. The application of frequency and time domain kurtosis to the assessment of hazardous noise exposures. J. Acoust. Soc. Am. 1994, 96, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.M.; Qiu, W.; Zeng, L.; Chen, S.S.; Cheng, X.R.; Davis, R.I.; Hamernik, R.P. Application of the kurtosis statistic to the evaluation of the risk of hearing loss in workers exposed to high-level complex noise. Ear Hear. 2010, 31, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kitamura, T.; Sakai, H.; Ando, Y. The loudness of “complex noise” in relation to the factors extracted from the auto-correlation function. J. Sound Vib. 2001, 241, 97–103. [Google Scholar] [CrossRef]
- Erdreich, J. A distribution based definition of impulse noise. J. Acoust. Soc. Am. 1986, 79, 990–998. [Google Scholar] [CrossRef]
- Xie, H.W.; Qiu, W.; Heyer, N.J.; Zhang, M.B.; Zhang, P.; Zhao, Y.M.; Hamernik, R.P. The use of the kurtosis-adjusted cumulative noise exposure metric in evaluating the hearing loss risk for complex noise. Ear Hear. 2016, 37, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.I.; Qiu, W.; Heyer, N.J.; Zhao, Y.; Qiuling Yang, M.S.; Li, N.; Tao, L.; Zhu, L.; Zeng, L.; Yao, D. The use of the kurtosis metric in the evaluation of occupational hearing loss in workers in china: Implications for hearing risk assessment. Noise Health 2012, 14, 330–342. [Google Scholar] [CrossRef]
- Kaur, T.; Borse, V.; Sheth, S.; Sheehan, K.; Ghosh, S.; Tupal, S.; Jajoo, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Adenosine a1 receptor protects against cisplatin ototoxicity by suppressing the nox3/stat1 inflammatory pathway in the cochlea. J. Neurosci. 2016, 36, 3962–3977. [Google Scholar] [CrossRef]
- Peralta-Videa, J.R.; Zhao, L.; Lopez-Moreno, M.L.; de la Rosa, G.; Hong, J.; Gardea-Torresdey, J.L. Nanomaterials and the environment: A review for the biennium 2008–2010. J. Hazard. Mater. 2011, 186, 1–15. [Google Scholar] [CrossRef]
- Roda, E.; Bottone, M.G.; Biggiogera, M.; Milanesi, G.; Coccini, T. Pulmonary and hepatic effects after low dose exposure to nanosilver: Early and long-lasting histological and ultrastructural alterations in rat. Toxicol. Rep. 2019, 6, 1047–1060. [Google Scholar] [CrossRef]
- Recordati, C.; De Maglie, M.; Bianchessi, S.; Argentiere, S.; Cella, C.; Mattiello, S.; Cubadda, F.; Aureli, F.; D’Amato, M.; Raggi, A. Tissue distribution and acute toxicity of silver after single intravenous administration in mice: Nano-specific and size-dependent effects. Part. Fibre Toxicol. 2015, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, X.; Takemura, T.; Hanagata, N.; Wu, G.; Chou, L.L. Genotoxicity and molecular response of silver nanoparticle (np)-based hydrogel. J. Nanobiotechnol. 2012, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, Z.; Nemmar, A. Health impact of silver nanoparticles: A review of the biodistribution and toxicity following various routes of exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef] [PubMed]
- AshaRani, P.V.; Low Kah Mun, G.; Hande, M.P.; Valiyaveettil, S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 2009, 3, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Kawata, K.; Osawa, M.; Okabe, S. In vitro toxicity of silver nanoparticles at noncytotoxic doses to hepg2 human hepatoma cells. Environ. Sci. Technol. 2009, 43, 6046–6051. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, M.J.; Lee, S.Y.; Oh, S.M.; Chung, K.H. Genotoxic effects of silver nanoparticles stimulated by oxidative stress in human normal bronchial epithelial (beas-2b) cells. Mutat. Res. 2011, 726, 129–135. [Google Scholar] [CrossRef]
- Neely, S.T.; Johnson, T.A.; Kopun, J.; Dierking, D.M.; Gorga, M.P. Distortion-product otoacoustic emission input/output characteristics in normal-hearing and hearing-impaired human ears. J. Acoust. Soc. Am. 2009, 126, 728–738. [Google Scholar] [CrossRef]
- Martin, G.K.; Stagner, B.B.; Chung, Y.S.; Lonsbury-Martin, B.L. Characterizing distortion-product otoacoustic emission components across four species. J. Acoust. Soc. Am. 2011, 129, 3090–3103. [Google Scholar] [CrossRef]
- Shahtaheri, S.J.; Goodarzi, Z.; Karami, E.; Khavanin, A.; Khansari, M.G.; Kiani, M.; Rashidy-Pour, A. Effects of acute exposure to al(2)o(3)-nps (alpha and gamma) and white noise and their combination on cochlea structure and function in wistar rats. Environ. Sci. Pollut. Res. Int. 2023, 30, 89859–89876. [Google Scholar] [CrossRef]
- Ates, M.; Demir, V.; Arslan, Z.; Daniels, J.; Farah, I.O.; Bogatu, C. Evaluation of alpha and gamma aluminum oxide nanoparticle accumulation, toxicity, and depuration in artemia salina larvae. Environ. Toxicol. 2015, 30, 109–118. [Google Scholar] [CrossRef]
- Patlolla, A.K.; Kumari, S.A.; Madhusudhanachary, P.; Turner, T.; Tchounwou, P.B. Biochemical and histopathological evaluation of al(2)o(3) nanomaterials in kidney of wistar rats. Curr. Top. Biochem. Res. 2018, 19, 1–12. [Google Scholar] [PubMed]
- Jacukowicz-Sobala, I.; Ocinski, D.; Kociolek-Balawejder, E. Iron and aluminium oxides containing industrial wastes as adsorbents of heavy metals: Application possibilities and limitations. Waste Manag. Res. 2015, 33, 612–629. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.M.; Brown, A.M.; Willitsford, A.H.; Dinello-Fass, R.; Airola, M.B.; Siegrist, K.M.; Thomas, M.E.; Chang, Y. Lidar measurements of solid rocket propellant fire particle plumes. Appl. Opt. 2016, 55, 4657–4669. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.; Milton, R.; Perry, K.M.A.; Thompson, D.R. Effect of aluminium and alumina on the lung in grinders of duralumin aeroplane propellers. Br. J. Ind. Med. 1944, 1, 159–164. [Google Scholar] [CrossRef]
- Sikkeland, L.; Alexis, N.E.; Fry, R.C.; Martin, E.; Danielsen, T.E.; Sostrand, P.; Kongerud, J. Inflammation in induced sputum after aluminium oxide exposure: An experimental chamber study. Occup. Environ. Med. 2016, 73, 199–205. [Google Scholar] [CrossRef]
- Xing, M.; Zou, H.; Gao, X.; Chang, B.; Tang, S.; Zhang, M. Workplace exposure to airborne alumina nanoparticles associated with separation and packaging processes in a pilot factory. Environ. Sci. Process Impacts 2015, 17, 656–666. [Google Scholar] [CrossRef]
- Lu, J.; Li, W.; Du, X.; Ewert, D.L.; West, M.B.; Stewart, C.; Floyd, R.A.; Kopke, R.D. Antioxidants reduce cellular and functional changes induced by intense noise in the inner ear and cochlear nucleus. J. Assoc. Res. Otolaryngol. JARO 2014, 15, 353–372. [Google Scholar] [CrossRef] [PubMed]
- Fram, R.J. Cisplatin and platinum analogues: Recent advances. Curr. Opin. Oncol. 1992, 4, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Boulikas, T.; Vougiouka, M. Recent clinical trials using cisplatin, carboplatin and their combination chemotherapy drugs (review). Oncol. Rep. 2004, 11, 559–595. [Google Scholar] [CrossRef] [PubMed]
- Laurell, G. Pharmacological intervention in the field of ototoxicity. Hno 2019, 67, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lippard, S.J. Cellular processing of platinum anticancer drugs. Nat. Rev. Drug Discov. 2005, 4, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.A.; Schwade, N.D.; Truelson, J.M. Fosfomycin does not inhibit the tumoricidal efficacy of cisplatinum. Laryngoscope 1999, 109, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; El-Demerdash, E. Protective effects of l-arginine against cisplatin-induced renal oxidative stress and toxicity: Role of nitric oxide. Basic Clin. Pharmacol. Toxicol. 2005, 97, 91–97. [Google Scholar] [CrossRef]
- Bodenner, D.L.; Dedon, P.C.; Keng, P.C.; Borch, R.F. Effect of diethyldithiocarbamate on cis-diamminedichloroplatinum(ii)-induced cytotoxicity, DNA cross-linking, and gamma-glutamyl transpeptidase inhibition. Cancer Res. 1986, 46, 2745–2750. [Google Scholar]
- Barabas, K.; Milner, R.; Lurie, D.; Adin, C. Cisplatin: A review of toxicities and therapeutic applications. Vet. Comp. Oncol. 2008, 6, 1–18. [Google Scholar] [CrossRef]
- Hartmann, J.T.; Lipp, H.P. Toxicity of platinum compounds. Expert Opin. Pharmacother. 2003, 4, 889–901. [Google Scholar] [CrossRef]
- Schacht, J.; Talaska, A.E.; Rybak, L.P. Cisplatin and aminoglycoside antibiotics: Hearing loss and its prevention. Anat. Rec. 2012, 295, 1837–1850. [Google Scholar] [CrossRef]
- Koike, N.; Sasaki, A.; Murakami, T.; Suzuki, K. Effect of edaravone against cisplatin-induced chronic renal injury. Drug Chem. Toxicol. 2021, 44, 437–446. [Google Scholar] [CrossRef]
- Thadhani, R.; Pascual, M.; Bonventre, J.V. Acute renal failure. N. Engl. J. Med. 1996, 334, 1448–1460. [Google Scholar] [CrossRef]
- Rybak, L.P.; Mukherjea, D.; Jajoo, S.; Ramkumar, V. Cisplatin ototoxicity and protection: Clinical and experimental studies. Tohoku J. Exp. Med. 2009, 219, 177–186. [Google Scholar] [CrossRef] [PubMed]
- McKeage, M.J. Comparative adverse effect profiles of platinum drugs. Drug Saf. 1995, 13, 228–244. [Google Scholar] [CrossRef] [PubMed]
- Bokemeyer, C.; Berger, C.C.; Hartmann, J.T.; Kollmannsberger, C.; Schmoll, H.J.; Kuczyk, M.A.; Kanz, L. Analysis of risk factors for cisplatin-induced ototoxicity in patients with testicular cancer. Br. J. Cancer 1998, 77, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.; Teh, B.S.; Strother, D.R.; Davis, Q.G.; Chiu, J.K.; Lu, H.H.; Carpenter, L.S.; Mai, W.Y.; Chintagumpala, M.M.; South, M.; et al. Intensity-modulated radiation therapy for pediatric medulloblastoma: Early report on the reduction of ototoxicity. Int. J. Radiat. Oncol. Biol. Phys. 2002, 52, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Anniko, M.; Sobin, A. Cisplatin: Evaluation of its ototoxic potential. Am. J. Otolaryngol. 1986, 7, 276–293. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, N.; Whitworth, C.A.; Rybak, L.P. Acute changes in cochlear potentials due to cisplatin. Hear. Res. 2000, 149, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.I.; Hess, A.; Bloch, W.; Michel, O. Nitric oxide synthase inhibitor suppresses the ototoxic side effect of cisplatin in guinea pigs. Anticancer Drugs 2000, 11, 401–406. [Google Scholar] [CrossRef]
- Hong, O.; Kerr, M.J.; Poling, G.L.; Dhar, S. Understanding and preventing noise-induced hearing loss. Dis.-A-Mon. DM 2013, 59, 110–118. [Google Scholar] [CrossRef]
- Waissbluth, S.; Salehi, P.; He, X.; Daniel, S.J. Systemic dexamethasone for the prevention of cisplatin-induced ototoxicity. Eur. Arch. Oto-Rhino-Laryngol. 2013, 270, 1597–1605. [Google Scholar] [CrossRef]
- Fleischman, R.W.; Stadnicki, S.W.; Ethier, M.F.; Schaeppi, U. Ototoxicity of cis-dichlorodiammine platinum (ii) in the guinea pig. Toxicol. Appl. Pharmacol. 1975, 33, 320–332. [Google Scholar] [CrossRef]
- Stadnicki, S.W.; Fleischman, R.W.; Schaeppi, U.; Merriam, P. Cis-dichlorodiammineplatinum (ii) (nsc-119875): Hearing loss and other toxic effects in rhesus monkeys. Cancer Chemother. Rep. 1975, 59, 467–480. [Google Scholar]
- Staecker, H.; Liu, W.; Malgrange, B.; Lefebvre, P.P.; Van De Water, T.R. Vector-mediated delivery of bcl-2 prevents degeneration of auditory hair cells and neurons after injury. ORL 2007, 69, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Cai, J.; Xu, L.; Wang, H.; Liu, W. Cisplatin-induced stria vascularis damage is associated with inflammation and fibrosis. Neural Plast. 2020, 2020, 8851525. [Google Scholar] [CrossRef] [PubMed]
- Sluyter, S.; Klis, S.F.; de Groot, J.C.; Smoorenburg, G.F. Alterations in the stria vascularis in relation to cisplatin ototoxicity and recovery. Hear. Res. 2003, 185, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, J.; Ouchi, T. Steroid-responsive bilateral sensorineural hearing loss and immune complexes. Arch. Oto-Rhino-Laryngol. 1981, 230, 5–9. [Google Scholar] [CrossRef]
- Yoshida, K.; Ichimiya, I.; Suzuki, M.; Mogi, G. Effect of proinflammatory cytokines on cultured spiral ligament fibrocytes. Hear. Res. 1999, 137, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Firestein, G.S.; Billings, P.B.; Harris, J.P.; Keithley, E.M. Proinflammatory cytokine expression in the endolymphatic sac during inner ear inflammation. J. Assoc. Res. Otolaryngol. JARO 2003, 4, 139–147. [Google Scholar] [CrossRef]
- Kim, H.J.; So, H.S.; Lee, J.H.; Park, C.; Lee, J.B.; Youn, M.J.; Kim, S.J.; Yang, S.H.; Lee, K.M.; Kwon, K.B.; et al. Role of proinflammatory cytokines in cisplatin-induced vestibular hair cell damage. Head Neck 2008, 30, 1445–1456. [Google Scholar] [CrossRef]
- So, H.; Kim, H.; Kim, Y.; Kim, E.; Pae, H.O.; Chung, H.T.; Kim, H.J.; Kwon, K.B.; Lee, K.M.; Lee, H.Y.; et al. Evidence that cisplatin-induced auditory damage is attenuated by downregulation of pro-inflammatory cytokines via nrf2/ho-1. J. Assoc. Res. Otolaryngol. JARO 2008, 9, 290–306. [Google Scholar] [CrossRef]
- Watanabe, K.; Inai, S.; Jinnouchi, K.; Bada, S.; Hess, A.; Michel, O.; Yagi, T. Nuclear-factor kappa b (nf-kappa b)-inducible nitric oxide synthase (inos/nos ii) pathway damages the stria vascularis in cisplatin-treated mice. Anticancer. Res. 2002, 22, 4081–4085. [Google Scholar]
- Ries, F.; Klastersky, J. Nephrotoxicity induced by cancer chemotherapy with special emphasis on cisplatin toxicity. Am. J. Kidney Dis. 1986, 8, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Leibbrandt, M.E.; Wolfgang, G.H.; Metz, A.L.; Ozobia, A.A.; Haskins, J.R. Critical subcellular targets of cisplatin and related platinum analogs in rat renal proximal tubule cells. Kidney Int. 1995, 48, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Hayakawa, M.; Kato, T.; Hanaki, Y.; Shimizu, K.; Ozawa, T. Adverse effects of anti-tumor drug, cisplatin, on rat kidney mitochondria: Disturbances in glutathione peroxidase activity. Biochem. Biophys. Res. Commun. 1989, 159, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- So, H.S.; Park, C.; Kim, H.J.; Lee, J.H.; Park, S.Y.; Lee, J.H.; Lee, Z.W.; Kim, H.M.; Kalinec, F.; Lim, D.J.; et al. Protective effect of t-type calcium channel blocker flunarizine on cisplatin-induced death of auditory cells. Hear. Res. 2005, 204, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, M.S.; Silveira, A.F.; Teixeira, A.R.; Hyppolito, M.A. Mechanisms of cisplatin ototoxicity: Theoretical review. J. Laryngol. Otol. 2013, 127, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, H.; Yonemura, K.; Ohishi, K.; Hishida, A. The role of oxygen free radicals in cisplatin-induced acute renal failure in rats. J. Lab. Clin. Med. 1998, 131, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Rybak, L.P.; Whitworth, C.; Somani, S. Application of antioxidants and other agents to prevent cisplatin ototoxicity. Laryngoscope 1999, 109, 1740–1744. [Google Scholar] [CrossRef]
- Sergi, B.; Ferraresi, A.; Troiani, D.; Paludetti, G.; Fetoni, A.R. Cisplatin ototoxicity in the guinea pig: Vestibular and cochlear damage. Hear. Res. 2003, 182, 56–64. [Google Scholar] [CrossRef]
- Takumida, M.; Anniko, M. Simultaneous detection of both nitric oxide and reactive oxygen species in guinea pig vestibular sensory cells. ORL 2002, 64, 143–147. [Google Scholar] [CrossRef]
- Darlington, C.L.; Smith, P.F. Vestibulotoxicity following aminoglycoside antibiotics and its prevention. Curr. Opin. Investig. Drugs 2003, 4, 841–846. [Google Scholar] [PubMed]
- Rivolta, M.N.; Grix, N.; Lawlor, P.; Ashmore, J.F.; Jagger, D.J.; Holley, M.C. Auditory hair cell precursors immortalized from the mammalian inner ear. Proceedings. Biol. Sci. 1998, 265, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Benkafadar, N.; Menardo, J.; Bourien, J.; Nouvian, R.; Francois, F.; Decaudin, D.; Maiorano, D.; Puel, J.L.; Wang, J. Reversible p53 inhibition prevents cisplatin ototoxicity without blocking chemotherapeutic efficacy. EMBO Mol. Med. 2017, 9, 7–26. [Google Scholar] [CrossRef]
- Fredholm, B.B.; AP, I.J.; Jacobson, K.A.; Linden, J.; Muller, C.E. International union of basic and clinical pharmacology. Lxxxi. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef]
- Ramkumar, V.; Ravi, R.; Wilson, M.C.; Gettys, T.W.; Whitworth, C.; Rybak, L.P. Identification of a1 adenosine receptors in rat cochlea coupled to inhibition of adenylyl cyclase. Am. J. Physiol. 1994, 267, C731–C737. [Google Scholar] [CrossRef] [PubMed]
- Ford, M.S.; Nie, Z.; Whitworth, C.; Rybak, L.P.; Ramkumar, V. Up-regulation of adenosine receptors in the cochlea by cisplatin. Hear. Res. 1997, 111, 143–152. [Google Scholar] [CrossRef]
- Ford, M.S.; Maggirwar, S.B.; Rybak, L.P.; Whitworth, C.; Ramkumar, V. Expression and function of adenosine receptors in the chinchilla cochlea. Hear. Res. 1997, 105, 130–140. [Google Scholar] [CrossRef]
- Bryant, G.M.; Barron, S.E.; Norris, C.H.; Guth, P.S. Adenosine is a modulator of hair cell-afferent neurotransmission. Hear. Res. 1987, 30, 231–237. [Google Scholar] [CrossRef]
- Nario, K.; Kitano, I.; Mori, N.; Matsunaga, T. The effect of adenosine on cochlear potentials in the guinea pig. Eur. Arch. Oto-Rhino-Laryngol. 1994, 251, 428–433. [Google Scholar] [CrossRef]
- Hight, N.G.; McFadden, S.L.; Henderson, D.; Burkard, R.F.; Nicotera, T. Noise-induced hearing loss in chinchillas pre-treated with glutathione monoethylester and r-pia. Hear. Res. 2003, 179, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Vlajkovic, S.M.; Guo, C.X.; Telang, R.; Wong, A.C.; Paramananthasivam, V.; Boison, D.; Housley, G.D.; Thorne, P.R. Adenosine kinase inhibition in the cochlea delays the onset of age-related hearing loss. Exp. Gerontol. 2011, 46, 905–914. [Google Scholar] [CrossRef]
- Whitworth, C.A.; Ramkumar, V.; Jones, B.; Tsukasaki, N.; Rybak, L.P. Protection against cisplatin ototoxicity by adenosine agonists. Biochem. Pharmacol. 2004, 67, 1801–1807. [Google Scholar] [CrossRef]
- Kaur, T.; Mukherjea, D.; Sheehan, K.; Jajoo, S.; Rybak, L.P.; Ramkumar, V. Short interfering rna against stat1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death Dis. 2011, 2, e180. [Google Scholar] [CrossRef]
- Demmler, G.J. Infectious diseases society of america and centers for disease control. Summary of a workshop on surveillance for congenital cytomegalovirus disease. Rev. Infect. Dis. 1991, 13, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Pass, R.F.; Cloud, G.; Britt, W.J.; Henderson, R.E.; Walton, P.D.; Veren, D.A.; Page, F.; Alford, C.A. Primary cytomegalovirus infection in pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA 1986, 256, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Barbi, M.; Binda, S.; Caroppo, S.; Primache, V. Neonatal screening for congenital cytomegalovirus infection and hearing loss. J. Clin. Virol. 2006, 35, 206–209. [Google Scholar] [CrossRef]
- Dreher, A.M.; Arora, N.; Fowler, K.B.; Novak, Z.; Britt, W.J.; Boppana, S.B.; Ross, S.A. Spectrum of disease and outcome in children with symptomatic congenital cytomegalovirus infection. J. Pediatr. 2014, 164, 855–859. [Google Scholar] [CrossRef]
- Shi, X.; Dong, Y.; Li, Y.; Zhao, Z.; Li, H.; Qiu, S.; Li, Y.; Guo, W.; Qiao, Y. Inflammasome activation in mouse inner ear in response to mcmv induced hearing loss. J. Otol. 2015, 10, 143–149. [Google Scholar] [CrossRef]
- Schachtele, S.J.; Mutnal, M.B.; Schleiss, M.R.; Lokensgard, J.R. Cytomegalovirus-induced sensorineural hearing loss with persistent cochlear inflammation in neonatal mice. J. Neurovirol. 2011, 17, 201–211. [Google Scholar] [CrossRef]
- Neag, M.A.; Mocan, A.; Echeverria, J.; Pop, R.M.; Bocsan, C.I.; Crisan, G.; Buzoianu, A.D. Berberine: Botanical occurrence, traditional uses, extraction methods, and relevance in cardiovascular, metabolic, hepatic, and renal disorders. Front. Pharmacol. 2018, 9, 557. [Google Scholar] [CrossRef]
- Katsarkas, A.; Ayukawa, H. Hearing loss due to aging (presbycusis). J. Otolaryngol. 1986, 15, 239–244. [Google Scholar]
- Gennis, V.; Garry, P.J.; Haaland, K.Y.; Yeo, R.A.; Goodwin, J.S. Hearing and cognition in the elderly. New findings and a review of the literature. Arch. Intern. Med. 1991, 151, 2259–2264. [Google Scholar] [CrossRef] [PubMed]
- Gates, G.A.; Mills, J.H. Presbycusis. Lancet 2005, 366, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Solfrizzi, V.; Logroscino, G. Age-related hearing impairment-a risk factor and frailty marker for dementia and ad. Nat. Rev. Neurol. 2015, 11, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, B.R.; Brewster, K.; Golub, J.S.; Kim, A.H.; Roose, S.P. Sensation and psychiatry: Linking age-related hearing loss to late-life depression and cognitive decline. Am. J. Psychiatry 2018, 175, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Schrepfer, T.; Schacht, J. Age-related hearing impairment and the triad of acquired hearing loss. Front. Cell. Neurosci. 2015, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Scholtz, A.W.; Kammen-Jolly, K.; Felder, E.; Hussl, B.; Rask-Andersen, H.; Schrott-Fischer, A. Selective aspects of human pathology in high-tone hearing loss of the aging inner ear. Hear. Res. 2001, 157, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhong, Y.; Peng, W.; Sun, Y.; Kong, W.J. Age-related changes in the central auditory system: Comparison of d-galactose-induced aging rats and naturally aging rats. Brain Res. 2010, 1344, 43–53. [Google Scholar] [CrossRef]
- Chen, B.; Zhong, Y.; Peng, W.; Sun, Y.; Hu, Y.J.; Yang, Y.; Kong, W.J. Increased mitochondrial DNA damage and decreased base excision repair in the auditory cortex of d-galactose-induced aging rats. Mol. Biol. Rep. 2011, 38, 3635–3642. [Google Scholar] [CrossRef]
- Du, Z.; Yang, Y.; Hu, Y.; Sun, Y.; Zhang, S.; Peng, W.; Zhong, Y.; Huang, X.; Kong, W. A long-term high-fat diet increases oxidative stress, mitochondrial damage and apoptosis in the inner ear of d-galactose-induced aging rats. Hear. Res. 2012, 287, 15–24. [Google Scholar] [CrossRef]
- Cui, X.; Zuo, P.; Zhang, Q.; Li, X.; Hu, Y.; Long, J.; Packer, L.; Liu, J. Chronic systemic d-galactose exposure induces memory loss, neurodegeneration, and oxidative damage in mice: Protective effects of r-alpha-lipoic acid. J. Neurosci. Res. 2006, 83, 1584–1590. [Google Scholar] [CrossRef]
- Hua, X.; Lei, M.; Zhang, Y.; Ding, J.; Han, Q.; Hu, G.; Xiao, M. Long-term d-galactose injection combined with ovariectomy serves as a new rodent model for alzheimer’s disease. Life Sci. 2007, 80, 1897–1905. [Google Scholar] [CrossRef]
- Lu, J.; Zheng, Y.L.; Wu, D.M.; Luo, L.; Sun, D.X.; Shan, Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by d-galactose. Biochem. Pharmacol. 2007, 74, 1078–1090. [Google Scholar] [CrossRef]
- Chen, C.F.; Lang, S.Y.; Zuo, P.P.; Yang, N.; Wang, X.Q.; Xia, C. Effects of d-galactose on the expression of hippocampal peripheral-type benzodiazepine receptor and spatial memory performances in rats. Psychoneuroendocrinology 2006, 31, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Dogra, S. Naringin alleviates cognitive impairment, mitochondrial dysfunction and oxidative stress induced by d-galactose in mice. Food Chem. Toxicol. 2010, 48, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Lu, J.; Zheng, Y.L.; Hu, B.; Fan, S.H.; Wu, D.M.; Zheng, Z.H.; Shan, Q.; Liu, C.M. Purple sweet potato color protects mouse liver against d-galactose-induced apoptosis via inhibiting caspase-3 activation and enhancing pi3k/akt pathway. Food Chem. Toxicol. 2010, 48, 2500–2507. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Hua, X.; Xiao, M.; Ding, J.; Han, Q.; Hu, G. Impairments of astrocytes are involved in the d-galactose-induced brain aging. Biochem. Biophys. Res. Commun. 2008, 369, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.M.; Wu, W.M.; Hu, M.L. Soy isoflavones attenuate oxidative stress and improve parameters related to aging and alzheimer’s disease in c57bl/6j mice treated with d-galactose. Food Chem. Toxicol. 2009, 47, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, L.; Zuo, P.; Han, Z.; Fang, Z.; Li, W.; Liu, J. D-galactose-caused life shortening in drosophila melanogaster and musca domestica is associated with oxidative stress. Biogerontology 2004, 5, 317–325. [Google Scholar] [CrossRef]
- Kong, W.J.; Wang, Y.; Wang, Q.; Hu, Y.J.; Han, Y.C.; Liu, J. The relation between d-galactose injection and mitochondrial DNA 4834 bp deletion mutation. Exp. Gerontol. 2006, 41, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.J.; Hu, Y.J.; Wang, Q.; Wang, Y.; Han, Y.C.; Cheng, H.M.; Kong, W.; Guan, M.X. The effect of the mtdna4834 deletion on hearing. Biochem. Biophys. Res. Commun. 2006, 344, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Hu, Y.J.; Yang, Y.; Peng, W.; Sun, Y.; Chen, B.; Huang, X.; Kong, W.J. Contribution of common deletion to total deletion burden in mitochondrial DNA from inner ear of d-galactose-induced aging rats. Mutat. Res. 2011, 712, 11–19. [Google Scholar] [CrossRef]
- Zhong, Y.; Hu, Y.J.; Chen, B.; Peng, W.; Sun, Y.; Yang, Y.; Zhao, X.Y.; Fan, G.R.; Huang, X.; Kong, W.J. Mitochondrial transcription factor a overexpression and base excision repair deficiency in the inner ear of rats with d-galactose-induced aging. FEBS J. 2011, 278, 2500–2510. [Google Scholar] [CrossRef] [PubMed]
- Yamasoba, T.; Someya, S.; Yamada, C.; Weindruch, R.; Prolla, T.A.; Tanokura, M. Role of mitochondrial dysfunction and mitochondrial DNA mutations in age-related hearing loss. Hear. Res. 2007, 226, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Druzhyna, N.M.; Wilson, G.L.; LeDoux, S.P. Mitochondrial DNA repair in aging and disease. Mech. Ageing Dev. 2008, 129, 383–390. [Google Scholar] [CrossRef]
- Meissner, C.; Bruse, P.; Mohamed, S.A.; Schulz, A.; Warnk, H.; Storm, T.; Oehmichen, M. The 4977 bp deletion of mitochondrial DNA in human skeletal muscle, heart and different areas of the brain: A useful biomarker or more? Exp. Gerontol. 2008, 43, 645–652. [Google Scholar] [CrossRef]
- Markaryan, A.; Nelson, E.G.; Hinojosa, R. Quantification of the mitochondrial DNA common deletion in presbycusis. Laryngoscope 2009, 119, 1184–1189. [Google Scholar] [CrossRef]
- Hiona, A.; Leeuwenburgh, C. The role of mitochondrial DNA mutations in aging and sarcopenia: Implications for the mitochondrial vicious cycle theory of aging. Exp. Gerontol. 2008, 43, 24–33. [Google Scholar] [CrossRef]
- Basu, S.; Xie, X.; Uhler, J.P.; Hedberg-Oldfors, C.; Milenkovic, D.; Baris, O.R.; Kimoloi, S.; Matic, S.; Stewart, J.B.; Larsson, N.G.; et al. Accurate mapping of mitochondrial DNA deletions and duplications using deep sequencing. PLoS Genet. 2020, 16, e1009242. [Google Scholar] [CrossRef]
- Schon, E.A.; Rizzuto, R.; Moraes, C.T.; Nakase, H.; Zeviani, M.; DiMauro, S. A direct repeat is a hotspot for large-scale deletion of human mitochondrial DNA. Science 1989, 244, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Pang, C.Y.; Hsu, H.S.; Wei, Y.H. Differential accumulations of 4,977 bp deletion in mitochondrial DNA of various tissues in human ageing. Biochim. Biophys. Acta 1994, 1226, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Hanke, T.; Erasmi, A.W.; Bechtel, M.J.; Scharfschwerdt, M.; Meissner, C.; Sievers, H.H.; Gosslau, A. Mitochondrial DNA deletions and the aging heart. Exp. Gerontol. 2006, 41, 508–517. [Google Scholar] [CrossRef]
- Nakano, Y.; Longo-Guess, C.M.; Bergstrom, D.E.; Nauseef, W.M.; Jones, S.M.; Banfi, B. Mutation of the cyba gene encoding p22phox causes vestibular and immune defects in mice. J. Clin. Investig. 2008, 118, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F.; Salmon, P.; Bredl, S.; Cherpin, O.; Coelho, M.; Myburgh, R.; Alessandrini, M.; Perny, M.; Roccio, M.; Speck, R.F.; et al. Optimizing synthetic mirna minigene architecture for efficient mirna hairpin concatenation and multi-target gene knockdown. Mol. Ther. Nucleic Acids 2019, 14, 351–363. [Google Scholar] [CrossRef]
- Zhang, S.; Xing, J.; Gong, Y.; Li, P.; Wang, B.; Xu, L. Downregulation of vdr in benign paroxysmal positional vertigo patients inhibits otolith-associated protein expression levels. Mol. Med. Rep. 2021, 24, 591. [Google Scholar] [CrossRef]
- Froehling, D.A.; Silverstein, M.D.; Mohr, D.N.; Beatty, C.W.; Offord, K.P.; Ballard, D.J. Benign positional vertigo: Incidence and prognosis in a population-based study in olmsted county, minnesota. Mayo Clin. Proc. 1991, 66, 596–601. [Google Scholar] [CrossRef]
- Imai, T.; Higashi-Shingai, K.; Takimoto, Y.; Masumura, C.; Hattori, K.; Inohara, H. New scoring system of an interview for the diagnosis of benign paroxysmal positional vertigo. Acta Oto-Laryngol. 2016, 136, 283–288. [Google Scholar] [CrossRef]
- Fife, T.D.; Iverson, D.J.; Lempert, T.; Furman, J.M.; Baloh, R.W.; Tusa, R.J.; Hain, T.C.; Herdman, S.; Morrow, M.J.; Gronseth, G.S.; et al. Practice parameter: Therapies for benign paroxysmal positional vertigo (an evidence-based review): Report of the quality standards subcommittee of the american academy of neurology. Neurology 2008, 70, 2067–2074. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Gubbels, S.P.; Schwartz, S.R.; Edlow, J.A.; El-Kashlan, H.; Fife, T.; Holmberg, J.M.; Mahoney, K.; Hollingsworth, D.B.; Roberts, R.; et al. Clinical practice guideline: Benign paroxysmal positional vertigo (update). Otolaryngol.-Head Neck Surg. 2017, 156, S1–S47. [Google Scholar] [CrossRef]
- Jang, Y.S.; Hwang, C.H.; Shin, J.Y.; Bae, W.Y.; Kim, L.S. Age-related changes on the morphology of the otoconia. Laryngoscope 2006, 116, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Baloh, R.W.; Honrubia, V.; Jacobson, K. Benign positional vertigo: Clinical and oculographic features in 240 cases. Neurology 1987, 37, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Pyykko, I.; Arroll, M.A.; Casselbrant, M.L.; Foster, C.A.; Manzoor, N.F.; Megerian, C.A.; Naganawa, S.; Young, Y.H. Meniere’s disease. Nat. Rev. Dis. Primers 2016, 2, 16028. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kim, J.S.; Shin, J.W.; Kim, S.; Lee, H.; Lee, A.Y.; Kim, J.M.; Jo, H.; Song, J.; Ghim, Y. Decreased serum vitamin d in idiopathic benign paroxysmal positional vertigo. J. Neurol. 2013, 260, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Gu, C.; Han, W.; Lu, X.; Chen, C.; Fan, Z. Reduction of bone mineral density in native chinese female idiopathic benign paroxysmal positional vertigo patients. Am J Otolaryngol 2018, 39, 31–33. [Google Scholar] [CrossRef] [PubMed]
- Yoda, S.; Cureoglu, S.; Yildirim-Baylan, M.; Morita, N.; Fukushima, H.; Harada, T.; Paparella, M.M. Association between type 1 diabetes mellitus and deposits in the semicircular canals. Otolaryngol. Head Neck Surg. 2011, 145, 458–462. [Google Scholar] [CrossRef]
- Holick, M.F. Resurrection of vitamin d deficiency and rickets. J. Clin. Investig. 2006, 116, 2062–2072. [Google Scholar] [CrossRef]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin d: Metabolism, molecular mechanism of action, and pleiotropic effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef]
- Editorial Board of Chinese Journal of Otorhinolaryngology Head and Neck Surgery; Society of Otorhinolaryngology Head and Neck Surgery Chinese Medical Association. Guideline of diagnosis and treatment of benign paroxysmal positional vertigo (2017). Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi Chin. J. Otorhinolaryngol. Head Neck Surg. 2017, 52, 173–177. [Google Scholar]
- Barnes, P.J.; Shapiro, S.D.; Pauwels, R.A. Chronic obstructive pulmonary disease: Molecular and cellular mechanisms. Eur. Respir. J. 2003, 22, 672–688. [Google Scholar] [CrossRef]
- Lokke, A.; Lange, P.; Scharling, H.; Fabricius, P.; Vestbo, J. Developing copd: A 25 year follow up study of the general population. Thorax 2006, 61, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, T.; Cho, C.Y.; Thimmulappa, R.K.; Zhen, L.; Srisuma, S.S.; Kensler, T.W.; Yamamoto, M.; Petrache, I.; Tuder, R.M.; Biswal, S. Genetic ablation of nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J. Clin. Investig. 2004, 114, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- MacNee, W. Pulmonary and systemic oxidant/antioxidant imbalance in chronic obstructive pulmonary disease. Proc. Am. Thorac. Soc. 2005, 2, 50–60. [Google Scholar] [CrossRef]
- Taggart, C.; Cervantes-Laurean, D.; Kim, G.; McElvaney, N.G.; Wehr, N.; Moss, J.; Levine, R.L. Oxidation of either methionine 351 or methionine 358 in alpha 1-antitrypsin causes loss of anti-neutrophil elastase activity. J. Biol. Chem. 2000, 275, 27258–27265. [Google Scholar] [CrossRef]
- Zhang, X.; Shan, P.; Sasidhar, M.; Chupp, G.L.; Flavell, R.A.; Choi, A.M.; Lee, P.J. Reactive oxygen species and extracellular signal-regulated kinase 1/2 mitogen-activated protein kinase mediate hyperoxia-induced cell death in lung epithelium. Am. J. Respir. Cell Mol. Biol. 2003, 28, 305–315. [Google Scholar] [CrossRef]
- Ishii, Y.; Itoh, K.; Morishima, Y.; Kimura, T.; Kiwamoto, T.; Iizuka, T.; Hegab, A.E.; Hosoya, T.; Nomura, A.; Sakamoto, T.; et al. Transcription factor nrf2 plays a pivotal role in protection against elastase-induced pulmonary inflammation and emphysema. J. Immunol. 2005, 175, 6968–6975. [Google Scholar] [CrossRef]
- Pons, J.; Sauleda, J.; Regueiro, V.; Santos, C.; Lopez, M.; Ferrer, J.; Agusti, A.G.; Bengoechea, J.A. Expression of toll-like receptor 2 is up-regulated in monocytes from patients with chronic obstructive pulmonary disease. Respir. Res. 2006, 7, 64. [Google Scholar] [CrossRef]
- Sukkar, M.B.; Xie, S.; Khorasani, N.M.; Kon, O.M.; Stanbridge, R.; Issa, R.; Chung, K.F. Toll-like receptor 2, 3, and 4 expression and function in human airway smooth muscle. J. Allergy Clin. Immunol. 2006, 118, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008, 133, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Marwick, J.A. Molecular mechanisms of oxidative stress in airways and lungs with reference to asthma and chronic obstructive pulmonary disease. Ann. N. Y. Acad. Sci. 2010, 1203, 85–91. [Google Scholar] [CrossRef]
- Pratico, D.; Basili, S.; Vieri, M.; Cordova, C.; Violi, F.; Fitzgerald, G.A. Chronic obstructive pulmonary disease is associated with an increase in urinary levels of isoprostane f2alpha-iii, an index of oxidant stress. Am. J. Respir. Crit. Care Med. 1998, 158, 1709–1714. [Google Scholar] [CrossRef]
- Lius, E.E.; Syafaah, I. Hyperoxia in the management of respiratory failure: A literature review. Ann. Med. Surg. 2022, 81, 104393. [Google Scholar] [CrossRef] [PubMed]
- Lilien, T.A.; van Meenen, D.M.P.; Schultz, M.J.; Bos, L.D.J.; Bem, R.A. Hyperoxia-induced lung injury in acute respiratory distress syndrome: What is its relative impact? Am. J. Physiol. Lung Cell. Mol. Physiol. 2023, 325, L9–L16. [Google Scholar] [CrossRef]
- Minkove, S.; Dhamapurkar, R.; Cui, X.; Li, Y.; Sun, J.; Cooper, D.; Eichacker, P.Q.; Torabi-Parizi, P. Effect of low-to-moderate hyperoxia on lung injury in preclinical animal models: A systematic review and meta-analysis. Intensive Care Med. Exp. 2023, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Kallet, R.H.; Matthay, M.A. Hyperoxic acute lung injury. Respir. Care 2013, 58, 123–141. [Google Scholar] [CrossRef]
- Zhang, X.; Shan, P.; Qureshi, S.; Homer, R.; Medzhitov, R.; Noble, P.W.; Lee, P.J. Cutting edge: Tlr4 deficiency confers susceptibility to lethal oxidant lung injury. J. Immunol. 2005, 175, 4834–4838. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Shan, P.; Hunt, C.R.; Pandita, T.K.; Lee, P.J. A protective hsp70-tlr4 pathway in lethal oxidant lung injury. J. Immunol. 2013, 191, 1393–1403. [Google Scholar] [CrossRef]
- Hunt, C.R.; Dix, D.J.; Sharma, G.G.; Pandita, R.K.; Gupta, A.; Funk, M.; Pandita, T.K. Genomic instability and enhanced radiosensitivity in hsp70.1- and hsp70.3-deficient mice. Mol. Cell. Biol. 2004, 24, 899–911. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Palsson-McDermott, E.M.; Bowie, A.G.; Jefferies, C.A.; Mansell, A.S.; Brady, G.; Brint, E.; Dunne, A.; Gray, P.; Harte, M.T.; et al. Mal (myd88-adapter-like) is required for toll-like receptor-4 signal transduction. Nature 2001, 413, 78–83. [Google Scholar] [CrossRef]
- Rajpoot, S.; Wary, K.K.; Ibbott, R.; Liu, D.; Saqib, U.; Thurston, T.L.M.; Baig, M.S. Tirap in the mechanism of inflammation. Front. Immunol. 2021, 12, 697588. [Google Scholar] [CrossRef]
- Ruwanpura, S.M.; McLeod, L.; Lilja, A.R.; Brooks, G.; Dousha, L.F.; Seow, H.J.; Bozinovski, S.; Vlahos, R.; Hertzog, P.J.; Anderson, G.P.; et al. Non-essential role for tlr2 and its signaling adaptor mal/tirap in preserving normal lung architecture in mice. PLoS ONE 2013, 8, e78095. [Google Scholar] [CrossRef] [PubMed]
- Ruwanpura, S.M.; McLeod, L.; Miller, A.; Jones, J.; Bozinovski, S.; Vlahos, R.; Ernst, M.; Armes, J.; Bardin, P.G.; Anderson, G.P.; et al. Interleukin-6 promotes pulmonary emphysema associated with apoptosis in mice. Am. J. Respir. Cell Mol. Biol. 2011, 45, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Ruwanpura, S.M.; McLeod, L.; Miller, A.; Jones, J.; Vlahos, R.; Ramm, G.; Longano, A.; Bardin, P.G.; Bozinovski, S.; Anderson, G.P.; et al. Deregulated stat3 signaling dissociates pulmonary inflammation from emphysema in gp130 mutant mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L627–L639. [Google Scholar] [CrossRef] [PubMed]
- Hantos, Z.; Adamicza, A.; Janosi, T.Z.; Szabari, M.V.; Tolnai, J.; Suki, B. Lung volumes and respiratory mechanics in elastase-induced emphysema in mice. J. Appl. Physiol. 2008, 105, 1864–1872. [Google Scholar] [CrossRef] [PubMed]
- King, T.E., Jr.; Schwarz, M.I.; Brown, K.; Tooze, J.A.; Colby, T.V.; Waldron, J.A., Jr.; Flint, A.; Thurlbeck, W.; Cherniack, R.M. Idiopathic pulmonary fibrosis: Relationship between histopathologic features and mortality. Am. J. Respir. Crit. Care Med. 2001, 164, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Navaratnam, V.; Fleming, K.M.; West, J.; Smith, C.J.; Jenkins, R.G.; Fogarty, A.; Hubbard, R.B. The rising incidence of idiopathic pulmonary fibrosis in the UK. Thorax 2011, 66, 462–467. [Google Scholar] [CrossRef]
- Zisman, D.A.; Keane, M.P.; Belperio, J.A.; Strieter, R.M.; Lynch, J.P. Pulmonary fibrosis. In Fibrosis Research: Methods and Protocols; Varga, J., Brenner, D.A., Phan, S.H., Eds.; Humana Press: Totowa, NJ, USA, 2005; pp. 3–44. [Google Scholar]
- Bocchino, M.; Agnese, S.; Fagone, E.; Svegliati, S.; Grieco, D.; Vancheri, C.; Gabrielli, A.; Sanduzzi, A.; Avvedimento, E.V. Reactive oxygen species are required for maintenance and differentiation of primary lung fibroblasts in idiopathic pulmonary fibrosis. PLoS ONE 2010, 5, e14003. [Google Scholar] [CrossRef]
- Manoury, B.; Nenan, S.; Leclerc, O.; Guenon, I.; Boichot, E.; Planquois, J.M.; Bertrand, C.P.; Lagente, V. The absence of reactive oxygen species production protects mice against bleomycin-induced pulmonary fibrosis. Respir. Res. 2005, 6, 11. [Google Scholar] [CrossRef]
- Murthy, S.; Adamcakova-Dodd, A.; Perry, S.S.; Tephly, L.A.; Keller, R.M.; Metwali, N.; Meyerholz, D.K.; Wang, Y.; Glogauer, M.; Thorne, P.S.; et al. Modulation of reactive oxygen species by rac1 or catalase prevents asbestos-induced pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L846–L855. [Google Scholar] [CrossRef]
- Kinnula, V.L.; Fattman, C.L.; Tan, R.J.; Oury, T.D. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am. J. Respir. Crit. Care Med. 2005, 172, 417–422. [Google Scholar] [CrossRef]
- Korfei, M.; von der Beck, D.; Henneke, I.; Markart, P.; Ruppert, C.; Mahavadi, P.; Ghanim, B.; Klepetko, W.; Fink, L.; Meiners, S.; et al. Comparative proteome analysis of lung tissue from patients with idiopathic pulmonary fibrosis (ipf), non-specific interstitial pneumonia (nsip) and organ donors. J. Proteom. 2013, 85, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Montuschi, P.; Ciabattoni, G.; Paredi, P.; Pantelidis, P.; du Bois, R.M.; Kharitonov, S.A.; Barnes, P.J. 8-isoprostane as a biomarker of oxidative stress in interstitial lung diseases. Am. J. Respir. Crit. Care Med. 1998, 158, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.; Allan, G.J.; Lochrie, J.D.; Flint, D.J. Insulin-like growth factor-binding protein-5 (igfbp-5): A critical member of the igf axis. Biochem. J. 2006, 395, 1–19. [Google Scholar] [CrossRef]
- Han, N.; Zhang, F.; Li, G.; Zhang, X.; Lin, X.; Yang, H.; Wang, L.; Cao, Y.; Du, J.; Fan, Z. Local application of igfbp5 protein enhanced periodontal tissue regeneration via increasing the migration, cell proliferation and osteo/dentinogenic differentiation of mesenchymal stem cells in an inflammatory niche. Stem Cell Res. Ther. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, K.; Lemire, M.; Potvin, C.; Tremblay, A.; Hunninghake, G.M.; Raby, B.A.; Hudson, T.J.; Perez-Iratxeta, C.; Andrade-Navarro, M.A.; Laprise, C. Genes to diseases (g2d) computational method to identify asthma candidate genes. PLoS ONE 2008, 3, e2907. [Google Scholar] [CrossRef]
- Perez-Iratxeta, C.; Wjst, M.; Bork, P.; Andrade, M.A. G2d: A tool for mining genes associated with disease. BMC Genet 2005, 6, 45. [Google Scholar] [CrossRef]
- Perez-Iratxeta, C.; Bork, P.; Andrade-Navarro, M.A. Update of the g2d tool for prioritization of gene candidates to inherited diseases. Nucleic Acids Res. 2007, 35, W212–W216. [Google Scholar] [CrossRef]
- Heyer, E.; Tremblay, M. Variability of the genetic contribution of quebec population founders associated to some deleterious genes. Am. J. Hum. Genet. 1995, 56, 970–978. [Google Scholar]
- Richter, A.; Rioux, J.D.; Bouchard, J.P.; Mercier, J.; Mathieu, J.; Ge, B.; Poirier, J.; Julien, D.; Gyapay, G.; Weissenbach, J.; et al. Location score and haplotype analyses of the locus for autosomal recessive spastic ataxia of charlevoix-saguenay, in chromosome region 13q11. Am. J. Hum. Genet. 1999, 64, 768–775. [Google Scholar] [CrossRef]
- Yin, C.; Li, K.; Yu, Y.; Huang, H.; Yu, Y.; Wang, Z.; Yan, J.; Pu, Y.; Li, Z.; Li, D.; et al. Genome-wide association study identifies loci and candidate genes for non-idiopathic pulmonary hypertension in eastern chinese han population. BMC Pulm. Med. 2018, 18, 158. [Google Scholar] [CrossRef]
- Barman, S.A.; Chen, F.; Li, X.; Haigh, S.; Stepp, D.W.; Kondrikov, D.; Mahboubi, K.; Bordan, Z.; Traber, P.; Su, Y.; et al. Galectin-3 promotes vascular remodeling and contributes to pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2018, 197, 1488–1492. [Google Scholar] [CrossRef]
- Soubrier, F.; Chung, W.K.; Machado, R.; Grunig, E.; Aldred, M.; Geraci, M.; Loyd, J.E.; Elliott, C.G.; Trembath, R.C.; Newman, J.H.; et al. Genetics and genomics of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2013, 62, D13–D21. [Google Scholar] [CrossRef]
- Cantu, E.; Shah, R.J.; Lin, W.; Daye, Z.J.; Diamond, J.M.; Suzuki, Y.; Ellis, J.H.; Borders, C.F.; Andah, G.A.; Beduhn, B.; et al. Oxidant stress regulatory genetic variation in recipients and donors contributes to risk of primary graft dysfunction after lung transplantation. J. Thorac. Cardiovasc. Surg. 2015, 149, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Charles, E.J.; Kron, I.L. One step closer to the elimination of primary graft dysfunction. J. Thorac. Cardiovasc. Surg. 2015, 149, 602–603. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.D.; Carby, M.; Bag, R.; Corris, P.; Hertz, M.; Weill, D. Report of the ISHLT Working Group on Primary Lung Graft Dysfunction Part II: Definition. A consensus statement of the international society for heart and lung transplantation. J. Heart Lung Transplant 2005, 24, 1454–1459. [Google Scholar] [CrossRef] [PubMed]
- Christie, J.D.; Edwards, L.B.; Kucheryavaya, A.Y.; Aurora, P.; Dobbels, F.; Kirk, R.; Rahmel, A.O.; Stehlik, J.; Hertz, M.I. The registry of the international society for heart and lung transplantation: Twenty-seventh official adult lung and heart-lung transplant report—2010. J. Heart Lung Transplant. 2010, 29, 1104–1118. [Google Scholar] [CrossRef]
- Suzuki, Y.; Cantu, E.; Christie, J.D. Primary graft dysfunction. Semin. Respir. Crit. Care Med. 2013, 34, 305–319. [Google Scholar] [CrossRef] [PubMed]
- de Perrot, M.; Liu, M.; Waddell, T.K.; Keshavjee, S. Ischemia-reperfusion-induced lung injury. Am. J. Respir. Crit. Care Med. 2003, 167, 490–511. [Google Scholar] [CrossRef]
- Williams, A.; Riise, G.C.; Anderson, B.A.; Kjellstrom, C.; Schersten, H.; Kelly, F.J. Compromised antioxidant status and persistent oxidative stress in lung transplant recipients. Free Radic. Res. 1999, 30, 383–393. [Google Scholar] [CrossRef]
- Rega, F.R.; Wuyts, W.A.; Vanaudenaerde, B.M.; Jannis, N.C.; Neyrinck, A.P.; Verleden, G.M.; Lerut, T.E.; Van Raemdonck, D.E. Nebulized n-acetyl cysteine protects the pulmonary graft inside the non-heart-beating donor. J. Heart Lung Transplant. 2005, 24, 1369–1377. [Google Scholar] [CrossRef]
- Kozower, B.D.; Christofidou-Solomidou, M.; Sweitzer, T.D.; Muro, S.; Buerk, D.G.; Solomides, C.C.; Albelda, S.M.; Patterson, G.A.; Muzykantov, V.R. Immunotargeting of catalase to the pulmonary endothelium alleviates oxidative stress and reduces acute lung transplantation injury. Nat. Biotechnol. 2003, 21, 392–398. [Google Scholar] [CrossRef]
- Cho, H.Y.; Reddy, S.P.; Kleeberger, S.R. Nrf2 defends the lung from oxidative stress. Antioxid. Redox Signal. 2006, 8, 76–87. [Google Scholar] [CrossRef]
- Lubos, E.; Mahoney, C.E.; Leopold, J.A.; Zhang, Y.Y.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 modulates lipopolysaccharide-induced adhesion molecule expression in endothelial cells by altering cd14 expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 2525–2532. [Google Scholar]
- Hamanishi, T.; Furuta, H.; Kato, H.; Doi, A.; Tamai, M.; Shimomura, H.; Sakagashira, S.; Nishi, M.; Sasaki, H.; Sanke, T.; et al. Functional variants in the glutathione peroxidase-1 (gpx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in japanese type 2 diabetic patients. Diabetes 2004, 53, 2455–2460. [Google Scholar] [CrossRef]
- Liu, P.; Shi, L.; Cang, X.; Huang, J.; Wu, X.; Yan, J.; Chen, L.; Cui, S.; Ye, X. Ctbp2 ameliorates palmitate-induced insulin resistance in hepg2 cells through ros mediated jnk pathway. Gen. Comp. Endocrinol. 2017, 247, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Styskal, J.; Van Remmen, H.; Richardson, A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radic. Biol. Med. 2012, 52, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling, and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, E.J.; Diamond-Stanic, M.K.; Marchionne, E.M. Oxidative stress and the etiology of insulin resistance and type 2 diabetes. Free Radic. Biol. Med. 2011, 51, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metab. Clin. Exp. 2008, 57, 1071–1077. [Google Scholar] [CrossRef]
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Zhou, B.G.; Sheng, J.Q.; Chen, Y.; Cao, Y.Q.; Chen, C. Molecular mechanisms of hepatic insulin resistance in nonalcoholic fatty liver disease and potential treatment strategies. Pharmacol. Res. 2020, 159, 104984. [Google Scholar] [CrossRef]
- Santoleri, D.; Titchenell, P.M. Resolving the paradox of hepatic insulin resistance. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 447–456. [Google Scholar] [CrossRef]
- Boden, G.; Shulman, G.I. Free fatty acids in obesity and type 2 diabetes: Defining their role in the development of insulin resistance and beta-cell dysfunction. Eur. J. Clin. Investig. 2002, 32, 14–23. [Google Scholar] [CrossRef]
- Collaboration, N.C.D.R.F. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Montonen, J.; Knekt, P.; Harkanen, T.; Jarvinen, R.; Heliovaara, M.; Aromaa, A.; Reunanen, A. Dietary patterns and the incidence of type 2 diabetes. Am. J. Epidemiol. 2005, 161, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Parks, E.; Yki-Jarvinen, H.; Hawkins, M. Out of the frying pan: Dietary saturated fat influences nonalcoholic fatty liver disease. J. Clin. Investig. 2017, 127, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the international diabetes federation diabetes atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef]
- Magnusson, I.; Rothman, D.L.; Katz, L.D.; Shulman, R.G.; Shulman, G.I. Increased rate of gluconeogenesis in type ii diabetes mellitus. A 13c nuclear magnetic resonance study. J. Clin. Investig. 1992, 90, 1323–1327. [Google Scholar] [CrossRef]
- Bock, G.; Chittilapilly, E.; Basu, R.; Toffolo, G.; Cobelli, C.; Chandramouli, V.; Landau, B.R.; Rizza, R.A. Contribution of hepatic and extrahepatic insulin resistance to the pathogenesis of impaired fasting glucose: Role of increased rates of gluconeogenesis. Diabetes 2007, 56, 1703–1711. [Google Scholar] [CrossRef]
- Basu, R.; Barosa, C.; Jones, J.; Dube, S.; Carter, R.; Basu, A.; Rizza, R.A. Pathogenesis of prediabetes: Role of the liver in isolated fasting hyperglycemia and combined fasting and postprandial hyperglycemia. J. Clin. Endocrinol. Metab. 2013, 98, E409–E417. [Google Scholar] [CrossRef]
- Funatsu, H.; Yamashita, H. Molecular biology in development and progression of diabetic retinopathy. Nihon Rinsho Jpn. J. Clin. Med. 2002, 60, 162–166. [Google Scholar]
- Wilkinson, C.P.; Ferris, F.L., 3rd; Klein, R.E.; Lee, P.P.; Agardh, C.D.; Davis, M.; Dills, D.; Kampik, A.; Pararajasegaram, R.; Verdaguer, J.T.; et al. Proposed international clinical diabetic retinopathy and diabetic macular edema disease severity scales. Ophthalmology 2003, 110, 1677–1682. [Google Scholar] [CrossRef]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuniga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7. [Google Scholar] [CrossRef]
- Bettaieb, A.; Vazquez Prieto, M.A.; Rodriguez Lanzi, C.; Miatello, R.M.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (-)-epicatechin mitigates high-fructose-associated insulin resistance by modulating redox signaling and endoplasmic reticulum stress. Free Radic. Biol. Med. 2014, 72, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Cremonini, E.; Kang, H.; Kang, J.; Haj, F.G.; Oteiza, P.I. Anti-inflammatory actions of (-)-epicatechin in the adipose tissue of obese mice. Int. J. Biochem. Cell Biol. 2016, 81, 383–392. [Google Scholar] [CrossRef]
- Cremonini, E.; Wang, Z.; Bettaieb, A.; Adamo, A.M.; Daveri, E.; Mills, D.A.; Kalanetra, K.M.; Haj, F.G.; Karakas, S.; Oteiza, P.I. (-)-epicatechin protects the intestinal barrier from high fat diet-induced permeabilization: Implications for steatosis and insulin resistance. Redox Biol. 2018, 14, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Loh, Y.P.; Cheng, Y.; Mahata, S.K.; Corti, A.; Tota, B. Chromogranin a and derived peptides in health and disease. J. Mol. Neurosci. MN 2012, 48, 347–356. [Google Scholar] [CrossRef]
- Hossain, Z.; Valicherla, G.R.; Gupta, A.P.; Syed, A.A.; Riyazuddin, M.; Chandra, S.; Siddiqi, M.I.; Gayen, J.R. Discovery of pancreastatin inhibitor psti8 for the treatment of insulin resistance and diabetes: Studies in rodent models of diabetes mellitus. Sci. Rep. 2018, 8, 8715. [Google Scholar] [CrossRef]
- Sanchez-Margalet, V.; Gonzalez-Yanes, C. Pancreastatin inhibits insulin action in rat adipocytes. Am. J. Physiol. 1998, 275, E1055–E1060. [Google Scholar] [CrossRef]
- Gayen, J.R.; Saberi, M.; Schenk, S.; Biswas, N.; Vaingankar, S.M.; Cheung, W.W.; Najjar, S.M.; O’Connor, D.T.; Bandyopadhyay, G.; Mahata, S.K. A novel pathway of insulin sensitivity in chromogranin a null mice: A crucial role for pancreastatin in glucose homeostasis. J. Biol. Chem. 2009, 284, 28498–28509. [Google Scholar] [CrossRef] [PubMed]
- Tatemoto, K.; Efendic, S.; Mutt, V.; Makk, G.; Feistner, G.J.; Barchas, J.D. Pancreastatin, a novel pancreatic peptide that inhibits insulin secretion. Nature 1986, 324, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, G.K.; Lu, M.; Avolio, E.; Siddiqui, J.A.; Gayen, J.R.; Wollam, J.; Vu, C.U.; Chi, N.W.; O’Connor, D.T.; Mahata, S.K. Pancreastatin-dependent inflammatory signaling mediates obesity-induced insulin resistance. Diabetes 2015, 64, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Broedbaek, K.; Hilsted, L. Chromogranin a as biomarker in diabetes. Biomark. Med. 2016, 10, 1181–1189. [Google Scholar] [CrossRef]
- Allu, P.K.; Chirasani, V.R.; Ghosh, D.; Mani, A.; Bera, A.K.; Maji, S.K.; Senapati, S.; Mullasari, A.S.; Mahapatra, N.R. Naturally occurring variants of the dysglycemic peptide pancreastatin: Differential potencies for multiple cellular functions and structure-function correlation. J. Biol. Chem. 2014, 289, 4455–4469. [Google Scholar] [CrossRef]
- Caturano, A.; D’Angelo, M.; Mormone, A.; Russo, V.; Mollica, M.P.; Salvatore, T.; Galiero, R.; Rinaldi, L.; Vetrano, E.; Marfella, R.; et al. Oxidative stress in type 2 diabetes: Impacts from pathogenesis to lifestyle modifications. Curr. Issues Mol. Biol. 2023, 45, 6651–6666. [Google Scholar] [CrossRef]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Rajan, S.; Shankar, K.; Beg, M.; Varshney, S.; Gupta, A.; Srivastava, A.; Kumar, D.; Mishra, R.K.; Hussain, Z.; Gayen, J.R.; et al. Chronic hyperinsulinemia reduces insulin sensitivity and metabolic functions of brown adipocyte. J. Endocrinol. 2016, 230, 275–290. [Google Scholar] [CrossRef]
- Garg, R.; Gupta, A.P.; Katekar, R.; Verma, S.; Goand, U.K.; Dadge, S.; Gayen, J.R. Pancreastatin inhibitor psti8 prevents free fatty acid-induced oxidative stress and insulin resistance by modulating jnk pathway: In vitro and in vivo findings. Life Sci. 2022, 289, 120221. [Google Scholar] [CrossRef]
- Mouche, S.; Mkaddem, S.B.; Wang, W.; Katic, M.; Tseng, Y.H.; Carnesecchi, S.; Steger, K.; Foti, M.; Meier, C.A.; Muzzin, P.; et al. Reduced expression of the NADPH oxidase nox4 is a hallmark of adipocyte differentiation. Biochim. Biophys. Acta 2007, 1773, 1015–1027. [Google Scholar] [CrossRef]
- Schroder, K.; Wandzioch, K.; Helmcke, I.; Brandes, R.P. Nox4 acts as a switch between differentiation and proliferation in preadipocytes. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.A.; Acharya, J.D.; Mehendale, N.K.; Kamat, S.S.; Ghaskadbi, S.S. Pterostilbene reverses palmitic acid mediated insulin resistance in hepg2 cells by reducing oxidative stress and triglyceride accumulation. Free Radic. Res. 2019, 53, 815–827. [Google Scholar] [CrossRef]
- Remsberg, C.M.; Yanez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. PTR 2008, 22, 169–179. [Google Scholar] [CrossRef]
- Acharya, J.D.; Ghaskadbi, S.S. Protective effect of pterostilbene against free radical mediated oxidative damage. BMC Complement. Altern. Med. 2013, 13, 238. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, M.A.; Pari, L. Effect of pterostilbene on lipids and lipid profiles in streptozotocin-nicotinamide induced type 2 diabetes mellitus. J. Appl. Biomed. 2008, 6, 31–37. [Google Scholar] [CrossRef]
- Elango, B.; Dornadula, S.; Paulmurugan, R.; Ramkumar, K.M. Pterostilbene ameliorates streptozotocin-induced diabetes through enhancing antioxidant signaling pathways mediated by nrf2. Chem. Res. Toxicol. 2016, 29, 47–57. [Google Scholar] [CrossRef]
- Gomez-Zorita, S.; Fernandez-Quintela, A.; Aguirre, L.; Macarulla, M.T.; Rimando, A.M.; Portillo, M.P. Pterostilbene improves glycaemic control in rats fed an obesogenic diet: Involvement of skeletal muscle and liver. Food Funct. 2015, 6, 1968–1976. [Google Scholar] [CrossRef]
- Pihlajaniemi, T.; Myllyla, R.; Kivirikko, K.I.; Tryggvason, K. Effects of streptozotocin diabetes, glucose, and insulin on the metabolism of type iv collagen and proteoglycan in murine basement membrane-forming ehs tumor tissue. J. Biol. Chem. 1982, 257, 14914–14920. [Google Scholar] [CrossRef]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive oxygen species (ros) and wound healing: The functional role of ros and emerging ros-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, H.Y.; Jang, J.Y.; Lee, H.R.; Shin, Y.S.; Kim, C.H. Redox treatment ameliorates diabetes mellitus-induced skin flap necrosis via inhibiting apoptosis and promoting neoangiogenesis. Exp. Biol. Med. 2021, 246, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, K.; Fujii, K.; Kamouchi, M.; Nakane, H.; Arihiro, S.; Okada, Y.; Ibayashi, S.; Iida, M. Free radical scavenger, edaravone, in stroke with internal carotid artery occlusion. J. Neurol. Sci. 2004, 221, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, D.; Luo, W.; Yu, J.; Li, J.; Yu, Y.; Zhang, X.; Chen, J.; Wu, X.R.; Huang, C. E3 ligase activity of xiap ring domain is required for xiap-mediated cancer cell migration, but not for its rhogdi binding activity. PLoS ONE 2012, 7, e35682. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Lau, D.C. The obesity epidemic and its impact on hypertension. Can. J. Cardiol. 2012, 28, 326–333. [Google Scholar] [CrossRef]
- Franklin, S.S.; Gustin, W.t.; Wong, N.D.; Larson, M.G.; Weber, M.A.; Kannel, W.B.; Levy, D. Hemodynamic patterns of age-related changes in blood pressure. The framingham heart study. Circulation 1997, 96, 308–315. [Google Scholar] [CrossRef]
- Pravenec, M.; Wallace, C.; Aitman, T.J.; Kurtz, T.W. Gene expression profiling in hypertension research: A critical perspective. Hypertension 2003, 41, 3–8. [Google Scholar] [CrossRef]
- Radkowski, P.; Wator, G.; Skupien, J.; Bogdali, A.; Wolkow, P. Analysis of gene expression to predict dynamics of future hypertension incidence in type 2 diabetic patients. BMC Proc. 2016, 10, 113–117. [Google Scholar] [CrossRef]
- Chen, G.; Adeyemo, A.A.; Zhou, J.; Chen, Y.; Doumatey, A.; Lashley, K.; Huang, H.; Amoah, A.; Agyenim-Boateng, K.; Eghan, B.A., Jr.; et al. A genome-wide search for linkage to renal function phenotypes in west africans with type 2 diabetes. Am. J. Kidney Dis. 2007, 49, 394–400. [Google Scholar] [CrossRef]
- Kwak, S.H.; Hernandez-Cancela, R.B.; DiCorpo, D.A.; Condon, D.E.; Merino, J.; Wu, P.; Brody, J.A.; Yao, J.; Guo, X.; Ahmadizar, F.; et al. Time-to-event genome-wide association study for incident cardiovascular disease in people with type 2 diabetes mellitus. medRxiv 2023. [Google Scholar] [CrossRef]
- Monteiro, R.; Azevedo, I. Chronic inflammation in obesity and the metabolic syndrome. Mediat. Inflamm. 2010, 2010, 289645. [Google Scholar] [CrossRef]
- Hotamisligil, G.S.; Arner, P.; Caro, J.F.; Atkinson, R.L.; Spiegelman, B.M. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J. Clin. Investig. 1995, 95, 2409–2415. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Talior, I.; Yarkoni, M.; Bashan, N.; Eldar-Finkelman, H. Increased glucose uptake promotes oxidative stress and pkc-delta activation in adipocytes of obese, insulin-resistant mice. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E295–E302. [Google Scholar] [CrossRef] [PubMed]
- Wajngarten, M.; Silva, G.S. Hypertension and stroke: Update on treatment. Eur. Cardiol. 2019, 14, 111–115. [Google Scholar] [CrossRef]
- Johansson, B.B. Hypertension mechanisms causing stroke. Clin. Exp. Pharmacol. Physiol. 1999, 26, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Neaton, J.D.; Blackburn, H.; Jacobs, D.; Kuller, L.; Lee, D.J.; Sherwin, R.; Shih, J.; Stamler, J.; Wentworth, D. Serum cholesterol level and mortality findings for men screened in the multiple risk factor intervention trial. Multiple risk factor intervention trial research group. Arch. Intern. Med. 1992, 152, 1490–1500. [Google Scholar] [CrossRef] [PubMed]
- Peeling, J.; Yan, H.J.; Chen, S.G.; Campbell, M.; Del Bigio, M.R. Protective effects of free radical inhibitors in intracerebral hemorrhage in rat. Brain Res. 1998, 795, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Aronowski, J.; Zhao, X. Molecular pathophysiology of cerebral hemorrhage: Secondary brain injury. Stroke 2011, 42, 1781–1786. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Yamamoto, K.; Morita, N.; Ohta, Y.; Chikugo, T.; Higashizawa, T.; Suzuki, T. Establishment and use of the m strain of stroke-prone spontaneously hypertensive rat. J. Hypertension. Suppl. 1986, 4, S21–S24. [Google Scholar]
- Iritani, N.; Fukuda, E.; Nara, Y.; Yamori, Y. Lipid metabolism in spontaneously hypertensive rats (shr). Atherosclerosis 1977, 28, 217–222. [Google Scholar] [CrossRef]
- Tanito, M.; Nakamura, H.; Kwon, Y.W.; Teratani, A.; Masutani, H.; Shioji, K.; Kishimoto, C.; Ohira, A.; Horie, R.; Yodoi, J. Enhanced oxidative stress and impaired thioredoxin expression in spontaneously hypertensive rats. Antioxid. Redox Signal. 2004, 6, 89–97. [Google Scholar] [CrossRef]
- Michihara, A.; Shimatani, M.; Anraku, M.; Tomida, H.; Akasaki, K. High levels of oxidative stress exist in the brain than serum or kidneys in stroke-prone spontaneously hypertensive rats at ten weeks of age. Biol. Pharm. Bull. 2010, 33, 518–521. [Google Scholar] [CrossRef]
- Loft, S.; Vistisen, K.; Ewertz, M.; Tjonneland, A.; Overvad, K.; Poulsen, H.E. Oxidative DNA damage estimated by 8-hydroxydeoxyguanosine excretion in humans: Influence of smoking, gender and body mass index. Carcinogenesis 1992, 13, 2241–2247. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamamoto, E.; Kataoka, K.; Yamashita, T.; Tokutomi, Y.; Dong, Y.F.; Matsuba, S.; Ogawa, H.; Kim-Mitsuyama, S. Pioglitazone exerts protective effects against stroke in stroke-prone spontaneously hypertensive rats, independently of blood pressure. Stroke 2007, 38, 3016–3022. [Google Scholar] [CrossRef] [PubMed]
- Nediani, C.; Borchi, E.; Giordano, C.; Baruzzo, S.; Ponziani, V.; Sebastiani, M.; Nassi, P.; Mugelli, A.; d’Amati, G.; Cerbai, E. NADPH oxidase-dependent redox signaling in human heart failure: Relationship between the left and right ventricle. J. Mol. Cell Cardiol. 2007, 42, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Borchi, E.; Bargelli, V.; Stillitano, F.; Giordano, C.; Sebastiani, M.; Nassi, P.A.; d’Amati, G.; Cerbai, E.; Nediani, C. Enhanced ros production by NADPH oxidase is correlated to changes in antioxidant enzyme activity in human heart failure. Biochim. Biophys. Acta 2010, 1802, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Maack, C.; Kartes, T.; Kilter, H.; Schafers, H.J.; Nickenig, G.; Bohm, M.; Laufs, U. Oxygen free radical release in human failing myocardium is associated with increased activity of rac1-gtpase and represents a target for statin treatment. Circulation 2003, 108, 1567–1574. [Google Scholar] [CrossRef]
- Zhang, M.; Perino, A.; Ghigo, A.; Hirsch, E.; Shah, A.M. NADPH oxidases in heart failure: Poachers or gamekeepers? Antioxid. Redox Signal. 2013, 18, 1024–1041. [Google Scholar] [CrossRef]
- Fenyo, I.M.; Florea, I.C.; Raicu, M.; Manea, A. Tyrphostin ag490 reduces napdh oxidase activity and expression in the aorta of hypercholesterolemic apolipoprotein e-deficient mice. Vasc. Pharmacol. 2011, 54, 100–106. [Google Scholar] [CrossRef]
- Sirker, A.; Murdoch, C.E.; Protti, A.; Sawyer, G.J.; Santos, C.X.; Martin, D.; Zhang, X.; Brewer, A.C.; Zhang, M.; Shah, A.M. Cell-specific effects of nox2 on the acute and chronic response to myocardial infarction. J. Mol. Cell Cardiol. 2016, 98, 11–17. [Google Scholar] [CrossRef]
- Bell, R.M.; Cave, A.C.; Johar, S.; Hearse, D.J.; Shah, A.M.; Shattock, M.J. Pivotal role of nox-2-containing NADPH oxidase in early ischemic preconditioning. FASEB J. 2005, 19, 2037–2039. [Google Scholar] [CrossRef]
- Wind, S.; Beuerlein, K.; Armitage, M.E.; Taye, A.; Kumar, A.H.; Janowitz, D.; Neff, C.; Shah, A.M.; Wingler, K.; Schmidt, H.H. Oxidative stress and endothelial dysfunction in aortas of aged spontaneously hypertensive rats by nox1/2 is reversed by NADPH oxidase inhibition. Hypertension 2010, 56, 490–497. [Google Scholar] [CrossRef]
- Bkaily, G.; Najibeddine, W.; Jacques, D. Increase of NADPH oxidase 3 in heart failure of hereditary cardiomyopathy (1). Can. J. Physiol. Pharmacol. 2019, 97, 902–908. [Google Scholar] [CrossRef]
- Dhalla, A.K.; Hill, M.F.; Singal, P.K. Role of oxidative stress in transition of hypertrophy to heart failure. J. Am. Coll. Cardiol. 1996, 28, 506–514. [Google Scholar] [CrossRef]
- Wu, R.; Yin, D.; Sadekova, N.; Deschepper, C.F.; de Champlain, J.; Girouard, H. Protective effects of aspirin from cardiac hypertrophy and oxidative stress in cardiomyopathic hamsters. Oxidative Med. Cell. Longev. 2012, 2012, 761710. [Google Scholar] [CrossRef] [PubMed]
- Mollnau, H.; Oelze, M.; August, M.; Wendt, M.; Daiber, A.; Schulz, E.; Baldus, S.; Kleschyov, A.L.; Materne, A.; Wenzel, P.; et al. Mechanisms of increased vascular superoxide production in an experimental model of idiopathic dilated cardiomyopathy. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2554–2559. [Google Scholar] [CrossRef] [PubMed]
- Jacques, D.; Bkaily, G. Endocardial endothelial cell hypertrophy takes place during the development of hereditary cardiomyopathy. Mol. Cell Biochem. 2019, 453, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, N.S.; Elmoselhi, A.B.; Hata, T.; Makino, N. Status of myocardial antioxidants in ischemia-reperfusion injury. Cardiovasc. Res. 2000, 47, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Bolli, R.; Jeroudi, M.O.; Patel, B.S.; Aruoma, O.I.; Halliwell, B.; Lai, E.K.; McCay, P.B. Marked reduction of free radical generation and contractile dysfunction by antioxidant therapy begun at the time of reperfusion. Evidence that myocardial “stunning” is a manifestation of reperfusion injury. Circ. Res. 1989, 65, 607–622. [Google Scholar] [CrossRef]
- Zhao, Z.Q.; Nakamura, M.; Wang, N.P.; Wilcox, J.N.; Shearer, S.; Ronson, R.S.; Guyton, R.A.; Vinten-Johansen, J. Reperfusion induces myocardial apoptotic cell death. Cardiovasc. Res. 2000, 45, 651–660. [Google Scholar] [CrossRef]
- Frangogiannis, N.G.; Smith, C.W.; Entman, M.L. The inflammatory response in myocardial infarction. Cardiovasc. Res. 2002, 53, 31–47. [Google Scholar] [CrossRef]
- Kloner, R.A.; Jennings, R.B. Consequences of brief ischemia: Stunning, preconditioning, and their clinical implications: Part 2. Circulation 2001, 104, 3158–3167. [Google Scholar] [CrossRef]
- Hayashidani, S.; Tsutsui, H.; Shiomi, T.; Ikeuchi, M.; Matsusaka, H.; Suematsu, N.; Wen, J.; Egashira, K.; Takeshita, A. Anti-monocyte chemoattractant protein-1 gene therapy attenuates left ventricular remodeling and failure after experimental myocardial infarction. Circulation 2003, 108, 2134–2140. [Google Scholar] [CrossRef]
- Kaikita, K.; Hayasaki, T.; Okuma, T.; Kuziel, W.A.; Ogawa, H.; Takeya, M. Targeted deletion of cc chemokine receptor 2 attenuates left ventricular remodeling after experimental myocardial infarction. Am. J. Pathol. 2004, 165, 439–447. [Google Scholar] [CrossRef]
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. Ccl2/monocyte chemoattractant protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ. Res. 2005, 96, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Hirose, M.; Takahashi, M.; Kawaguchi, M.; Ise, H.; Kolattukudy, P.E.; Yamada, M.; Ikeda, U. Mcp-1 induces cardioprotection against ischaemia/reperfusion injury: Role of reactive oxygen species. Cardiovasc. Res. 2008, 78, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Kolattukudy, P.E.; Quach, T.; Bergese, S.; Breckenridge, S.; Hensley, J.; Altschuld, R.; Gordillo, G.; Klenotic, S.; Orosz, C.; Parker-Thornburg, J. Myocarditis induced by targeted expression of the mcp-1 gene in murine cardiac muscle. Am. J. Pathol. 1998, 152, 101–111. [Google Scholar] [PubMed]
- Vats, S.; Sundquist, K.; Sundquist, J.; Zhang, N.; Wang, X.; Acosta, S.; Gottsater, A.; Memon, A.A. Oxidative stress-related genetic variation and antioxidant vitamin intake in intact and ruptured abdominal aortic aneurysm: A swedish population-based retrospective cohort study. Eur. J. Prev. Cardiol. 2024, 31, 61–74. [Google Scholar] [CrossRef]
- Manjer, J.; Elmstahl, S.; Janzon, L.; Berglund, G. Invitation to a population-based cohort study: Differences between subjects recruited using various strategies. Scand. J. Public Health 2002, 30, 103–112. [Google Scholar] [CrossRef]
- Wanhainen, A.; Verzini, F.; Van Herzeele, I.; Allaire, E.; Bown, M.; Cohnert, T.; Dick, F.; van Herwaarden, J.; Karkos, C.; Koelemay, M.; et al. Editor’s choice—European society for vascular surgery (ESVS) 2019 clinical practice guidelines on the management of abdominal aorto-iliac artery aneurysms. Eur. J. Vasc. Endovasc. Surg. 2019, 57, 8–93. [Google Scholar] [CrossRef]
- Howard, D.P.; Banerjee, A.; Fairhead, J.F.; Handa, A.; Silver, L.E.; Rothwell, P.M.; Oxford Vascular, S. Age-specific incidence, risk factors and outcome of acute abdominal aortic aneurysms in a defined population. Br. J. Surg. 2015, 102, 907–915. [Google Scholar] [CrossRef]
- Reimerink, J.J.; van der Laan, M.J.; Koelemay, M.J.; Balm, R.; Legemate, D.A. Systematic review and meta-analysis of population-based mortality from ruptured abdominal aortic aneurysm. Br. J. Surg. 2013, 100, 1405–1413. [Google Scholar] [CrossRef]
- Song, P.; He, Y.; Adeloye, D.; Zhu, Y.; Ye, X.; Yi, Q.; Rahimi, K.; Rudan, I.; Global Health Epidemiology Research Group. The global and regional prevalence of abdominal aortic aneurysms: A systematic review and modeling analysis. Ann. Surg. 2023, 277, 912–919. [Google Scholar] [CrossRef]
- Sweeting, M.J.; Thompson, S.G.; Brown, L.C.; Powell, J.T.; Collaborators, R. Meta-analysis of individual patient data to examine factors affecting growth and rupture of small abdominal aortic aneurysms. Br. J. Surg. 2012, 99, 655–665. [Google Scholar] [CrossRef]
- Guzik, B.; Sagan, A.; Ludew, D.; Mrowiecki, W.; Chwala, M.; Bujak-Gizycka, B.; Filip, G.; Grudzien, G.; Kapelak, B.; Zmudka, K.; et al. Mechanisms of oxidative stress in human aortic aneurysms--association with clinical risk factors for atherosclerosis and disease severity. Int. J. Cardiol. 2013, 168, 2389–2396. [Google Scholar] [CrossRef] [PubMed]
- McCormick, M.L.; Gavrila, D.; Weintraub, N.L. Role of oxidative stress in the pathogenesis of abdominal aortic aneurysms. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Blomkalns, A.L.; Ogbi, M.; Thomas, M.; Gavrila, D.; Neltner, B.S.; Cassis, L.A.; Thompson, R.W.; Weiss, R.M.; Lindower, P.D.; et al. Role of myeloperoxidase in abdominal aortic aneurysm formation: Mitigation by taurine. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H1168–H1179. [Google Scholar] [CrossRef] [PubMed]
- Ludvigsson, J.F.; Andersson, E.; Ekbom, A.; Feychting, M.; Kim, J.L.; Reuterwall, C.; Heurgren, M.; Olausson, P.O. External review and validation of the swedish national inpatient register. BMC Public Health 2011, 11, 450. [Google Scholar] [CrossRef] [PubMed]
- Caramori, M.L.; Fioretto, P.; Mauer, M. Low glomerular filtration rate in normoalbuminuric type 1 diabetic patients: An indicator of more advanced glomerular lesions. Diabetes 2003, 52, 1036–1040. [Google Scholar] [CrossRef]
- Cockcroft, D.W.; Gault, M.H. Prediction of creatinine clearance from serum creatinine. Nephron 1976, 16, 31–41. [Google Scholar] [CrossRef]
- Hunt, S.C.; Hasstedt, S.J.; Coon, H.; Camp, N.J.; Cawthon, R.M.; Wu, L.L.; Hopkins, P.N. Linkage of creatinine clearance to chromosome 10 in utah pedigrees replicates a locus for end-stage renal disease in humans and renal failure in the fawn-hooded rat. Kidney Int. 2002, 62, 1143–1148. [Google Scholar] [CrossRef]
- Rokutan, K.; Kawahara, T.; Kuwano, Y.; Tominaga, K.; Nishida, K.; Teshima-Kondo, S. Nox enzymes and oxidative stress in the immunopathology of the gastrointestinal tract. Semin. Immunopathol. 2008, 30, 315–327. [Google Scholar] [CrossRef]
- Salles, N.; Szanto, I.; Herrmann, F.; Armenian, B.; Stumm, M.; Stauffer, E.; Michel, J.P.; Krause, K.H. Expression of mrna for ros-generating NADPH oxidases in the aging stomach. Exp. Gerontol. 2005, 40, 353–357. [Google Scholar] [CrossRef]
- Augusto, A.C.; Miguel, F.; Mendonca, S.; Pedrazzoli, J., Jr.; Gurgueira, S.A. Oxidative stress expression status associated to Helicobacter pylori virulence in gastric diseases. Clin. Biochem. 2007, 40, 615–622. [Google Scholar] [CrossRef]
- El Hassani, R.A.; Benfares, N.; Caillou, B.; Talbot, M.; Sabourin, J.C.; Belotte, V.; Morand, S.; Gnidehou, S.; Agnandji, D.; Ohayon, R.; et al. Dual oxidase2 is expressed all along the digestive tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G933–G942. [Google Scholar] [CrossRef]
- Plantinga, T.S.; Arts, P.; Knarren, G.H.; Mulder, A.H.; Wakelkamp, I.M.; Hermus, A.R.; Joosten, L.A.; Netea, M.G.; Bisschop, P.H.; de Herder, W.W.; et al. Rare nox3 variants confer susceptibility to agranulocytosis during thyrostatic treatment of graves’ disease. Clin. Pharmacol. Ther. 2017, 102, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhong, J.; Xiao, X.H.; Zhou, L.Z.; Chen, Y.J.; Liu, J.H.; Cao, R.X.; Wen, G.B. The relationship between bone marrow characteristics and the clinical prognosis of antithyroid drug-induced agranulocytosis. Endocr. J. 2013, 60, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, N.; Ohnishi, H.; Nishiuma, T.; Doukuni, R.; Umezawa, K.; Oozone, S.; Kuramoto, E.; Yoshimura, S.; Kinami, S. Antithyroid drug-induced agranulocytosis complicated by pneumococcal sepsis and upper airway obstruction. Intern. Med. 2013, 52, 2355–2359. [Google Scholar] [CrossRef] [PubMed]
- Kubota, Y.; Toh Yoon, E.W. Severe drug-induced agranulocytosis successfully treated with recombinant human granulocyte colony-stimulating factor. Case Rep. Med. 2018, 2018, 8439791. [Google Scholar]
- Andrès, E.; Maloisel, F. Antibiotic-induced agranulocytosis: A monocentric study of 21 cases. Arch. Intern. Med. 2001, 161, 2619. [Google Scholar] [CrossRef]
- di Fonzo, H.; Villegas Gutsch, M.; Castroagudin, A.; Cabrera, M.V.; Mazzei, M.E.; Rueda, D. Agranulocytosis induced by vancomycin. Case report and literature review. Am. J. Case Rep. 2018, 19, 1053–1056. [Google Scholar] [CrossRef]
- Weissel, M. Propylthiouracil: Clinical overview of its efficacy and its side effects more than 50 years after the introduction of its use in thyrostatic treatment. Exp. Clin. Endocrinol. Diabetes 2010, 118, 101–104. [Google Scholar] [CrossRef]
- Sato, S.; Noh, J.Y.; Sato, S.; Suzuki, M.; Yasuda, S.; Matsumoto, M.; Kunii, Y.; Mukasa, K.; Sugino, K.; Ito, K.; et al. Comparison of efficacy and adverse effects between methimazole 15 mg+inorganic iodine 38 mg/day and methimazole 30 mg/day as initial therapy for graves’ disease patients with moderate to severe hyperthyroidism. Thyroid 2015, 25, 43–50. [Google Scholar] [CrossRef]
- Taylor, P.N.; Vaidya, B. Side effects of anti-thyroid drugs and their impact on the choice of treatment for thyrotoxicosis in pregnancy. Eur. Thyroid J. 2012, 1, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Andersohn, F.; Konzen, C.; Garbe, E. Systematic review: Agranulocytosis induced by nonchemotherapy drugs. Ann. Intern. Med. 2007, 146, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Takata, K.; Kubota, S.; Fukata, S.; Kudo, T.; Nishihara, E.; Ito, M.; Amino, N.; Miyauchi, A. Methimazole-induced agranulocytosis in patients with graves’ disease is more frequent with an initial dose of 30 mg daily than with 15 mg daily. Thyroid 2009, 19, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Pan, C.M.; Zhao, S.X.; Liang, J.; Gao, G.Q.; Zhang, X.M.; Yuan, G.Y.; Li, C.G.; Xue, L.Q.; Shen, M.; et al. A genome-wide association study identifies two new risk loci for graves’ disease. Nat. Genet. 2011, 43, 897–901. [Google Scholar] [PubMed]
- Liu, W.; Wang, H.N.; Gu, Z.H.; Yang, S.Y.; Ye, X.P.; Pan, C.M.; Zhao, S.X.; Xue, L.Q.; Xie, H.J.; Yu, S.S.; et al. Identification of bach2 as a susceptibility gene for graves’ disease in the chinese han population based on a three-stage genome-wide association study. Hum. Genet. 2014, 133, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Roderique, J.D.; Josef, C.S.; Feldman, M.J.; Spiess, B.D. A modern literature review of carbon monoxide poisoning theories, therapies, and potential targets for therapy advancement. Toxicology 2015, 334, 45–58. [Google Scholar] [CrossRef]
- Varma, D.R.; Mulay, S.; Chemtob, S. Chapter 20—Carbon monoxide: From public health risk to painless killer. In Handbook of Toxicology of Chemical Warfare Agents; Gupta, R.C., Ed.; Academic Press: San Diego, CA, USA, 2009; pp. 271–292. [Google Scholar]
- Ginsberg, M.D. Delayed neurological deterioration following hypoxia. Adv. Neurol. 1979, 26, 21–44. [Google Scholar]
- Sun, Q.; Cai, J.; Zhou, J.; Tao, H.; Zhang, J.H.; Zhang, W.; Sun, X.J. Hydrogen-rich saline reduces delayed neurologic sequelae in experimental carbon monoxide toxicity. Crit. Care Med. 2011, 39, 765–769. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Piantadosi, C.A.; Zhang, J.; Demchenko, I.T. Production of hydroxyl radical in the hippocampus after co hypoxia or hypoxic hypoxia in the rat. Free Radic. Biol. Med. 1997, 22, 725–732. [Google Scholar] [CrossRef]
- Hara, S.; Kobayashi, M.; Kuriiwa, F.; Mukai, T.; Mizukami, H. Different mechanisms of hydroxyl radical production susceptible to purine p2 receptor antagonists between carbon monoxide poisoning and exogenous atp in rat striatum. Free Radic. Res. 2014, 48, 1322–1333. [Google Scholar] [CrossRef]
- Hara, S.; Kobayash, M.; Kuriiwa, F.; Kurosaki, K.; Mizukami, H. Gene expression in rat striatum following carbon monoxide poisoning. Genom Data 2017, 12, 74–75. [Google Scholar] [CrossRef]
- Thom, S.R.; Ohnishi, S.T.; Ischiropoulos, H. Nitric oxide released by platelets inhibits neutrophil b2 integrin function following acute carbon monoxide poisoning. Toxicol. Appl. Pharmacol. 1994, 128, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Katoh, A.; Ishimaru, H.; Yoneda, Y.; Ogita, K.; Murase, K.; Ohtsuka, H.; Inari, K.; Fukuta, T.; Kameyama, T. Carbon monoxide-induced delayed amnesia, delayed neuronal death and change in acetylcholine concentration in mice. J. Pharmacol. Exp. Ther. 1991, 256, 378–384. [Google Scholar] [PubMed]
- Fujita, K.; Seike, T.; Yutsudo, N.; Ohno, M.; Yamada, H.; Yamaguchi, H.; Sakumi, K.; Yamakawa, Y.; Kido, M.A.; Takaki, A.; et al. Hydrogen in drinking water reduces dopaminergic neuronal loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of parkinson’s disease. PLoS ONE 2009, 4, e7247. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, L.; Holopainen, S.; Pessa-Morikawa, T.; Lappalainen, A.K.; Hytonen, M.K.; Lohi, H.; Iivanainen, A. Genetic dissection of canine hip dysplasia phenotypes and osteoarthritis reveals three novel loci. BMC Genom. 2019, 20, 1027. [Google Scholar] [CrossRef] [PubMed]
- Skurkova, L.; Hluchy, M.; Lackova, M.; Mihalova, M.; Ledecky, V. Relation of the norberg angle and position of the femoral head centre to the dorsal acetabular edge in evaluation of canine hip dysplasia. Vet. Comp. Orthop. Traumatol. 2010, 23, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Broeckx, B.J.G.; Vezzoni, A.; Bogaerts, E.; Bertal, M.; Bosmans, T.; Stock, E.; Deforce, D.; Peelman, L.; Saunders, J.H. Comparison of three methods to quantify laxity in the canine hip joint. Vet. Comp. Orthop. Traumatol. V.C.O.T 2018, 31, 23–29. [Google Scholar] [PubMed]
- Sanchez-Molano, E.; Woolliams, J.A.; Pong-Wong, R.; Clements, D.N.; Blott, S.C.; Wiener, P. Quantitative trait loci mapping for canine hip dysplasia and its related traits in uk labrador retrievers. BMC Genom. 2014, 15, 833. [Google Scholar] [CrossRef]
- Lavrijsen, I.C.; Leegwater, P.A.; Martin, A.J.; Harris, S.J.; Tryfonidou, M.A.; Heuven, H.C.; Hazewinkel, H.A. Genome wide analysis indicates genes for basement membrane and cartilage matrix proteins as candidates for hip dysplasia in labrador retrievers. PLoS ONE 2014, 9, e87735. [Google Scholar] [CrossRef]
- Fels, L.; Distl, O. Identification and validation of quantitative trait loci (qtl) for canine hip dysplasia (chd) in german shepherd dogs. PLoS ONE 2014, 9, e96618. [Google Scholar] [CrossRef]
- Pfahler, S.; Distl, O. Identification of quantitative trait loci (qtl) for canine hip dysplasia and canine elbow dysplasia in bernese mountain dogs. PLoS ONE 2012, 7, e49782. [Google Scholar] [CrossRef]
- Zhou, Z.; Sheng, X.; Zhang, Z.; Zhao, K.; Zhu, L.; Guo, G.; Friedenberg, S.G.; Hunter, L.S.; Vandenberg-Foels, W.S.; Hornbuckle, W.E.; et al. Differential genetic regulation of canine hip dysplasia and osteoarthritis. PLoS ONE 2010, 5, e13219. [Google Scholar] [CrossRef]
- Carlstrom, K.E.; Ewing, E.; Granqvist, M.; Gyllenberg, A.; Aeinehband, S.; Enoksson, S.L.; Checa, A.; Badam, T.V.S.; Huang, J.; Gomez-Cabrero, D.; et al. Therapeutic efficacy of dimethyl fumarate in relapsing-remitting multiple sclerosis associates with ros pathway in monocytes. Nat. Commun. 2019, 10, 3081. [Google Scholar] [CrossRef] [PubMed]
- Gopal, S.; Mikulskis, A.; Gold, R.; Fox, R.J.; Dawson, K.T.; Amaravadi, L. Evidence of activation of the nrf2 pathway in multiple sclerosis patients treated with delayed-release dimethyl fumarate in the phase 3 define and confirm studies. Mult. Scler. 2017, 23, 1875–1883. [Google Scholar] [CrossRef]
- Gold, R.; Kappos, L.; Arnold, D.L.; Bar-Or, A.; Giovannoni, G.; Selmaj, K.; Tornatore, C.; Sweetser, M.T.; Yang, M.; Sheikh, S.I.; et al. Placebo-controlled phase 3 study of oral bg-12 for relapsing multiple sclerosis. N. Engl. J. Med. 2012, 367, 1098–1107. [Google Scholar] [CrossRef]
- Kraaij, M.D.; Savage, N.D.; van der Kooij, S.W.; Koekkoek, K.; Wang, J.; van den Berg, J.M.; Ottenhoff, T.H.; Kuijpers, T.W.; Holmdahl, R.; van Kooten, C.; et al. Induction of regulatory t cells by macrophages is dependent on production of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2010, 107, 17686–17691. [Google Scholar] [CrossRef]
- George-Chandy, A.; Nordstrom, I.; Nygren, E.; Jonsson, I.M.; Postigo, J.; Collins, L.V.; Eriksson, K. Th17 development and autoimmune arthritis in the absence of reactive oxygen species. Eur. J. Immunol. 2008, 38, 1118–1126. [Google Scholar] [CrossRef]
- Mossberg, N.; Movitz, C.; Hellstrand, K.; Bergstrom, T.; Nilsson, S.; Andersen, O. Oxygen radical production in leukocytes and disease severity in multiple sclerosis. J. Neuroimmunol. 2009, 213, 131–134. [Google Scholar] [CrossRef]
- Mix, E.; Stefan, K.; Hoppner, J.; Klauer, T.; Zettl, U.K.; Strauss, U.; Meyer-Rienecker, H.J.; Rolfs, A. Lymphocyte subpopulations, oxidative burst and apoptosis in peripheral blood cells of patients with multiple sclerosis-effect of interferon-beta. Autoimmunity 2003, 36, 291–305. [Google Scholar] [CrossRef]
- Becanovic, K.; Jagodic, M.; Sheng, J.R.; Dahlman, I.; Aboul-Enein, F.; Wallstrom, E.; Olofsson, P.; Holmdahl, R.; Lassmann, H.; Olsson, T. Advanced intercross line mapping of eae5 reveals ncf-1 and cldn4 as candidate genes for experimental autoimmune encephalomyelitis. J. Immunol. 2006, 176, 6055–6064. [Google Scholar] [CrossRef]
- Linker, R.A.; Lee, D.H.; Ryan, S.; van Dam, A.M.; Conrad, R.; Bista, P.; Zeng, W.; Hronowsky, X.; Buko, A.; Chollate, S.; et al. Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the nrf2 antioxidant pathway. Brain A J. Neurol. 2011, 134, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Sheng, Q.; Zeng, H.; Li, W.; Wang, Q.; Ma, G.; Xu, X.; Qiu, M.; Zhang, W.; Shan, C. Specific genetic aberrations of parathyroid in chinese patients with tertiary hyperparathyroidism using whole-exome sequencing. Front. Endocrinol. 2023, 14, 1221060. [Google Scholar] [CrossRef] [PubMed]
- Messa, P.; Alfieri, C.M. Secondary and tertiary hyperparathyroidism. Front. Horm. Res. 2019, 51, 91–108. [Google Scholar] [PubMed]
- Alfieri, C.; Mattinzoli, D.; Messa, P. Tertiary and postrenal transplantation hyperparathyroidism. Endocrinol. Metab. Clin. North Am. 2021, 50, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Dream, S.; Chen, H.; Lindeman, B. Tertiary hyperparathyroidism: Why the delay? Ann. Surg. 2021, 273, e120–e122. [Google Scholar] [CrossRef] [PubMed]
- Rybak, L.P.; Mukherjea, D.; Jajoo, S.; Kaur, T.; Ramkumar, V. Sirna-mediated knock-down of nox3: Therapy for hearing loss? Cell. Mol. Life Sci. 2012, 69, 2429–2434. [Google Scholar] [CrossRef] [PubMed]
- Southall, M.D.; Li, T.; Gharibova, L.S.; Pei, Y.; Nicol, G.D.; Travers, J.B. Activation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytes. J. Pharmacol. Exp. Ther. 2003, 304, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Puntambekar, P.; Mukherjea, D.; Jajoo, S.; Ramkumar, V. Essential role of rac1/NADPH oxidase in nerve growth factor induction of trpv1 expression. J. Neurochem. 2005, 95, 1689–1703. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, Y.; Hamilton, T.A. Requirement for stat1 in lps-induced gene expression in macrophages. J. Leukoc. Biol. 2001, 69, 598–604. [Google Scholar] [CrossRef]
- Sugawara, I.; Yamada, H.; Mizuno, S. Stat1 knockout mice are highly susceptible to pulmonary mycobacterial infection. Tohoku J. Exp. Med. 2004, 202, 41–50. [Google Scholar] [CrossRef]
- Yoshimura, A. Signal transduction of inflammatory cytokines and tumor development. Cancer Sci. 2006, 97, 439–447. [Google Scholar] [CrossRef]
- Porta, C.; Hadj-Slimane, R.; Nejmeddine, M.; Pampin, M.; Tovey, M.G.; Espert, L.; Alvarez, S.; Chelbi-Alix, M.K. Interferons alpha and gamma induce p53-dependent and p53-independent apoptosis, respectively. Oncogene 2005, 24, 605–615. [Google Scholar] [CrossRef]
- Al Aameri, R.F.H.; Alanisi, E.M.A.; Oluwatosin, A.; Al Sallami, D.; Sheth, S.; Alberts, I.; Patel, S.; Rybak, L.P.; Ramkumar, V. Targeting cxcl1 chemokine signaling for treating cisplatin ototoxicity. Front. Immunol. 2023, 14, 1125948. [Google Scholar] [CrossRef]
- Dai, M.; Yang, Y.; Omelchenko, I.; Nuttall, A.L.; Kachelmeier, A.; Xiu, R.; Shi, X. Bone marrow cell recruitment mediated by inducible nitric oxide synthase/stromal cell-derived factor-1alpha signaling repairs the acoustically damaged cochlear blood-labyrinth barrier. Am. J. Pathol. 2010, 177, 3089–3099. [Google Scholar] [CrossRef]
- Tan, W.J.; Thorne, P.R.; Vlajkovic, S.M. Characterisation of cochlear inflammation in mice following acute and chronic noise exposure. Histochem. Cell Biol. 2016, 146, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.B.; Zuo, J. The contribution of immune infiltrates to ototoxicity and cochlear hair cell loss. Front. Cell. Neurosci. 2017, 11, 106. [Google Scholar] [CrossRef]
- Ou, H.C.; Raible, D.W.; Rubel, E.W. Cisplatin-induced hair cell loss in zebrafish (danio rerio) lateral line. Hear. Res. 2007, 233, 46–53. [Google Scholar] [CrossRef]
- Pang, J.; Xiong, H.; Zhan, T.; Cheng, G.; Jia, H.; Ye, Y.; Su, Z.; Chen, H.; Lin, H.; Lai, L.; et al. Sirtuin 1 and autophagy attenuate cisplatin-induced hair cell death in the mouse cochlea and zebrafish lateral line. Front. Cell. Neurosci. 2018, 12, 515. [Google Scholar] [CrossRef] [PubMed]
- Coffin, A.B.; Ou, H.; Owens, K.N.; Santos, F.; Simon, J.A.; Rubel, E.W.; Raible, D.W. Chemical screening for hair cell loss and protection in the zebrafish lateral line. Zebrafish 2010, 7, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Pyun, J.H.; Kang, S.U.; Hwang, H.S.; Oh, Y.T.; Kang, S.H.; Lim, Y.A.; Choo, O.S.; Kim, C.H. Epicatechin inhibits radiation-induced auditory cell death by suppression of reactive oxygen species generation. Neuroscience 2011, 199, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Herr, I.; Ucur, E.; Herzer, K.; Okouoyo, S.; Ridder, R.; Krammer, P.H.; von Knebel Doeberitz, M.; Debatin, K.M. Glucocorticoid cotreatment induces apoptosis resistance toward cancer therapy in carcinomas. Cancer Res. 2003, 63, 3112–3120. [Google Scholar] [PubMed]
- Bas, E.; Gupta, C.; Van De Water, T.R. A novel organ of corti explant model for the study of cochlear implantation trauma. Anat. Rec. 2012, 295, 1944–1956. [Google Scholar] [CrossRef]
- Dinh, C.T.; Haake, S.; Chen, S.; Hoang, K.; Nong, E.; Eshraghi, A.A.; Balkany, T.J.; Van De Water, T.R. Dexamethasone protects organ of corti explants against tumor necrosis factor-alpha-induced loss of auditory hair cells and alters the expression levels of apoptosis-related genes. Neuroscience 2008, 157, 405–413. [Google Scholar] [CrossRef]
- Calli, C.; Pinar, E.; Oncel, S.; Alper Bagriyanik, H.; Umut Sakarya, E. Recovery of hearing in cisplatin-induced ototoxicity in the guinea pig with intratympanic dexamethasone. Indian J. Otolaryngol. Head Neck Surg. 2012, 64, 46–50. [Google Scholar] [CrossRef]
- Shafik, A.G.; Elkabarity, R.H.; Thabet, M.T.; Soliman, N.B.; Kalleny, N.K. Effect of intratympanic dexamethasone administration on cisplatin-induced ototoxicity in adult guinea pigs. Auris Nasus Larynx 2013, 40, 51–60. [Google Scholar] [CrossRef]
- Murphy, D.; Daniel, S.J. Intratympanic dexamethasone to prevent cisplatin ototoxicity: A guinea pig model. Otolaryngol. Head Neck Surg. 2011, 145, 452–457. [Google Scholar] [CrossRef]
- Paksoy, M.; Ayduran, E.; Sanli, A.; Eken, M.; Aydin, S.; Oktay, Z.A. The protective effects of intratympanic dexamethasone and vitamin e on cisplatin-induced ototoxicity are demonstrated in rats. Med. Oncol. 2011, 28, 615–621. [Google Scholar] [CrossRef]
- Daldal, A.; Odabasi, O.; Serbetcioglu, B. The protective effect of intratympanic dexamethasone on cisplatin-induced ototoxicity in guinea pigs. Otolaryngol. Head Neck Surg. 2007, 137, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Hill, G.W.; Morest, D.K.; Parham, K. Cisplatin-induced ototoxicity: Effect of intratympanic dexamethasone injections. Otol. Neurotol. 2008, 29, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Marshak, T.; Steiner, M.; Kaminer, M.; Levy, L.; Shupak, A. Prevention of cisplatin-induced hearing loss by intratympanic dexamethasone: A randomized controlled study. Otolaryngol. Head Neck Surg. 2014, 150, 983–990. [Google Scholar] [CrossRef]
- Dinh, C.T.; Chen, S.; Bas, E.; Dinh, J.; Goncalves, S.; Telischi, F.; Angeli, S.; Eshraghi, A.A.; Van De Water, T. Dexamethasone protects against apoptotic cell death of cisplatin-exposed auditory hair cells in vitro. Otol. Neurotol. 2015, 36, 1566–1571. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, C.; Lee, J.N.; Park, R. Protective roles of fenofibrate against cisplatin-induced ototoxicity by the rescue of peroxisomal and mitochondrial dysfunction. Toxicol. Appl. Pharmacol. 2018, 353, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Keating, G.M.; Croom, K.F. Fenofibrate: A review of its use in primary dyslipidaemia, the metabolic syndrome and type 2 diabetes mellitus. Drugs 2007, 67, 121–153. [Google Scholar] [CrossRef]
- Park, C.; Ji, H.M.; Kim, S.J.; Kil, S.H.; Lee, J.N.; Kwak, S.; Choe, S.K.; Park, R. Fenofibrate exerts protective effects against gentamicin-induced toxicity in cochlear hair cells by activating antioxidant enzymes. Int. J. Mol. Med. 2017, 39, 960–968. [Google Scholar] [CrossRef]
- Thongnuanjan, P.; Soodvilai, S.; Chatsudthipong, V.; Soodvilai, S. Fenofibrate reduces cisplatin-induced apoptosis of renal proximal tubular cells via inhibition of jnk and p38 pathways. J. Toxicol. Sci. 2016, 41, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Daynes, R.A.; Jones, D.C. Emerging roles of ppars in inflammation and immunity. Nat. Rev. Immunol. 2002, 2, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Portilla, D.; Dai, G.; McClure, T.; Bates, L.; Kurten, R.; Megyesi, J.; Price, P.; Li, S. Alterations of pparalpha and its coactivator pgc-1 in cisplatin-induced acute renal failure. Kidney Int. 2002, 62, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Morizono, T. Calcium transport mechanism in the endolymph of the chinchilla. Hear. Res. 1988, 34, 307–311. [Google Scholar] [CrossRef]
- Fridberger, A.; Flock, A.; Ulfendahl, M.; Flock, B. Acoustic overstimulation increases outer hair cell Ca2+ concentrations and causes dynamic contractions of the hearing organ. Proc. Natl. Acad. Sci. USA 1998, 95, 7127–7132. [Google Scholar] [CrossRef]
- Chan, D.K.; Rouse, S.L. Sound-induced intracellular Ca2+ dynamics in the adult hearing cochlea. PLoS ONE 2016, 11, e0167850. [Google Scholar] [CrossRef]
- Li, W.; Zhao, L.; Jiang, S.; Gu, R. Effects of high intensity impulse noise on ionic concentrations in cochlear endolymph of the guinea pig. Chin. Med. J. 1997, 110, 883–886. [Google Scholar]
- Hirose, K.; Discolo, C.M.; Keasler, J.R.; Ransohoff, R. Mononuclear phagocytes migrate into the murine cochlea after acoustic trauma. J. Comp. Neurol. 2005, 489, 180–194. [Google Scholar] [CrossRef]
- So, H.; Kim, H.; Lee, J.H.; Park, C.; Kim, Y.; Kim, E.; Kim, J.K.; Yun, K.J.; Lee, K.M.; Lee, H.Y.; et al. Cisplatin cytotoxicity of auditory cells requires secretions of proinflammatory cytokines via activation of erk and nf-kappab. J. Assoc. Res. Otolaryngol. JARO 2007, 8, 338–355. [Google Scholar] [CrossRef]
- Bas, E.; Martinez-Soriano, F.; Lainez, J.M.; Marco, J. An experimental comparative study of dexamethasone, melatonin and tacrolimus in noise-induced hearing loss. Acta Oto-Laryngol. 2009, 129, 385–389. [Google Scholar] [CrossRef]
- Arslan, H.H.; Satar, B.; Serdar, M.; Yilmaz, E. Changes in proinflammatory cytokines in the cochlea in relation to hearing thresholds in noise-exposed rats. J. Int. Adv. Otol. 2017, 13, 308–312. [Google Scholar] [CrossRef]
- Infante, E.B.; Channer, G.A.; Telischi, F.F.; Gupta, C.; Dinh, J.T.; Vu, L.; Eshraghi, A.A.; Van De Water, T.R. Mannitol protects hair cells against tumor necrosis factor alpha-induced loss. Otol. Neurotol. 2012, 33, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zheng, J. Understand spiciness: Mechanism of trpv1 channel activation by capsaicin. Protein Cell 2017, 8, 169–177. [Google Scholar] [CrossRef]
- Mohler, K.M.; Torrance, D.S.; Smith, C.A.; Goodwin, R.G.; Stremler, K.E.; Fung, V.P.; Madani, H.; Widmer, M.B. Soluble tumor necrosis factor (tnf) receptors are effective therapeutic agents in lethal endotoxemia and function simultaneously as both tnf carriers and tnf antagonists. J. Immunol. 1993, 151, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Oishi, N.; Chen, F.Q.; Zheng, H.W.; Sha, S.H. Intra-tympanic delivery of short interfering rna into the adult mouse cochlea. Hear. Res. 2013, 296, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: Microrna biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.S.; Hutvagner, G.; Du, T.; Xu, Z.; Aronin, N.; Zamore, P.D. Asymmetry in the assembly of the rnai enzyme complex. Cell 2003, 115, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Chandradoss, S.D.; Schirle, N.T.; Szczepaniak, M.; MacRae, I.J.; Joo, C. A dynamic search process underlies microrna targeting. Cell 2015, 162, 96–107. [Google Scholar] [CrossRef]
- Han, J.; Lee, Y.; Yeom, K.H.; Nam, J.W.; Heo, I.; Rhee, J.K.; Sohn, S.Y.; Cho, Y.; Zhang, B.T.; Kim, V.N. Molecular basis for the recognition of primary micrornas by the drosha-dgcr8 complex. Cell 2006, 125, 887–901. [Google Scholar] [CrossRef]
- Sullenger, B.A.; Nair, S. From the rna world to the clinic. Science 2016, 352, 1417–1420. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic mirna and sirna: Moving from bench to clinic as next generation medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef]
- Aiuti, A.; Biasco, L.; Scaramuzza, S.; Ferrua, F.; Cicalese, M.P.; Baricordi, C.; Dionisio, F.; Calabria, A.; Giannelli, S.; Castiello, M.C.; et al. Lentiviral hematopoietic stem cell gene therapy in patients with wiskott-aldrich syndrome. Science 2013, 341, 1233151. [Google Scholar] [CrossRef]
- Musacchio, T.; Torchilin, V.P. Sirna delivery: From basics to therapeutic applications. Front. Biosci. 2013, 18, 58–79. [Google Scholar]
- Boudreau, R.L.; Monteys, A.M.; Davidson, B.L. Minimizing variables among hairpin-based rnai vectors reveals the potency of shrnas. RNA 2008, 14, 1834–1844. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, R.L.; Martins, I.; Davidson, B.L. Artificial micrornas as sirna shuttles: Improved safety as compared to shrnas in vitro and in vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2009, 17, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Glover, S.J.; Burgess, P.I.; Cohen, D.B.; Harding, S.P.; Hofland, H.W.; Zijlstra, E.E.; Allain, T.J. Prevalence of diabetic retinopathy, cataract and visual impairment in patients with diabetes in sub-saharan africa. Br. J. Ophthalmol. 2012, 96, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Min, H.; Kim, S.J.; Oh, S.; Kim, K.; Yu, H.G.; Park, T.; Kim, Y. Development of diagnostic biomarkers for detecting diabetic retinopathy at early stages using quantitative proteomics. J. Diabetes Res. 2016, 2016, 6571976. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, T.; Mackey, G.; Lusk, J.; Kyle, U.G.; Fontenot, T.; Marshall, P.; Shekerdemian, L.S.; Coss-Bu, J.A.; Nishigaki, A.; Yatabe, T.; et al. Esicm lives 2016: Part three. Intensive Care Med. Exp. 2016, 4, 28. [Google Scholar] [CrossRef]
- Liu, N.; Zhao, N.; Chen, L.; Cai, N. Survivin contributes to the progression of diabetic retinopathy through hif-1alpha pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 9161–9167. [Google Scholar]
- Dorrell, M.; Uusitalo-Jarvinen, H.; Aguilar, E.; Friedlander, M. Ocular neovascularization: Basic mechanisms and therapeutic advances. Surv. Ophthalmol. 2007, 52 (Suppl. S1), S3–S19. [Google Scholar] [CrossRef]
- Wan, T.T.; Li, X.F.; Sun, Y.M.; Li, Y.B.; Su, Y. Recent advances in understanding the biochemical and molecular mechanism of diabetic retinopathy. Biomed. Pharmacother. 2015, 74, 145–147. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chuang, L.M. The role of oxidative stress in the pathogenesis of type 2 diabetes: From molecular mechanism to clinical implication. Am. J. Transl. Res. 2010, 2, 316–331. [Google Scholar]
- Naudi, A.; Jove, M.; Ayala, V.; Cassanye, A.; Serrano, J.; Gonzalo, H.; Boada, J.; Prat, J.; Portero-Otin, M.; Pamplona, R. Cellular dysfunction in diabetes as maladaptive response to mitochondrial oxidative stress. Exp. Diabetes Res. 2012, 2012, 696215. [Google Scholar] [CrossRef] [PubMed]
- Malendowicz, L.K.; Macchi, C.; Nussdorfer, G.G.; Nowak, K.W.; Zyterska, A.; Ziolkowska, A. Effects of prolonged exendin-4 administration on entero-insular axis of normal and streptozotocin-induced diabetic rats. Int. J. Mol. Med. 2003, 11, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Kleine, N.; Orskov, C.; Holst, J.J.; Willms, B.; Creutzfeldt, W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7–36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia 1993, 36, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Nystrom, T. The potential beneficial role of glucagon-like peptide-1 in endothelial dysfunction and heart failure associated with insulin resistance. Horm. Metab. Res. 2008, 40, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Oeseburg, H.; de Boer, R.A.; Buikema, H.; van der Harst, P.; van Gilst, W.H.; Sillje, H.H. Glucagon-like peptide 1 prevents reactive oxygen species-induced endothelial cell senescence through the activation of protein kinase a. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- You, P.; Wu, H.; Deng, M.; Peng, J.; Li, F.; Yang, Y. Brevilin a induces apoptosis and autophagy of colon adenocarcinoma cell ct26 via mitochondrial pathway and pi3k/akt/mtor inactivation. Biomed. Pharmacother. 2018, 98, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Du, Y.; Nan, J.; Zhang, X.; Qin, X.; Wang, Y.; Hou, J.; Wang, Q.; Yang, J. Brevilin a, a novel natural product, inhibits janus kinase activity and blocks stat3 signaling in cancer cells. PLoS ONE 2013, 8, e63697. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, M.; Cui, X.; Lv, D.; Jin, L.; Khan, M.; Ma, T. Brevilin a promotes oxidative stress and induces mitochondrial apoptosis in u87 glioblastoma cells. OncoTargets Ther. 2018, 11, 7031–7040. [Google Scholar] [CrossRef]
- Choi, J.H.; Oh, J.; Lee, M.J.; Bae, H.; Ko, S.G.; Nah, S.Y.; Cho, I.H. Inhibition of lysophosphatidic acid receptor 1–3 deteriorates experimental autoimmune encephalomyelitis by inducing oxidative stress. J. Neuroinflamm. 2021, 18, 240. [Google Scholar] [CrossRef]
- Zarzuelo Romero, M.J.; Perez Ramirez, C.; Carrasco Campos, M.I.; Sanchez Martin, A.; Calleja Hernandez, M.A.; Ramirez Tortosa, M.C.; Jimenez Morales, A. Therapeutic value of single nucleotide polymorphisms on the efficacy of new therapies in patients with multiple sclerosis. J. Pers. Med. 2021, 11, 335. [Google Scholar] [CrossRef]
- Adamczyk, B.; Adamczyk-Sowa, M. New insights into the role of oxidative stress mechanisms in the pathophysiology and treatment of multiple sclerosis. Oxidative Med. Cell. Longev. 2016, 2016, 1973834. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.W.; Wang, J.; Zhang, Q.; Wang, R.; Dhandapani, K.M.; Vadlamudi, R.K.; Brann, D.W. NADPH oxidase in brain injury and neurodegenerative disorders. Mol. Neurodegener. 2017, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Han, J.; Song, S.Y.; Kim, W.S.; Shin, S.; Kim, J.H.; Ahn, H.; Jeong, J.H.; Hwang, S.J.; Sung, J.H. Lysophosphatidic acid increases the proliferation and migration of adipose-derived stem cells via the generation of reactive oxygen species. Mol. Med. Rep. 2015, 12, 5203–5210. [Google Scholar] [CrossRef]
- Lin, C.C.; Lin, C.E.; Lin, Y.C.; Ju, T.K.; Huang, Y.L.; Lee, M.S.; Chen, J.H.; Lee, H. Lysophosphatidic acid induces reactive oxygen species generation by activating protein kinase c in pc-3 human prostate cancer cells. Biochem. Biophys. Res. Commun. 2013, 440, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Saunders, J.A.; Rogers, L.C.; Klomsiri, C.; Poole, L.B.; Daniel, L.W. Reactive oxygen species mediate lysophosphatidic acid induced signaling in ovarian cancer cells. Free Radic. Biol. Med. 2010, 49, 2058–2067. [Google Scholar] [CrossRef] [PubMed]
- Yung, Y.C.; Stoddard, N.C.; Mirendil, H.; Chun, J. Lysophosphatidic acid signaling in the nervous system. Neuron 2015, 85, 669–682. [Google Scholar] [CrossRef]
- Yung, Y.C.; Stoddard, N.C.; Chun, J. Lpa receptor signaling: Pharmacology, physiology, and pathophysiology. J. Lipid Res. 2014, 55, 1192–1214. [Google Scholar] [CrossRef]
- Geraldo, L.H.M.; Spohr, T.; Amaral, R.F.D.; Fonseca, A.; Garcia, C.; Mendes, F.A.; Freitas, C.; dos Santos, M.F.; Lima, F.R.S. Role of lysophosphatidic acid and its receptors in health and disease: Novel therapeutic strategies. Signal Transduct. Target. Ther. 2021, 6, 45. [Google Scholar] [CrossRef]
- Schmitz, K.; Brunkhorst, R.; de Bruin, N.; Mayer, C.A.; Haussler, A.; Ferreiros, N.; Schiffmann, S.; Parnham, M.J.; Tunaru, S.; Chun, J.; et al. Dysregulation of lysophosphatidic acids in multiple sclerosis and autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2017, 5, 42. [Google Scholar] [CrossRef]
- Dittel, B.N. Cd4 t cells: Balancing the coming and going of autoimmune-mediated inflammation in the cns. Brain Behav. Immun. 2008, 22, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, G.G.; Pacheco-Moises, F.P.; Macias-Islas, M.A.; Flores-Alvarado, L.J.; Mireles-Ramirez, M.A.; Gonzalez-Renovato, E.D.; Hernandez-Navarro, V.E.; Sanchez-Lopez, A.L.; Alatorre-Jimenez, M.A. Role of the blood-brain barrier in multiple sclerosis. Arch. Med. Res. 2014, 45, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Breitzig, M.; Bhimineni, C.; Lockey, R.; Kolliputi, N. 4-hydroxy-2-nonenal: A critical target in oxidative stress? Am. J. Physiol. Cell Physiol. 2016, 311, C537–C543. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The enzyme family of NADPH oxidases (Nox) consists of seven members, namely Nox1, Nox2, Nox3, Nox4, Nox5 and the Dual oxidases Duox1 and Duox2. The core structure consists of six α-helical domains. At the C-terminal end of the core, the structure binding regions for nicotinamide adenine dinucleotide phosphate (NADPH) and flavin adenine dinucleotide (FAD) are located, which mediate electron delivery and translocation. The isoforms strongly vary in terms of activation and utilization of adaptor subunits necessary for ROS production. Nox1 recruits the organizer subunit NADPH oxidase organizer 1 (NOXO1) and the activator subunit NADPH oxidase activator 1 (NOXA1). Nox2 needs p47phox as organizer and p67phox as activator subunit, which both are tethered together via the scaffold-like protein p40phox. Nox1 and Nox2 strictly also need the guanosine triphosphate phosphohydrolase (GTPase) Ras-related C3 botulinum toxin substrate (Rac) for full activity. Without these adaptor subunits, both Nox isoforms are quiescent. Nox4, on the contrary, is constitutively active, but the ROS production can be regulated either via changes in Nox4 protein expression or via regulatory factor polymerase (DNA-directed) delta interacting protein 2 (Poldip2). The structural membrane-bound subunit p22phox is crucial for enzymatic activity and maturation of Nox1, Nox2 and Nox4. Nox5, Duox1 and Duox2 do not need p22phox or any activating subunit but are both activated by Ca2+-binding to their cytosolic EF-hand domains. The stabilizing factors Dual Oxidase Maturation Factor 1/2 (DUOXA1/2) are crucial for the maturation and transportation of Duox1/2 but not for activation of the enzymes. Duox1 and Duox2 also contain a peroxidase-like domain on the extracellular side, which utilizes H2O2 for oxidation.
Figure 1.
The enzyme family of NADPH oxidases (Nox) consists of seven members, namely Nox1, Nox2, Nox3, Nox4, Nox5 and the Dual oxidases Duox1 and Duox2. The core structure consists of six α-helical domains. At the C-terminal end of the core, the structure binding regions for nicotinamide adenine dinucleotide phosphate (NADPH) and flavin adenine dinucleotide (FAD) are located, which mediate electron delivery and translocation. The isoforms strongly vary in terms of activation and utilization of adaptor subunits necessary for ROS production. Nox1 recruits the organizer subunit NADPH oxidase organizer 1 (NOXO1) and the activator subunit NADPH oxidase activator 1 (NOXA1). Nox2 needs p47phox as organizer and p67phox as activator subunit, which both are tethered together via the scaffold-like protein p40phox. Nox1 and Nox2 strictly also need the guanosine triphosphate phosphohydrolase (GTPase) Ras-related C3 botulinum toxin substrate (Rac) for full activity. Without these adaptor subunits, both Nox isoforms are quiescent. Nox4, on the contrary, is constitutively active, but the ROS production can be regulated either via changes in Nox4 protein expression or via regulatory factor polymerase (DNA-directed) delta interacting protein 2 (Poldip2). The structural membrane-bound subunit p22phox is crucial for enzymatic activity and maturation of Nox1, Nox2 and Nox4. Nox5, Duox1 and Duox2 do not need p22phox or any activating subunit but are both activated by Ca2+-binding to their cytosolic EF-hand domains. The stabilizing factors Dual Oxidase Maturation Factor 1/2 (DUOXA1/2) are crucial for the maturation and transportation of Duox1/2 but not for activation of the enzymes. Duox1 and Duox2 also contain a peroxidase-like domain on the extracellular side, which utilizes H2O2 for oxidation.

Figure 2.
Nox3 is unique among the Nox isoforms since it shows a robust and constitutive ROS production without any organizer or activator subunit, similar to Nox4. Nevertheless, like Nox1, Nox2 and Nox4, Nox3 critically needs the membrane-bound subunit p22phox for enzymatic activity as well as proper protein synthesis and cellular localization. Remarkably, Nox3 shows the most flexible possibilities of adaptor subunit usage among all Nox isoforms. Nox3 can utilize the adaptor subunits of both Nox1 and Nox2 in any thinkable combination. In human cells, the combination of adaptor subunits strongly affects the ROS production of Nox3. The subunits of Nox2, namely p67phox or p47phox, either not or only weakly enhance Nox3-derived ROS production, respectively. The same applies for the activator subunit of Nox1 NOXA1. While Rac is crucial for Nox1 and Nox2 activation, it is not needed for Nox3 activation per se, but, in combination with p67phox, it can enhance Nox3-derived ROS production. The Nox1 organizer subunit NOXO1 induces the strongest ROS production, which can be initiated by a single adaptor subunit together with Nox3, while the combination of either the Nox2 or the Nox1 adaptor subunits together lead to maximal ROS output by Nox3.
Figure 2.
Nox3 is unique among the Nox isoforms since it shows a robust and constitutive ROS production without any organizer or activator subunit, similar to Nox4. Nevertheless, like Nox1, Nox2 and Nox4, Nox3 critically needs the membrane-bound subunit p22phox for enzymatic activity as well as proper protein synthesis and cellular localization. Remarkably, Nox3 shows the most flexible possibilities of adaptor subunit usage among all Nox isoforms. Nox3 can utilize the adaptor subunits of both Nox1 and Nox2 in any thinkable combination. In human cells, the combination of adaptor subunits strongly affects the ROS production of Nox3. The subunits of Nox2, namely p67phox or p47phox, either not or only weakly enhance Nox3-derived ROS production, respectively. The same applies for the activator subunit of Nox1 NOXA1. While Rac is crucial for Nox1 and Nox2 activation, it is not needed for Nox3 activation per se, but, in combination with p67phox, it can enhance Nox3-derived ROS production. The Nox1 organizer subunit NOXO1 induces the strongest ROS production, which can be initiated by a single adaptor subunit together with Nox3, while the combination of either the Nox2 or the Nox1 adaptor subunits together lead to maximal ROS output by Nox3.

Figure 3.
So far, only a few studies described detailed pathways, which lead to Nox3 activation and subsequent ROS production. (A) Li et al. demonstrated that Nox3 shows basal ROS production in unstimulated HepG2 cells. (B) Nox3-derived ROS production can be enhanced via tumor necrosis factor (TNF) by two different mechanisms. (1) TNF- and protein kinase C (PKC)-mediated signaling results in the translocation of p47phox to the Nox3 core complex and subsequently activates Nox3-derived ROS production directly and (2) as many other exogenous or endogenous factors; also, TNF signaling leads to the up-regulation of Nox3 mRNA expression, protein synthesis and finally the increase of ROS production [396]. (C) Similar to TNF, other endogenous factors, such as growth factors or hormones, as well as exogenous factors like carbon monoxide, cisplatin or noise, lead to the up-regulation of Nox3 mRNA expression and subsequent increase of the Nox3 protein, which is often correlated to an increased ROS production.
Figure 3.
So far, only a few studies described detailed pathways, which lead to Nox3 activation and subsequent ROS production. (A) Li et al. demonstrated that Nox3 shows basal ROS production in unstimulated HepG2 cells. (B) Nox3-derived ROS production can be enhanced via tumor necrosis factor (TNF) by two different mechanisms. (1) TNF- and protein kinase C (PKC)-mediated signaling results in the translocation of p47phox to the Nox3 core complex and subsequently activates Nox3-derived ROS production directly and (2) as many other exogenous or endogenous factors; also, TNF signaling leads to the up-regulation of Nox3 mRNA expression, protein synthesis and finally the increase of ROS production [396]. (C) Similar to TNF, other endogenous factors, such as growth factors or hormones, as well as exogenous factors like carbon monoxide, cisplatin or noise, lead to the up-regulation of Nox3 mRNA expression and subsequent increase of the Nox3 protein, which is often correlated to an increased ROS production.

Figure 4.
Nox3-derived ROS fulfill various important functions in the organism. (A) Insulin treatment of HepG2 cells leads to Nox3-derived ROS production, which, in turn, induce a extracellular signal-regulated kinase 1/2 (ERK1/2)-mediated translocation of the transcription factor Specific protein 1 (Sp1). Sp1 binds to the promoter of the VGEFA gene and induces expression of Vascular Endothelial Growth Factor (VGEF)-A mRNA and VGEF-A protein production. Neither the exact steps of the insulin-induced signaling cascade, which activates Nox3, nor the exact targets of Nox3-derived ROS that activate the ERK1/2 pathway are known in detail [487]. (B) Treatment of HepG2 cells with palmitate induces (1) Nox3 mRNA expression and protein synthesis by an unknown mechanism. The increased basal Nox3-derived ROS production then (2) activates a signaling cascade, which involves phosphorylation of the mitogen-activated protein kinases (MAPK) C-Jun-N-terminal Kinase 1/2 (JNK1/2), p38, phosphoinositide 3-kinases (PI3K) and the protein kinase B, also known as Akt, which ultimately leads to gluconeogenesis [537]. (C) In the inner ear, the vestibular system is responsible for detection of acceleration and gravity sensing. Three semicircular canals detect and rotational acceleration. In the vestibule, consisting of the saccule and the utricle, changes in gravity and linear acceleration are detected. In the vestibule, the maculae are responsible for this function. A gelatinous matrix is located on top of the maculae. Otoconia, solid crystalline structures, are formed in this matrix directly above the sensory hair cells. Movement of otoconia in this matrix stimulates the hair cells, which transfer the sensory information to the ganglion cells. The main inorganic compound in otoconia is calcium carbonate (CaCO3) but otoconia are not completely inorganic. Various proteins are necessary for proper otoconial formation. The major protein component is otoconin 90/95 (OC-90/95) [429,430,431], which is produced by the non-sensory epithelial cells. OC-90/95 is crucial for proper formation of the inorganic CaCO3 crystallites. The most current mechanism describes Nox3-derived ROS as crucial mediators of disulfide linkage and subsequent multimerization of OC-90. The OC-90 multimers then function as nucleation points for calcium ions (Ca2+) and CO32− to form CaCO3. Without Nox3-derived ROS, no OC-90 multimers are present as nucleation points, Ca2+ and CO32− remain in solution and otoconia are not formed [464,466]. The lack of otoconia leads to the most obvious phenotype of Nox3-deficient mice, i.e., strong balancing deficits [370].
Figure 4.
Nox3-derived ROS fulfill various important functions in the organism. (A) Insulin treatment of HepG2 cells leads to Nox3-derived ROS production, which, in turn, induce a extracellular signal-regulated kinase 1/2 (ERK1/2)-mediated translocation of the transcription factor Specific protein 1 (Sp1). Sp1 binds to the promoter of the VGEFA gene and induces expression of Vascular Endothelial Growth Factor (VGEF)-A mRNA and VGEF-A protein production. Neither the exact steps of the insulin-induced signaling cascade, which activates Nox3, nor the exact targets of Nox3-derived ROS that activate the ERK1/2 pathway are known in detail [487]. (B) Treatment of HepG2 cells with palmitate induces (1) Nox3 mRNA expression and protein synthesis by an unknown mechanism. The increased basal Nox3-derived ROS production then (2) activates a signaling cascade, which involves phosphorylation of the mitogen-activated protein kinases (MAPK) C-Jun-N-terminal Kinase 1/2 (JNK1/2), p38, phosphoinositide 3-kinases (PI3K) and the protein kinase B, also known as Akt, which ultimately leads to gluconeogenesis [537]. (C) In the inner ear, the vestibular system is responsible for detection of acceleration and gravity sensing. Three semicircular canals detect and rotational acceleration. In the vestibule, consisting of the saccule and the utricle, changes in gravity and linear acceleration are detected. In the vestibule, the maculae are responsible for this function. A gelatinous matrix is located on top of the maculae. Otoconia, solid crystalline structures, are formed in this matrix directly above the sensory hair cells. Movement of otoconia in this matrix stimulates the hair cells, which transfer the sensory information to the ganglion cells. The main inorganic compound in otoconia is calcium carbonate (CaCO3) but otoconia are not completely inorganic. Various proteins are necessary for proper otoconial formation. The major protein component is otoconin 90/95 (OC-90/95) [429,430,431], which is produced by the non-sensory epithelial cells. OC-90/95 is crucial for proper formation of the inorganic CaCO3 crystallites. The most current mechanism describes Nox3-derived ROS as crucial mediators of disulfide linkage and subsequent multimerization of OC-90. The OC-90 multimers then function as nucleation points for calcium ions (Ca2+) and CO32− to form CaCO3. Without Nox3-derived ROS, no OC-90 multimers are present as nucleation points, Ca2+ and CO32− remain in solution and otoconia are not formed [464,466]. The lack of otoconia leads to the most obvious phenotype of Nox3-deficient mice, i.e., strong balancing deficits [370].

Figure 5.
Overproduction of ROS or ROS production in the wrong location can lead to oxidative distress, cellular damage, malfunctioning of tissues and organs and finally manifest in diseases. (A) The cochlea is the organ responsible for hearing and in contrast to the vestibular system, loss of Nox3 leads to a rather protective outcome for the tissue and the hearing capacity. In the cochlea, the organ of Corti is responsible for detection of sound waves and neuronal processing. For that, outer and inner hair cells detect movements of the tectorial membrane, which are induced by incoming sound waves. Under healthy conditions, hair cells and supporting cells function normally; however, under exogenous or endogenous stress conditions, supporting cells up-regulate Nox3. The subsequent ROS overproduction leads to hair cell death and contributes to age-, noise- and drug-induced hearing loss [462,476,478]. (B) In WT mice, the development of lung emphysemas with increasing age is inhibited by a complicated signaling cascade in lung endothelial cells [395,483]. (1) Heat shock protein 70 (Hsp70) activates Toll-like receptor 4 (TLR4)-mediated signaling, which finally leads to activation and translocation of the transcription factor Interferon regulatory factor 3 (IFR3) into the nucleus. IFR3 induces the expression and production of Interferon-β (IFN-β), which is subsequently secreted and (2) activates the lung cells via binding to the IFN receptor in an autocrine manner. The IFN receptor-induced signaling cascade results in activation and translocation of the transcription factor Signal transducer and activator of transcription 3 (STAT3), which then binds to the promoter of the NOX3 gene in result inhibiting the expression of Nox3. (C) In TLR4-deficient animals, this autocrine signaling cascade does not activate, which leads to increased mRNA expression and synthesis of Nox3 and subsequently to an increased ROS production of lung endothelial cells. The accumulating oxidative damage results in destruction of the alveolar structures and subsequently to the development of lung emphysemas in TLR4-deficient mice observed with increasing age.
Figure 5.
Overproduction of ROS or ROS production in the wrong location can lead to oxidative distress, cellular damage, malfunctioning of tissues and organs and finally manifest in diseases. (A) The cochlea is the organ responsible for hearing and in contrast to the vestibular system, loss of Nox3 leads to a rather protective outcome for the tissue and the hearing capacity. In the cochlea, the organ of Corti is responsible for detection of sound waves and neuronal processing. For that, outer and inner hair cells detect movements of the tectorial membrane, which are induced by incoming sound waves. Under healthy conditions, hair cells and supporting cells function normally; however, under exogenous or endogenous stress conditions, supporting cells up-regulate Nox3. The subsequent ROS overproduction leads to hair cell death and contributes to age-, noise- and drug-induced hearing loss [462,476,478]. (B) In WT mice, the development of lung emphysemas with increasing age is inhibited by a complicated signaling cascade in lung endothelial cells [395,483]. (1) Heat shock protein 70 (Hsp70) activates Toll-like receptor 4 (TLR4)-mediated signaling, which finally leads to activation and translocation of the transcription factor Interferon regulatory factor 3 (IFR3) into the nucleus. IFR3 induces the expression and production of Interferon-β (IFN-β), which is subsequently secreted and (2) activates the lung cells via binding to the IFN receptor in an autocrine manner. The IFN receptor-induced signaling cascade results in activation and translocation of the transcription factor Signal transducer and activator of transcription 3 (STAT3), which then binds to the promoter of the NOX3 gene in result inhibiting the expression of Nox3. (C) In TLR4-deficient animals, this autocrine signaling cascade does not activate, which leads to increased mRNA expression and synthesis of Nox3 and subsequently to an increased ROS production of lung endothelial cells. The accumulating oxidative damage results in destruction of the alveolar structures and subsequently to the development of lung emphysemas in TLR4-deficient mice observed with increasing age.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Herb, M. NADPH Oxidase 3: Beyond the Inner Ear. Antioxidants 2024, 13, 219. https://doi.org/10.3390/antiox13020219
AMA Style
Herb M. NADPH Oxidase 3: Beyond the Inner Ear. Antioxidants. 2024; 13(2):219. https://doi.org/10.3390/antiox13020219
Chicago/Turabian StyleHerb, Marc. 2024. "NADPH Oxidase 3: Beyond the Inner Ear" Antioxidants 13, no. 2: 219. https://doi.org/10.3390/antiox13020219
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.