
Exploring Influence of Production Area and Harvest Time on Specialized Metabolite Content of Glycyrrhiza glabra Leaves and Evaluation of Antioxidant and Anti-Aging Properties
 , , , , ,
, , , , , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Standards
2.2. Samples
2.3. Ultrasound-Assisted Extraction (UAE)
2.4. Quantitative Analysis
2.5. ABTS Assay
2.6. Yeast Strains and Media
2.7. Chronological Lifespan Experiments (CLS)
2.8. Analysis of Reactive Oxygen Species (ROS) Levels
2.9. Statistical Analysis
3. Results and Discussion
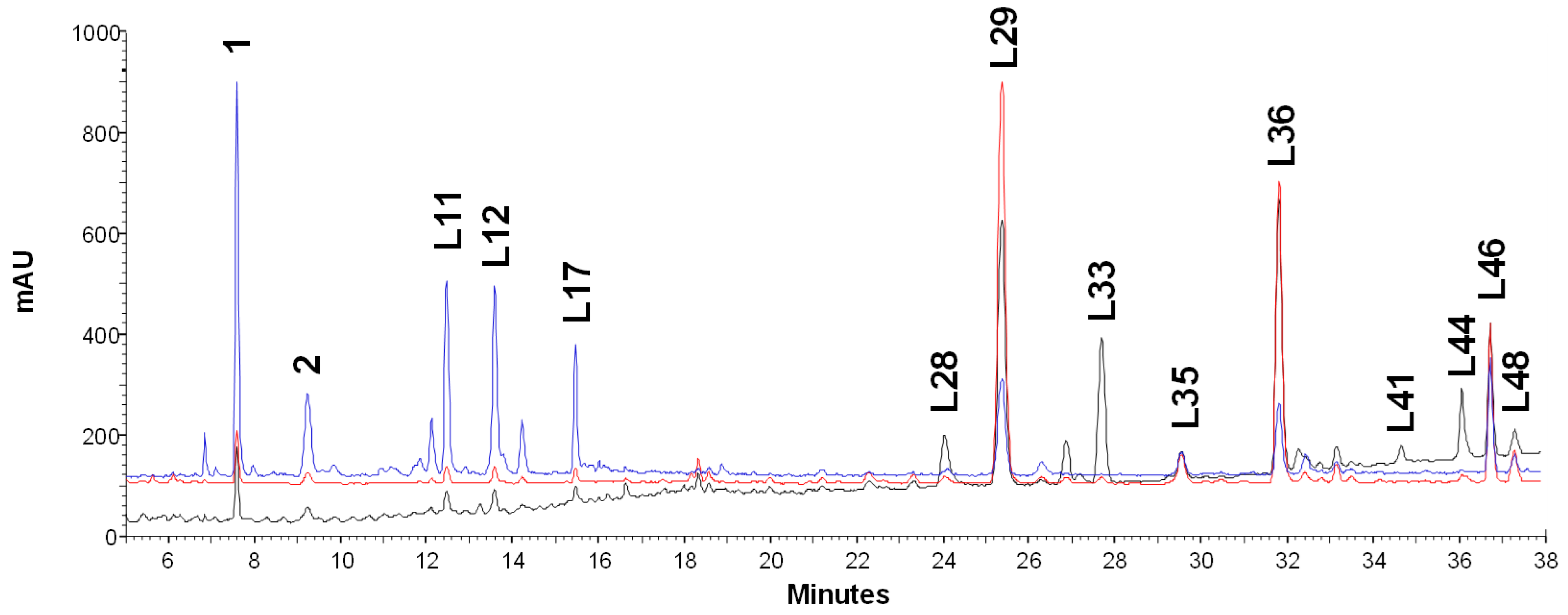
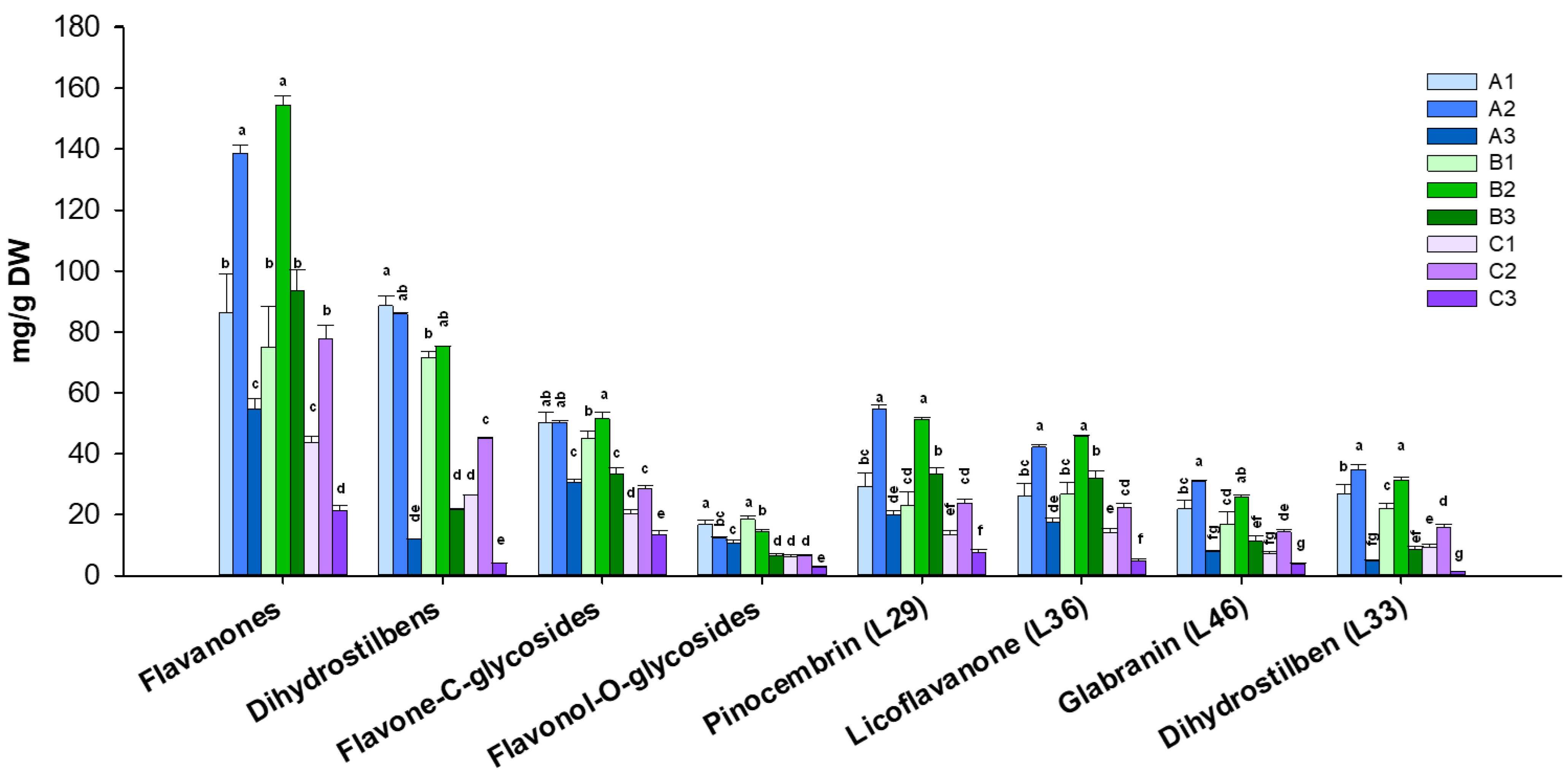
3.1. Quantitative Profile of Glycyrrhiza glabra Leaves
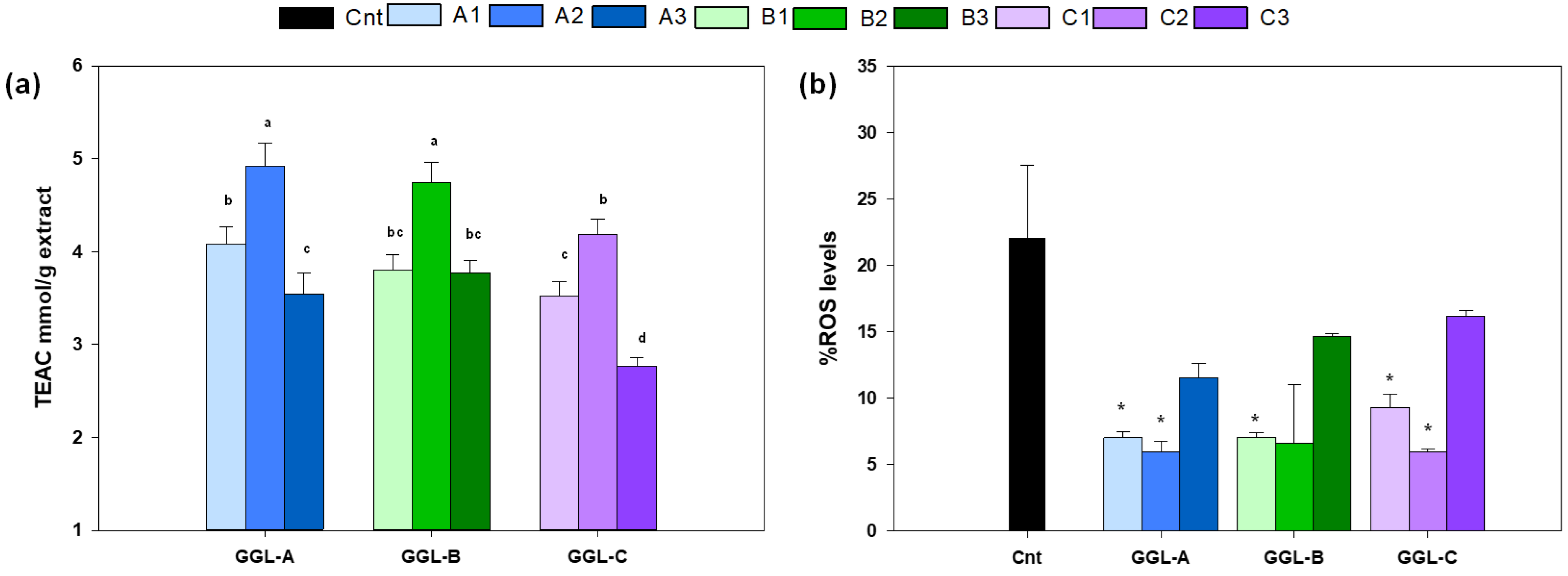
3.2. Antioxidant Activity In Vitro and In Vivo of Glycyrrhiza glabra Extracts
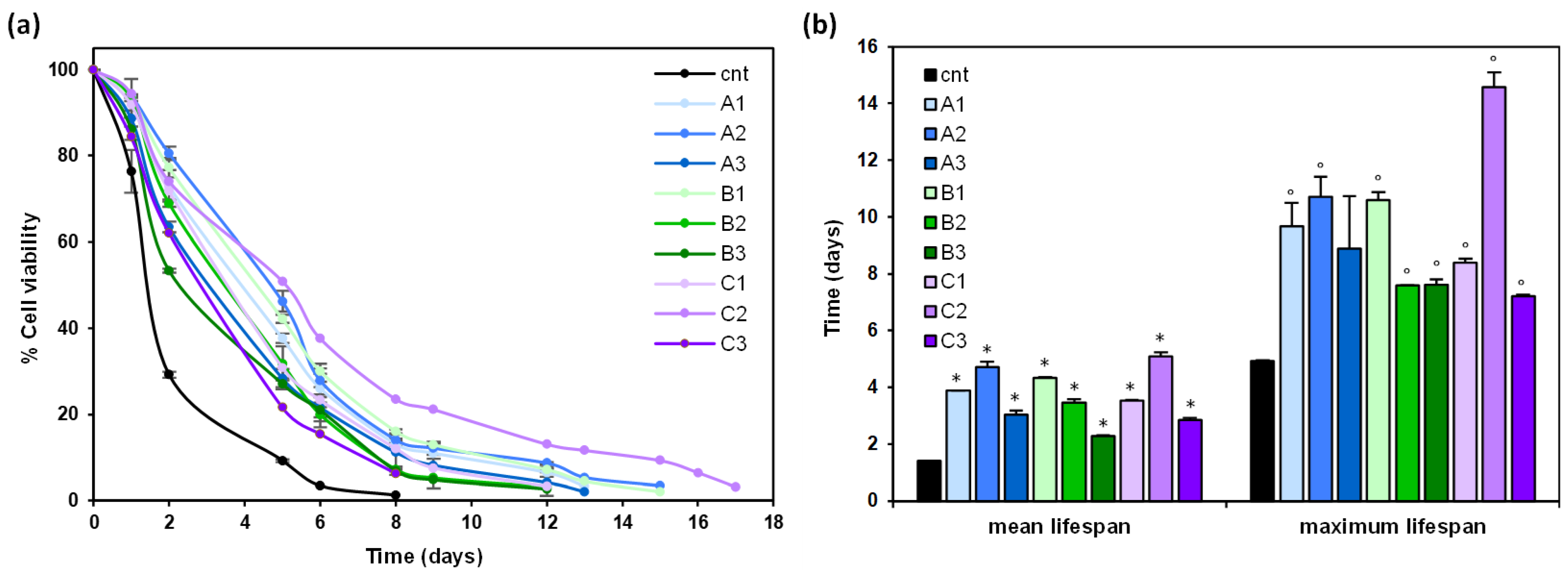
3.3. Anti-Aging Effects of Glycyrrhiza glabra Extracts in Yeast Cells Expressing Human α-Synuclein
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef]
- Karkanis, A.; Martins, N.; Petropoulos, S.; Ferreira, I.C. Phytochemical composition, health effects, and crop management of liquorice (Glycyrrhiza glabra L.): A medicinal plant. Food Rev. Int. 2018, 34, 182–203. [Google Scholar] [CrossRef]
- Ceccuzzi, G.; Rapino, A.; Perna, B.; Costanzini, A.; Farinelli, A.; Fiorica, I.; Marziani, B.; Cianci, A.; Rossin, F.; Cesaro, A.E. Liquorice Toxicity: A Comprehensive Narrative Review. Nutrients 2023, 15, 3866. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Serra, D.; Suraci, F.; Di Sanzo, R.; Fuda, S.; Postorino, S. The potential of e-nose aroma profiling for identifying the geographical origin of licorice (Glycyrrhiza glabra L.) roots. Food Chem. 2014, 165, 467–474. [Google Scholar] [CrossRef]
- Cheel, J.; Tůmová, L.; Areche, C.; Van Antwerpen, P.; Nève, J.; Zouaoui-Boudjeltia, K.; San Martin, A.; Vokřál, I.; Wsól, V.; Neugebauerová, J. Variations in the chemical profile and biological activities of licorice (Glycyrrhiza glabra L.), as influenced by harvest times. Acta Physiol. Plant. 2013, 35, 1337–1349. [Google Scholar] [CrossRef]
- Beszterda, M.; Frański, R. Seasonal Qualitative Variations of Phenolic Content in the Stem Bark of Prunus persica var. nucipersica—Implication for the Use of the Bark as a Source of Bioactive Compounds. ChemistrySelect 2022, 7, e202200418. [Google Scholar] [CrossRef]
- He, G.; Hou, X.; Han, M.; Qiu, S.; Li, Y.; Qin, S.; Chen, X. Discrimination and polyphenol compositions of green teas with seasonal variations based on UPLC-QTOF/MS combined with chemometrics. J. Food Compos. Anal. 2022, 105, 104267. [Google Scholar] [CrossRef]
- Ju, Y.; Wang, Y.; Ma, L.; Kang, L.; Liu, H.; Ma, X.; Zhao, D. Comparative Analysis of Polyphenols in Lycium barbarum Fruits Using UPLC-IM-QTOF-MS. Molecules 2023, 28, 4930. [Google Scholar] [CrossRef]
- Milião, G.L.; de Oliveira, A.P.H.; de Souza Soares, L.; Arruda, T.R.; Vieira, É.N.R.; de Castro Leite, B.R., Jr. Unconventional food plants: Nutritional aspects and perspectives for industrial applications. Future Foods 2022, 5, 100124. [Google Scholar] [CrossRef]
- Wojdyło, A.; Nowicka, P.; Turkiewicz, I.P.; Tkacz, K.; Hernandez, F. Comparison of bioactive compounds and health promoting properties of fruits and leaves of apple, pear and quince. Sci. Rep. 2021, 11, 20253. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.H.; Ye, R.; Xu, H.Y.; Feng, X.H.; Ma, C.M. Structures and in vitro antihepatic fibrosis activities of prenylated dihydrostilbenes and flavonoids from Glycyrrhiza uralensis leaves. J. Food Sci. 2019, 84, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Frattaruolo, L.; Carullo, G.; Brindisi, M.; Mazzotta, S.; Bellissimo, L.; Rago, V.; Curcio, R.; Dolce, V.; Aiello, F.; Cappello, A.R. Antioxidant and anti-inflammatory activities of flavanones from Glycyrrhiza glabra L. (licorice) leaf phytocomplexes: Identification of licoflavanone as a modulator of NF-kB/MAPK pathway. Antioxidants 2019, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, L.; Saija, A.; Cristani, M.; Cimino, F.; D’Arrigo, M.; Trombetta, D.; Rao, F.; Ruberto, G. Phytocomplexes from liquorice (Glycyrrhiza glabra L.) leaves—Chemical characterization and evaluation of their antioxidant, anti-genotoxic and anti-inflammatory activity. Fitoterapia 2011, 82, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, D.; Giofrè, S.V.; Tomaino, A.; Raciti, R.; Saija, A.; Cristani, M.; Romeo, R.; Siracusa, L.; Ruberto, G. Selective COX-2 inhibitory properties of dihydrostilbenes from liquorice leaves—In vitro assays and structure/activity relationship study. Nat. Prod. Commun. 2014, 9, 1934578X1400901226. [Google Scholar] [CrossRef]
- Ye, R.; Fan, Y.-H.; Ma, C.-M. Identification and enrichment of α-glucosidase-inhibiting dihydrostilbene and flavonoids from Glycyrrhiza uralensis leaves. J. Agric. Food Chem. 2017, 65, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Celano, R.; Docimo, T.; Piccinelli, A.L.; Rizzo, S.; Campone, L.; Di Sanzo, R.; Carabetta, S.; Rastrelli, L.; Russo, M. Specialized metabolite profiling of different Glycyrrhiza glabra organs by untargeted UHPLC-HRMS. Ind. Crops Prod. 2021, 170, 113688. [Google Scholar] [CrossRef]
- Chen, X.; Mukwaya, E.; Wong, M.-S.; Zhang, Y. A systematic review on biological activities of prenylated flavonoids. Pharm. Biol. 2014, 52, 655–660. [Google Scholar] [CrossRef]
- Chang, S.K.; Jiang, Y.; Yang, B. An update of prenylated phenolics: Food sources, chemistry and health benefits. Trends Food Sci. Technol. 2021, 108, 197–213. [Google Scholar] [CrossRef]
- Wen, L.; Zhou, T.; Jiang, Y.; Gong, L.; Yang, B. Identification of prenylated phenolics in mulberry leaf and their neuroprotective activity. Phytomedicine 2021, 90, 153641. [Google Scholar] [CrossRef]
- Jo, Y.H.; Lee, S.; Yeon, S.W.; Turk, A.; Lee, J.H.; Hong, S.-M.; Han, Y.K.; Lee, K.Y.; Hwang, B.Y.; Kim, S.Y. Anti-diabetic potential of Masclura tricuspidata leaves: Prenylated isoflavonoids with α-glucosidase inhibitory and anti-glycation activity. Bioorg. Chem. 2021, 114, 105098. [Google Scholar] [CrossRef]
- Song, Y.-H.; Cai, H.; Gu, N.; Qian, C.-F.; Cao, S.-P.; Zhao, Z.-M. Icariin attenuates cardiac remodelling through down-regulating myocardial apoptosis and matrix metalloproteinase activity in rats with congestive heart failure. J. Pharm. Pharmacol. 2011, 63, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Dodel, R.; Csoti, I.; Ebersbach, G.; Fuchs, G.; Hahne, M.; Kuhn, W.; Oechsner, M.; Jost, W.; Reichmann, H.; Schulz, J.B. Lewy body dementia and Parkinson’s disease with dementia. J. Neurol. 2008, 255, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, D.F.; Pavlou, M.A.S.; Outeiro, T.F. Cellular models as tools for the study of the role of alpha-synuclein in Parkinson’s disease. Exp. Neurol. 2017, 298, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.B.S.S.; Guedes, A.; Vasilevskiy, I.; Gonçalves, S.; Outeiro, T.F.; Winderickx, J.; Burhans, W.C.; Ludovico, P. α-Synuclein toxicity in yeast and human cells is caused by cell cycle re-entry and autophagy degradation of ribonucleotide reductase 1. Aging Cell 2019, 18, e12922. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Marques, B.; Pereira, H.; Santos, A.R.; Teixeira, A.; Ludovico, P. Caloric restriction rescues yeast cells from alpha-synuclein toxicity through autophagic control of proteostasis. Aging 2018, 10, 3821. [Google Scholar] [CrossRef] [PubMed]
- Tenreiro, S.; Franssens, V.; Winderickx, J.; Outeiro, T.F. Yeast models of Parkinson’s disease-associated molecular pathologies. Curr. Opin. Genet. Dev. 2017, 44, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, F.; Falletta, E.; Leri, M.; Angeloni, C.; Beghelli, D.; Giusti, L.; Milanesi, R.; Sampaio-Marques, B.; Ludovico, P.; Goppa, L. Anti-aging and neuroprotective properties of Grifola frondosa and Hericium erinaceus extracts. Nutrients 2022, 14, 4368. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, F.; Lombardi, L.; Guzzetti, L.; Panzeri, D.; Milanesi, R.; Leri, M.; Bucciantini, M.; Angeloni, C.; Beghelli, D.; Hrelia, S. Protective effect of Vigna unguiculata extract against aging and neurodegeneration. Aging 2020, 12, 19785. [Google Scholar] [CrossRef]
- Piccinelli, A.L.; García Mesa, M.; Armenteros, D.M.; Alfonso, M.A.; Arevalo, A.C.; Campone, L.; Rastrelli, L. HPLC-PDA-MS and NMR characterization of C-glycosyl flavones in a hydroalcoholic extract of Citrus aurantifolia leaves with antiplatelet activity. J. Agric. Food Chem. 2008, 56, 1574–1581. [Google Scholar] [CrossRef]
- Celano, R.; Piccinelli, A.L.; Pagano, I.; Roscigno, G.; Campone, L.; De Falco, E.; Russo, M.; Rastrelli, L. Oil distillation wastewaters from aromatic herbs as new natural source of antioxidant compounds. Food Res. Int. 2017, 99, 298–307. [Google Scholar] [CrossRef]
- Li, Y.; Zidorn, C. Seasonal variations of natural products in European herbs. Phytochem. Rev. 2022, 21, 1549–1575. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are flavonoids effective antioxidants in plants? Twenty years of our investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Woo, H.R.; Nam, H.G. Toward systems understanding of leaf senescence: An integrated multi-omics perspective on leaf senescence research. Mol. Plant 2016, 9, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, H.; Li, X.; Zhang, F.; Liu, C.; Du, Y.; Gao, X.; Zhang, Z.; Zhang, X.; Hou, Z. Intergrative metabolomic and transcriptomic analyses unveil nutrient remobilization events in leaf senescence of tobacco. Sci. Rep. 2017, 7, 12126. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Chen, P.; Ma, M. Feeding preference of Altica deserticola for leaves of Glycyrrhiza glabra and G. uralensis and its mechanism. Sci. Rep. 2020, 10, 1534. [Google Scholar] [CrossRef]
- Diaz Napal, G.N.; Palacios, S.M. Bioinsecticidal effect of the flavonoids pinocembrin and quercetin against Spodoptera frugiperda. J. Pest Sci. 2015, 88, 629–635. [Google Scholar] [CrossRef]
- Rasul, A.; Millimouno, F.M.; Ali Eltayb, W.; Ali, M.; Li, J.; Li, X. Pinocembrin: A novel natural compound with versatile pharmacological and biological activities. BioMed Res. Int. 2013, 2013, 379850. [Google Scholar] [CrossRef]
- Hanawa, F.; Yamada, T.; Nakashima, T. Phytoalexins from Pinus strobus bark infected with pinewood nematode, Bursaphelenchus xylophilus. Phytochemistry 2001, 57, 223–228. [Google Scholar] [CrossRef]
- Napal, G.N.D.; Carpinella, M.C.; Palacios, S.M. Antifeedant activity of ethanolic extract from Flourensia oolepis and isolation of pinocembrin as its active principle compound. Bioresource Technol. 2009, 100, 3669–3673. [Google Scholar] [CrossRef]
- Shain, L.; Miller, J.B. Pinocembrin: An antifungal compound secreted by leaf glands of eastern cottonwood. Phytopathology 1982, 72, 877–880. [Google Scholar] [CrossRef]
- Hermann, S.; Orlik, M.; Boevink, P.; Stein, E.; Scherf, A.; Kleeberg, I.; Schmitt, A.; Schikora, A. Biocontrol of plant diseases using Glycyrrhiza glabra leaf extract. Plant Dis. 2022, 106, 3133–3144. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Zhao, F.; Ye, N. Exploration of the effects of different blue LED light intensities on flavonoid and lipid metabolism in tea plants via transcriptomics and metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Sgrò, F.; Raimondo, F.M.; Ingegneri, M.; Mastracci, L.; D’Angelo, V.; Germanò, M.P.; Trombetta, D.; Smeriglio, A. Pedoclimatic Conditions Influence the Morphological, Phytochemical and Biological Features of Mentha pulegium L. Plants 2022, 12, 24. [Google Scholar] [CrossRef]
- Biondi, D.M.; Rocco, C.; Ruberto, G. New Dihydrostilbene Derivatives from the Leaves of Glycyrrhiza glabra and Evaluation of Their Antioxidant Activity. J. Nat. Prod. 2003, 66, 477–480. [Google Scholar] [CrossRef]
- Biondi, D.M.; Rocco, C.; Ruberto, G. Dihydrostilbene derivatives from Glycyrrhiza glabra leaves. J. Nat. Prod. 2005, 68, 1099–1102. [Google Scholar] [CrossRef]
- Reddy, V.P. Oxidative Stress in Health and Disease. Biomedicines 2023, 11, 2925. [Google Scholar] [CrossRef]
- Van Den Eeden, S.K.; Tanner, C.M.; Bernstein, A.L.; Fross, R.D.; Leimpeter, A.; Bloch, D.A.; Nelson, L.M. Incidence of Parkinson’s disease: Variation by age, gender, and race/ethnicity. Am. J. Epidemiol. 2003, 157, 1015–1022. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Flavanones | Dihydrostilbenes | Flavones | Flavonols | L29 | L36 | L46 | L33 | TEAC |
|---|---|---|---|---|---|---|---|---|---|
| P (2) b | 183 * | 132 * | 417 * | 294 * | 210 * | 211 * | 101 * | 216 * | 40 * |
| HT (2) b | 239 * | 318 * | 202 * | 182 * | 250 * | 171 * | 168 * | 531 * | 114 * |
| P × HT (4) b | 15 * | 21 * | 9 * | 27 * | 26 * | 13 * | 11 * | 25 * | 3.4 * |
| Compound | TEAC (mmol TE/mmol ± SD) |
|---|---|
| Vincenin 2 | 1.12 ± 0.01 |
| Rutin | 6.00 ± 0.04 |
| Isoquercitrin | 6.45 ± 0.03 |
| Pinocembrin | 3.21 ± 0.01 |
| Licoflavanone | 9.55 ± 0.04 |
| Glabranin | 4.08 ± 0.03 |
| Prenylstilbene (L33) | 5.09 ± 0.01 |
| Prenylstilbene (L44) | 2.50 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Docimo, T.; Celano, R.; Lambiase, A.; Di Sanzo, R.; Serio, S.; Santoro, V.; Coccetti, P.; Russo, M.; Rastrelli, L.; Piccinelli, A.L. Exploring Influence of Production Area and Harvest Time on Specialized Metabolite Content of Glycyrrhiza glabra Leaves and Evaluation of Antioxidant and Anti-Aging Properties. Antioxidants 2024, 13, 93. https://doi.org/10.3390/antiox13010093
Docimo T, Celano R, Lambiase A, Di Sanzo R, Serio S, Santoro V, Coccetti P, Russo M, Rastrelli L, Piccinelli AL. Exploring Influence of Production Area and Harvest Time on Specialized Metabolite Content of Glycyrrhiza glabra Leaves and Evaluation of Antioxidant and Anti-Aging Properties. Antioxidants. 2024; 13(1):93. https://doi.org/10.3390/antiox13010093
Chicago/Turabian StyleDocimo, Teresa, Rita Celano, Alessia Lambiase, Rosa Di Sanzo, Simona Serio, Valentina Santoro, Paola Coccetti, Mariateresa Russo, Luca Rastrelli, and Anna Lisa Piccinelli. 2024. "Exploring Influence of Production Area and Harvest Time on Specialized Metabolite Content of Glycyrrhiza glabra Leaves and Evaluation of Antioxidant and Anti-Aging Properties" Antioxidants 13, no. 1: 93. https://doi.org/10.3390/antiox13010093