
Combined Effect of Salicylic Acid and Proline Mitigates Drought Stress in Rice (Oryza sativa L.) through the Modulation of Physiological Attributes and Antioxidant Enzymes
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Pot Preparation and Fertilizer Application
2.3. Treatment Imposition
2.4. Assessment of Growth and Yield Parameters
2.5. Gas Exchange Characteristics
2.6. Estimation of Total Chlorophyll and Carotenoid Content
2.7. Estimation of Leaf Relative Water Content and Membrane Stability Index
2.8. Osmoprotectant Measurements
2.9. Hydrogen Peroxide and Melondealdehyde Determination
2.10. Estimation of Antioxidant Enzymatic Activity
2.11. Measurements of Nutrient Content
2.12. Statistical Analysis
3. Results
3.1. Growth and Biomass of Rice
3.2. Agro-Morphology and Yield of Rice
3.3. Gas Exchange Characteristics and Photosynthetic Pigments of Rice
3.4. Relative Water Content and Membrane Stability Index
3.5. Proline, Soluble Sugar and Starch Content
3.6. Hydrogen Peroxide and Malondialdehyde Content
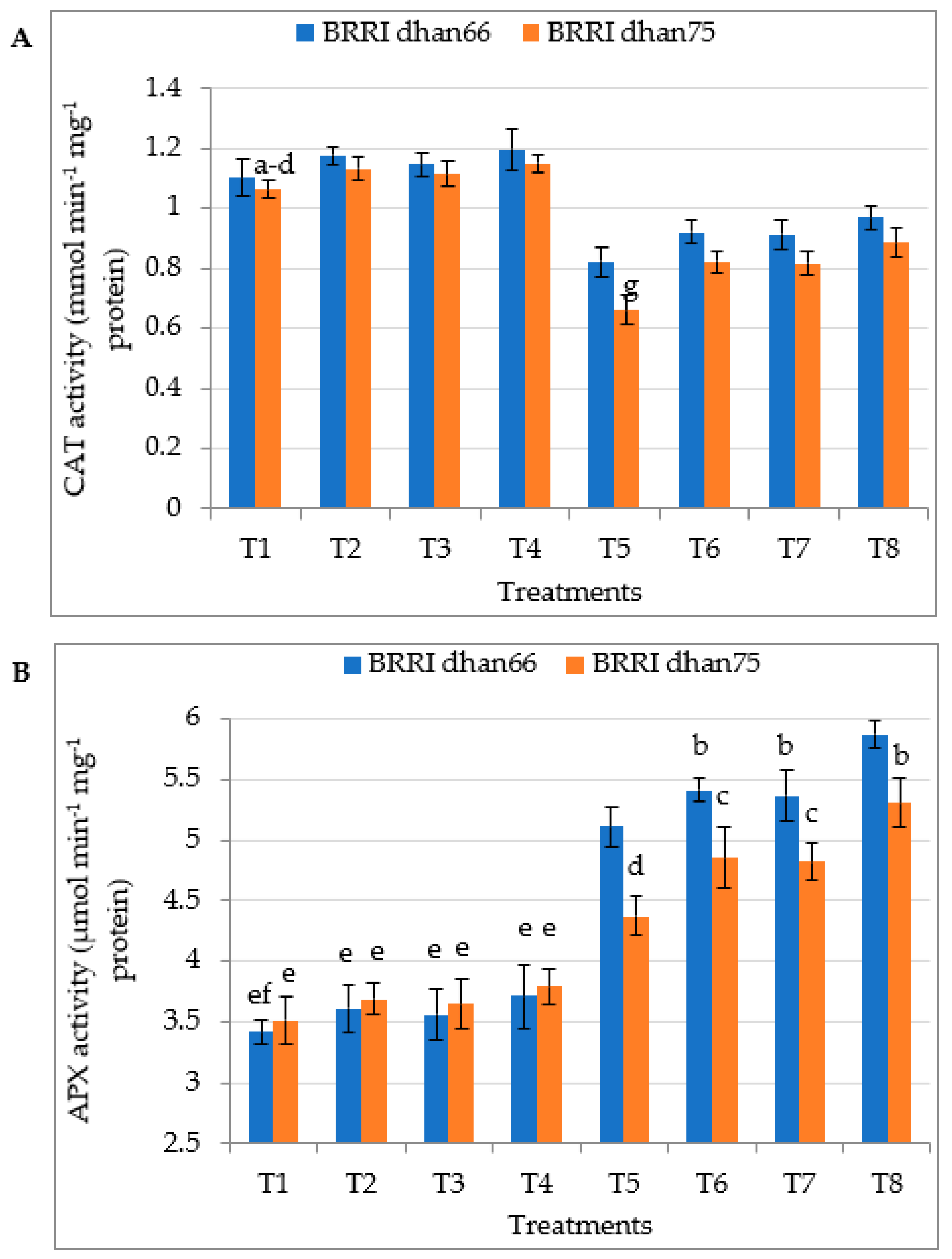
3.7. Antioxidant Enzyme Activity
3.8. Nutrients Concentrations
4. Discussion
4.1. Growth and Biomass of Rice
4.2. Agro-Morphology and Yield of Rice
4.3. Gas Exchange Characteristics and Photosynthetic Pigments of Rice
4.4. Relative Water Content and Membrane Stability Index
4.5. Proline, Soluble Sugar and Starch Content
4.6. Hydrogen Peroxide and Malondialdehyde Content
4.7. Antioxidant Enzyme Activities
4.8. Nutrients Concentrations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prasad, R.; Shivay, Y.S.; Kumar, D. Current status, challenges, and opportunities in rice production. In Rice Production Worldwide; Chauhan, B., Jabran, K., Mahajan, G., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–32. [Google Scholar]
- Molotoks, A.; Smith, P.; Dawson, T.P. Impacts of land use, population, and climate change on global food security. Food Energy Secur. 2021, 10, e261. [Google Scholar] [CrossRef]
- Islam, M.M.; Jahan, K.; Sen, A.; Urmi, T.A.; Haque, M.M.; Ali, H.M.; Siddiqui, M.H.; Murata, Y. Exogenous application of calcium ameliorates salinity stress tolerance of tomato (Solanum lycopersicum L.) and enhances fruit quality. Antioxidants 2023, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.C.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahad, S.C.; Adnan, M.; Noor, M.; Arif, M.; Alam, M.; Khan, I.A.; Ullah, H.; Wahid, F.; Mian, I.A.; Jamal, Y.; et al. Major constraints for global rice production. In Advances in Rice Research for Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Nahar, K., Biswas, J., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 1–22. [Google Scholar]
- AQUASTAT, AQUASTAT—FAO’s Global Information System on Water and Agriculture. Food and Agriculture Organization. 2019. Available online: http://www.fao.org/nr/water/aquastat/didyouknow/index3.stm (accessed on 15 March 2023).
- GIEWS, Global Information and Early Warning System. 2022. Available online: http://www.fao.org/giews/countrybrief/country.jsp?code=MYS (accessed on 22 December 2022).
- Zu, X.; Lu, Y.; Wang, Q.; Chu, P.; Miao, W.; Wang, H.; La, H. A new method for evaluating the drought tolerance of upland rice cultivars. Crop J. 2017, 5, 488–498. [Google Scholar] [CrossRef]
- Jayaweera, J.K.P.T.P.; Herath, H.M.V.G.; Jayatilake, D.V.; Udumulla, G.S.; Wickramasinghe, H.A.M. Physiological, Biochemical and Proteomic Responses of Rice (Oryza sativa L.) Varieties Godaheenati and Pokkali for Drought Stress at the Seedling Stage. Trop. Agric. Res. 2016, 27, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Larkunthod, P.; Nounjan, N.; Siangliw, J.L.; Toojinda, T.; Sanitchon, J.; Jongdee, B.; Theerakulpisut, P. Physiological Responses under Drought Stress of Improved Drought-Tolerant Rice Lines and their Parents. Not. Bot. Horti Agrobot. 2018, 46, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Sade, N.; Arzani, A.; Rubio Wilhelmi, M.D.M.; Coe, K.M.; Li, B.; Blumwald, E. Effects of abiotic stress on physiological plasticity and water use of Setaria viridis (L.). Plant Sci. 2016, 251, 128–138. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic adjustment and plant adaptation to drought stress. In Drought Stress Tolerance in Plants; Hossainh, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 1, pp. 105–143. [Google Scholar]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Tamanna, T.; Islam, M.M.; Chaity, A.R.; Shams, S.N.U.; Rasel, M.A.; Haque, M.M.; Miah, M.G.; Alamri, S.; Murata, Y. Water Relation, Gas Exchange Characteristics and Yield Performance of Selected Mungbean Genotypes under Low Soil Moisture Condition. Agronomy 2023, 13, 1068. [Google Scholar] [CrossRef]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Brestic, M.; Afrin, S.; Sakil, M.A.; Hossain, M.T.; Hossain, M.A.; Hossain, M.A. Exogenous salicylic acid and hydrogen peroxide attenuate drought stress in rice. Plant Soil Environ. 2020, 66, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Hurry, V.; Huner, N.P. Interaction of glycine betaine and plant hormones protection of the photosynthetic apparatus during abiotic stress. In Photosynthesis: Structures, Mechanisms, and Applications; Hou, H.J.M., Najafpour, M., Moore, G., Allakhverdiev, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 185–202. [Google Scholar]
- Siddiqui, M.H.; Alamri, S.; Al-Khaishany, M.Y.; Khan, M.N.; Al-Amri, A.; Ali, H.M.; Alaraidh, I.A.; Alsahli, A.A. Exogenous melatonin counteracts NaCl-induced damage by regulating the antioxidant system, proline and carbohydrates metabolism in tomato seedlings. Int. J. Mol. Sci. 2019, 20, 353. [Google Scholar] [CrossRef] [Green Version]
- Dien, D.C.; Mochizuki, T.; Yamakawa, T. Effect of various drought stresses and subsequent recovery on proline, total soluble sugar and starch metabolisms in Rice (Oryza sativa L.) varieties. Plant Prod. Sci. 2019, 22, 530–545. [Google Scholar] [CrossRef] [Green Version]
- Maruri-López, I.; Aviles-Baltazar, N.Y.; Buchala, A.; Serrano, M. Intra and extracellular journey of the phytohormone salicylic acid. Front. Plant Sci. 2019, 10, 423. [Google Scholar] [CrossRef] [Green Version]
- Khokon, M.A.R.; Salam, M.A.; Jammes, F.; Ye, W.; Hossain, M.A.; Okuma, E.; Nakamura, Y.; Mori, I.C.; Kwak, J.M.; Murata, Y. MPK9 and MPK12 function in SA-induced stomatal closure in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2017, 81, 1394–1400. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef] [Green Version]
- Movahhedi-Dehnavi, M.; Behzadi, Y.; Niknam, N.; Mohtashami, R. Salicylic acid mitigates the effects of drought and salinity on nutrient and dry matter accumulation of Linseed. J. Plant Process Funct. 2019, 8, 31–44. [Google Scholar]
- Rady, M.O.A.; Semida, W.M.; El-mageed, T.A.A.; Hemida, K.A.; Rady, M.M. Upregulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Saha, S.; Begum, H.H.; Nasrin, S. Effects of drought stress on growth and accumulation of proline in five rice varieties (Oryza sativa L.). J. Asiat. Soc. Bangladesh Sci. 2019, 45, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Kumar, V.; Burritt, D.J.; Fujita, M.; Mäkelä, P. Osmoprotectant-mediated abiotic stress tolerance in plants. In Proline Metabolism and Its Functions in Development and Stress Tolerance; Springer Nature: Cham, Switzerland, 2019; pp. 41–72. [Google Scholar]
- Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.; Rady, M.M. Natural bee-honey based biostimulants confer salt tolerance in onion via modulation of the antioxidant defence system. J. Hortic. Sci. Biotechnol. 2019, 94, 632–642. [Google Scholar] [CrossRef]
- Merwad, A.R.M.; Desoky, E.S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; Attia, K.A.; Alamery, S.F.; El-Afry, M.M.; Ghazy, A.I.; Tantawy, D.S.; Al-Doss, A.A.; El-Shawy, E.S.E.; Abu-Elsaoud, A.M.; Hafez, Y.M. Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plants associated with physiological and histological characters. Sustainability 2020, 12, 1736. [Google Scholar] [CrossRef] [Green Version]
- Patade, V.Y.; Lokhande, V.H.; Suprasanna, P. Exogenous application of proline alleviates salt induced oxidative stress more efficiently than glycine betaine in sugarcane cultured cells. Sugar Tech. 2014, 16, 22–29. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- BRRI, Modern Rice Cultivation (Adunik Dhaner Chash), 23rd ed.; Bangladesh Rice Research Institute: Gazipur, Bangladesh, 2020; pp. 1–103.
- Fadeel, A.A. Location and properties of chloroplasts and pigment determination in roots. Physiol. Plant. 1962, 15, 130–146. [Google Scholar] [CrossRef]
- Fairoj, S.A.; Islam, M.M.; Islam, M.A.; Zaman, E.; Momtaz, M.B.; Hossain, M.S.; Jahan, N.A.; Shams, S.N.U.; Urmi, T.A.; Rasel, M.A.; et al. Salicylic Acid Improves Agro-Morphology, Yield and Ion Accumulation of Two Wheat (Triticum aestivum L.) Genotypes by Ameliorating the Impact of Salt Stress. Agronomy 2022, 13, 25. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Sen, A.; Islam, M.; Zaman, E.; Ghosh, U.K.; Momtaz, M.B.; Islam, M.A.; Urmi, T.A.; Mamun, M.A.A.; Rahman, M.M.; Kamal, M.Z.U.; et al. Agro-Morphological, Yield and Biochemical Responses of Selected Wheat (Triticum aestivum L.) Genotypes to Salt Stress. Agronomy 2022, 12, 3027. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Zhang, W.; Zhang, B.; Xie, F. Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 2020, 146, 1–12. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Dhindsa, R.S.; Matowe, W. Drought Tolerance in Two Mosses: Correlated with Enzymatic Defence Against Lipid Peroxidation. J. Exp. Bot. 1981, 32, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.M.; Mulvaney, C.S. Total nitrogen. In Methods of Soil Analysis, Part 2; Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeny, D.R., Eds.; American Society of Agronomy and Soil Science Society of America, Inc.: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Jones, J.B., Jr.; Case, V.W. Sampling, handling and analyzing plant tissue samples. In Soil Testing and Plant Analysis, 3rd ed.; Westermaan, W.S., Ed.; Soil Science Society of America: Madison, WI, USA, 1990; pp. 389–427. [Google Scholar]
- Watson, M.E.; Isaac, R.A. Analytical instruments for soil and plant analysis. In Soil Testing and Plant Analysis, 3rd ed.; Westermaan, W.S., Ed.; Soil Science Society of America: Madison, WI, USA, 1990; pp. 691–740. [Google Scholar]
- Tani, E.; Chronopoulou, E.G.; Labrou, N.E.; Sarri, E.; Goufa, Μ.; Vaharidi, X.; Tornesaki, A.; Psychogiou, M.; Bebeli, P.J.; Abraham, Ε.M. Growth, physiological, biochemical, and transcriptional responses to drought stress in seedlings of Medicago sativa L., Medicago arborea L. and their hybrid (Alborea). Agronomy 2019, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Kayesh, E.; Zaman, E.; Urmi, T.A.; Haque, M.M. Evaluation of rice (Oryza sativa L.) genotypes for drought tolerance at germination and early seedling stage. Agriculturists 2018, 16, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Sokoto, M.B.; Muhammad, A. Response of rice varieties to water stress in Sokoto, Sudan Savannah, Nigeria. J. Biosci. Med. 2014, 2, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Tayyab, N.; Naz, R.; Yasmin, H.; Nosheen, A.; Keyani, R.; Sajjad, M.; Hassan, M.N.; Roberts, T.H. Combined seed and foliar pre-treatments with exogenous methyl jasmonate and salicylic acid mitigate drought-induced stress in maize. PLoS ONE 2020, 15, e0232269. [Google Scholar] [CrossRef]
- Kibria, M.G.; Farzana, K.; Matin, M.A.; Hoque, M.A. Mitigating water stress in wheat (BARI Gom-26) by exogenous application of proline. Fundam. Appl. Agric. 2016, 1, 118–123. [Google Scholar]
- Loutfy, N.; El–Tayeb, M.A.; Hassanen, A.M.; Moustafa, M.F.; Sakuma, Y.; Inouhe, M. Changes in the water status and osmotic solute contents in response to drought and salicylic acid treatments in four different cultivars of wheat (Triticum aestivum). J. Plant Res. 2012, 125, 173–184. [Google Scholar] [CrossRef]
- Shukla, N.; Awasthi, R.P.; Rawat, L.; Kumar, J. Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 2012, 54, 78–88. [Google Scholar] [CrossRef]
- Swain, P.; Raman, A.; Singh, S.P.; Kumar, A. Breeding drought tolerant rice for shallow rainfed ecosystem of eastern India. Field Crop. Res. 2017, 209, 168–178. [Google Scholar] [CrossRef]
- Singh, B.; Reddy, K.R.; Redoña, E.D.; Walker, T. Screening of rice cultivars for morpho-physiological responses to early-season soil moisture stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Islam, M.M.; Ahmed, S.; Urmi, T.A.; Raihan, M.S.; Islam, M.R. Evaluation of Moisture Regime on Agronomic Traits of Rice Genotypes. Ann. Bangladesh Agric. 2021, 25, 89–104. [Google Scholar] [CrossRef]
- Islam, M.M.; Ye, W.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, S.; Mano, J.I.; Murata, Y. Reactive carbonyl species mediate ABA signaling in guard cells. Plant Cell Physiol. 2016, 57, 2552–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.M.; Ye, W.; Matsushima, D.; Rhaman, M.S.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.S.; Mano, J.I.; Murata, Y. Reactive carbonyl species function as signal mediators downstream of H2O2 production and regulate [Ca2+]cyt elevation in ABA signal pathway in Arabidopsis guard cells. Plant Cell Physiol. 2019, 60, 1146–1159. [Google Scholar] [CrossRef]
- Du, T.; Kang, S.; Zhang, J.; Davies, W.J. Deficit irrigation and sustainable water resource strategies in agriculture for China’s food security. J. Exp. Bot. 2015, 66, 2253–2269. [Google Scholar] [CrossRef]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Talla, S.K.; Panigrahy, M.; Kappara, S.; Nirosha, P.; Neelamraju, S.; Ramanan, R. Cytokinin delays dark-induced senescence in rice by maintaining the chlorophyll cycle and photosynthetic complexes. J. Exp. Bot. 2016, 67, 1839–1851. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yu, J.; Qian, Q.; Shang, L. Enhancement of Heat and Drought Stress Tolerance in Rice by Genetic Manipulation: A Systematic Review. Rice 2022, 15, 67. [Google Scholar] [CrossRef]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.S.; Behera, P.K.; Kumar, V.; Lenka, S.K.; Panda, D. Physiological characterization and allelic diversity of selected drought tolerant traditional rice (Oryza sativa L.) landraces of Koraput, India. Physiol. Mol. Biol. Plants 2018, 24, 1035–1046. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Zhu, R.; Wu, F.Y.; Zhou, S.; Hu, T.; Huang, J.; Gao, Y. Cumulative effects of drought-flood abrupt alternation on the photosynthetic characteristics of rice. Environ. Exp. Bot. 2020, 169, 103901. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, S.; Zhang, Y.; Xu, T.; Guo, F.; Tang, H.; Li, X.; Wang, P.; Qian, W.; Xue, Y. A high temperature-dependent mitochondrial lipase EXTRA GLUME1 promotes floral phenotypic robustness against temperature fluctuation in rice (Oryza sativa L.). PLoS Genet. 2016, 12, e1006152. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Han, H.; Chen, M.; Yang, W.; Liu, L.; Li, N.; Ding, X.; Chu, Z. Overexpression of OsDT11, which encodes a novel cysteine-rich peptide, enhances drought tolerance and increases ABA concentration in rice. Plant Mol. Biol. 2017, 93, 21–34. [Google Scholar] [CrossRef]
- Croft, H.; Chen, J.M.; Luo, X.; Bartlett, P.; Chen, B.; Staebler, R.M. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Glob. Chang. Biol. 2017, 23, 3513–3524. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Mishra, S.S.; Behera, P.K.; Panda, D. Genotypic variability for drought tolerance-related morpho-physiological traits among indigenous rice landraces of Jeypore tract of Odisha, India. J. Crop Improv. 2019, 33, 254–278. [Google Scholar] [CrossRef]
- Sarwar, J.M.; Nozulaidi, B.N.M.; Khairi, B.C.L.M.; Mohd, K.Y. Effects of water stress on rice production: Bioavailability of potassium in soil. J. Stress Physiol. Biochem. 2013, 9, 97–107. [Google Scholar]
- Nahar, S.; Sahoo, L.; Tanti, B. Screening of drought tolerant rice through morpho-physiological and biochemical approaches. Biocatal. Agric. Biotechnol. 2018, 15, 150–159. [Google Scholar] [CrossRef]
- Kumar, A.; Basu, S.; Ramegowda, V.; Pereira, A. Mechanisms of drought tolerance in rice. In Achieving Sustainable Cultivation of Rice; Sasaki, T., Ed.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2016; Volume 11. [Google Scholar]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought tolerance in rice: Focus on recent mechanisms and approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef] [PubMed]
- Semida, W.M.; Abd El-Mageed, T.A.A.; Mohamed, S.E.; El-Sawah, N.A. Combined effect of deficit irrigation and foliar-applied salicylic acid on physiological responses, yield, and water-use efficiency of onion plants in saline calcareous soil. Arch. Agron. Soil Sci. 2017, 63, 1227–1239. [Google Scholar] [CrossRef]
- Gao, S.; Wang, Y.; Yu, S.; Huang, Y.; Liu, H.; Chen, W.; He, X. Effects of drought stress on growth, physiology and secondary metabolites of Two Adonis species in Northeast China. Sci. Hortic. 2020, 259, 108795. [Google Scholar] [CrossRef]
- Li, L.; Liu, Y.; Liu, Y.; He, B.; Wang, M.; Yu, C.; Weng, M. Physiological response and resistance of three cultivars of Acer rubrum L. To continuous drought stress. Acta Ecol. Sin. 2015, 35, 196–202. [Google Scholar] [CrossRef]
- Siaut, M.; Cuiné, S.; Cagnon, C.; Fessler, B.; Nguyen, M.; Carrier, P.; Beyly, A.; Beisson, F.; Triantaphylidès, C.; Li-Beisson, Y.; et al. Oil accumulation in the model green alga Chlamydomonas reinhardtii: Characterization, variability between common laboratory strains and relationship with starch reserves. BMC Biotechnol. 2011, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.R.; Islam, M.T.; Park, S.H.; Jung, H.I.; Bae, D.W.; Kim, T.H. Characterization of salicylic acid-mediated modulation of the drought stress responses: Reactive oxygen species, proline, and redox state in Brassica napus. Environ. Exp. Bot. 2019, 157, 1–10. [Google Scholar]
- Islam, M.M.; Ye, W.; Akter, F.; Rhaman, M.S.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.S.; Mano, J.I.; et al. Reactive carbonyl species mediate methyl jasmonate-induced stomatal closure. Plant Cell Physiol. 2020, 61, 1788–1797. [Google Scholar] [CrossRef]
- Melandri, G.; AbdElgawad, H.; Riewe, D.; Hageman, J.A.; Asard, H.; Beemster, G.T.; Kadam, N.; Jagadish, K.; Altmann, T.; Ruyter-Spira, C.; et al. Biomarkers for grain yield stability in rice under drought stress. J. Exp. Bot. 2020, 71, 669–683. [Google Scholar] [CrossRef] [Green Version]
- Picaud, J.C.; Steghens, J.P.; Auxenfans, C.; Barbieux, A.; Laborie, C.O. Lipid peroxidation assessment by malondialdehyde measurement in parenteral nutrition solutions for new born infants: A pilot study. Acta Paediatr. 2004, 93, 241–245. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Saxena, S.C.; Kaur, H.; Verma, P.; Petla, B.P.; Andugula, V.R.; Majee, M. Osmoprotectants: Potential for crop improvement under adverse conditions. In Plant Acclimation to Environmental Stress; Tuteja, N., Gill, S.S., Eds.; Springer: New York, NY, USA, 2013; pp. 197–232. [Google Scholar]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef]
- EL-Bauome, H.A.; Abdeldaym, E.A.; Abd El-Hady, M.A.; Darwish, D.B.E.; Alsubeie, M.S.; El-Mogy, M.M.; Basahi, M.A.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Alzuaibr, F.M.; et al. Exogenous Proline, Methionine, and Melatonin Stimulate Growth, Quality, and Drought Tolerance in Cauliflower Plants. Agriculture 2022, 12, 1301. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Fujita, M. Hydrogen peroxide priming stimulates drought tolerance in mustard Brassica juncea L. seedlings. Plant Gene Trait. 2013, 4, 109–123. [Google Scholar]
- Mishra, S.S.; Panda, D. Leaf traits and antioxidant defense for drought tolerance during early growth stage in some popular traditional rice landraces from Koraput, India. Rice Sci. 2017, 24, 207–217. [Google Scholar] [CrossRef]
- Alam, M.M.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous salicylic acid ameliorates short-term drought stress in mustard (Brassica juncea L.) seedlings by up-regulating the antioxidant defense and glyoxalase system. Aust. J. Crop Sci. 2013, 7, 1053–1063. [Google Scholar]
- Ghafoor, R.; Akram, N.A.; Rashid, M.; Ashraf, M.; Iqbal, M.; Lixin, Z. Exogenously applied proline induced changes in key anatomical features and physio-biochemical attributes in water stressed oat (Avena sativa L.) plants. Physiol. Mol. Biol. Plants 2019, 25, 1121–1135. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M.; Shahbaz, M.; Humera, H. Ameliorating effect of foliar applied proline on nutrient uptake in water stressed maize (Zea mays L.) plants. Pak. J. Bot 2008, 40, 211–219. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Treatment | Plant Height (cm) | Fresh Weight (g)/Plant | Dry Weight (g)/Plant |
|---|---|---|---|---|
| BRRI dhan66 | T1 | 58.8 ± 2.15 bc | 0.836 ± 0.03 a | 0.152 ± 0.02 abc |
| T2 | 60.4 ± 1.99 ab | 0.851 ± 0.01 a | 0.156 ± 0.01 ab | |
| T3 | 60.1 ± 1.99 ab | 0.843 ± 0.03 a | 0.155 ± 0.01 ab | |
| T4 | 62.4 ± 2.74 a | 0.862 ± 0.04 a | 0.159 ± 0.00 a | |
| T5 | 53.9 ± 1.78 de | 0.646 ± 0.01 c | 0.133 ± 0.01 efg | |
| T6 | 55.2 ± 2.83 de | 0.712 ± 0.03 b | 0.139 ± 0.02 cdef | |
| T7 | 54.7 ± 1.77 de | 0.701 ± 0.07 bc | 0.138 ± 0.02 def | |
| T8 | 56.7 ± 2.70 cd | 0.743 ± 0.03 b | 0.143 ± 0.01 bcde | |
| BRRI dhan75 | T1 | 53.7 ± 1.37 e | 0.817 ± 0.01 a | 0.151 ± 0.03 abc |
| T2 | 55.5 ± 1.41 de | 0.831 ± 0.06 a | 0.155 ± 0.01 ab | |
| T3 | 55.7 ± 1.44 de | 0.824 ± 0.04 a | 0.155 ± 0.01 ab | |
| T4 | 56.7 ± 2.34 cd | 0.84 ± 0.03 a | 0.158 ± 0.02 a | |
| T5 | 37.9 ± 2.08 h | 0.271 ± 0.02 f | 0.103 ± 0.01 i | |
| T6 | 43.8 ± 1.23 g | 0.473 ± 0.02 e | 0.121 ± 0.02 gh | |
| T7 | 43.6 ± 1.44 g | 0.436 ± 0.05 e | 0.119 ± 0.01 g | |
| T8 | 47.6 ± 2.53 f | 0.551 ± 0.02 d | 0.128 ± 0.01 fgh |
| Variety | Treatment | Plant Height (cm) | Total Tillers per Hill | Effective Tillers per Hill | Panicle Length (cm) | Filled Grain per Panicle | Unfilled Grain per Panicle | Thousand-Seed Weight (g) | Seed Yield (g) per Hill |
|---|---|---|---|---|---|---|---|---|---|
| BRRI dhan66 | T1 | 116.3 ± 3.51 ab | 15.7 ± 0.58 ab | 14.3 ± 0.36 abc | 25.1 ± 0.65 abc | 177.7 ± 3.51 bc | 32.3 ± 1.15 ghi | 22.1 ± 1.06 ab | 21.5 ± 0.51 a |
| T2 | 118.5 ± 4.21 a | 16.3 ± 0.41 a | 14.7 ± 0.58 ab | 26.1 ± 1.01 ab | 185.3 ± 4.13 a | 29.3 ± 1.76 hi | 22.5 ± 0.50 a | 21.8 ± 1.03 a | |
| T3 | 117.8 ± 3.30 a | 15.7 ± 0.58 ab | 14.7 ± 0.65 ab | 25.9 ± 1.02 ab | 182.3 ± 4.41 abc | 31.7 ± 1.10 ght | 22.3 ± 0.55 ab | 21.7 ± 1.25 a | |
| T4 | 119.3 ± 3.51 a | 16.7 ± 1.53 a | 15.0 ± 1.50 a | 26.3 ± 2.33 a | 187.2 ± 4.00 a | 28.1 ± 1.87 i | 22.6 ± 1.30 a | 22.1 ± 1.13 a | |
| T5 | 107.0 ± 2.65 d | 13.7 ± 1.00 bc | 12.0 ± 1.00 c | 21.3 ± 0.95 d | 146.3 ± 2.89 f | 78.0 ± 1.73 d | 19.8 ± 0.73 b | 15.7 ± 0.93 d | |
| T6 | 110.2 ± 3.02 cd | 14.7 ± 0.58 ab | 13.0 ± 0.58 abc | 21.9 ± 0.62 d | 160.7 ± 4.16 de | 64.3 ± 1.53 e | 20.6 ± 0.54 ab | 17.5 ± 1.45 cd | |
| T7 | 109.3 ± 4.93 cd | 14.7 ± 0.63 ab | 12.3 ± 1.13 bc | 21.7 ± 0.66 d | 157.3 ± 3.30 e | 68.3 ± 1.53 e | 20.6 ± 0.28 ab | 17.1 ± 1.20 cd | |
| T8 | 112.7 ± 3.06 bc | 15.3 ± 1.26 ab | 13.3 ± 1.00 abc | 22.3 ± 1.53 d | 166.3 ± 3.62 d | 57.4 ± 2.46 f | 21.3 ± 1.70 ab | 18.7 ± 0.81 bc | |
| BRRI dhan75 | T1 | 108.3 ± 3.11 cd | 14.7 ± 0.58 ab | 13.3 ± 0.58 abc | 22.9 ± 0.85 cd | 176.0 ± 2.65 c | 35.3 ± 1.15 g | 20.6 ± 0.89 ab | 21.0 ± 0.50 ab |
| T2 | 110.3 ± 3.79 cd | 15.0 ± 1.00 ab | 13.7 ± 1.09 abc | 23.6 ± 0.52 bcd | 184.3 ± 3.21 ab | 34.3 ± 1.53 gh | 20.9 ± 0.49 ab | 21.4 ± 1.36 a | |
| T3 | 109.5 ± 4.82 cd | 15.0 ± 1.15 ab | 13.7 ± 1.53 abc | 23.5 ± 0.50 bcd | 182.4 ± 4.87 abc | 34.0 ± 2.00 gh | 20.7 ± 0.57 ab | 21.2 ± 1.24 a | |
| T4 | 111.4 ± 4.50 cd | 15.3 ± 1.57 ab | 14.1 ± 1.46 abc | 23.9 ± 1.84 abcd | 187.3 ± 4.00 a | 33.6 ± 2.04 gh | 21.3 ± 0.87 ab | 21.7 ± 1.52 a | |
| T5 | 63.7 ± 2.52 f | 8.3 ± 1.00 e | 5.7 ± 0.61 e | 11.6 ± 1.30 f | 56.3 ± 2.89 i | 146.3 ± 4.21 a | 13.0 ± 0.70 d | 4.2 ± 0.32 g | |
| T6 | 79.3 ± 2.63 e | 10.3 ± 1.16 de | 7.3 ± 0.40 de | 14.7 ± 0.53 e | 78.3 ± 3.06 h | 105.3 ± 3.13 c | 15.9 ± 0.43 c | 7.7 ± 0.35 f | |
| T7 | 77.3 ± 2.52 e | 10.3 ± 0.72 de | 7.0 ± 0.00 de | 14.4 ± 0.58 e | 71.7 ± 3.51 h | 111.6 ± 3.21 b | 15.5 ± 0.35 cd | 7.3 ± 0.32 f | |
| T8 | 81.6 ± 4.13 e | 11.6 ± 1.06 cd | 8.7 ± 0.76 d | 16.4 ± 1.30 e | 114.7 ± 5.03 g | 81.6 ± 3.90 d | 16.7 ± 0.80 c | 9.6 ± 0.210 e |
| Variety | Treatment | Photosynthetic Rate (µmol m−2 s−1) | Stomatal Conductance (mmol m−2 s−1) | Transpiration Rate (mmol m−2 s−1) | Relative Water Content |
|---|---|---|---|---|---|
| BRRI dhan66 | T1 | 15.7 ± 0.50 a | 0.414 ± 0.02 a | 6.58 ± 0.37 a | 90.6 ± 2.51 a |
| T2 | 16.2 ± 0.83 a | 0.426 ± 0.03 a | 6.81 ± 0.46 a | 93.8 ± 3.02 a | |
| T3 | 16.1 ± 1.08 a | 0.421 ± 0.02 a | 6.74 ± 0.50 a | 93.3 ± 2.53 a | |
| T4 | 16.4 ± 1.35 a | 0.437 ± 0.05 a | 6.95 ± 0.46 a | 94.4 ± 3.64 a | |
| T5 | 11.2 ± 0.84 e | 0.276 ± 0.03 cd | 4.06 ± 0.20 bcd | 70.4 ± 1.95 f | |
| T6 | 13.4 ± 0.40 cd | 0.313 ± 0.02 bc | 4.62 ± 0.40 bc | 81.4 ± 3.00 cd | |
| T7 | 13.1 ± 0.27 d | 0.305 ± 0.03 bc | 4.48 ± 0.52 bc | 80.1 ± 3.57 cd | |
| T8 | 13.8 ± 0.80 bcd | 0.351 ± 0.03 b | 5.04 ± 0.52 b | 84.3 ± 3.25 bc | |
| BRRI dhan75 | T1 | 15.1 ± 0.35 abc | 0.411 ± 0.01 a | 6.46 ± 0.30 a | 88.9 ± 3.72 ab |
| T2 | 15.5 ± 1.08 ab | 0.422 ± 0.02 a | 6.68 ± 0.31 a | 92.7 ± 1.95 a | |
| T3 | 15.5 ± 0.60 ab | 0.419 ± 0.02 a | 6.57 ± 0.41 a | 91.7 ± 2.05 a | |
| T4 | 15.7 ± 1.09 a | 0.430 ± 0.03 a | 6.81 ± 0.36 a | 92.4 ± 3.44 a | |
| T5 | 6.1 ± 0.42 g | 0.106 ± 0.00 f | 1.89 ± 0.06 f | 57.3 ± 1.05 g | |
| T6 | 9.3 ± 0.62 f | 0.186 ± 0.01 e | 3.13 ± 0.20 de | 71.2 ± 2.17 ef | |
| T7 | 9.2 ± 0.78 f | 0.175 ± 0.03 e | 2.91 ± 0.19 ef | 70.4 ± 2.39 f | |
| T8 | 10.5 ± 0.62 ef | 0.229 ± 0.03 de | 3.65 ± 0.46 cde | 76.6 ± 3.20 de |
| Variety | Treatment | Proline Content (µg g−1 FW) | Soluble Sugar Content (mg g−1 FW) | Starch Content (mg g−1 FW) |
|---|---|---|---|---|
| BRRI dhan66 | T1 | 1.36 ± 0.06 g | 31.6 ± 1.30 ab | 15.9 ± 0.95 abcd |
| T2 | 1.41 ± 0.19 g | 32.7 ± 2.37 a | 16.4 ± 1.40 ab | |
| T3 | 1.39 ± 0.22 g | 32.5 ± 2.46 a | 16.2 ± 1.20 ab | |
| T4 | 1.45 ± 0.15 g | 32.8 ± 2.18 a | 16.7 ± 1.18 a | |
| T5 | 3.75 ± 0.21 cd | 25.6 ± 1.70 d | 14.4 ± 1.36 ef | |
| T6 | 4.12 ± 0.24 ab | 27.1 ± 2.20 d | 15.1 ± 1.14 cde | |
| T7 | 4.06 ± 0.17 abc | 26.9 ± 1.10 d | 14.9 ± 1.21 de | |
| T8 | 4.25 ± 0.39 a | 29.6 ± 2.40 c | 15.5 ± 1.35 bcd | |
| BRRI dhan75 | T1 | 1.32 ± 0.25 g | 30.2 ± 1.60 bc | 16.0 ± 1.32 abc |
| T2 | 1.37 ± 0.20 g | 31.1 ± 1.95 abc | 16.5 ± 1.00 a | |
| T3 | 1.34 ± 0.29 g | 31.1 ± 1.15 abc | 16.4 ± 1.25 ab | |
| T4 | 1.41 ± 0.20 g | 31.5 ± 1.41 ab | 16.7 ± 1.18 a | |
| T5 | 2.88 ± 0.25 f | 20.3 ± 1.15 f | 10.3 ± 0.88 h | |
| T6 | 3.41 ± 0.34 de | 23.1 ± 1.36 e | 12.1 ± 0.67 g | |
| T7 | 3.37 ± 0.20 e | 22.8 ± 1.11 e | 11.9 ± 0.79 g | |
| T8 | 3.78 ± 0.21 bc | 25.3 ± 1.25 d | 13.9 ± 0.89 f |
| Variety | Treatment | %N | %P | %K | |||
|---|---|---|---|---|---|---|---|
| Roots | Shoots | Roots | Shoots | Roots | Shoots | ||
| BRRI dhan66 | T1 | 1.183 ± 0.03 a | 0.4072 ± 0.01 a | 0.1557 ± 0.01 cd | 0.0778 ± 0.00 c | 0.2447 ± 0.02 bc | 0.8217 ± 0.01 de |
| T2 | 1.185 ± 0.06 a | 0.4120 ± 0.03 a | 0.1590 ± 0.01 ab | 0.0789 ± 0.00 bc | 0.2480 ± 0.01 ab | 0.8320 ± 0.02 ab | |
| T3 | 1.184 ± 0.04 a | 0.410 ± 0.03 a | 0.1583 ± 0.01 abc | 0.0788 ± 0.00 bc | 0.2470 ± 0.00 abc | 0.8310 ± 0.03 ab | |
| T4 | 1.192 ± 0.03 a | 0.4170 ± 0.04 a | 0.1599 ± 0.03 a | 0.0791 ± 0.00 bc | 0.2510 ± 0.00 a | 0.8350 ± 0.01 a | |
| T5 | 1.112 ± 0.01 d | 0.3650 ± 0.1 c | 0.1465 ± 0.02 e | 0.0613 ± 0.00 f | 0.2360 ± 0.01 ef | 0.7260 ± 0.01 h | |
| T6 | 1.135 ± 0.01 c | 0.3750 ± 0.01 bc | 0.1490 ± 0.02 e | 0.0650 ± 0.00 e | 0.2390 ± 0.02 de | 0.7420 ± 0.01 fg | |
| T7 | 1.132 ± 0.03 c | 0.3720 ± 0.01 bc | 0.1480 ± 0.01 e | 0.0640 ± 0.00 ef | 0.2370 ± 0.03 de | 0.7390 ± 0.02 g | |
| T8 | 1.153 ± 0.01 b | 0.3860 ± 0.01 b | 0.1530 ± 0.01 d | 0.0690 ± 0.00 d | 0.2420 ± 0.01 cd | 0.7480 ± 0.03 f | |
| BRRI dhan75 | T1 | 1.181 ± 0.05 a | 0.4058 ± 0.03 a | 0.1570 ± 0.02 bc | 0.0811 ± 0.01 ab | 0.2450 ± 0.02 bc | 0.8157 ± 0.03 e |
| T2 | 1.184 ± 0.03 a | 0.4110 ± 0.04 a | 0.1598 ± 0.03 a | 0.0825 ± 0.00 a | 0.2481 ± 0.02 ab | 0.8260 ± 0.02 bcd | |
| T3 | 1.183 ± 0.06 a | 0.4090 ± 0.04 a | 0.1593 ± 0.03 ab | 0.0823 ± 0.01 a | 0.2476 ± 0.00 ab | 0.8240 ± 0.02 cd | |
| T4 | 1.189 ± 0.02 a | 0.4160 ± 0.03 a | 0.1606 ± 0.01 a | 0.0826 ± 0.00 a | 0.2492 ± 0.01 ab | 0.8290 ± 0.01 abc | |
| T5 | 0.938 ± 0.01 g | 0.2058 ± 0.02 f | 0.1240 ± 0.01 i | 0.0336 ± 0.00 i | 0.2161 ± 0.02 i | 0.5705 ± 0.01 k | |
| T6 | 0.971 ± 0.04 f | 0.2510 ± 0.01 e | 0.1310 ± 0.01 g | 0.0410 ± 0.00 h | 0.2260 ± 0.00 gh | 0.6130 ± 0.01 j | |
| T7 | 0.965 ± 0.01 f | 0.2430 ± 0.02 e | 0.1280 ± 0.01 h | 0.0380 ± 0.00 h | 0.2230 ± 0.01 h | 0.6110 ± 0.02 j | |
| T8 | 0.997 ± 0.02 e | 0.2830 ± 0.02 d | 0.1390 ± 0.02 f | 0.0520 ± 0.00 g | 0.2310 ± 0.01 fg | 0.6820 ± 0.03 i | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urmi, T.A.; Islam, M.M.; Zumur, K.N.; Abedin, M.A.; Haque, M.M.; Siddiqui, M.H.; Murata, Y.; Hoque, M.A. Combined Effect of Salicylic Acid and Proline Mitigates Drought Stress in Rice (Oryza sativa L.) through the Modulation of Physiological Attributes and Antioxidant Enzymes. Antioxidants 2023, 12, 1438. https://doi.org/10.3390/antiox12071438
Urmi TA, Islam MM, Zumur KN, Abedin MA, Haque MM, Siddiqui MH, Murata Y, Hoque MA. Combined Effect of Salicylic Acid and Proline Mitigates Drought Stress in Rice (Oryza sativa L.) through the Modulation of Physiological Attributes and Antioxidant Enzymes. Antioxidants. 2023; 12(7):1438. https://doi.org/10.3390/antiox12071438
Chicago/Turabian StyleUrmi, Tahmina Akter, Md. Moshiul Islam, Kamrun Naher Zumur, Md. Anwarul Abedin, M. Moynul Haque, Manzer H. Siddiqui, Yoshiyuki Murata, and Md. Anamul Hoque. 2023. "Combined Effect of Salicylic Acid and Proline Mitigates Drought Stress in Rice (Oryza sativa L.) through the Modulation of Physiological Attributes and Antioxidant Enzymes" Antioxidants 12, no. 7: 1438. https://doi.org/10.3390/antiox12071438