
Exogenous Allantoin Confers Rapeseed (Brassica campestris) Tolerance to Simulated Drought by Improving Antioxidant Metabolism and Physiology

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Exposure
2.2. Measurement of Growth Attributes
2.3. Estimation of Relative Water Content
2.4. Determination of Proline Content
2.5. Estimation of K+ Content
2.6. Determination of Electrolyte Leakage
2.7. Quantification of Malondialdehyde Content
2.8. Estimation of Hydrogen Peroxide Content
2.9. Estimation of Ascorbate and Glutathione Content
2.10. Enzyme Extraction, Protein Determination, and Antioxidant Enzyme Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Names of the Enzymes | Name of the Chemicals and Their Concentrations | Wavelength | Extinction Coefficient | References |
|---|---|---|---|---|
| Ascorbate peroxidase (APX; EC: 1.11.1.1) | L-ascorbic acid (Asc; 0.5 mM) Ethylenediaminetetraacetic acid (EDTA; 0.1 mM) Potassium phosphate (K-P) buffer (15 mM, pH 7.0) Hydrogen peroxide (H2O2; 0.1 mM) | 290 nm | 2.8 mM−1 cm−1 | [49] |
| Dehydroascobate reductase (DHAR; EC: 1.8.5.1) | Reduced glutathione (GSH; 2.5 mM) EDTA (0.1 mM) K-P buffer (50 mM, pH 7.0) Dehydroascorbate (DHA; 0.1 mM) | 265 nm | 14 mM−1 cm−1 | [49] |
| Glutathione reductase (GR; EC: 1.6.4.2) | Oxidized glutathione (GSSG; 1 mM) Nicotinamide adenine dinucleotide phosphate (NADPH; 0.2 mM) EDTA (1 mM) K-P buffer (0.1 M, pH 7.8) | 340 nm | 6.2 mM−1 cm−1 | [50] |
| Glutathione peroxidase (GPX; EC: 1.11.1.9) | NADPH (0.12 mM) GR (1 unit) GSH (2 mM) Sodium azide (1 mM) EDTA (1 mM) K-P buffer (100 mM, pH 7.0) | 340 nm | 6.62 mM−1 cm−1 | [50,51] |
| Catalase (CAT; EC: 1.11.1.6) | H2O2 (15 mM) K-P buffer (50 mM, pH 7.0) | 240 nm | 39.4 M−1 cm−1 | [50] |
| Glyoxalase I (Gly I; EC: 4.4.1.5) | GSH (100 mM) Magnesium phosphate (16 mM) Methylglyoxal (100 mM) Sodium phosphate buffer (0.1 M) | 240 nm | 3.37 mM−1 cm−1 | [50] |
| Glyoxalase II (Gly II; EC: 3.1.2.6) | 5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB; 0.2 mM) S-D-lactoylglutathione (1 mM) Tris-HCl buffer (100 mM, pH 7.2) | 412 nm | 13.6 mM−1 cm−1 | [52] |
2.11. Yield and Yield Attributes
2.12. Statistical Analysis
3. Results
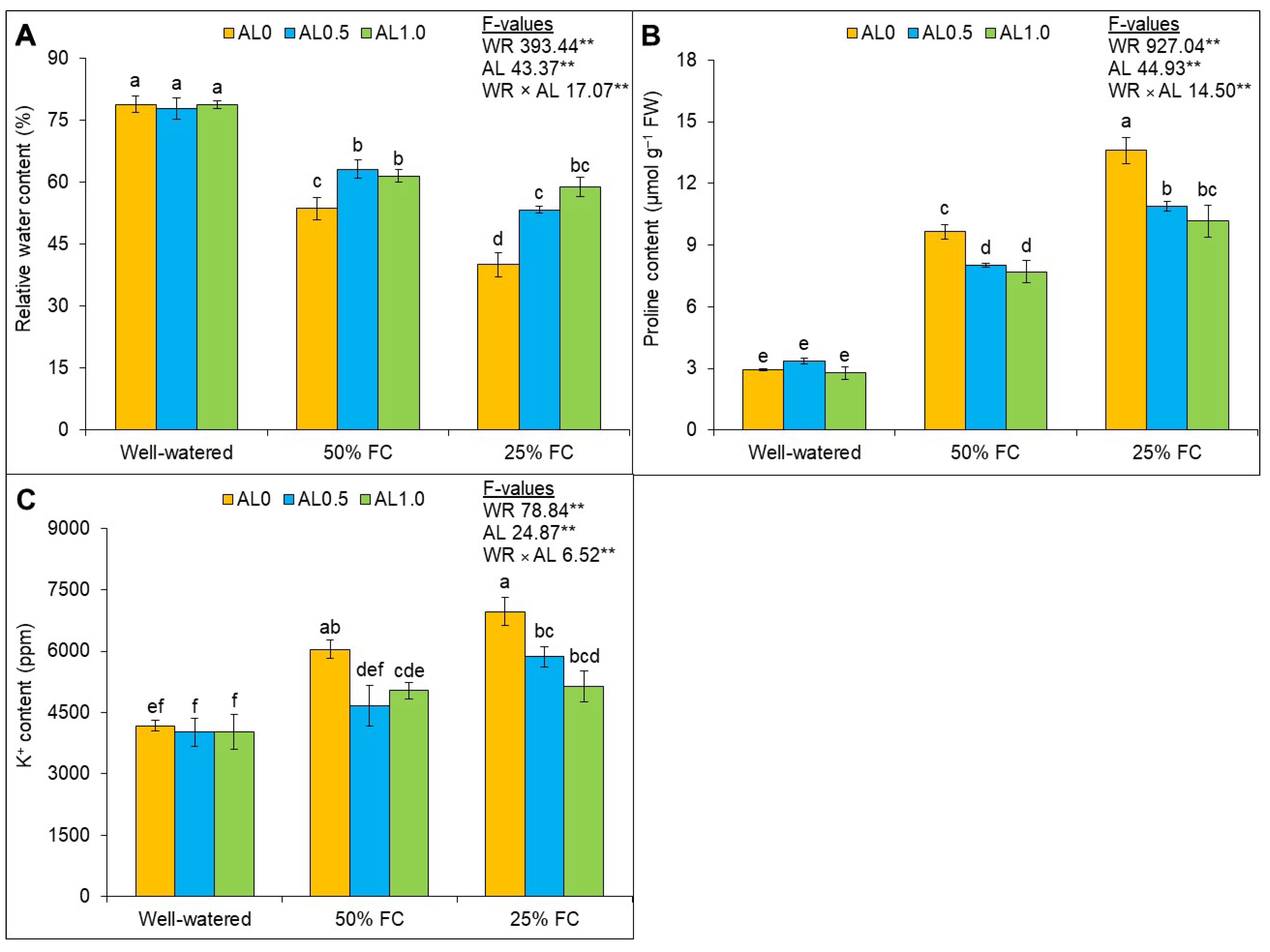
3.1. Allantoin Enhances Relative Water Availability, Maintains Osmoregulation, and Regulates Potassium Ion Accumulation in Plants under Drought
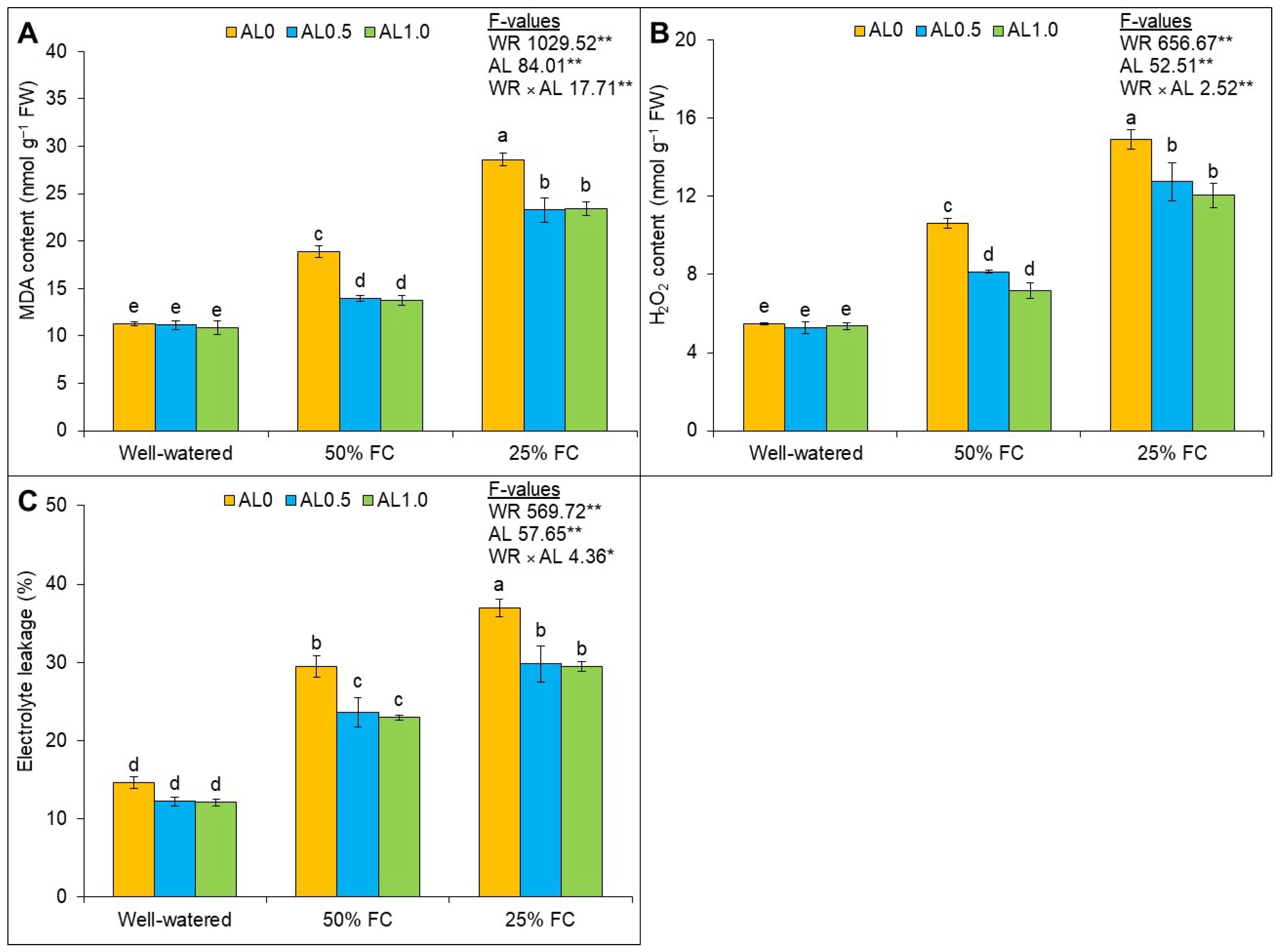
3.2. Exogenous Allantoin Protects ROS-Induced Cellular Damage and Increases Membrane Stability in Plants under Drought
3.3. Allantoin Regulates AsA-GSH Pool under Drought
3.4. Allantoin Causes Enhanced Antioxidant Activities and Boosts Drought Tolerance of Plants
3.5. Allantoin Improves Plant Growth and Biomass Accumulation under Drought
3.6. Effect of Allantoin on the Partitioning Fresh and Dry Weight of Rapeseed under Drought
3.7. Effect of Allantoin on the Yield and Yield Attributes of Rapeseed under Drought
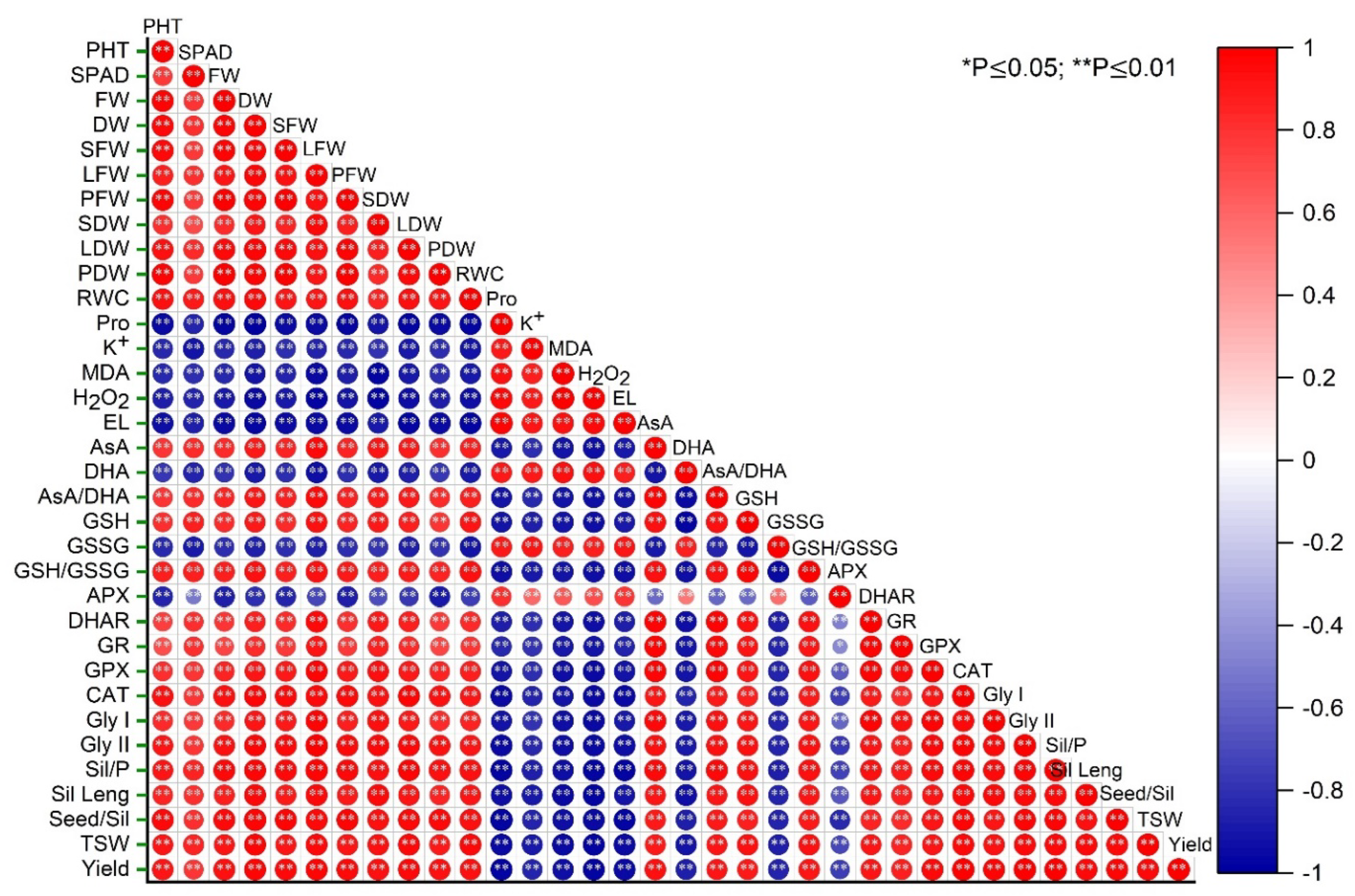
3.8. Correlation among Oxidative Stress Indicators, Antioxidant Defense System, Growth, and Yield Attributes of Rapeseed
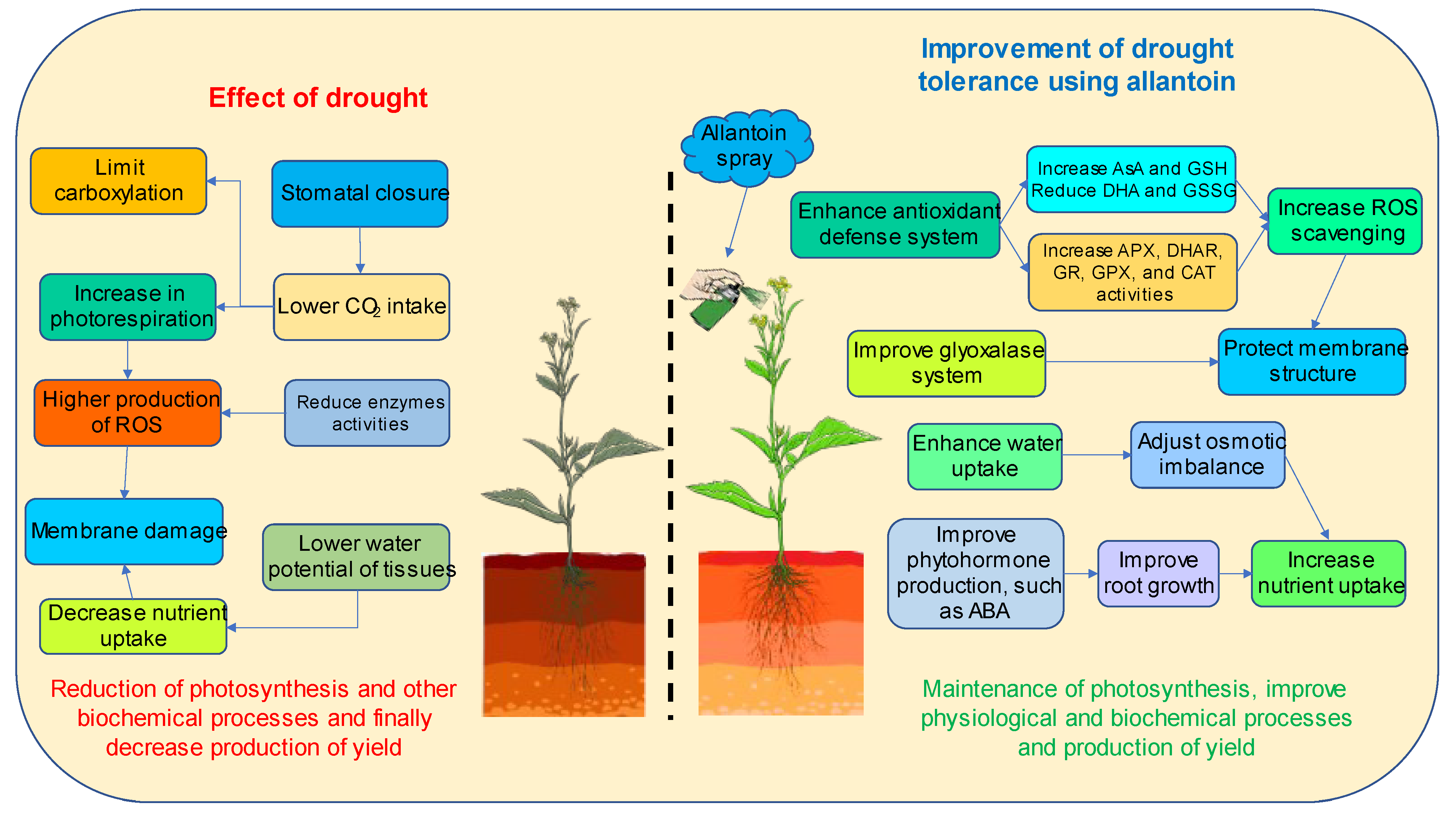
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and drought stresses in crops and approaches for their mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.; Modi, P.; Dave, A.; Vijapura, A.; Patel, D.; Patel, M. Effect of abiotic stress on crops. In Sustainable Crop Production; Hasanuzzaman, M., Nogueira, T.A.R., Eds.; IntechOpen: London, UK, 2020; p. 3. [Google Scholar] [CrossRef]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2016, 45, 523–539. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Wasaya, A.; Zhang, X.; Fang, Q.; Zongzheng, Y. Root phenotyping for drought tolerance: A review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Kuromori, T.; Urano, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Drought stress responses and resistance in plants: From cellular responses to long distance intercellular communication. Front. Plant Sci. 2020, 11, 556972. [Google Scholar] [CrossRef] [PubMed]
- Caser, M.; Angiolillo, F.D.; Chitarra, W.; Lovisolo, C.; Ruffoni, B.; Pistelli, L.; Pistelli, L.; Scariot, V. Ecophysiological and phytochemical responses of Salvia sinaloensis Fern. to drought stress. J. Plant Growth Regul. 2018, 84, 383–394. [Google Scholar] [CrossRef]
- Kohli, S.K.; Khanna, K.; Bhardwaj, R.; Abd_Allah, E.F.; Ahmad, P.; Corpas, F.J. Assessment of subcellular ROS and NO metabolism in higher plants: Multifunctional signaling molecules. Antioxidants 2019, 8, 641. [Google Scholar] [CrossRef] [Green Version]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Rastogi, A.; Yadav, D.K.; Szymańska, R.; Kruk, J.; Sedlářová, M.; Pospíšil, P. Singlet oxygen scavenging activity of tocopherol and plastochromanol in Arabidopsis thaliana: Relevance to photooxidative stress: 1O2 Scavenging by tocopherol and plastochromanol. Plant Cell Environ. 2014, 37, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Scirè, A.; Cianfruglia, L.; Minnelli, C.; Romaldi, B.; Laudadio, E.; Galeazzi, R.; Antognelli, C.; Armeni, T. Glyoxalase 2: Towards a broader view of the second player of the glyoxalase system. Antioxidants 2022, 11, 2131. [Google Scholar] [CrossRef] [PubMed]
- Garai, S.; Bhowal, B.; Kaur, C.; Singla-Pareek, S.L.; Sopory, S.K. What signals the glyoxalase pathway in plants? Physiol. Mol. Biol. Plants 2021, 27, 2407–2420. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Macalister, C.J. A new cell proliferant: Its clinical application in the treatment of ulcers. Br. Med. J. 1912, 1, 10–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Chowrasia, S.; Gaur, V.S.; Mondal, T.K. Allantoin: Emerging role in plant abiotic stress tolerance. Plant Mol. Biol. Rep. 2021, 39, 648–661. [Google Scholar] [CrossRef]
- Watanabe, S.; Matsumoto, M.; Hakomori, Y.; Takagi, H.; Shimada, H.; Sakamoto, A. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant Cell Environ. 2014, 37, 1022–1036. [Google Scholar] [CrossRef] [Green Version]
- Thu, S.W.; Lu, M.; Carter, A.M.; Collier, R.; Gandin, A.; Sitton, C.C.; Tegeder, M. Role of ureides in source-to-sink transport of photoassimilates in non-fixing soybean. J. Exp. Bot. 2020, 71, 4495–4511. [Google Scholar] [CrossRef]
- Fahad, S.; Khan, F.A.; Pandupuspitasari, N.; Hussain, S.; Khan, I.A.; Saeed, M.; Saud, S.; Hassan, S.; Adnan, M.; Arif, M.; et al. Suppressing photorespiration for the improvement in photosynthesis and crop yields: A review on the role of S-allantoin as a nitrogen source. J. Environ. Manag. 2019, 237, 644–651. [Google Scholar] [CrossRef]
- Yobi, A.; Wone, B.W.; Xu, W.; Alexander, D.C.; Guo, L.; Ryals, J.A.; Oliver, M.J.; Cushman, J.C. Metabolomic profiling in Selaginella lepidophylla at various hydration states provides new insights into the mechanistic basis of desiccation tolerance. Mol. Plant 2013, 6, 369–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casartelli, A.; Riewe, D.; Hubberten, H.M.; Altmann, T.; Hoefgen, R.; Heuer, S. Exploring traditional Aus-type rice for metabolites conferring drought tolerance. Rice 2018, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Nishad, J.; Panda, A.K.; Chowrasia, S.; Nirmala, C.; Mondal, T.K. Allantoin mediated regulation of miRNAs for short term salinity stress tolerance in Oryza sativa L. cv. IR-29. J. Plant Biochem. Biotechnol. 2022, 31, 953–960. [Google Scholar] [CrossRef]
- Dresler, S.; Hawrylak-Nowak, B.; Kováčik, J.; Pochwatka, M.; Hanaka, A.; Strzemski, M.; Sowa, I.; Wójciak-Kosior, M. Allantoin attenuates cadmium-induced toxicity in cucumber plants. Ecotoxicol. Environ. Saf. 2019, 170, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.F.A.; Tahjib-Ul-Arif, M.; Sohag, A.A.M.; Abdel Latef, A.A.H.; Ragaey, M.M. Mechanistic insight of allantoin in protecting tomato plants against ultraviolet C stress. Plants 2021, 10, 11. [Google Scholar] [CrossRef]
- Wang, P.; Kong, C.H.; Sun, B.; Xu, X.H. Distribution and function of allantoin (5-ureidohydantoin) in rice grains. J. Agric. Food Chem. 2012, 60, 2793–2798. [Google Scholar] [CrossRef]
- Redillas, M.C.F.R.; Bang, S.W.; Lee, D.; Kim, Y.S.; Jung, H.; Chung, P.J.; Suh, J.; Kim, J. Allantoin accumulation through overexpression of ureide permease1 improves rice growth under limited nitrogen conditions. Plant Biotechnol. J. 2018, 17, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Irani, S.; Todd, C.D. Exogenous allantoin increases Arabidopsis seedlings tolerance to NaCl stress and regulates expression of oxidative stress response genes. J. Plant Physiol. 2018, 221, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Channaoui, S.; Kahkahi, R.E.; Charafi, J.; Mazouz, H.; Fechtali, M.E.; Nabloussi, A. Germination and seedling growth of a set of rapeseed (Brassica napus) varieties under drought stress conditions. Int. J. Environ. Agric. Biotechnol. 2017, 2, 487–494. [Google Scholar] [CrossRef]
- Zhu, M.; Monroe, J.G.; Suhail, Y.; Villiers, F.; Mullen, J.; Pater, D.; Hauser, F.; Jeon, B.W.; Bader, J.S.; Kwak, J.M.; et al. Molecular and systems approaches towards drought-tolerant canola crops. New Phytol. 2016, 210, 1169–1189. [Google Scholar] [CrossRef] [Green Version]
- Ahmar, S.; Liaqat, N.; Hussain, M.; Salim, M.A.; Shabbir, M.A.; Ali, M.Y.; Noushahi, H.A.; Bilal, M.; Atta, B.; Rizwan, M. Effect of abiotic stresses on Brassica species and role of transgenic breeding for adaptation. Asian J. Res. Crop Sci. 2019, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different Brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Dawood, M.G.; Sadak, M.S. Physiological role of glycinebetaine in alleviating the deleterious effects of drought stress on canola plants (Brassica napus L.). Middle East J. Agric. Res. 2014, 3, 943–954. [Google Scholar]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, C.; Dinis, L.T.; Ferreira, H.; Coutinho, J.; Moutinho-Pereira, J.; Correia, C.M. Salicylic acid increases drought adaptability of young olive trees by changes on redox status and ionome. Plant Physiol. Biochem. 2019, 141, 315–324. [Google Scholar] [CrossRef]
- Hassan, M.U.; Chattha, M.U.; Chattha, M.B.; Mahmood, A.; Sahi, S.T. Chemical composition and methane yield of sorghum as influenced by planting methods and cultivars. J. Anim. Plant Sci. 2019, 29, 251–259. [Google Scholar]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Improving tolerance against drought in canola by penconazole and calcium. Pestic. Biochem. Physiol. 2018, 149, 123–136. [Google Scholar] [CrossRef] [PubMed]
- BARI (Bangladesh Agricultural Research Institute). Krishi Projukti Hatboi; Bangladesh Agricultural Research Institute: Gazipur, Bangladesh, 2021. (In Bengali) [Google Scholar]
- Liu, L.; Liu, D.; Wang, Z.; Zou, C.; Wang, B.; Zhang, H.; Gai, Z.; Zhang, P.; Wang, Y.; Li, C. Exogenous allantoin improves the salt tolerance of sugar beet by increasing putrescine metabolism and antioxidant activities. Plant Physiol. Biochem. 2020, 154, 699–713. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teari, D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplast. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide induced chilling tolerance in mung beans mediated through ABA independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; He, W.; Guo, J.; Chang, X.; Su, P.; Zhang, L. Increased sensitivity to salt stress in ascorbate-deficient Arabidopsis mutant. J. Exp. Bot. 2005, 56, 3041–3049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasanuzzaman, M.; Raihan, M.R.H.; Siddika, A.; Rahman, K.; Nahar, K. Supplementation with Ascophyllum nodosum extracts mitigates arsenic toxicity by modulating reactive oxygen species metabolism and reducing oxidative stress in rice. Ecotoxicol. Environ. Saf. 2023, 255, 114819. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Hasanuzzaman, M.; Raihan, M.R.H.; Khojah, E.; Samra, B.N.; Fujita, M.; Nahar, K. Biochar and chitosan regulate antioxidant defense and methylglyoxal detoxification systems and enhance salt tolerance in jute (Corchorus olitorius L.). Antioxidants 2021, 10, 2017. [Google Scholar] [CrossRef]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dorr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Principato, G.B.; Rosi, G.; Talesa, V.; Govannini, E.; Uolila, L. Purification and characterization of two forms of glyoxalase II from rat liver and brain of Wistar rats. Biochim. Biophys. Acta 1987, 911, 349–355. [Google Scholar] [CrossRef]
- CoStat. CoStat-Statistics Software, Version 6.400; CoHort Software: Monterey, CA, USA, 2008.
- SPSS. Stastical Package for Social Sciences, Version 27.0. IBM SPSS Statistics for Windows. IBM Crop.: Armonk, NY, USA, 2020.
- Alvarez, M.E.; Savouré, A.; Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2021, 27, 39–55. [Google Scholar] [CrossRef]
- Munns, R. Plant adaptations to salt and water stress: Differences and commonalities. Adv. Bot. Res. 2011, 57, 1–32. [Google Scholar]
- Ahmed, N.; Rahman, K.; Rahman, M.; Sathi, K.S.; Alam, M.M.; Nahar, K.; Islam, M.S.; Hasanuzzaman, M. Insight into the thiourea-induced drought tolerance in two chickpea varieties: Regulation of osmoprotection, reactive oxygen species metabolism and glyoxalase system. Plant Physiol. Biochem. 2021, 167, 449–458. [Google Scholar] [CrossRef]
- Dien, D.C.; Mochizuki, T.; Yamakawa, T. Effect of various drought stresses and subsequent recovery on proline, total soluble sugar and starch metabolisms in rice (Oryza sativa L.) varieties. Plant Prod. Sci. 2019, 22, 530–545. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.; Sandhu, P.S.; Chahal, G.K.; Jaidka, M. Foliar application of thiourea ameliorates drought stress by elevating antioxidant level in rainfed maize (Zea mays L.). Agric. Res. J. 2020, 57, 522–528. [Google Scholar] [CrossRef]
- Wu, H.; Liu, X.; You, L.; Zhang, L.; Yu, J.; Zhou, D.; Zhao, J.; Feng, J. Salinity-induced effects in the halophyte Suaeda salsa using NMR-based metabolomics. Plant Mol. Biol. Rep. 2012, 30, 590–598. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.J.; Guo, L.; Alexander, D.C.; Ryals, J.A.; Wone, B.W.M.; Cushman, J.C. A sister group contrast using untargeted global metabolomic analysis delineates the biochemical regulation underlying desiccation tolerance in Sporobolus stapfianus. Plant Cell 2011, 23, 1231–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Zhang, Y.; Liu, A.; Li, D.; Wang, X.; Dossa, K.; Zhou, R.; Yu, J.; Zhang, Y.; Wang, L.; et al. Transcriptomic and metabolomic profiling of drought tolerant and susceptible sesame genotypes in response to drought stress. BMC Plant Biol. 2019, 19, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, Y.; Wang, Y.; Du, F.; Zhang, Y.; Yin, M.; Zhao, X.; Xu, J.; Yang, Y.; Wang, W.; et al. Transcriptome and metabolome analyses reveal complex molecular mechanisms involved in the salt tolerance of rice induced by exogenous allantoin. Antioxidants 2022, 11, 2045. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Dong, G.; Ma, Y.; Zhao, S.; Liu, X.; Li, X.; Li, Y.; Hou, B. Rice glycosyltransferase gene UGT85E1 is involved in drought stress tolerance through enhancing abscisic acid response. Front. Plant Sci. 2021, 12, 790195. [Google Scholar] [CrossRef]
- Lu, S.; Jia, Z.; Meng, X.; Chen, Y.; Wang, S.; Fu, C.; Yang, L.; Zhou, R.; Wang, B.; Cao, Y. Combined metabolomic and transcriptomic analysis reveals allantoin enhances drought tolerance in rice. Int. J. Mol. Sci. 2022, 23, 14172. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–179. [Google Scholar] [CrossRef]
- Zhang, C.; Shi, S.; Liu, Z.; Yang, F.; Yin, G. Drought tolerance in alfalfa (Medicago sativa L.) varieties is associated with enhanced antioxidative protection and declined lipid peroxidation. J. Plant Physiol. 2019, 232, 226–240. [Google Scholar] [CrossRef]
- Laloi, C.; Apel, K.; Danon, A. Reactive oxygen signaling: The latest news. Curr. Opin. Plant Biol. 2004, 7, 323–328. [Google Scholar] [CrossRef]
- Werner, A.K.; Witte, C.-P. The biochemistry of nitrogen mobilization: Purine ring catabolism. Trends Plant Sci. 2011, 16, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Lamberto, I.; Percudani, R.; Gatti, R.; Folli, C.; Petrucco, S. Conserved alternative splicing of Arabidopsis transthyretin-like determines protein localization and S-allantoin synthesis in peroxisomes. Plant Cell 2010, 22, 1564–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, H.; Ishiga, Y.; Watanabe, S.; Konishi, T.; Egusa, M.; Akiyoshi, N.; Matsuura, T.; Mori, I.C.; Hirayama, T.; Kaminaka, H.; et al. Allantoin, a stress-related purine metabolite, can activate jasmonate signaling in a MYC2-regulated and abscisic acid-dependent manner. J. Exp. Bot. 2016, 67, 2519–2532. [Google Scholar] [CrossRef]
- Lescano, I.; Bogino, M.F.; Martini, C.; Tessi, T.M.; González, C.A.; Schumacher, K.; Desimone, M. Ureide permease 5 (AtUPS5) connects cell compartments involved in ureide metabolism. Plant Physiol. 2020, 182, 1310–1325. [Google Scholar] [CrossRef]
- Nourimand, M.; Todd, C.D. Allantoin contributes to the stress response in cadmium-treated Arabidopsis roots. Plant Physiol. Biochem. 2017, 119, 103–109. [Google Scholar] [CrossRef]
- Watanabe, S.; Nakagawa, A.; Izumi, S.; Shimada, H.; Sakamoto, A. RNA interference-mediated suppression of xanthine dehydrogenase reveals the role of purine metabolism in drought tolerance in Arabidopsis. FEBS Lett. 2010, 584, 1181–1186. [Google Scholar] [CrossRef] [Green Version]
- Alamillo, J.M.; Díaz-Leal, J.L.; Sánchez-Moran, M.V.; Pineda, M. Molecular analysis of ureide accumulation under drought stress in Phaseolus vulgaris L. Plant Cell Environ. 2010, 33, 1828–1837. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kumar, A.; Yadav, S.; Singh, I.K. Reactive oxygen species-mediated signaling during abiotic stress. Plant Gene 2019, 18, 100173. [Google Scholar] [CrossRef]
- Jovanović, S.V.; Kukavica, B.; Vidović, M.; Morina, F.; Menckhoff, L. Class III peroxidases: Functions, localization and redox regulation of isoenzymes. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 269–300. [Google Scholar]
- Seminario, A.; Song, L.; Zulet, A.; Nguyen, H.T.; Gonz´alez, E.M.; Larrainzar, E. Drought stress causes a reduction in the biosynthesis of ascorbic acid in soybean plants. Front. Plant Sci. 2017, 8, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gus’kov, E.P.; Shkurat, T.P.; Milyutina, N.P.; Prokof’ev, V.N.; Pokudina, I.O.; Mashkina, E.V.; Timofeeva, I.V. Effect of allantoin on the activity of enzymes providing regulation of the ROS-dependent status of an organism. Dokl. Biochem. Biophys. 2001, 379, 239–242. [Google Scholar] [CrossRef]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Bela, K.; Horváth, E.; Gallé, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef]
- Mehla, N.; Sindhi, V.; Josula, D.; Bisht, P.; Wani, S.H. An introduction to antioxidants and their roles in plant stress tolerance. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 1–23. [Google Scholar]
- Czarnocka, W.; Karpinski, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Rang, Z.W.; Jagadish, S.V.K.; Zhou, Q.M.; Craufurd, P.Q.; Heuer, S. Effect of heat and drought stress on pollen germination and spikelet fertility in rice. Environ. Exp. Bot. 2011, 70, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Qi, F.; Zhang, F. Cell cycle regulation in the plant response to stress. Front. Plant Sci. 2020, 10, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowtham, H.G.; Singh, S.B.; Shilpa, N.; Aiyaz, M.; Nataraj, K.; Udayashankar, A.C.; Amruthesh, K.N.; Murali, M.; Poczai, P.; Gafur, A.; et al. Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants 2022, 11, 1763. [Google Scholar] [CrossRef]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of fourteen bread wheat (Triticum aestivum L.) genotypes by observing gas exchange parameters, relative water and chlorophyll content, and yield attributes under drought stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Mohi-Ud-Din, M.; Hossain, M.A.; Rohman, M.M.; Uddin, M.N.; Haque, M.S.; Ahmed, J.U.; Hossain, A.; Hassan, M.M.; Mostofa, M.G. Multivariate analysis of morpho-physiological traits reveals differential drought tolerance potential of bread wheat genotypes at the seedling stage. Plants 2021, 10, 879. [Google Scholar] [CrossRef]
- Hopkins, H.G.; Hüner, N.P.A. Plants and Inorganic Nutrients. In Introduction to Plant Physiology, 4th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2009; pp. 61–76. [Google Scholar]
- Esmaili, E.; Kapourchal, S.A.; Malakouti, M.J.; Homaee, M. Interactive effect of salinity and two nitrogen fertilizers on growth and composition of sorghum. Plant Soil Environ. 2008, 54, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Sikder, R.K.; Wang, X.R.; Zhang, H.S.; Gui, H.P.; Dong, Q.; Jin, D.S.; Song, M.Z. Nitrogen enhances salt tolerance by modulating the antioxidant defense system and osmoregulation substance content in Gossypium hirsutum. Plants 2020, 9, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wang, X.; Lu, Q.; Zhang, H.; Chen, J.; Zhang, H.; Wang, Y.; Li, C. Allantoin, a purine metabolite, confers saline–alkaline tolerance to sugar beet by triggering a self-amplifying feedback loop comprising jasmonic acid and nitric oxide. Environ. Exp. Bot. 2023, 206, 105172. [Google Scholar] [CrossRef]










| Water Regimes (WRs) | Allantoin (AL) Doses | No. of Silique Plant−1 | Silique Length (cm) | No. of Seeds Silique−1 | 1000-Seed Weight (g) | Seed Yield (g Plant−1) |
|---|---|---|---|---|---|---|
| Well-watered | 0 mM | 39.33 a ± 1.72 | 4.60 a ± 0.28 | 22.53 a ± 0.93 | 2.60 a ± 0.10 | 2.28 a ± 0.11 |
| 0.5 mM | 40.07 a ± 2.04 | 4.25 ab ± 0.40 | 22.40 a ± 0.82 | 2.47 a ± 0.06 | 2.26 a ± 0.02 | |
| 1.0 mM | 42.67 a ± 0.12 | 4.30 ab ± 0.21 | 22.16 a ± 1.88 | 2.47 a ± 0.09 | 2.17 a ± 0.11 | |
| 50% FC | 0 mM | 23.17 d ± 0.68 | 3.07 c ± 0.10 | 12.37 c ± 0.50 | 1.72 c ± 0.11 | 1.14 d ± 0.04 |
| 0.5 mM | 27.87 c ± 1.22 | 3.78 b ± 0.21 | 14.20 bc ± 0.60 | 2.13 b ± 0.13 | 1.34 c ± 0.04 | |
| 1.0 mM | 32.98 b ± 2.39 | 3.95 b ± 0.09 | 14.97 b ± 0.55 | 2.18 b ± 0.04 | 1.80 b ± 0.12 | |
| 25% FC | 0 mM | 16.41 f ± 0.59 | 2.23 e ± 0.07 | 6.40 e ± 0.20 | 1.33 d ± 0.06 | 0.66 f ± 0.01 |
| 0.5 mM | 18.90 ef ± 0.26 | 2.41 de ± 0.11 | 8.30 de ± 0.20 | 1.57 cd ± 0.08 | 0.76 ef ± 0.05 | |
| 1.0 mM | 22.34 de ± 1.16 | 2.96 cd ± 0.18 | 9.22 d ± 0.34 | 1.71 c ± 0.06 | 0.91 e ± 0.02 | |
| F-values | WR | 568.64 ** | 176.52 ** | 687.55 ** | 292.41 ** | 980.07 ** |
| AL | 49.78 ** | 9.74 ** | 9.83 ** | 18.37 ** | 33.52 ** | |
| WR × AL | 4.39 * | 8.65 ** | 3.59 * | 12.89 ** | 24.04 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raihan, M.R.H.; Rahman, M.; Rastogi, A.; Fujita, M.; Hasanuzzaman, M. Exogenous Allantoin Confers Rapeseed (Brassica campestris) Tolerance to Simulated Drought by Improving Antioxidant Metabolism and Physiology. Antioxidants 2023, 12, 1508. https://doi.org/10.3390/antiox12081508
Raihan MRH, Rahman M, Rastogi A, Fujita M, Hasanuzzaman M. Exogenous Allantoin Confers Rapeseed (Brassica campestris) Tolerance to Simulated Drought by Improving Antioxidant Metabolism and Physiology. Antioxidants. 2023; 12(8):1508. https://doi.org/10.3390/antiox12081508
Chicago/Turabian StyleRaihan, Md. Rakib Hossain, Mira Rahman, Anshu Rastogi, Masayuki Fujita, and Mirza Hasanuzzaman. 2023. "Exogenous Allantoin Confers Rapeseed (Brassica campestris) Tolerance to Simulated Drought by Improving Antioxidant Metabolism and Physiology" Antioxidants 12, no. 8: 1508. https://doi.org/10.3390/antiox12081508