
Chemical Components and Antioxidant Activity of Geotrigona sp. and Tetragonisca fiebrigi Stingless Bee Cerumen Reduce Juglone-Induced Oxidative Stress in Caenorhabditis elegans

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Ethanolic Extracts of Cerumen from Geotrigona sp. (EEC-G) and Tetragonisca fiebrigi (EEC-T)
2.2. Chemical Composition
2.2.1. HPLC Analysis
2.2.2. GC Analysis
2.2.3. ICP-OES Analysis
2.3. In Vitro Antioxidant Activity
2.3.1. 2,2-Diphenyl-1-picrylhydrazyl (DPPH•) Free Radical Scavenging
2.3.2. Elimination of the Free Radical 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS•+)
2.4. Antioxidant Activity in Human Erythrocytes
2.4.1. Hemolysis and Oxidative Hemolysis
2.4.2. Malondialdehyde Dosage (MDA)
2.5. Antioxidant Activity in C. elegans
2.5.1. Acute Toxicity Assay
2.5.2. Oxidative Stress Resistance Assay
2.6. Statistical Analysis
3. Results
3.1. Chemical Composition
3.2. In Vitro Antioxidant Activity
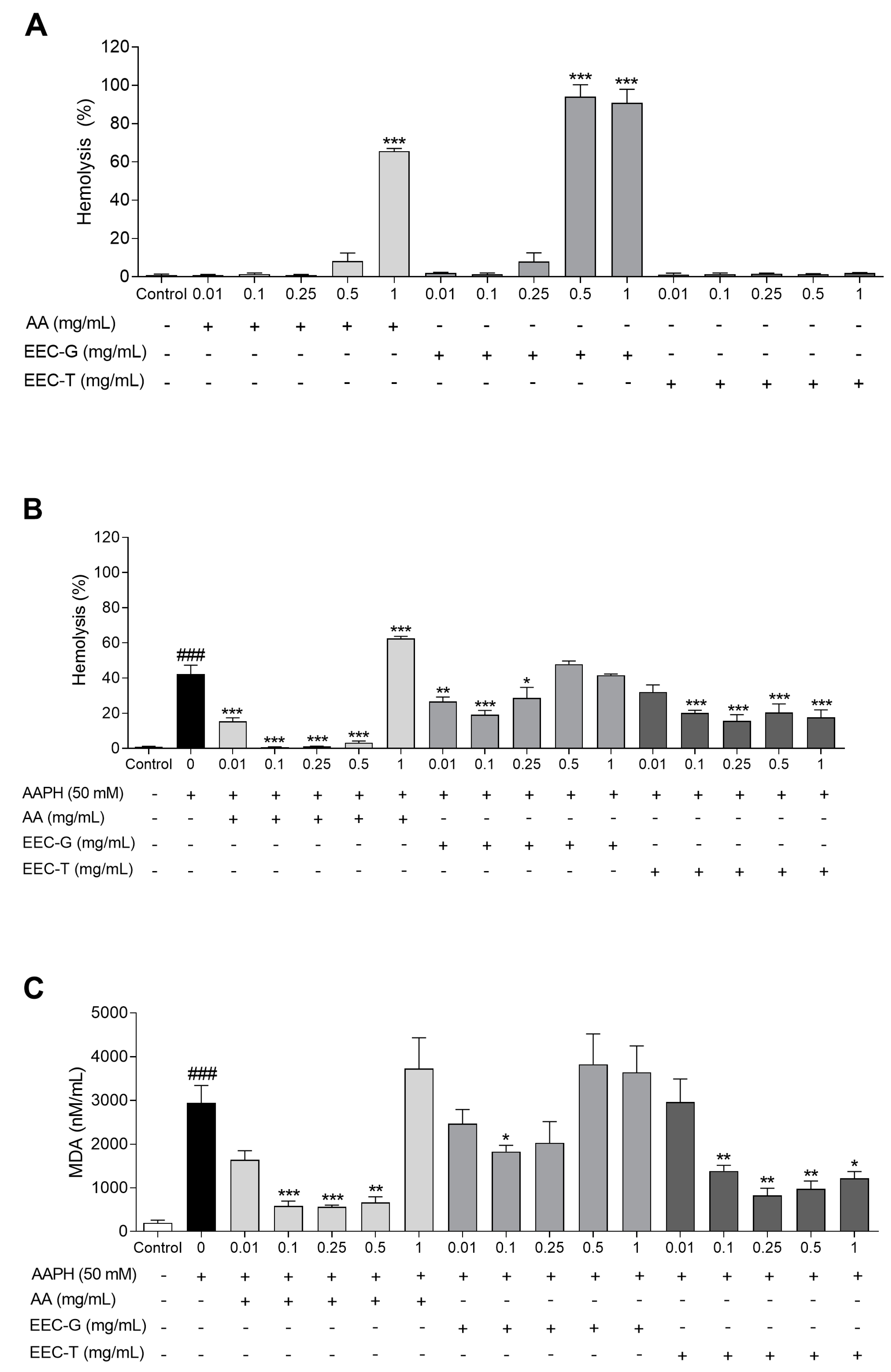
3.3. Antioxidant Activity in Human Erythrocytes
3.4. Acute Toxicity and Antioxidant Activity in C. elegans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grüter, C. Evolution and Diversity of Stingless Bees. In Stingless Bees; Fascinating Life Sciences; Springer: Cham, Switzerland, 2020; pp. 43–86. [Google Scholar] [CrossRef]
- Meléndez Ramírez, V.; Ayala, R.; Delfín González, H. Crop Pollination by Stingless Bees. In Pot-Pollen in Stingless Bee Melittology; Vit, P., Pedro, S., Roubik, D., Eds.; Springer: Cham, Switzerland, 2018; pp. 139–153. [Google Scholar] [CrossRef]
- Al-Hatamleh, M.A.I.; Boer, J.C.; Wilson, K.L.; Plebanski, M.; Mohamud, R.; Mustafa, M.Z. Antioxidant-Based Medicinal Properties of Stingless Bee Products: Recent Progress and Future Directions. Biomolecules 2020, 10, 923. [Google Scholar] [CrossRef]
- Hamilton, K.D.; Brooks, P.R.; Ogbourne, S.M.; Russell, F.D. Natural products isolated from Tetragonula carbonaria cerumen modulate free radical-scavenging and 5-lipoxygenase activities in vitro. BMC Complement. Altern. Med. 2017, 17, 232. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.F.; dos Santos, U.P.; Rocha, P.D.S.D.; Damião, M.J.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Estevinho, L.M.; Souza, K.D.P.; dos Santos, E.L. Antimicrobial, Antioxidant, Anti-Inflammatory, and Cytotoxic Activities of Propolis from the Stingless Bee Tetragonisca fiebrigi (Jataí). Evid. -Based Complement. Altern. Med. 2015, 2015, 296186. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.J.O.; Vasconcelos, C.C.; Garcia, J.B.S.; Pinheiro, M.S.D.; Pereira, F.A.N.; Camelo, D.D.S.; de Morais, S.V.; Freitas, J.R.B.; da Rocha, C.Q.; Ribeiro, M.N.D.S.; et al. Anti-Inflammatory and Antioxidant Activity of Pollen Extract Collected by Scaptotrigona affinis postica: In silico, in vitro, and in vivo Studies. Antioxidants 2020, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos, H.F.; Campos, J.F.; dos Santos, C.M.; Balestieri, J.B.P.; Silva, D.B.; Carollo, C.A.; Souza, K.D.P.; Estevinho, L.M.; Dos Santos, E.L. Chemical Profile and Antioxidant, Anti-Inflammatory, Antimutagenic and Antimicrobial Activities of Geopropolis from the Stingless Bee Melipona orbignyi. Int. J. Mol. Sci. 2017, 18, 953. [Google Scholar] [CrossRef] [Green Version]
- Martinello, M.; Mutinelli, F. Antioxidant Activity in Bee Products: A Review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Plutino, M.; Pignatti, G.; Karabagias, I.K.; Martinelli, E.; Souto, E.B.; Santini, A.; Lucini, L. Antioxidant Properties of Bee Products Derived from Medicinal Plants as Beekeeping Sources. Agriculture 2021, 11, 1136. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Concept and some Practical Aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 10 February 2023).
- Camargo, J.M.F.; Pedro, S.R.M.; Melo, G.A.R.; Meliponini Lepeletier. 1836. In Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region; Moure, J.S., Urban, D., Melo, G.A.R., (Orgs). 2013. Available online: http://www.moure.cria.org.br/catalogue (accessed on 10 February 2023).
- Camargo, J.M.F.; Moure, J.S. Meliponini neotropicais: O gênero Geotrigona Moure, 1943 (Apinae, Apidae, Hymenoptera), com especial referência à filogenia e biogeografia. Arq. Zool. 1996, 33, 95–161. [Google Scholar] [CrossRef]
- Roubik, D.W. Stingless bee nesting biology. Apidologie 2006, 37, 124–143. [Google Scholar] [CrossRef] [Green Version]
- Ishii, N.; Ishii, T.; Hartman, P.S. Oxidative Stress and C. elegans Models. In Aging Mechanisms; Mori, N., Mook-Jung, I., Eds.; Springer: Tokyo, Japan, 2015; pp. 111–122. [Google Scholar] [CrossRef]
- Alencar, S.; Oldoni, T.; Castro, M.; Cabral, I.; Costa-Neto, C.; Cury, J.; Rosalen, P.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 912–917. [Google Scholar] [CrossRef]
- Gupta, D.; Gupta, R.K. Bioprotective properties of Dragon’s blood resin: In vitro evaluation of antioxidant activity and antimicrobial activity. BMC Complement. Altern. Med. 2011, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Bonamigo, T.; Campos, J.F.; Alfredo, T.M.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Souza, K.D.P.; dos Santos, E.L. Antioxidant, Cytotoxic, and Toxic Activities of Propolis from Two Native Bees in Brazil: Scaptotrigona depilis and Melipona quadrifasciata anthidioides. Oxidative Med. Cell. Longev. 2017, 2017, 1038153. [Google Scholar] [CrossRef] [Green Version]
- Dengg, M.; van Meel, J.C. Caenorhabditis elegans as model system for rapid toxicity assessment of pharmaceutical compounds. J. Pharmacol. Toxicol. Methods 2004, 50, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Li, H.; Zhang, J.; Yang, F.; Huang, A.; Deng, J.; Liang, M.; Ma, F.; Hu, M.; Huang, Z. Salidroside Protects Caenorhabditis elegans Neurons from Polyglutamine-Mediated Toxicity by Reducing Oxidative Stress. Molecules 2014, 19, 7757–7769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [Green Version]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a Potent Antioxidant: Implications for Neurodegenerative Disorders. Oxidative Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta Rev. Cancer 2012, 1826, 443–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [Green Version]
- Caro, A.A.; Davis, A.; Fobare, S.; Horan, N.; Ryan, C.; Schwab, C. Antioxidant and pro-oxidant mechanisms of (+) catechin in microsomal CYP2E1-dependent oxidative stress. Toxicol. In Vitro 2019, 54, 1–9. [Google Scholar] [CrossRef]
- Lu, N.; Chen, P.; Yang, Q.; Peng, Y.-Y. Anti- and pro-oxidant effects of (+)-catechin on hemoglobin-induced protein oxidative damage. Toxicol. In Vitro 2011, 25, 833–838. [Google Scholar] [CrossRef]
- Park, H.-E.H.; Jung, Y.; Lee, S.-J.V. Survival assays using Caenorhabditis elegans. Mol. Cells 2017, 40, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Arriola, E.; Cárdenas-Rodríguez, N.; Coballase-Urrutia, E.; Pedraza-Chaverri, J.; Carmona-Aparicio, L.; Ortega-Cuellar, D. Caenorhabditis elegans: A Useful Model for Studying Metabolic Disorders in Which Oxidative Stress Is a Contributing Factor. Oxidative Med. Cell. Longev. 2014, 2014, 705253. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, L.M.; Machado, M.L.; da Silva, A.F.; Baptista, F.B.O.; da Silveira, T.L.; Soares, F.A.A.; Arantes, L.P. Rutin protects Huntington’s disease through the insulin/IGF1 (IIS) signaling pathway and autophagy activity: Study in Caenorhabditis elegans model. Food Chem. Toxicol. 2020, 141, 111323. [Google Scholar] [CrossRef]
- Cordeiro, L.M.; Soares, M.V.; da Silva, A.F.; Machado, M.L.; Baptista, F.B.O.; da Silveira, T.L.; Arantes, L.P.; Soares, F.A.A. Neuroprotective effects of rutin on ASH neurons in Caenorhabditis elegans model of Huntington’s disease. Nutr. Neurosci. 2022, 25, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Nkwonkam, C.G.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Investigations of protective effects of the flavonoids quercetin and rutin on stress resistance in the model organism Caenorhabditis elegans. Toxicology 2007, 234, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Kandemir, F.M.; Ozkaraca, M.; Yildirim, B.A.; Hanedan, B.; Kirbas, A.; Kilic, K.; Aktas, E.; Benzer, F. Rutin attenuates gentamicin-induced renal damage by reducing oxidative stress, inflammation, apoptosis, and autophagy in rats. Ren. Fail. 2015, 37, 518–525. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Protective effects of quercetin glycosides, rutin, and isoquercetrin against 6-hydroxydopamine (6-OHDA)-induced neurotoxicity in rat pheochromocytoma (PC-12) cells. Int. J. Immunopathol. Pharmacol. 2015, 29, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jattujan, P.; Srisirirung, S.; Watcharaporn, W.; Chumphoochai, K.; Kraokaew, P.; Sanguanphun, T.; Prasertsuksri, P.; Thongdechsri, S.; Sobhon, P.; Meemon, K. 2-Butoxytetrahydrofuran and Palmitic Acid from Holothuria scabra Enhance C. elegans Lifespan and Healthspan via DAF-16/FOXO and SKN-1/NRF2 Signaling Pathways. Pharmaceuticals 2022, 15, 1374. [Google Scholar] [CrossRef]
- Wei, C.-C.; Yen, P.-L.; Chang, S.-T.; Cheng, P.-L.; Lo, Y.-C.; Liao, V.H.-C. Antioxidative Activities of Both Oleic Acid and Camellia tenuifolia Seed Oil Are Regulated by the Transcription Factor DAF-16/FOXO in Caenorhabditis elegans. PLoS ONE 2016, 11, e0157195. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Compounds | EEC-G (mg/L) | EEC-T (mg/L) |
|---|---|---|
| Phenolic compounds | ||
| 3,4-hydroxycinnamic acid | 3.040 ± 0.032 | - |
| Catechin | 10.000 ± 0.044 | - |
| Gallic acid | - | 6.473 ± 0.020 |
| Rutin | - | 12.993 ± 0.022 |
| Vanillin | 4.020 ± 0.035 | - |
| Dicarboxylic acids | ||
| Maleic acid | 1.970 ± 0.023 | - |
| Fatty acids | ||
| Caprylic acid | 0.303 ± 0.003 | 0.347 ± 0.003 |
| Capric acid | 1.607 ± 0.007 | 1.503 ± 0.003 |
| Lauric acid | 2.590 ± 0.012 | 2.543 ± 0.009 |
| Myristic acid | 9.973 ± 0.035 | 10.583 ± 0.015 |
| Pentadecanoic acid | 0.103 ± 0.003 | 0.103 ± 0.003 |
| Palmitic acid | 37.193 ± 0.143 | 35.953 ± 0.046 |
| Margaric acid | 0.103 ± 0.003 | 0.107 ± 0.003 |
| Stearic acid | 10.983 ± 0.020 | 11.447 ± 0.009 |
| Arachidic acid | 3.347 ± 0.020 | 3.417 ± 0.018 |
| Behenic acid | 3.067 ± 0.023 | 2.987 ± 0.015 |
| Myristoleic acid | 7.447 ± 0.035 | 7.107 ± 0.020 |
| Palmitoleic acid | 8.683 ± 0.035 | 9.173 ± 0.032 |
| Oleic acid | 12.083 ± 0.071 | 12.027 ± 0.032 |
| Linoleic acid | 13.710 ± 0.059 | 13.523 ± 0.020 |
| Linolenic acid | 14.237 ± 0.041 | 14.023 ± 0.024 |
| Metallic minerals | ||
| Aluminum (Al) | 0.713 ± 0.012 | 0.593 ± 0.009 |
| Barium (Ba) | 0.263 ± 0.003 | 0.293 ± 0.009 |
| Calcium (Ca) | 1.510 ± 0.017 | 1.343 ± 0.231 |
| Copper (Cu) | 0.260 ± 0.006 | 0.290 ± 0.017 |
| Iron (Fe) | 0.713 ± 0.012 | 0.620 ± 0.015 |
| Potassium (K) | 1.623 ± 0.024 | 1.630 ± 0.035 |
| Magnesium (Mg) | 1.710 ± 0.015 | 1.630 ± 0.059 |
| Manganese (Mn) | 0.303 ± 0.007 | 0.303 ± 0.012 |
| Sodium (Na) | 0.310 ± 0.010 | 0.347 ± 0.015 |
| Zinc (Zn) | 0.227 ± 0.003 | 0.217 ± 0.003 |
| Non-metallic minerals | ||
| Phosphorus (P) | 0.777 ± 0.015 | 0.720 ± 0.015 |
| Sulfur (S) | 0.253 ± 0.003 | 0.267 ± 0.009 |
| Sample | DPPH• | ABTS•+ |
|---|---|---|
| IC50 (mg/mL) | IC50 (mg/mL) | |
| Ascorbic acid | 0.004 ± 0.00029 | 0.003 ± 0.00006 |
| BHT | 0.031 ± 0.005 | 0.009 ± 0.0009 |
| EEC-G | 1.001 ± 0.062 | 0.496 ± 0.040 |
| EEC-T | 1.251 ± 0.068 | 0.254 ± 0.023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, I.C.; Côrrea, R.C.D.; Orué, S.L.; Leite, D.F.; da Rocha, P.d.S.; Cardoso, C.A.L.; Mussury, R.M.; Vit, P.; de Picoli Souza, K.; dos Santos, E.L.; et al. Chemical Components and Antioxidant Activity of Geotrigona sp. and Tetragonisca fiebrigi Stingless Bee Cerumen Reduce Juglone-Induced Oxidative Stress in Caenorhabditis elegans. Antioxidants 2023, 12, 1276. https://doi.org/10.3390/antiox12061276
Ferreira IC, Côrrea RCD, Orué SL, Leite DF, da Rocha PdS, Cardoso CAL, Mussury RM, Vit P, de Picoli Souza K, dos Santos EL, et al. Chemical Components and Antioxidant Activity of Geotrigona sp. and Tetragonisca fiebrigi Stingless Bee Cerumen Reduce Juglone-Induced Oxidative Stress in Caenorhabditis elegans. Antioxidants. 2023; 12(6):1276. https://doi.org/10.3390/antiox12061276
Chicago/Turabian StyleFerreira, Isamara Carvalho, Raíssa Cristina Darroz Côrrea, Sarah Lam Orué, Daniel Ferreira Leite, Paola dos Santos da Rocha, Claudia Andrea Lima Cardoso, Rosilda Mara Mussury, Patricia Vit, Kely de Picoli Souza, Edson Lucas dos Santos, and et al. 2023. "Chemical Components and Antioxidant Activity of Geotrigona sp. and Tetragonisca fiebrigi Stingless Bee Cerumen Reduce Juglone-Induced Oxidative Stress in Caenorhabditis elegans" Antioxidants 12, no. 6: 1276. https://doi.org/10.3390/antiox12061276