
Carnosinase-1 Knock-Out Reduces Kidney Fibrosis in Type-1 Diabetic Mice on High Fat Diet
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
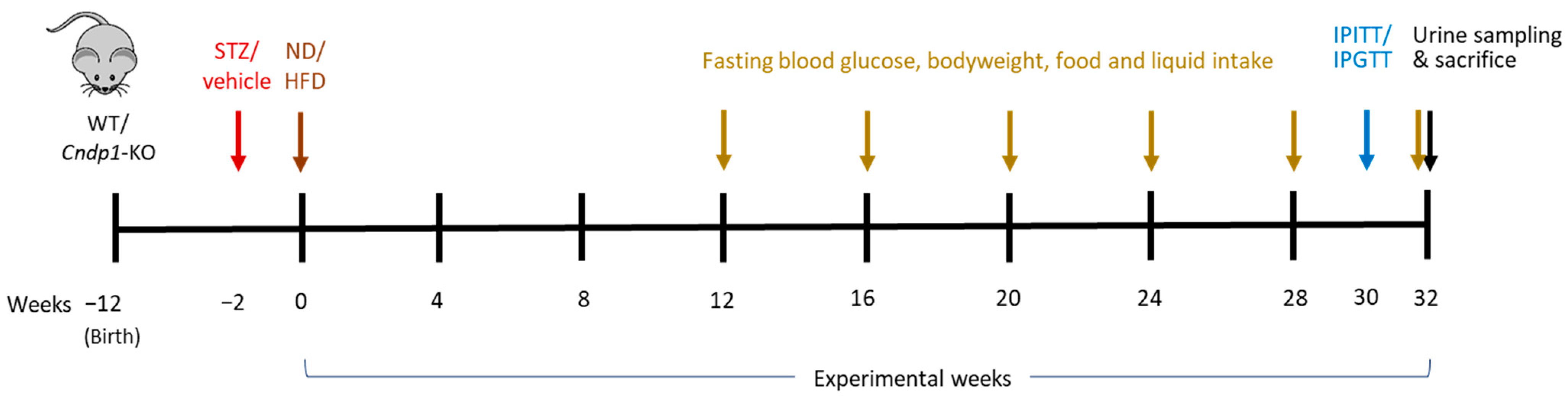
2.1. Animal Experiments
2.2. Quantitative Histopathology and Immunohistochemistry
2.3. Urine Albumin Quantification
2.4. Tissue RT-qPCR mRNA and Western Blotting
2.5. Tissue AGE and 4-HNE Quantification
2.6. Tissue Metabolomics
2.7. Statistical Analysis
3. Results
3.1. Body and Organ Weight
3.2. Glucose Homeostasis
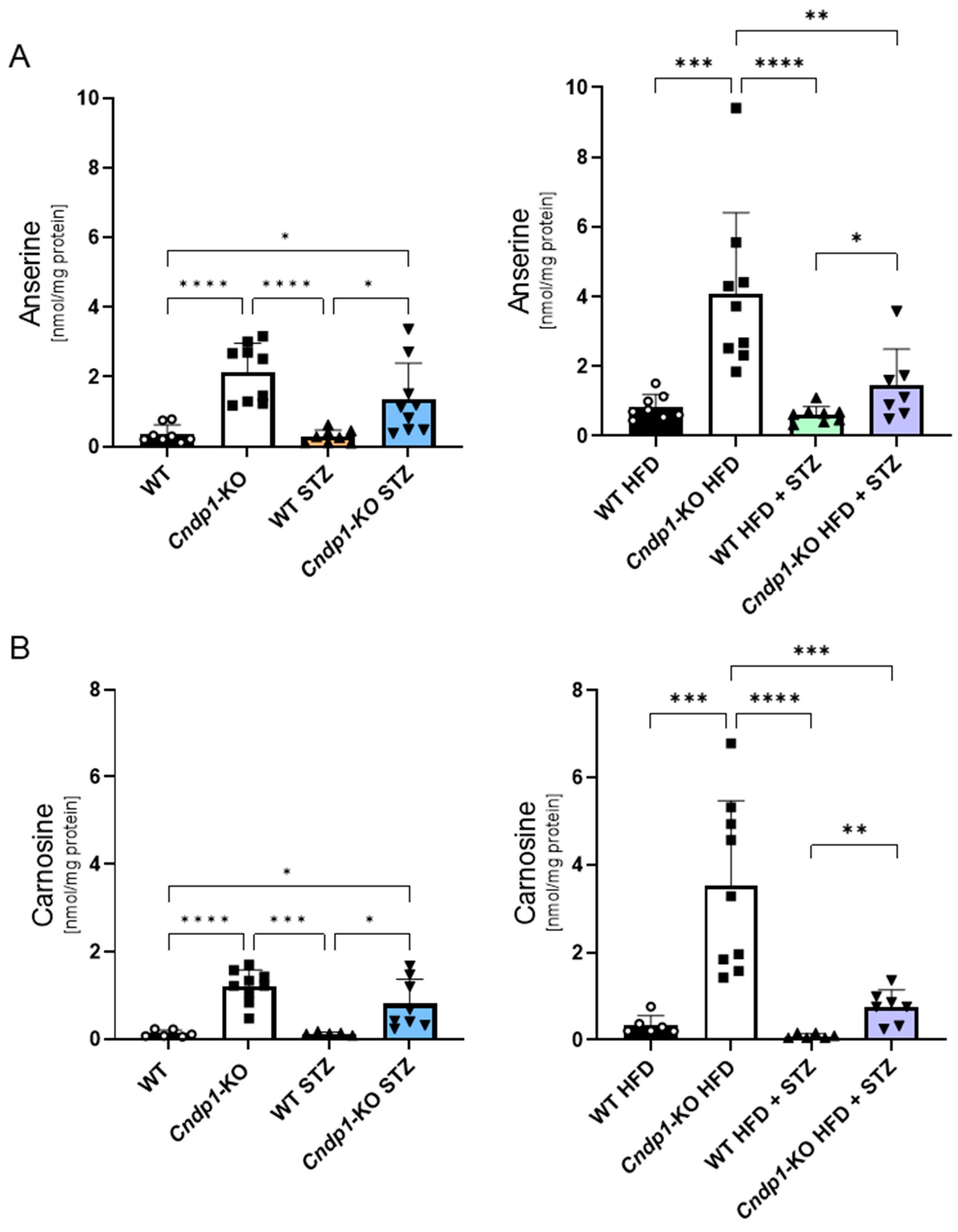
3.3. Tissue Anserine and Carnosine
3.4. Kidney Metabolome
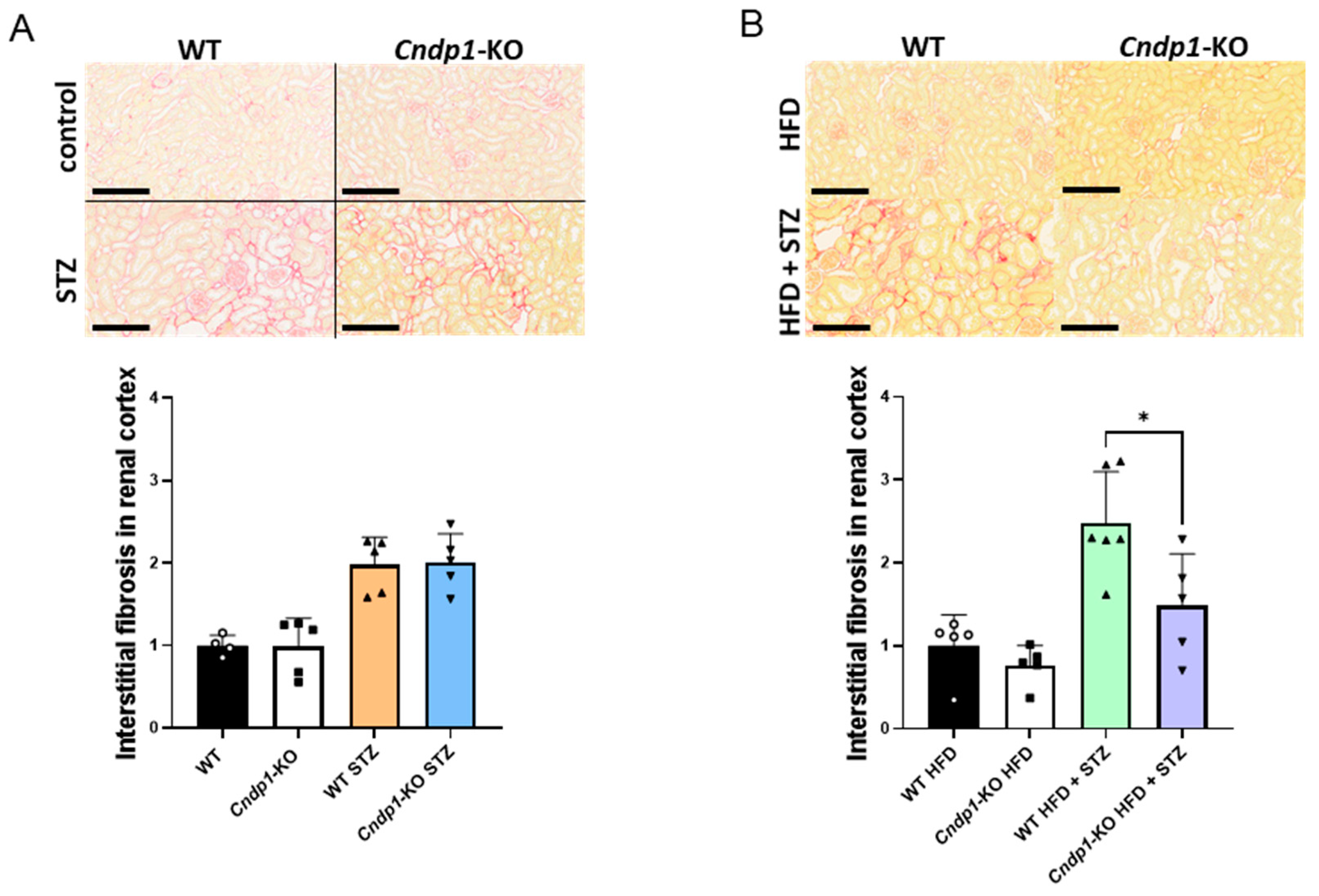
3.5. Kidney Morphology
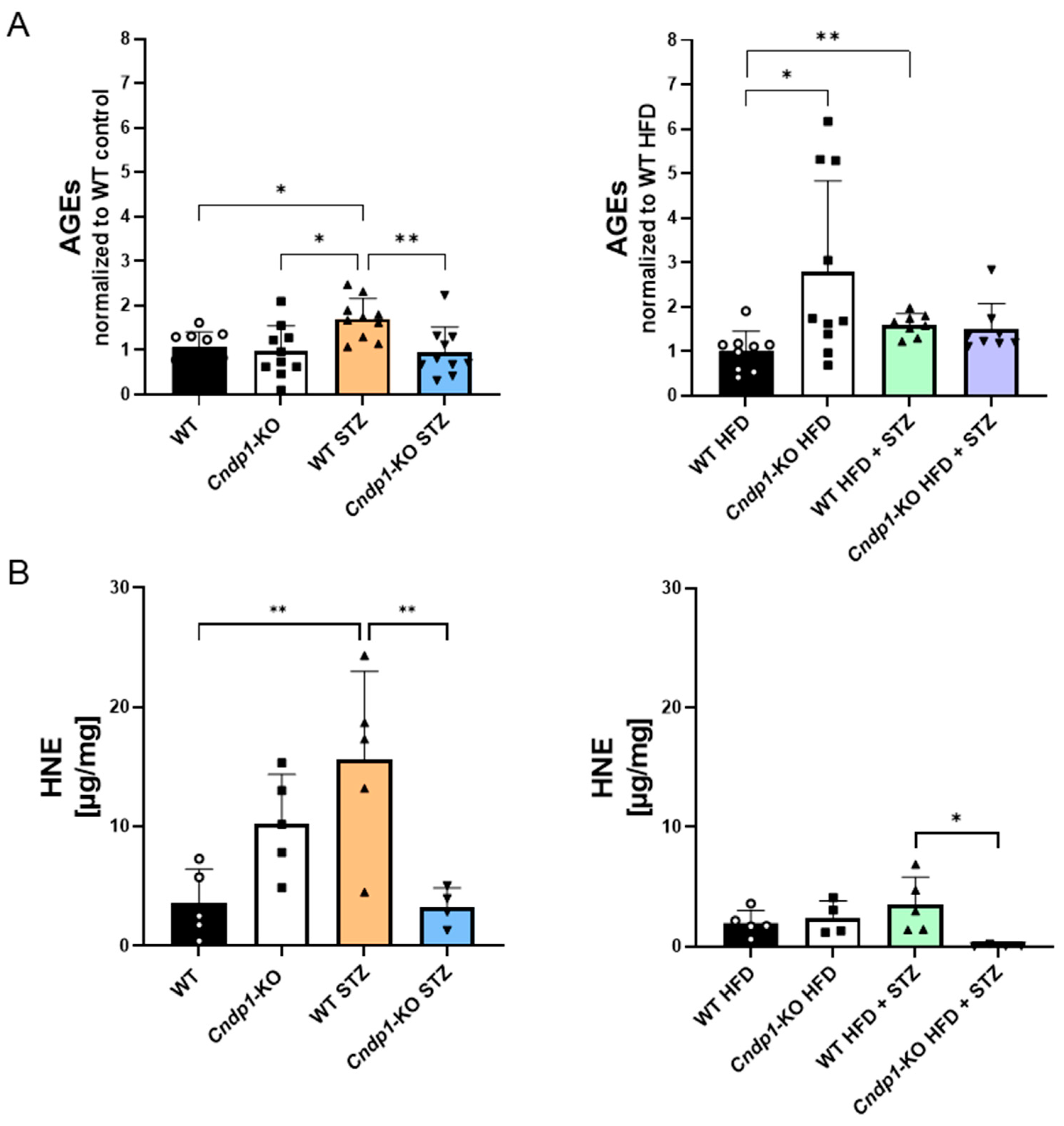
3.6. Kidney Stress Marker
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kilis-Pstrusinska, K. Carnosine and Kidney Diseases: What We Currently Know? Curr. Med. Chem. 2020, 27, 1764–1781. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Scuto, M.; Salinaro, A.T.; Dionisio, G.; Modafferi, S.; Ontario, M.L.; Greco, V.; Sciuto, S.; Schmitt, C.P.; Calabrese, E.J.; et al. Hydrogen Sulfide and Carnosine: Modulation of Oxidative Stress and Inflammation in Kidney and Brain Axis. Antioxidants 2020, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Solana-Manrique, C.; Sanz, F.J.; Martínez-Carrión, G.; Paricio, N. Antioxidant and Neuroprotective Effects of Carnosine: Therapeutic Implications in Neurodegenerative Diseases. Antioxidants 2022, 11, 848. [Google Scholar] [CrossRef] [PubMed]
- Schwank-Xu, C.; Forsberg, E.; Bentinger, M.; Zhao, A.; Ansurudeen, I.; Dallner, G.; Catrina, S.B.; Brismar, K.; Tekle, M. L-Carnosine Stimulation of Coenzyme Q10 Biosynthesis Promotes Improved Mitochondrial Function and Decreases Hepatic Steatosis in Diabetic Conditions. Antioxidants 2021, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’Amato, A.; Vistoli, G.; Carini, M. Understanding the antioxidant and carbonyl sequestering activity of carnosine: Direct and indirect mechanisms. Free. Radic. Res. 2021, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Vistoli, G.; Colzani, M.; Mazzolari, A.; Gilardoni, E.; Rivaletto, C.; Carini, M.; Aldini, G. Quenching activity of carnosine derivatives towards reactive carbonyl species: Focus on alpha-(methylglyoxal) and beta-(malondialdehyde) dicarbonyls. Biochem. Biophys. Res. Commun. 2017, 492, 487–492. [Google Scholar] [CrossRef]
- Anderson, E.J.; Vistoli, G.; Katunga, L.A.; Funai, K.; Regazzoni, L.; Monroe, T.B.; Gilardoni, E.; Cannizzaro, L.; Colzani, M.; De Maddis, D.; et al. A Carnosine Analog Mitigates Metabolic Disorders of Obesity by Reducing Carbonyl Stress. J. Clin. Investig. 2018, 128, 5280–5293. [Google Scholar] [CrossRef]
- Weigand, T.; Singler, B.; Fleming, T.; Nawroth, P.; Klika, K.D.; Thiel, C.; Baelde, H.; Garbade, S.F.; Wagner, A.H.; Hecker, M.; et al. Carnosine Catalyzes the Formation of the Oligo/Polymeric Products of Methylglyoxal. Cell. Physiol. Biochem. 2018, 46, 713–726. [Google Scholar] [CrossRef]
- Wetzel, C.; Pfeffer, T.; Bulkescher, R.; Zemva, J.; Modafferi, S.; Polimeni, A.; Salinaro, A.T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Anserine and Carnosine Induce HSP70-Dependent H2S Formation in Endothelial Cells and Murine Kidney. Antioxidants 2023, 12, 66. [Google Scholar] [CrossRef]
- Koppel, H.; RiedL, E.; Braunagel, M.; Sauerhoefer, S.; Ehnert, S.; Godoy, P.; Sternik, P.; Dooley, S.; Yard, B.A. L-carnosine inhibits high-glucose-mediated matrix accumulation in human mesangial cells by interfering with TGF-beta production and signalling. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2011, 26, 3852–3858. [Google Scholar]
- Scuto, M.; Trovato Salinaro, A.; Modafferi, S.; Polimeni, A.; Pfeffer, T.; Weigand, T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Carnosine Activates Cellular Stress Response in Podocytes and Reduces Glycative and Lipoperoxidative Stress. Biomedicines 2020, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, T.; Schilperoort, M.; Zhang, S.; Braun, J.D.; Qiu, J.; Rodriguez, A.; Pastene, D.O.; Kramer, B.K.; Koppel, H.; Baelde, H.; et al. Carnosine Attenuates the Development of both Type 2 Diabetes and Diabetic Nephropathy in BTBR ob/ob Mice. Sci. Rep. 2017, 7, 44492. [Google Scholar] [CrossRef] [PubMed]
- Iacobini, C.; Menini, S.; Blasetti Fantauzzi, C.; Pesce, C.M.; Giaccari, A.; Salomone, E.; Lapolla, A.; Orioli, M.; Aldini, G.; Pugliese, G. FL-926-16, a novel bioavailable carnosinase-resistant carnosine derivative, prevents onset and stops progression of diabetic nephropathy in db/db mice. Br. J. Pharm. 2018, 175, 53–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RiedL, E.; Pfister, F.; Braunagel, M.; Brinkkotter, P.; Sternik, P.; Deinzer, M.; Bakker, S.J.; Henning, R.H.; van den Born, J.; Kramer, B.K.; et al. Carnosine prevents apoptosis of glomerular cells and podocyte loss in STZ diabetic rats. Cell. Physiol. Biochem. 2011, 28, 279–288. [Google Scholar] [CrossRef]
- Rodriguez-Niño, A.; Pastene, D.O.; Hettler, S.A.; Qiu, J.; Albrecht, T.; Vajpayee, S.; Perciaccante, R.; Gretz, N.; Bakker, S.J.L.; Krämer, B.K.; et al. Influence of carnosine and carnosinase-1 on diabetes-induced afferent arteriole vasodilation: Implications for glomerular hemodynamics. Am. J. Physiol. Ren. Physiol. 2022, 323, F69–F80. [Google Scholar] [CrossRef]
- Iacobini, C.; Vitale, M.; Pugliese, G.; Menini, S. Normalizing HIF-1α Signaling Improves Cellular Glucose Metabolism and Blocks the Pathological Pathways of Hyperglycemic Damage. Biomedicines 2021, 9, 1139. [Google Scholar] [CrossRef]
- Lavilla, C.J.; Billacura, M.P.; Hanna, K.; Boocock, D.J.; Coveney, C.; Miles, A.K.; Foulds, G.A.; Murphy, A.; Tan, A.; Jackisch, L.; et al. Carnosine protects stimulus-secretion coupling through prevention of protein carbonyl adduction events in cells under metabolic stress. Free Radic. Biol. Med. 2021, 175, 65–79. [Google Scholar] [CrossRef]
- Matthews, J.J.; Turner, M.D.; Santos, L.; Elliott-Sale, K.J.; Sale, C. Carnosine increases insulin-stimulated glucose uptake and reduces methylglyoxal-modified proteins in type-2 diabetic human skeletal muscle cells. Amino Acids 2023, 55, 413–420. [Google Scholar] [CrossRef]
- Matthews, J.J.; Dolan, E.; Swinton, P.A.; Santos, L.; Artioli, G.G.; Turner, M.D.; Elliott-Sale, K.J.; Sale, C. Effect of Carnosine or β-Alanine Supplementation on Markers of Glycemic Control and Insulin Resistance in Humans and Animals: A Systematic Review and Meta-analysis. Adv. Nutr. 2021, 12, 2216–2231. [Google Scholar] [CrossRef]
- Menon, K.; Marquina, C.; Liew, D.; Mousa, A.; de Courten, B. Histidine-containing dipeptides reduce central obesity and improve glycaemic outcomes: A systematic review and meta-analysis of randomized controlled trials. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2020, 21, e12975. [Google Scholar] [CrossRef]
- Barca, A.; Gatti, F.; Spagnolo, D.; Ippati, S.; Vetrugno, C.; Verri, T. Responsiveness of Carnosine Homeostasis Genes in the Pancreas and Brain of Streptozotocin-Treated Mice Exposed to Dietary Carnosine. Int. J. Mol. Sci. 2018, 19, 1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, E.A.; Botusan, I.R.; Wang, J.; Peters, V.; Ansurudeen, I.; Brismar, K.; Catrina, S.B. Carnosine decreases IGFBP1 production in db/db mice through suppression of HIF-1. J. Endocrinol. 2015, 225, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6251–6531. [Google Scholar] [CrossRef] [Green Version]
- de Courten, B.; Jakubova, M.; de Courten, M.P.; Kukurova, I.J.; Vallova, S.; Krumpolec, P.; Valkovic, L.; Kurdiova, T.; Garzon, D.; Barbaresi, S.; et al. Effects of carnosine supplementation on glucose metabolism: Pilot clinical trial. Obesity 2016, 24, 1027–1034. [Google Scholar] [CrossRef]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatr. Diabetes 2018, 19, 470–477. [Google Scholar] [CrossRef]
- Siriwattanasit, N.; Satirapoj, B.; Supasyndh, O. Effect of Oral carnosine supplementation on urinary TGF-β in diabetic nephropathy: A randomized controlled trial. BMC Nephrol. 2021, 22, 236. [Google Scholar] [CrossRef]
- Qiu, J.; Yard, B.A.; Kramer, B.K.; van Goor, H.; van Dijk, P.; Kannt, A. Association Between Serum Carnosinase Concentration and Activity and Renal Function Impairment in a Type-2 Diabetes Cohort. Front. Pharm. 2022, 13, 899057. [Google Scholar] [CrossRef]
- Weigand, T.; Colbatzky, F.; Pfeffer, T.; Garbade, S.F.; Klingbeil, K.; Colbatzky, F.; Becker, M.; Zemva, J.; Bulkescher, R.; Schürfeld, R.; et al. A Global Cndp1-Knock-Out Selectively Increases Renal Carnosine and Anserine Concentrations in an Age- and Gender-Specific Manner in Mice. Int. J. Mol. Sci. 2020, 21, 4887. [Google Scholar] [CrossRef]
- Peters, V.; RiedL, E.; Braunagel, M.; Hoger, S.; Hauske, S.; Pfister, F.; Zschocke, J.; Lanthaler, B.; Benck, U.; Hammes, H.P.; et al. Carnosine treatment in combination with ACE inhibition in diabetic rats. Regul. Pept. 2014, 194–195, 36–40. [Google Scholar] [CrossRef]
- Peters, V.; Klessens, C.Q.; Baelde, H.J.; Singler, B.; Veraar, K.A.; Zutinic, A.; Drozak, J.; Zschocke, J.; Schmitt, C.P.; de Heer, E. Intrinsic carnosine metabolism in the human kidney. Amino Acids 2015, 47, 2541–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Hamada, C.; Nakayama, M.; Miyazaki, M.; Sherif, A.M.; Harada, T.; Hirano, H. Impact of uremia, diabetes, and peritoneal dialysis itself on the pathogenesis of peritoneal sclerosis: A quantitative study of peritoneal membrane morphology. Clin. J. Am. Soc. Nephrol. CJASN 2008, 3, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Weger, B.D.; Weger, M.; Gorling, B.; Schink, A.; Gobet, C.; Keime, C.; Poschet, G.; Jost, B.; Krone, N.; Hell, R.; et al. Extensive Regulation of Diurnal Transcription and Metabolism by Glucocorticoids. PLoS Genet. 2016, 12, e1006512. [Google Scholar] [CrossRef] [PubMed]
- Andresen, C.; Boch, T.; Gegner, H.M.; Mechtel, N.; Narr, A.; Birgin, E.; Rasbach, E.; Rahbari, N.; Trumpp, A.; Poschet, G.; et al. Comparison of extraction methods for intracellular metabolomics of human tissues. Front. Mol. Biosci. 2022, 9, 932261. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.N. Fast Stable Restricted Maximum Likelihood and Marginal Likelihood Estimation of Semiparametric Generalized Linear Models. J. R. Stat. Soc. Ser. B Stat. Methodol. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Peters, V.; Schmitt, C.P.; Zschocke, J.; Gross, M.L.; Brismar, K.; Forsberg, E. Carnosine treatment largely prevents alterations of renal carnosine metabolism in diabetic mice. Amino Acids 2012, 42, 2411–2416. [Google Scholar] [CrossRef]
- Oppermann, H.; Elsel, S.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Erythrocytes Prevent Degradation of Carnosine by Human Serum Carnosinase. Int. J. Mol. Sci. 2021, 22, 12802. [Google Scholar] [CrossRef]
- Braga, P.C.; Alves, M.G.; Rodrigues, A.S.; Oliveira, P.F. Mitochondrial Pathophysiology on Chronic Kidney Disease. Int. J. Mol. Sci. 2022, 23, 1776. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Diet | Non-Diabetic | Diabetic | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| nmol/mg Protein | WT | Cndp1-KO | p (vs. WT) | WT STZ | p (vs. WT) | Cndp1-KO STZ | p (vs. WT STZ) | |||
| Kidney # | Anserine | 0.36 ± 0.26 | 2.13 ± 0.83 | <0.001 * | 0.28 ± 0.19 | 0.996 | 1.35 ± 1.05 | 0.02 * | ||
| Carnosine | 0.12 ± 0.08 | 1.20 ± 0.38 | <0.001 * | 0.12 ± 0.03 | >0.999 | 0.80 ± 0.57 | 0.021 * | |||
| Heart | Anserine | 0.13 ± 0.09 | 0.11 ± 0.07 | 0.914 | 0.13 ± 0.04 | 0.913 | 0.22 ± 0.06 | 0.075 | ||
| Carnosine | 0.18 ± 0.11 | 0.15 ± 0.09 | 0.914 | 0.32 ± 0.19 | 0.336 | 0.57 ± 0.28 | 0.135 | |||
| Muscle | Anserine | 22.7 ± 8.0 | 18.2 ± 8.8 | 0.577 | 15.3 ± 8.8 | 0.265 | 17.0 ± 4.9 | 0.689 | ||
| Carnosine | 9.73 ± 3.82 | 7.71 ± 3.64 | 0.577 | 6.86 ± 3.64 | 0.265 | 8.08 ± 2.02 | 0.689 | |||
| Liver | Anserine | 0.08 ± 0.02 | 0.08 ± 0.02 | 0.884 | n.a. | n.a. | n.a. | n.a. | ||
| Carnosine | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |||
| Serum | Anserine | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.261 | 0.01 ± 0.00 | 0.416 | 0.02 ± 0.02 | 0.255 | ||
| Carnosine | 0.04 ± 0.01 | 0.05 ± 0.04 | 0.53 | 0.02 ± 0.01 | 0.126 | 0.06 ± 0.05 | 0.255 | |||
| High Fat Diet | Non-Diabetic | Diabetic | ||||||||
| nmol/mg Protein | WT HFD | Cndp1-KO HFD | p (vs. WT HFD) | p (vs. Cndp1-KO) | WT HFD STZ | p (vs. WT HFD) | Cndp1-KO HFD STZ | p (vs. WT STZ HFD) | p (vs. Cndp1-KO STZ) | |
| Kidney # | Anserine | 0.84 ± 0.36 | 4.08 ± 2.33 | 0.001 * | 0.007 * | 0.61 ± 0.24 | 0.987 | 1.4 ± 1.1 | 0.046 * ## | >0.999 |
| Carnosine | 0.34 ± 0.22 | 3.52 ± 1.95 | 0.001 * | <0.001 * | 0.09 ± 0.05 | 0.981 | 0.8 ± 0.38 | 0.002 * ## | >0.999 | |
| Heart | Anserine | 0.10 ± 0.02 | 0.13 ± 0.04 | 0.433 | 0.891 | 0.13 ± 0.04 | 0.368 | 0.10 ± 0.05 | 0.301 | 0.016 * |
| Carnosine | 0.17 ± 0.08 | 0.15 ± 0.04 | 0.774 | 0.966 | 0.23 ± 0.10 | 0.368 | 0.14 ± 0.05 | 0.301 | 0.021 * | |
| Muscle | Anserine | 20.4 ± 13.3 | 18.5 ± 5.8 | 0.936 | 0.999 | 14.9 ± 5.2 | 0.597 | 19.4 ± 8.2 | 0.481 | 0.799 |
| Carnosine | 8.13 ± 5.04 | 7.7 ± 2.3 | 0.936 | 0.999 | 6.1 ± 2.1 | 0.597 | 7.9 ± 3.6 | 0.481 | 0.895 | |
| Liver | Anserine | 0.08 ± 0.01 | 0.09 ± 0.04 | 0.814 | 0.542 | n.a. | n.a. | n.a. | n.a. | n.a. |
| Carnosine | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | |
| Serum | Anserine | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.204 | 0.847 | 0.02 ± 0.02 | 0.578 | 0.02 ± 0.02 | 0.702 | 0.797 |
| Carnosine | 0.04 ± 0.03 | 0.04 ± 0.02 | 0.795 | 0.847 | 0.06 ± 0.03 | 0.449 | 0.04 ± 0.02 | 0.534 | 0.733 | |
| Normal Diet | Non-Diabetic | Diabetic | |||||
|---|---|---|---|---|---|---|---|
| WT | Cndp1-KO | p (vs. WT) | WT STZ | p (vs. WT) | Cndp1-KO STZ | p (vs. WT STZ) | |
| Diabetic lesions | 0.2 ± 0.45 | 0 ± 0 | 0.882 | 1 ± 0 | 0.019 * | 1 ± 0 | 0.999 |
| Arterial lesions | 0 ± 0 | 0 ± 0 | 0.999 | 0 ± 0 | 0.999 | 0 ± 0 | 0.999 |
| Proteinaceous casts | 0.6 ± 0.55 | 0.5 ± 0.58 | 0.983 | 1.75 ± 0.5 | 0.001 * | 1 ± 0 | 0.032 |
| Chronical interstitial inflammation | 0.6 ± 0.55 | 0.5 ± 0.58 | 0.456 | 1.25 ± 0.5 | 0.082 | 1.4 ± 0.55 | 0.946 |
| Tubular dilation | 0 ± 0 | 0.25 ± 0.5 | 0.794 | 1 ± 0.82 | 0.002 * | 1.6 ± 0.55 | 0.125 |
| Tubular fibrosis and atrophy | 0 ± 0 | 0.25 ± 0.5 | 0.794 | 2 ± 0 | <0.001 * | 2 ± 0 | 0.999 |
| Interstitial fibrosis # | 1 ± 0.13 | 0.99 ± 0.35 | 0.999 | 1.98 ± 0.34 | 0.001 * | 2.02 ± 0.35 | 0.999 |
| Fibrosis of Bowman´s capsule | 0 ± 0 | 0 ± 0 | 0.999 | 1 ± 0 | 0.002 * | 0.8 ± 0.45 | 0.882 |
| Cystic glomeruli | 0 ± 0 | 0 ± 0 | 0.999 | 0 ± 0 | 0.999 | 0.6 ± 0.9 | 0.125 |
| Glomerular hyalinosis | 0.2 ± 0.45 | 0 ± 0 | 0.882 | 0.25 ± 0.5 | 0.998 | 0 ± 0 | 0.794 |
| Mesangial sclerosis | 0.2 ± 0.45 | 0 ± 0 | 0.884 | 1.5 ± 0.58 | <0.001 * | 1.4 ± 0.55 | 0.983 |
| Mesangial expansion # | 0.39 ± 0.02 | 0.49 ± 0.09 | 0.011 * | 0.47 ± 0.04 | 0.039 * | 0.5 ± 0.07 | 0.814 |
| Glomerular size # | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.823 | 0.07 ± 0.01 | 0.998 | 0.07 ± 0.01 | 0.789 |
| High Fat Diet | Non-Diabetic | Diabetic | |||||
| WT HFD | Cndp1-KO HFD | p (vs. WT HFD) | WT HFD STZ | p (vs. WT HFD) | Cndp1-KO HFD + STZ | p (vs. WT HFD STZ) | |
| Diabetic lesions | n.a. | n.a. | n.a. | 1 ± 0 | n.a. | 1 ± 0 | 0.999 |
| Arterial lesions | n.a. | n.a. | n.a. | 0 ± 0 | n.a. | 0 ± 0 | 0.999 |
| Proteinaceous casts | n.a. | n.a. | n.a. | 1.8 ± 0.45 | n.a. | 1 ± 0 | 0.004 |
| Chronic interstitial inflammation | n.a. | n.a. | n.a. | 2 ± 0.36 | n.a. | 1 ± 0.71 | 0.022 |
| Tubular dilation | n.a. | n.a. | n.a. | 2.8 ± 0.45 | n.a. | 2 ± 0.71 | 0.065 |
| Tubular fibrosis and atrophy | n.a. | n.a. | n.a. | 2 ± 0 | n.a. | 1.6 ± 0.55 | 0.141 |
| Interstitial fibrosis # | 1 ± 0.37 | 0.77 ± 0.24 | 0.8732 | 2.48 ± 0.62 | 0.001 | 1.49 ± 0.63 | 0.020 |
| Fibrosis of Bowman´s capsule | n.a. | n.a. | n.a. | 1 ± 0 | n.a. | 0.8 ± 0.45 | 0.347 |
| Cystic glomeruli | n.a. | n.a. | n.a. | 0.2 ± 0.45 | n.a. | 0 ± 0 | 0.347 |
| Glomerular hyalinosis | n.a. | n.a. | n.a. | 0 ± 0 | n.a. | 0.4 ± 0.55 | 0.141 |
| Mesangial sclerosis | n.a. | n.a. | n.a. | 1.2 ± 0.84 | n.a. | 1.6 ± 0.9 | 0.486 |
| Mesangial expansion # | 0.63 ± 0.06 | 0.59 ± 0.06 | 0.6202 | 0.41 ± 0.05 | <0.001 | 0.44 ± 0.09 | 0.625 |
| Glomerular size # | 0.08 ± 0.01 | 0.08 ± 0.01 | 0.9999 | 0.07 ± 0.01 | <0.001 | 0.07 ± 0.01 | 0.890 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeffer, T.; Wetzel, C.; Kirschner, P.; Bartosova, M.; Poth, T.; Schwab, C.; Poschet, G.; Zemva, J.; Bulkescher, R.; Damgov, I.; et al. Carnosinase-1 Knock-Out Reduces Kidney Fibrosis in Type-1 Diabetic Mice on High Fat Diet. Antioxidants 2023, 12, 1270. https://doi.org/10.3390/antiox12061270
Pfeffer T, Wetzel C, Kirschner P, Bartosova M, Poth T, Schwab C, Poschet G, Zemva J, Bulkescher R, Damgov I, et al. Carnosinase-1 Knock-Out Reduces Kidney Fibrosis in Type-1 Diabetic Mice on High Fat Diet. Antioxidants. 2023; 12(6):1270. https://doi.org/10.3390/antiox12061270
Chicago/Turabian StylePfeffer, Tilman, Charlotte Wetzel, Philip Kirschner, Maria Bartosova, Tanja Poth, Constantin Schwab, Gernot Poschet, Johanna Zemva, Ruben Bulkescher, Ivan Damgov, and et al. 2023. "Carnosinase-1 Knock-Out Reduces Kidney Fibrosis in Type-1 Diabetic Mice on High Fat Diet" Antioxidants 12, no. 6: 1270. https://doi.org/10.3390/antiox12061270