
Veronica persica Ethanol Extract Ameliorates Dinitrochlorobenzene-Induced Atopic Dermatitis-like Skin Inflammation in Mice, Likely by Inducing Nrf2/HO-1 Signaling
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Animal Experiments
2.3. Analysis of Serum Ig E, Histamine, and Immune Cytokines
2.4. Histological Examination of the AD-like Dermis
2.5. Flow Cytometry Analysis
2.6. Splenocyte Isolation and Cytokine Measurement
2.7. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction
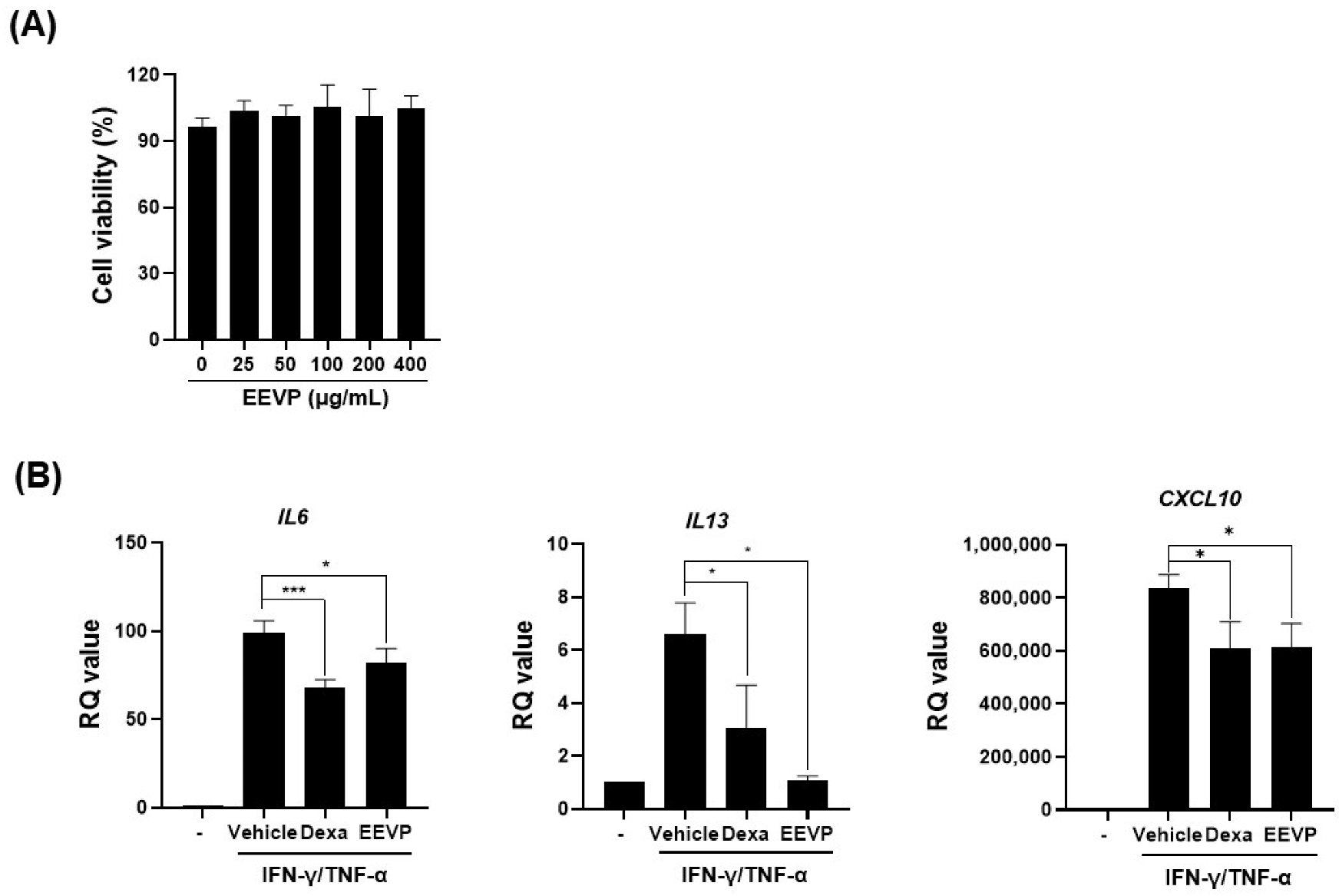
2.8. Cell Culture and Cell Viability Assay
2.9. Western Blot Analysis
2.10. Docking Analysis of The Kelch-Like ECH-Associated Protein (KEAP)-1 Kelch Domain Pocket and EEVP Components
2.11. Statistical Analysis
3. Results
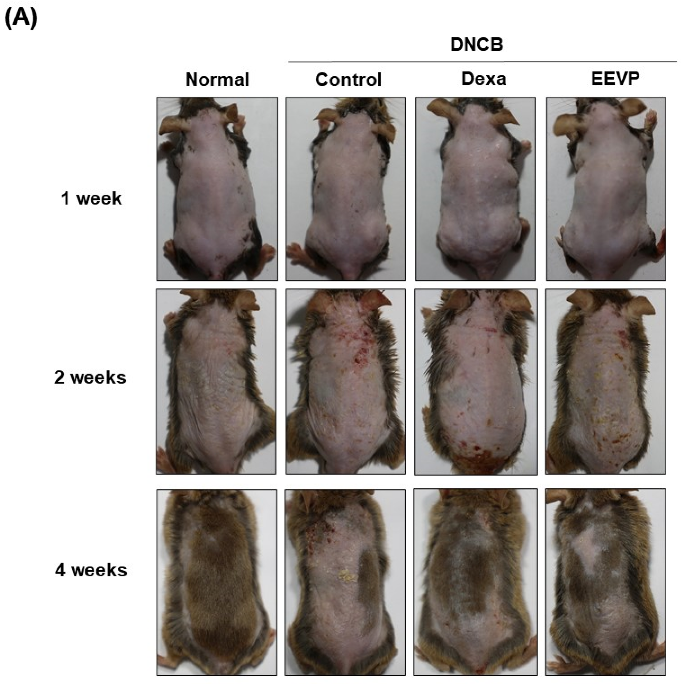
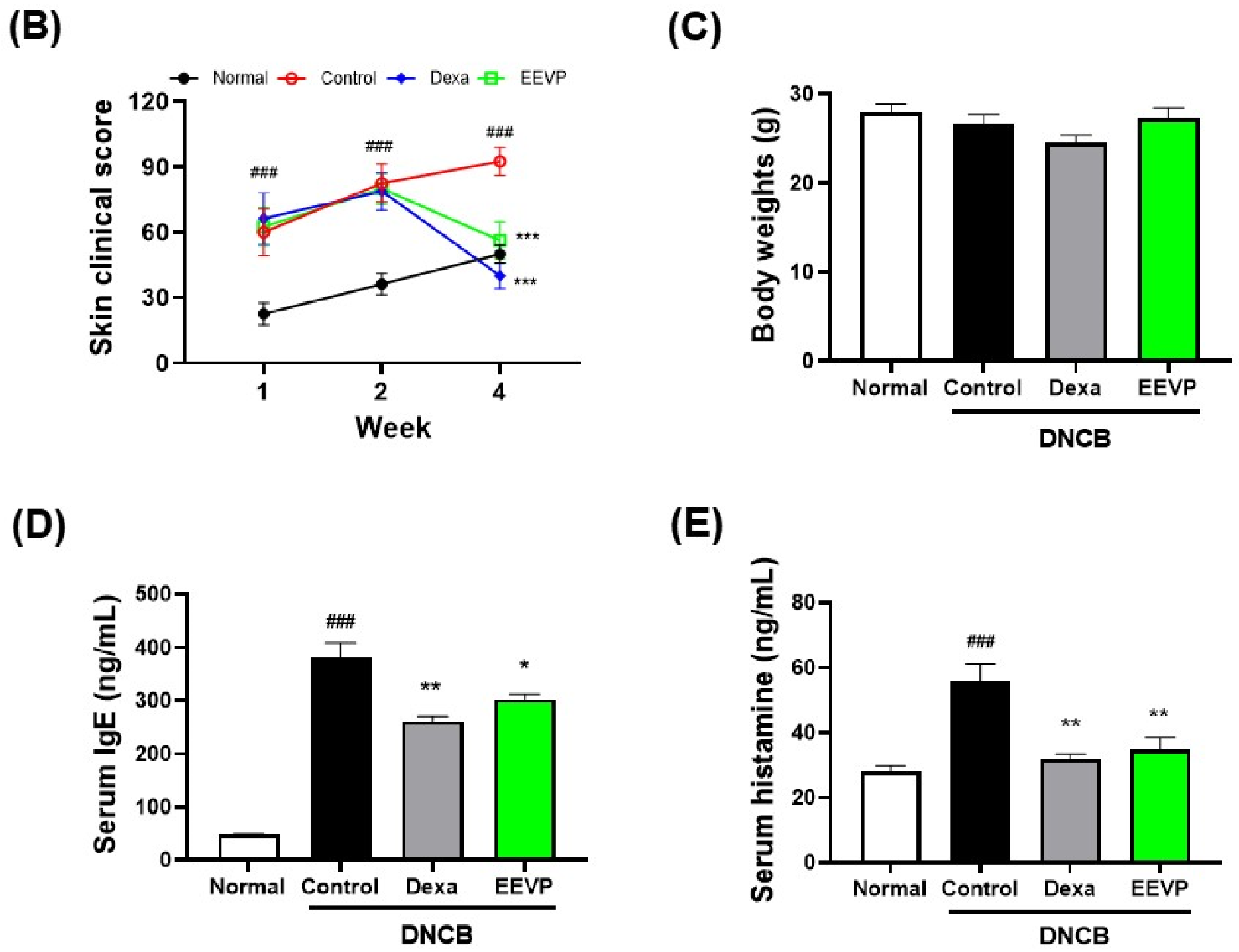
3.1. Effects of EEVP on DNCB-Induced IgE and Histamine Levels in Serum and AD-like Skin Lesions
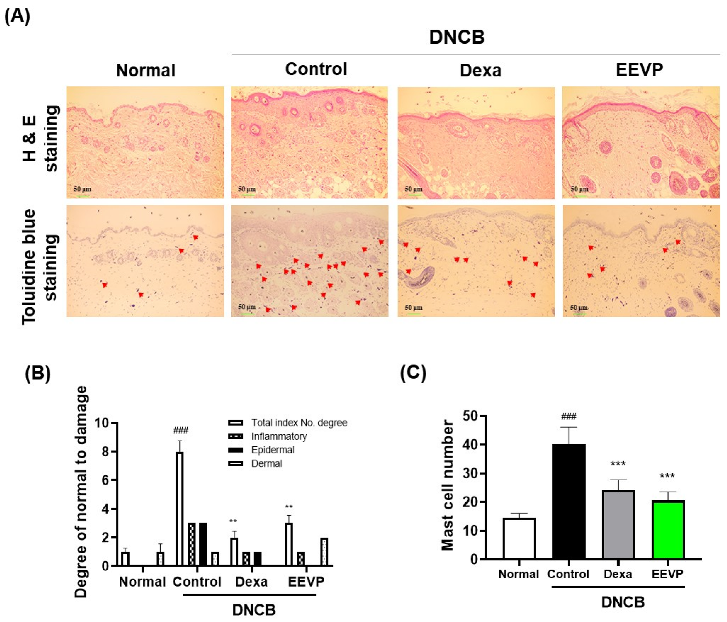
3.2. Effects of EEVP on DNCB-Induced Histological Changes in Dorsal Skin
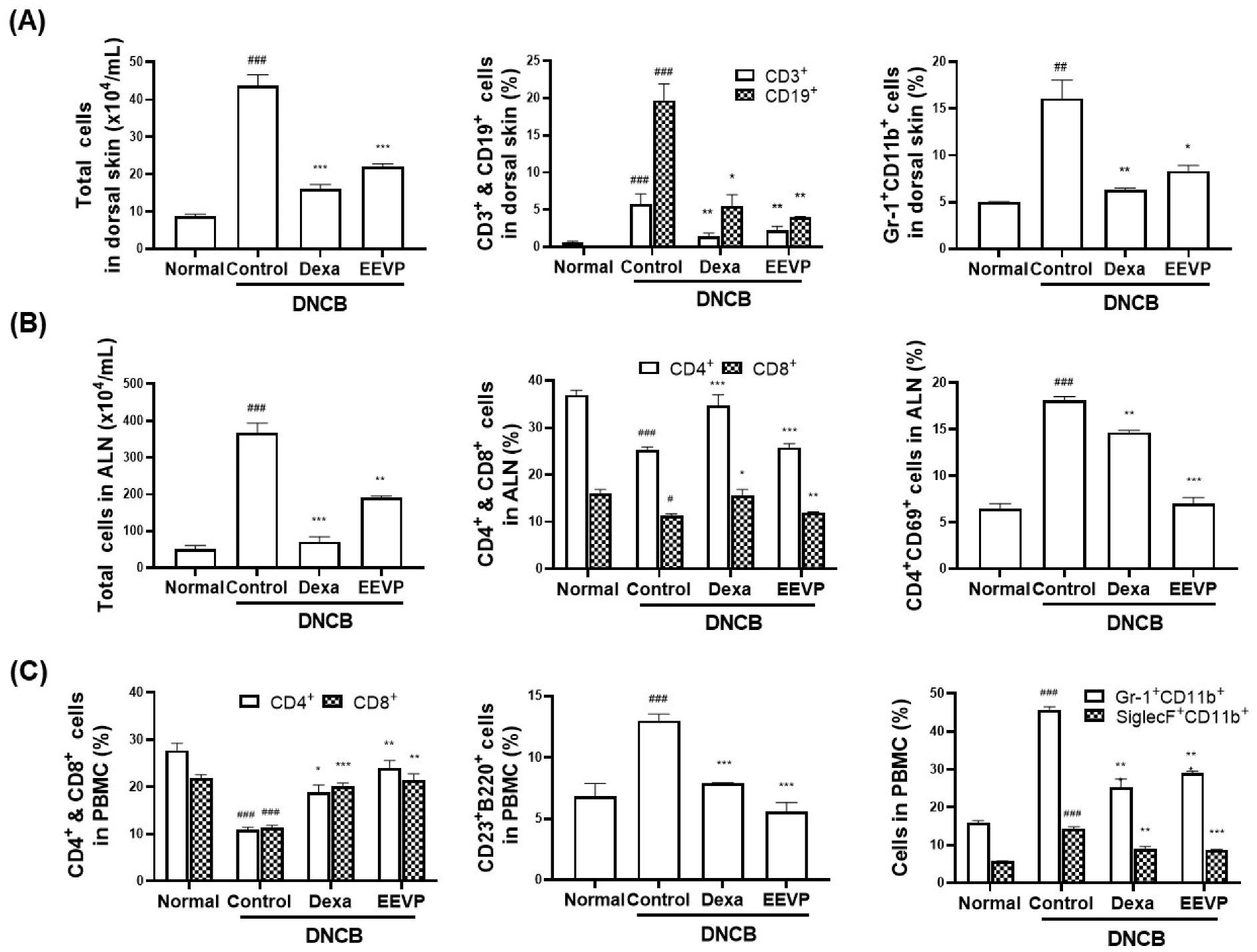
3.3. Effects of EEVP on Immune Cell Subtypes in PBMCs, Dorsal Skin, and ALNs
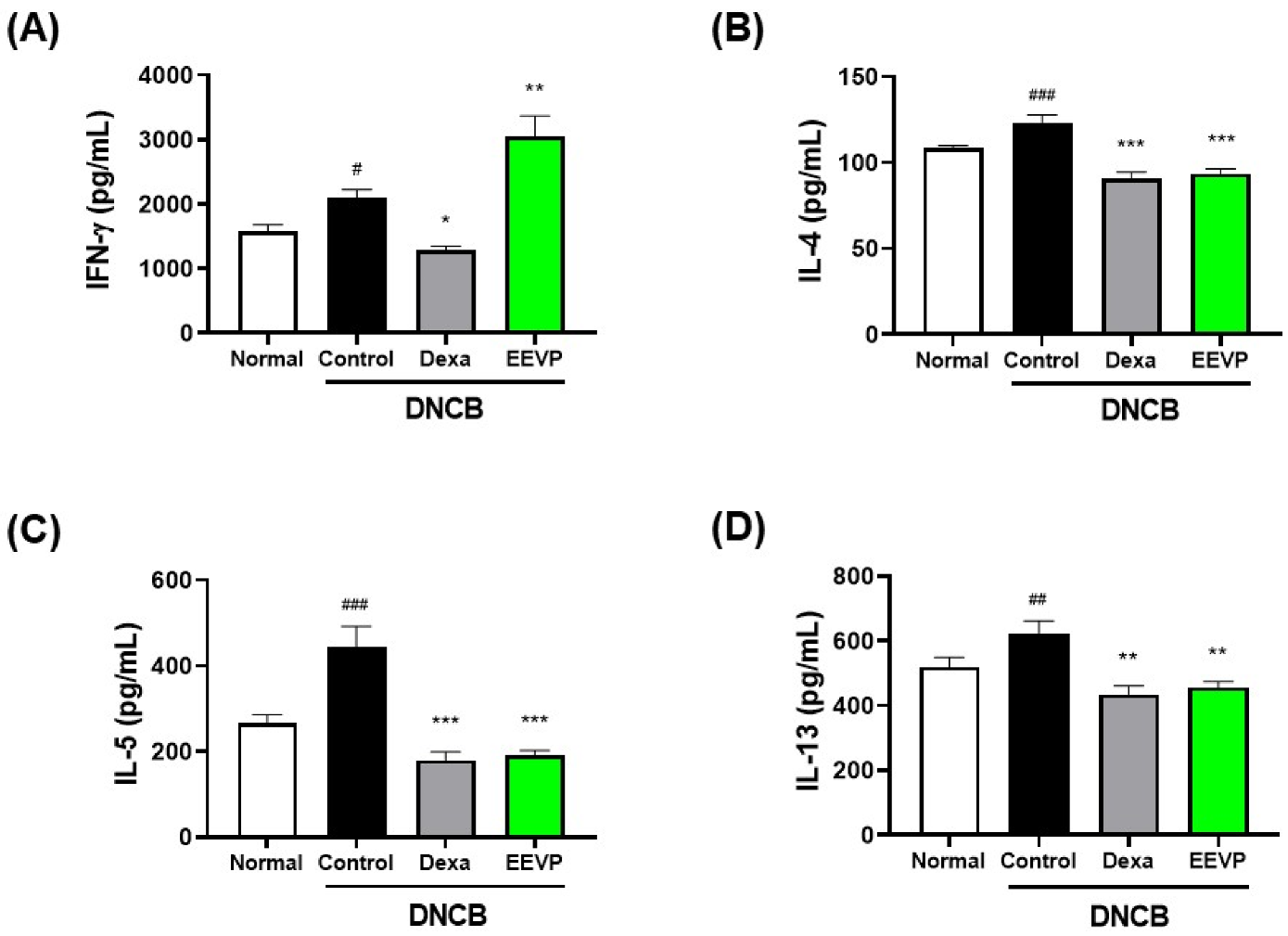
3.4. Effects of EEVP on Th1 and Th2 Cytokine Levels in Splenocyte Culture Medium and Dorsal Skin
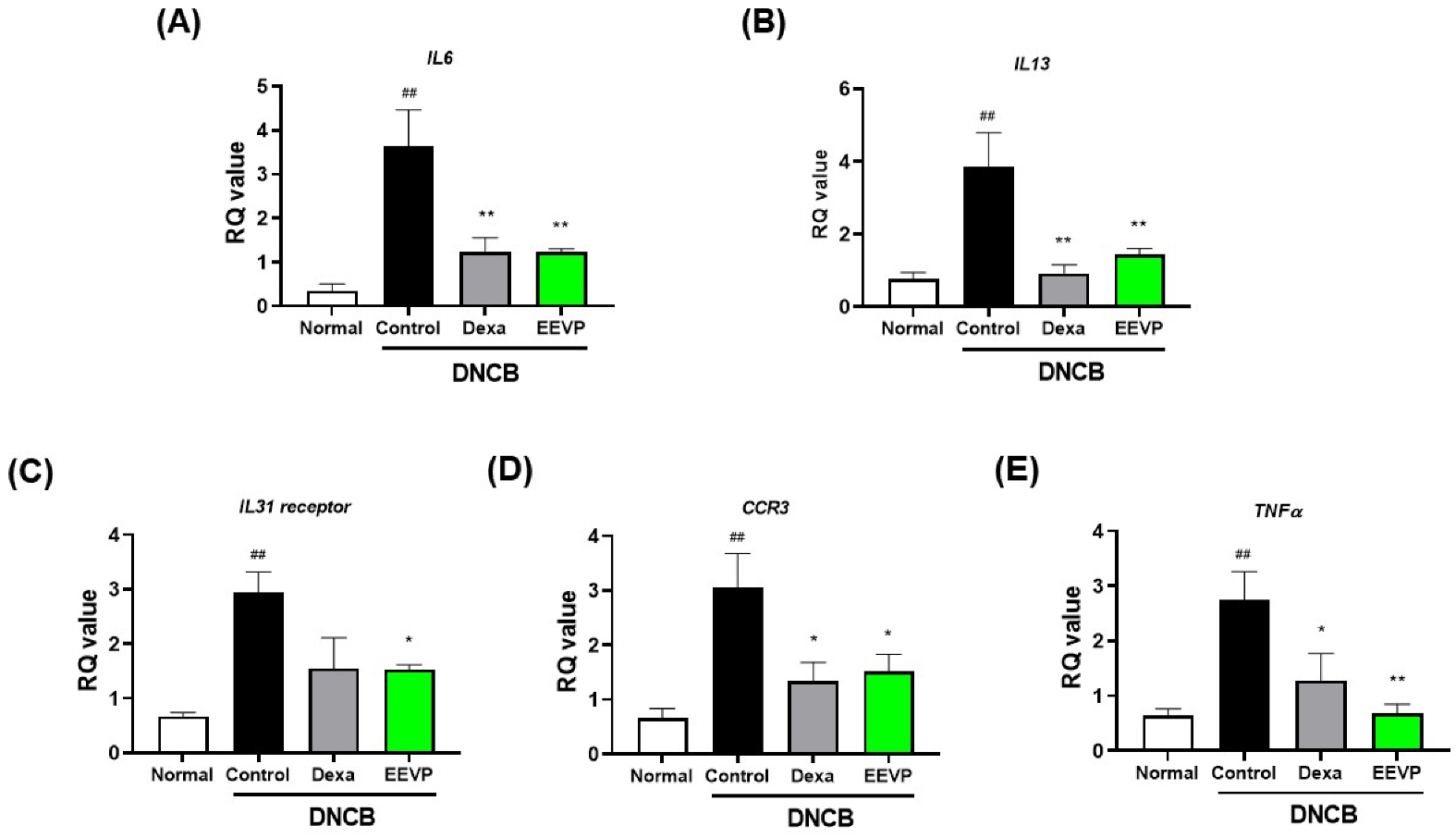
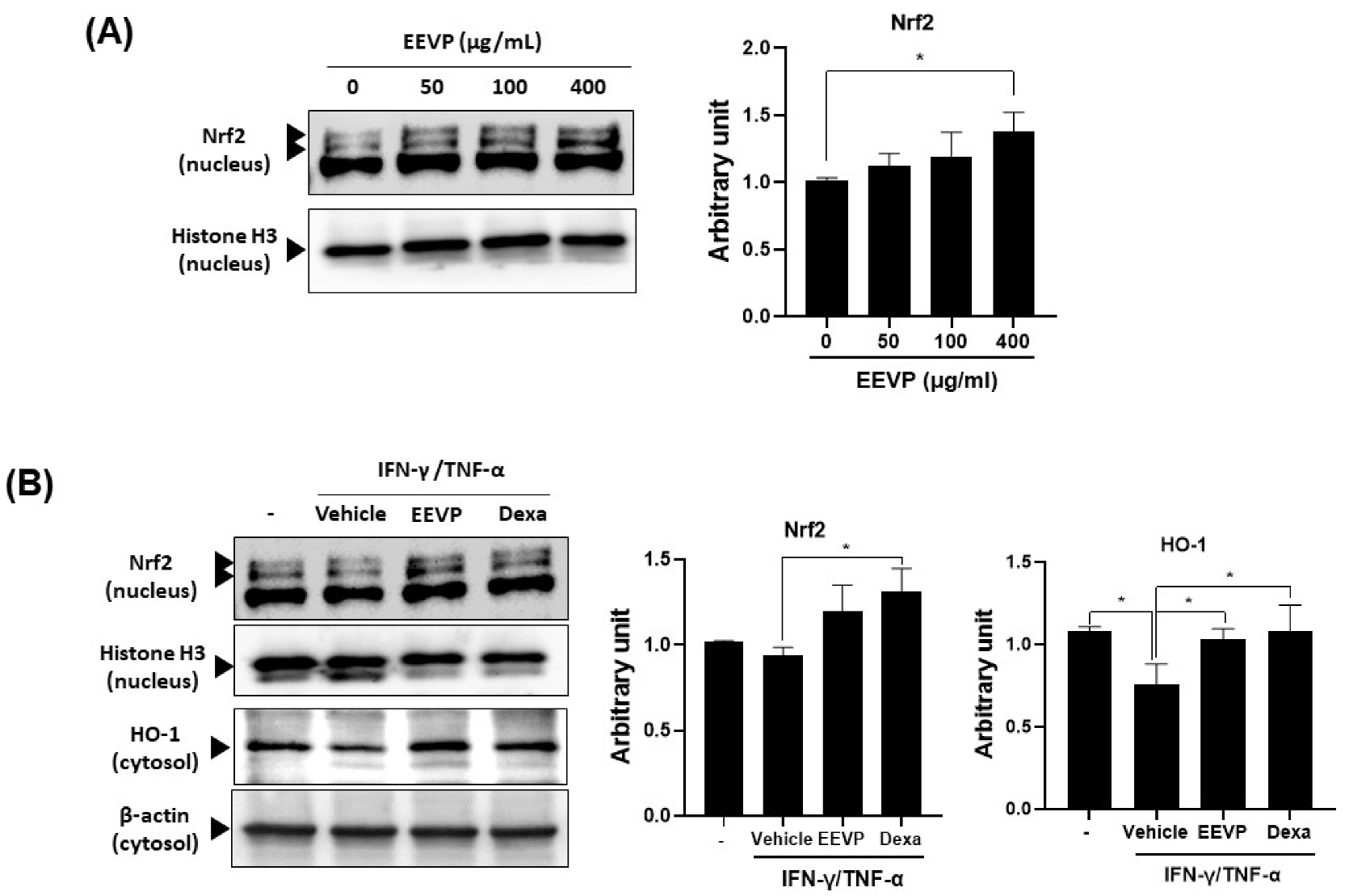
3.5. Effects of EEVP on Expression of Inflammatory Genes and the Nrf2/HO-1 Pathway in HaCaT Keratinocytes
3.6. Docking Analysis of Five Components in EEVP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bieber, T. Atopic dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, N.B.; Silverberg, J.I. Inside out or outside in: Does atopic dermatitis disrupt barrier function or does disruption of barrier function trigger atopic dermatitis? Cutis 2015, 96, 359–361. [Google Scholar] [PubMed]
- Gittler, J.K.; Shemer, A.; Suarez-Farinas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell. Immunol. 2011, 2, 110. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Torres, T. More than skin deep: The systemic nature of atopic dermatitis. Eur. J. Dermatol. 2019, 29, 250–258. [Google Scholar]
- Leung, D.Y.; Guttman-Yassky, E. Deciphering the complexities of atopic dermatitis: Shifting paradigms in treatment approaches. J. Allergy Clin. Immunol. 2014, 134, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.V. Treatment options for moderate to severe atopic dermatitis. Allergy Asthma Proc. 2022, 43, 474–493. [Google Scholar] [CrossRef]
- Saeki, H.; Ohya, Y.; Furuta, J.; Arakawa, H.; Ichiyama, S.; Katsunuma, T.; Katoh, N.; Tanaka, A.; Tsunemi, Y.; Nakahara, T.; et al. English Version of Clinical Practice Guidelines for the Management of Atopic Dermatitis 2021. J. Dermatol. 2022, 49, e315–e375. [Google Scholar] [CrossRef]
- Simpson, E.L.; Bieber, T.; Guttman-Yassky, E.; Beck, L.A.; Blauvelt, A.; Cork, M.J.; Silverberg, J.I.; Deleuran, M.; Kataoka, Y.; Lacour, J.P.; et al. Two Phase 3 Trials of Dupilumab versus Placebo in Atopic Dermatitis. N. Engl. J. Med. 2016, 375, 2335–2348. [Google Scholar] [CrossRef]
- Napolitano, M.; Fabbrocini, G.; Genco, L.; Martora, F.; Potestio, L.; Patruno, C. Rapid improvement in pruritus in atopic dermatitis patients treated with upadacitinib: A real-life experience. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1497–1498. [Google Scholar] [CrossRef]
- Gambardella, A.; Licata, G.; Calabrese, G.; De Rosa, A.; Alfano, R.; Argenziano, G. Dual Efficacy of Upadacitinib in 2 Patients with Concomitant Severe Atopic Dermatitis and Alopecia Areata. Dermatitis 2021, 32, e85–e86. [Google Scholar] [CrossRef]
- Park, B.K.; Park, Y.C.; Jung, I.C.; Kim, S.H.; Choi, J.J.; Do, M.; Kim, S.Y.; Jin, M. Gamisasangja-tang suppresses pruritus and atopic skin inflammation in the NC/Nga murine model of atopic dermatitis. J. Ethnopharmacol. 2015, 165, 54–60. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Yuk, H.J.; Yang, W.K.; Kim, S.H.; Kim, D.S. Siraitia grosvenorii Residual Extract Attenuates Atopic Dermatitis by Regulating Immune Dysfunction and Skin Barrier Abnormality. Nutrients 2020, 12, 3638. [Google Scholar] [CrossRef]
- Schafer, M.; Werner, S. Nrf2—A regulator of keratinocyte redox signaling. Free Radic. Biol. Med. 2015, 88, 243–252. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Chaitidis, P.; O’Donnell, V.; Kuban, R.J.; Bermudez-Fajardo, A.; Ungethuem, U.; Kuhn, H. Gene expression alterations of human peripheral blood monocytes induced by medium-term treatment with the TH2-cytokines interleukin-4 and -13. Cytokine 2005, 30, 366–377. [Google Scholar] [CrossRef]
- Lee, T.S.; Chau, L.Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef]
- Kirino, M.; Kirino, Y.; Takeno, M.; Nagashima, Y.; Takahashi, K.; Kobayashi, M.; Murakami, S.; Hirasawa, T.; Ueda, A.; Aihara, M.; et al. Heme oxygenase 1 attenuates the development of atopic dermatitis-like lesions in mice: Implications for human disease. J. Allergy Clin. Immunol. 2008, 122, 290–297.e8. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sharifi-Rad, M.; Salehi, B.; Iriti, M.; Roointan, A.; Mnayer, D.; Soltani-Nejad, A.; Afshari, A. In vitro and in vivo assessment of free radical scavenging and antioxidant activities of Veronica persica Poir. Cell. Mol. Biol. 2018, 64, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Kupeli, E.; Harput, U.S.; Varel, M.; Yesilada, E.; Saracoglu, I. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. J. Ethnopharmacol. 2005, 102, 170–176. [Google Scholar] [CrossRef]
- Harput, U.S.; Saracoglu, I.; Inoue, M.; Ogihara, Y. Anti-inflammatory and cytotoxic activities of five Veronica species. Biol. Pharm. Bull. 2002, 25, 483–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, K.S.; Song, H.K.; Hwang, Y.H.; Chae, S.; Kim, H.K.; Jang, S.; Kim, Y.H.; Choo, B.K.; Yang, W.K.; Kim, S.H.; et al. Ethanol extract of Veronica persica ameliorates house dust mite-induced asthmatic inflammation by inhibiting STAT-3 and STAT-6 activation. Biomed. Pharmacother. 2022, 152, 113264. [Google Scholar] [CrossRef] [PubMed]
- Eichenfield, L.F.; Hanifin, J.M.; Beck, L.A.; Lemanske, R.F., Jr.; Sampson, H.A.; Weiss, S.T.; Leung, D.Y. Atopic dermatitis and asthma: Parallels in the evolution of treatment. Pediatrics 2003, 111, 608–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal models of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Ryu, H.W.; Yang, W.K.; Park, M.H.; Park, Y.C.; Kim, D.Y.; Kwon, H.J.; Kim, S.Y.; Oh, S.R.; Kim, S.H. A combination of Olea europaea leaf extract and Spirodela polyrhiza extract alleviates atopic dermatitis by modulating immune balance and skin barrier function in a 1-chloro-2,4-dinitrobenzene-induced murine model. Phytomedicine 2021, 82, 153407. [Google Scholar] [CrossRef] [PubMed]
- Gallegos-Alcala, P.; Jimenez, M.; Cervantes-Garcia, D.; Salinas, E. The Keratinocyte as a Crucial Cell in the Predisposition, Onset, Progression, Therapy and Study of the Atopic Dermatitis. Int. J. Mol. Sci. 2021, 22, 661. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Kim, S.H.; Yang, W.K.; Park, Y.C.; Kim, H.K. Bleomycin Aggravates Atopic Dermatitis via Lung Inflammation in 2,4-Dinitrochlorobenzene-Induced NC/Nga Mice. Front. Pharmacol. 2018, 9, 578. [Google Scholar] [CrossRef]
- Mohd Kasim, V.N.K.; Noble, S.M.; Liew, K.Y.; Tan, J.W.; Israf, D.A.; Tham, C.L. Management of Atopic Dermatitis via Oral and Topical Administration of Herbs in Murine Model: A Systematic Review. Front. Pharmacol. 2022, 13, 785782. [Google Scholar] [CrossRef]
- Zhang, E.Y.; Zhu, B.T. Estriol strongly inhibits DNCB-induced contact dermatitis: Role of antigen-specific antibodies in pathogenesis. Endocr. Connect. 2014, 3, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Lee, C.H.; Kim, J.R.; Kwon, J.Y.; Seo, S.G.; Han, J.G.; Kim, B.G.; Kim, J.E.; Lee, K.W. Chlorella vulgaris Attenuates Dermatophagoides Farinae-Induced Atopic Dermatitis-Like Symptoms in NC/Nga Mice. Int. J. Mol. Sci. 2015, 16, 21021–21034. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, L.; Harvima, I.T. Mast cells of psoriatic and atopic dermatitis skin are positive for TNF-alpha and their degranulation is associated with expression of ICAM-1 in the epidermis. Arch. Dermatol. Res. 1998, 290, 353–359. [Google Scholar] [CrossRef]
- Kashiwakura, J.; Otani, I.M.; Kawakami, T. Monomeric IgE and mast cell development, survival and function. Adv. Exp. Med. Biol. 2011, 716, 29–46. [Google Scholar] [CrossRef]
- Malissen, B.; Tamoutounour, S.; Henri, S. The origins and functions of dendritic cells and macrophages in the skin. Nat. Rev. Immunol. 2014, 14, 417–428. [Google Scholar] [CrossRef]
- Pastore, S.; Corinti, S.; La Placa, M.; Didona, B.; Girolomoni, G. Interferon-gamma promotes exaggerated cytokine production in keratinocytes cultured from patients with atopic dermatitis. J. Allergy Clin. Immunol. 1998, 101, 538–544. [Google Scholar] [CrossRef]
- Trautmann, A.; Akdis, M.; Schmid-Grendelmeier, P.; Disch, R.; Brocker, E.B.; Blaser, K.; Akdis, C.A. Targeting keratinocyte apoptosis in the treatment of atopic dermatitis and allergic contact dermatitis. J. Allergy Clin. Immunol. 2001, 108, 839–846. [Google Scholar] [CrossRef]
- Rahman, M.S.; Alam, M.B.; Kim, Y.K.; Madina, M.H.; Fliss, I.; Lee, S.H.; Yoo, J.C. Activation of Nrf2/HO-1 by Peptide YD1 Attenuates Inflammatory Symptoms through Suppression of TLR4/MYyD88/NF-kappaB Signaling Cascade. Int. J. Mol. Sci. 2021, 22, 5161. [Google Scholar] [CrossRef]
- Augments Ogawa, T.; Ishitsuka, Y. The Role of KEAP1-NRF2 System in Atopic Dermatitis and Psoriasis. Antioxidants 2022, 11, 1397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Peng, L.; Fu, Y.; Wang, W.; Wang, P.; Zhou, F. Ginnalin A Binds to the Subpockets of Keap1 Kelch Domain to Activate the Nrf2-Regulated Antioxidant Defense System in SH-SY5Y Cells. ACS Chem. Neurosci. 2021, 12, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.L.; Weinberg, A.D.; English, M.; Huston, G. IL-4 directs the development of Th2-like helper effectors. J. Immunol. 1990, 145, 3796–3806. [Google Scholar] [CrossRef] [PubMed]
- Iezzi, G.; Boni, A.; Degl’Innocenti, E.; Grioni, M.; Bertilaccio, M.T.; Bellone, M. Type 2 cytotoxic T lymphocytes modulate the activity of dendritic cells toward type 2 immune responses. J. Immunol. 2006, 177, 2131–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zink, A.; Gensbaur, A.; Zirbs, M.; Seifert, F.; Suarez, I.L.; Mourantchanian, V.; Weidinger, S.; Mempel, M.; Ring, J.; Ollert, M. Targeting IgE in Severe Atopic Dermatitis with a Combination of Immunoadsorption and Omalizumab. Acta Derm. Venereol. 2016, 96, 72–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakonarson, H.; Maskeri, N.; Carter, C.; Chuang, S.; Grunstein, M.M. Autocrine interaction between IL-5 and IL-1beta mediates altered responsiveness of atopic asthmatic sensitized airway smooth muscle. J. Clin. Investig. 1999, 104, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Frempah, B.; Luckett-Chastain, L.R.; Gallucci, R.M. IL-6 Negatively Regulates IL-22Ralpha Expression on Epidermal Keratinocytes: Implications for Irritant Contact Dermatitis. J. Immunol. Res. 2019, 2019, 6276254. [Google Scholar] [CrossRef] [Green Version]
- Bieber, T. Interleukin-13: Targeting an underestimated cytokine in atopic dermatitis. Allergy 2020, 75, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Kasutani, K.; Fujii, E.; Ohyama, S.; Adachi, H.; Hasegawa, M.; Kitamura, H.; Yamashita, N. Anti-IL-31 receptor antibody is shown to be a potential therapeutic option for treating itch and dermatitis in mice. Br. J. Pharmacol. 2014, 171, 5049–5058. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.F.; Roan, F.; Bell, B.D.; Stoklasek, T.A.; Kitajima, M.; Han, H. The biology of thymic stromal lymphopoietin (TSLP). Adv. Pharmacol. 2013, 66, 129–155. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.R.; The, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Simon, D.; Vassina, E.; Yousefi, S.; Kozlowski, E.; Braathen, L.R.; Simon, H.U. Reduced dermal infiltration of cytokine-expressing inflammatory cells in atopic dermatitis after short-term topical tacrolimus treatment. J. Allergy Clin. Immunol. 2004, 114, 887–895. [Google Scholar] [CrossRef]
- Hyung, K.E.; Kim, S.J.; Jang, Y.W.; Lee, D.K.; Hyun, K.H.; Moon, B.S.; Kim, B.; Ahn, H.; Park, S.Y.; Sohn, U.D.; et al. Therapeutic effects of orally administered CJLP55 for atopic dermatitis via the regulation of immune response. Korean J. Physiol. Pharmacol. 2017, 21, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Kim, D.J.; Nam, S.; Lim, S.; Hwang, J.S.; Park, K.S.; Hong, H.S.; Shin, M.K.; Chung, E.; Son, Y. Manifestation of atopic dermatitis-like skin in TNCB-induced NC/Nga mice is ameliorated by topical treatment of substance P, possibly through blockade of allergic inflammation. Exp. Dermatol. 2018, 27, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Miller, H.L.; Andhey, P.S.; Swiecki, M.K.; Rosa, B.A.; Zaitsev, K.; Villani, A.C.; Mitreva, M.; Artyomov, M.N.; Gilfillan, S.; Cella, M.; et al. Altered ratio of dendritic cell subsets in skin-draining lymph nodes promotes Th2-driven contact hypersensitivity. Proc. Natl. Acad. Sci. USA 2021, 118, e2021364118. [Google Scholar] [CrossRef]
- Wagener, F.A.; van Beurden, H.E.; von den Hoff, J.W.; Adema, G.J.; Figdor, C.G. The heme-heme oxygenase system: A molecular switch in wound healing. Blood 2003, 102, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Chauveau, C.; Remy, S.; Royer, P.J.; Hill, M.; Tanguy-Royer, S.; Hubert, F.X.; Tesson, L.; Brion, R.; Beriou, G.; Gregoire, M.; et al. Heme oxygenase-1 expression inhibits dendritic cell maturation and proinflammatory function but conserves IL-10 expression. Blood 2005, 106, 1694–1702. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.S.; Shim, S.M.; Heo, J.; Pae, H.O.; Seo, B.Y.; Han, S.Y.; Sohn, D.H.; Jang, S.I.; Chung, H.T. Wogonin suppresses TARC expression induced by mite antigen via heme oxygenase 1 in human keratinocytes. Suppressive effect of wogonin on mite antigen-induced TARC expression. J. Dermatol. Sci. 2007, 46, 31–40. [Google Scholar] [CrossRef]
- Schafer, M.; Farwanah, H.; Willrodt, A.H.; Huebner, A.J.; Sandhoff, K.; Roop, D.; Hohl, D.; Bloch, W.; Werner, S. Nrf2 links epidermal barrier function with antioxidant defense. EMBO Mol. Med. 2012, 4, 364–379. [Google Scholar] [CrossRef]
- Napolitano, M.; Fabbrocini, G.; Martora, F.; Picone, V.; Morelli, P.; Patruno, C. Role of Aryl Hydrocarbon Receptor Activation in Inflammatory Chronic Skin Diseases. Cells 2021, 10, 3559. [Google Scholar] [CrossRef]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Uchi, H.; Takahara, M.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T.; Furue, M. Antioxidant soybean tar Glyteer rescues T-helper-mediated downregulation of filaggrin expression via aryl hydrocarbon receptor. J. Dermatol. 2015, 42, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bogaard, E.H.; Bergboer, J.G.; Vonk-Bergers, M.; van Vlijmen-Willems, I.M.; Hato, S.V.; van der Valk, P.G.; Schroder, J.M.; Joosten, I.; Zeeuwen, P.L.; Schalkwijk, J. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J. Clin. Investig. 2013, 123, 917–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.H.; Jayawickreme, C.; Rickard, D.J.; Nicodeme, E.; Bui, T.; Simmons, C.; Coquery, C.M.; Neil, J.; Pryor, W.M.; Mayhew, D.; et al. Tapinarof Is a Natural AhR Agonist that Resolves Skin Inflammation in Mice and Humans. J. Investig. Dermatol. 2017, 137, 2110–2119. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Xu, W.; Headley, M.B.; Jessup, H.K.; Lee, K.S.; Omori, M.; Comeau, M.R.; Marshak-Rothstein, A.; Ziegler, S.F. Thymic stromal lymphopoietin (TSLP)-mediated dermal inflammation aggravates experimental asthma. Mucosal Immunol. 2012, 5, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Tan, S.; Gu, W.; Li, F.; Hua, W.; Zhang, S.; Chen, F.; Tang, L. Phytochemical composition, isolation and hepatoprotective activity of active fraction from Veronica ciliata against acetaminophen-induced acute liver injury via p62-Keap1-Nrf2 signaling pathway. J. Ethnopharmacol. 2019, 243, 112089. [Google Scholar] [CrossRef]
- Moon, M.K.; Choi, B.M.; Oh, G.S.; Pae, H.O.; Kim, J.D.; Oh, H.; Oh, C.S.; Kim, D.H.; Rho, Y.D.; Shin, M.K.; et al. Catalposide protects Neuro 2A cells from hydrogen peroxide-induced cytotoxicity via the expression of heme oxygenase-1. Toxicol. Lett. 2003, 145, 46–54. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Chen, Z.; Liu, X. Picroside II attenuates ischemia/reperfusion testicular injury by alleviating oxidative stress and apoptosis through reducing nitric oxide synthesis. Acta Cir. Bras. 2019, 34, e201901102. [Google Scholar] [CrossRef] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, K.-S.; Park, M.; Yang, W.-K.; Lee, H.; Kim, S.-H.; Choo, B.-K.; Chae, S.; Kim, H.-K.; Kim, T.; Kim, K.-M. Veronica persica Ethanol Extract Ameliorates Dinitrochlorobenzene-Induced Atopic Dermatitis-like Skin Inflammation in Mice, Likely by Inducing Nrf2/HO-1 Signaling. Antioxidants 2023, 12, 1267. https://doi.org/10.3390/antiox12061267
Shim K-S, Park M, Yang W-K, Lee H, Kim S-H, Choo B-K, Chae S, Kim H-K, Kim T, Kim K-M. Veronica persica Ethanol Extract Ameliorates Dinitrochlorobenzene-Induced Atopic Dermatitis-like Skin Inflammation in Mice, Likely by Inducing Nrf2/HO-1 Signaling. Antioxidants. 2023; 12(6):1267. https://doi.org/10.3390/antiox12061267
Chicago/Turabian StyleShim, Ki-Shuk, Musun Park, Won-Kyung Yang, Hanbyeol Lee, Seung-Hyung Kim, Byung-Kil Choo, Sungwook Chae, Ho-Kyoung Kim, Taesoo Kim, and Ki-Mo Kim. 2023. "Veronica persica Ethanol Extract Ameliorates Dinitrochlorobenzene-Induced Atopic Dermatitis-like Skin Inflammation in Mice, Likely by Inducing Nrf2/HO-1 Signaling" Antioxidants 12, no. 6: 1267. https://doi.org/10.3390/antiox12061267