
Physical Exercise Mitigates Salivary Gland and Saliva Damages in Rats Exposed to Binge-like Ethanol Pattern
 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
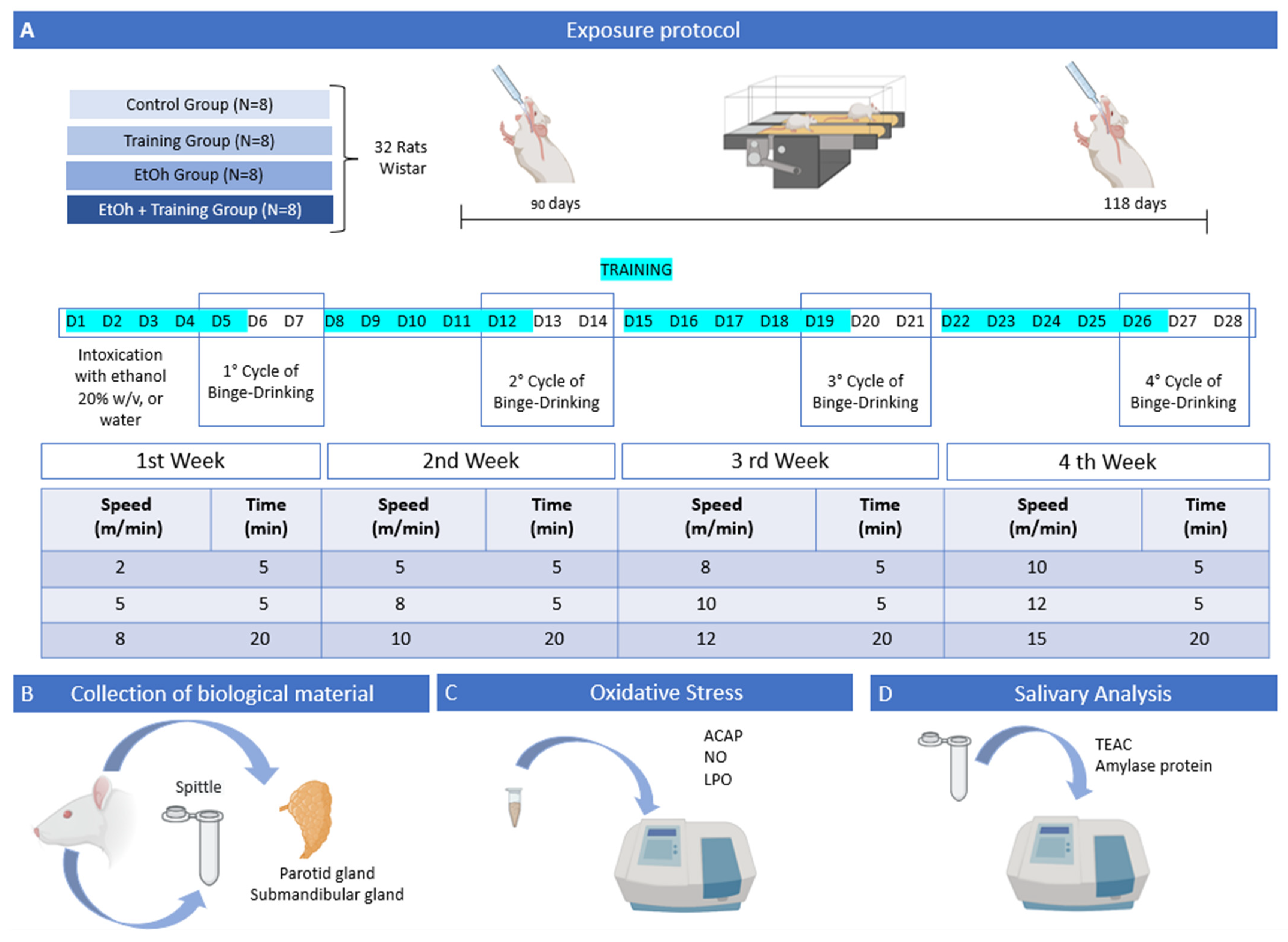
2.1. Animals and Experimental Groups
- The animal refuses to run;
- Running without constancy (runs and stops or runs in the wrong direction);
- Regular race;
- Good run (occasionally runs at the back of the treadmill);
- Excellent running (runs permanently at the front of the treadmill).
2.2. Binge-like EtOH Treatment
2.3. Aerobic Physical Training Protocol
2.4. Collection of Saliva and Salivary Glands
2.5. Salivary Gland Oxidative Biochemistry Analysis
2.5.1. Analysis of Antioxidant Capacity against Peroxyl Radicals (Antioxidant Capacity against Peroxyl Radicals—ACAP)
2.5.2. Lipid Peroxidation Assay
2.5.3. Analysis of Nitrite Concentration
2.6. Saliva Biochemical Analysis
2.6.1. Trolox-Equivalent Antioxidant Capacity (TEAC) Level
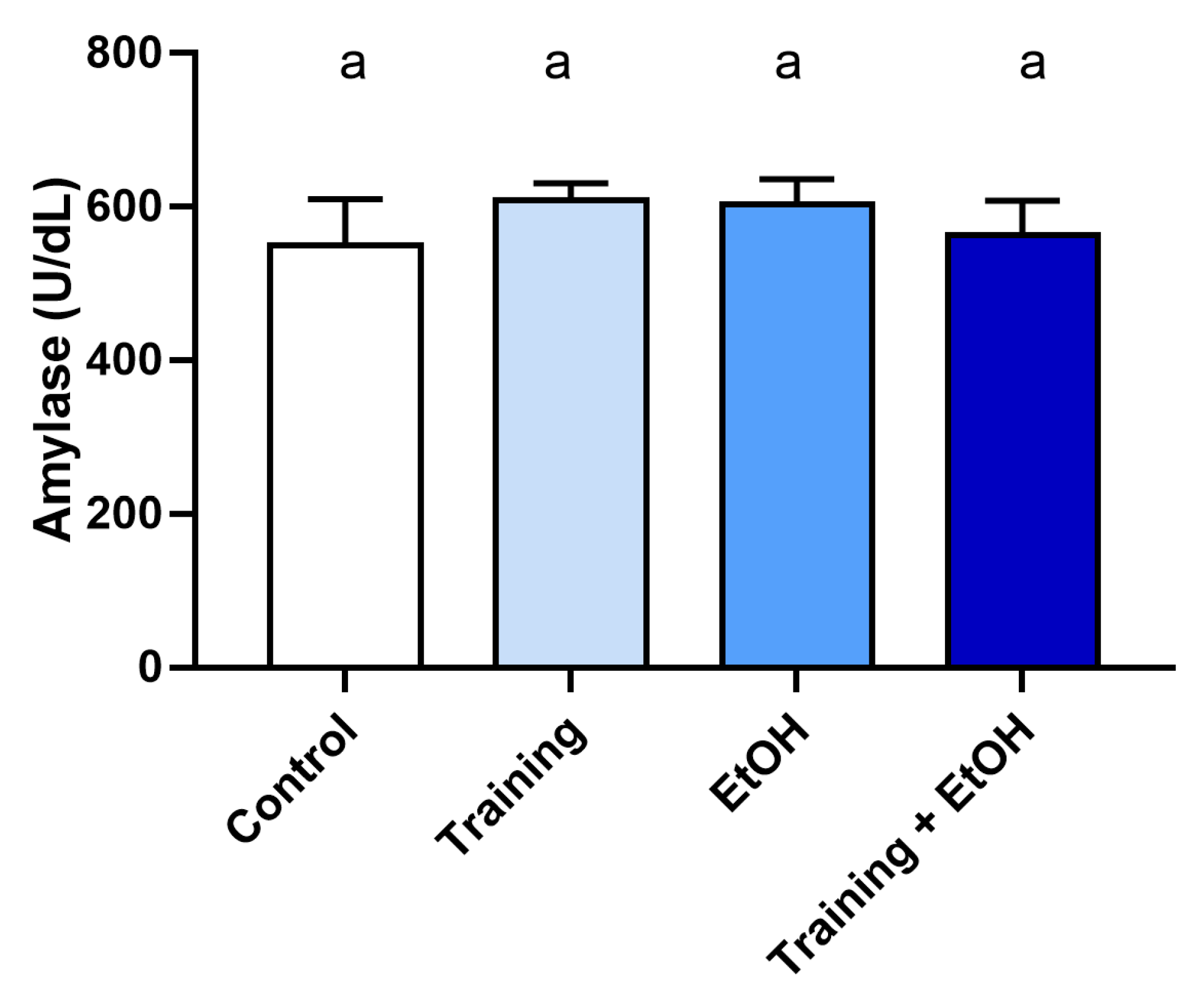
2.6.2. Amylase Activity
2.7. Statistical Analysis
3. Results
3.1. Physical Training Was Able to Attenuate the Oxidative Stress Generated by the EtOH Consumption in the Salivary Glands in a Binge-Drinking Model in Rats
3.2. The Intense and Episodic EtOH Consumption Promoted Alterations in the Salivary Oxidative Biochemistry of the Animals, but Aerobic Physical Training Could Not Minimize the Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Alcohol. World Health Organization. Available online: https://www.who.int/news-room/fact-sheets/detail/alcohol (accessed on 15 February 2023).
- Chikritzhs, T.; Livingston, M. Alcohol and the Risk of Injury. Nutrients 2021, 13, 2777. [Google Scholar] [CrossRef]
- Hendriks, H.F.J. Alcohol and Human Health: What Is the Evidence? Annu. Rev. Food Sci. Technol. 2020, 11, 1–21. [Google Scholar] [CrossRef]
- Krieger, H.; Young, C.M.; Anthenien, A.M.; Neighbors, C. The Epidemiology of Binge Drinking Among College-Age Individuals in the United States. Alcohol Res. Curr. Rev. 2018, 39, 23–30. [Google Scholar]
- Tapia-Rojas, C.; Mira, R.G.; Torres, A.K.; Jara, C.; Pérez, M.J.; Vergara, E.H.; Cerpa, W.; Quintanilla, R.A. Alcohol consumption during adolescence: A link between mitochondrial damage and ethanol brain intoxication. Birth Defects Res. 2017, 109, 1623–1639. [Google Scholar] [CrossRef] [PubMed]
- Drinking Patterns and Their Definitions. Alcohol Res. Curr. Rev. 2018, 39, 17–18.
- Fernandes, L.; de Andrade Jr, E.F.; Monteiro, M.; Cartágenes, S.; Lima, R.; Prediger, R.; Maia, C. Ethanol: Neurotoxicity and brain disorders. In Addictive Substances and Neurological Disease; Elsevier: Amsterdam, The Netherlands, 2017; pp. 201–215. [Google Scholar]
- Kamal, H.; Tan, G.C.; Ibrahim, S.F.; Shaikh, M.F.; Mohamed, I.N.; Mohamed, R.M.P.; Hamid, A.A.; Ugusman, A.; Kumar, J. Alcohol Use Disorder, Neurodegeneration, Alzheimer’s and Parkinson’s Disease: Interplay Between Oxidative Stress, Neuroimmune Response and Excitotoxicity. Front. Cell. Neurosci. 2020, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Han, J.; Lee, C.; Yoon, M.; Jung, Y. Pathophysiological Aspects of Alcohol Metabolism in the Liver. Int. J. Mol. Sci. 2021, 22, 5717. [Google Scholar] [CrossRef] [PubMed]
- Muñiz Hernandez, S.; Velazquez Fernandez, J.B.; Diaz Chavez, J.; Lopez Sanchez, R.C.; Hernandez, J.A.; Rendon Ramirez, A. Alcoholism: Common and oxidative damage biomarkers. J. Clin. Toxicol 2014, 1–8. [Google Scholar] [CrossRef]
- Fernandes, L.M.P.; Bezerra, F.R.; Monteiro, M.C.; Silva, M.L.; de Oliveira, F.R.; Lima, R.R.; Fontes-Júnior, E.A.; Maia, C.S.F. Thiamine deficiency, oxidative metabolic pathways and ethanol-induced neurotoxicity: How poor nutrition contributes to the alcoholic syndrome, as Marchiafava-Bignami disease. Eur. J. Clin. Nutr. 2017, 71, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Dal Prá, K.J.; de Fatima Moraes da Silva, A.; Chaves-Neto, A.H.; Soubhia, A.M.P. Effects of cachaça, a typical Brazilian alcoholic beverage, on submandibular glands of rats: A histomorphometric and biochemical study. Clin. Oral Investig. 2020, 24, 4283–4290. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, N.C.; Fernandes, L.M.; Paraense, R.S.; de Farias-Junior, P.M.; Teixeira, F.B.; Alves-Junior, S.M.; Pinheiro Jde, J.; Crespo-López, M.E.; Maia, C.S.; Lima, R.R. Binge Drinking of Ethanol during Adolescence Induces Oxidative Damage and Morphological Changes in Salivary Glands of Female Rats. Oxidative Med. Cell. Longev. 2016, 2016, 7323627. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, L.M.; Teixeira, F.B.; Alves-Junior, S.M.; Pinheiro Jde, J.; Maia, C.S.; Lima, R.R. Immunohistochemical changes and atrophy after chronic ethanol intoxication in rat salivary glands. Histol. Histopathol. 2015, 30, 1069–1078. [Google Scholar] [CrossRef]
- Chodzko-Zajko, W.J.; Proctor, D.N.; Fiatarone Singh, M.A.; Minson, C.T.; Nigg, C.R.; Salem, G.J.; Skinner, J.S. American College of Sports Medicine position stand. Exercise and physical activity for older adults. Med. Sci. Sport. Exerc. 2009, 41, 1510–1530. [Google Scholar] [CrossRef]
- Garber, C.E.; Blissmer, B.; Deschenes, M.R.; Franklin, B.A.; Lamonte, M.J.; Lee, I.M.; Nieman, D.C.; Swain, D.P. American College of Sports Medicine position stand. Quantity and quality of exercise for developing and maintaining cardiorespiratory, musculoskeletal, and neuromotor fitness in apparently healthy adults: Guidance for prescribing exercise. Med. Sci. Sport. Exerc. 2011, 43, 1334–1359. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Deminice, R.; Ozdemir, M.; Yoshihara, T.; Bomkamp, M.P.; Hyatt, H. Exercise-induced oxidative stress: Friend or foe? J. Sport Health Sci. 2020, 9, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lamarão-Vieira, K.; Pamplona-Santos, D.; Nascimento, P.C.; Corrêa, M.G.; Bittencourt, L.O.; Dos Santos, S.M.; Cartágenes, S.C.; Fernandes, L.M.P.; Monteiro, M.C.; Maia, C.S.F.; et al. Physical Exercise Attenuates Oxidative Stress and Morphofunctional Cerebellar Damages Induced by the Ethanol Binge Drinking Paradigm from Adolescence to Adulthood in Rats. Oxidative Med. Cell. Longev. 2019, 2019, 6802424. [Google Scholar] [CrossRef]
- Pamplona-Santos, D.; Lamarão-Vieira, K.; Nascimento, P.C.; Bittencourt, L.O.; Corrêa, M.G.; Dos Santos, S.M.; Cartágenes, S.C.; Fernandes, L.M.P.; Monteiro, M.C.; Maia, C.S.F.; et al. Aerobic Physical Exercise as a Neuroprotector Strategy for Ethanol Binge-Drinking Effects in the Hippocampus and Systemic Redox Status in Rats. Oxidative Med. Cell. Longev. 2019, 2019, 2415243. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.C.C.; Ferreira, R.O.; Frazão, D.R.; de Souza Né, Y.G.; Mendes, P.F.S.; Marañón-Vásquez, G.; Royes, L.F.F.; Fagundes, N.C.F.; Maia, L.C.; Lima, R.R. The Relationship between Exercise and Salivary Oxidative Stress: A Systematic Review. Antioxidants 2022, 11, 1489. [Google Scholar] [CrossRef]
- Dishman, R.K.; Armstrong, R.B.; Delp, M.D.; Graham, R.E.; Dunn, A.L. Open-field behavior is not related to treadmill performance in exercising rats. Physiol. Behav. 1988, 43, 541–546. [Google Scholar] [CrossRef]
- Arida, R.M.; Scorza, F.A.; de Lacerda, A.F.; Gomes da Silva, S.; Cavalheiro, E.A. Physical training in developing rats does not influence the kindling development in the adult life. Physiol. Behav. 2007, 90, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, L.M.P.; Lopes, K.S.; Santana, L.N.S.; Fontes-Júnior, E.A.; Ribeiro, C.; Silva, M.C.F.; de Oliveira Paraense, R.S.; Crespo-López, M.E.; Gomes, A.R.Q.; Lima, R.R.; et al. Repeated Cycles of Binge-Like Ethanol Intake in Adolescent Female Rats Induce Motor Function Impairment and Oxidative Damage in Motor Cortex and Liver, but Not in Blood. Oxidative Med. Cell. Longev. 2018, 2018, 3467531. [Google Scholar] [CrossRef] [PubMed]
- Arida, R.M.; Scorza, F.A.; Gomes da Silva, S.; Cysneiros, R.M.; Cavalheiro, E.A. Exercise paradigms to study brain injury recovery in rodents. Am. J. Phys. Med. Rehabil. 2011, 90, 452–465. [Google Scholar] [CrossRef]
- Amado, L.L.; Garcia, M.L.; Ramos, P.B.; Freitas, R.F.; Zafalon, B.; Ferreira, J.L.; Yunes, J.S.; Monserrat, J.M. A method to measure total antioxidant capacity against peroxyl radicals in aquatic organisms: Application to evaluate microcystins toxicity. Sci. Total Environ. 2009, 407, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Cheeseman, K.H. Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4-hydroxynonenal. Methods Enzymol. 1990, 186, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Ruiz de Luzuriaga, K.; Wagner, D.A.; Rand, W.; Istfan, N.; Young, V.R.; Tannenbaum, S.R. Nitrate biosynthesis in man. Proc. Natl. Acad. Sci. USA 1981, 78, 7764–7768. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Caraway, W.T. A stable starch substrate for the determination of amylase in serum and other body fluids. Am. J. Clin. Pathol. 1959, 32, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.N.; Hoffman, M.P. Salivary gland development: A template for regeneration. Semin. Cell Dev. Biol. 2014, 25, 52–60. [Google Scholar] [CrossRef]
- de Paula, F.; Teshima, T.H.N.; Hsieh, R.; Souza, M.M.; Nico, M.M.S.; Lourenco, S.V. Overview of Human Salivary Glands: Highlights of Morphology and Developing Processes. Anat. Rec. (Hoboken N. J. 2007) 2017, 300, 1180–1188. [Google Scholar] [CrossRef]
- Holmberg, K.V.; Hoffman, M.P. Anatomy, biogenesis and regeneration of salivary glands. Monogr. Oral Sci. 2014, 24, 1–13. [Google Scholar] [CrossRef]
- Amano, O.; Mizobe, K.; Bando, Y.; Sakiyama, K. Anatomy and histology of rodent and human major salivary glands: -overview of the Japan salivary gland society-sponsored workshop. Acta Histochem. Et Cytochem. 2012, 45, 241–250. [Google Scholar] [CrossRef]
- Proctor, G.B.; Carpenter, G.H. Regulation of salivary gland function by autonomic nerves. Auton. Neurosci. 2007, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Tucker, A. Salivary gland development. In Proceedings of the Seminars in Cell & Developmental Biology; 2007; pp. 237–244. [Google Scholar]
- Busch, L.; Sterin-Borda, L.; Borda, E. Differences in the regulatory mechanism of amylase release by rat parotid and submandibular glands. Arch. Oral Biol. 2002, 47, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.; Rehman, A.; Shah, K.; Kamran, M.; Mashal, S.; Rustam, S.; Sabir, M.; Nayab, A.; Muzammal, M. Composition and function of saliva: A review. World J. Pharm. Pharm. Sci 2020, 9, 1552–1567. [Google Scholar]
- Wu, A.M.; Csako, G.; Herp, A. Structure, biosynthesis, and function of salivary mucins. Mol. Cell. Biochem. 1994, 137, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Maddu, N. Functions of saliva. In Saliva and Salivary Diagnostics; IntechOpen: London, UK, 2019. [Google Scholar]
- Hernández, J.A.; López-Sánchez, R.C.; Rendón-Ramírez, A. Lipids and Oxidative Stress Associated with Ethanol-Induced Neurological Damage. Oxidative Med. Cell. Longev. 2016, 2016, 1543809. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K.; Sugimoto, N.; Katakura, M.; Sumiyoshi, E.; Hara, T.; Hashimoto, M.; Shido, O. Daily voluntary exercise enhances pilocarpine-induced saliva secretion and aquaporin 1 expression in rat submandibular glands. FEBS Open Bio. 2018, 8, 85–93. [Google Scholar] [CrossRef]
- Ferreira, R.O.; Aragão, W.A.B.; Bittencourt, L.O.; Fernandes, L.P.M.; Balbinot, K.M.; Alves-Junior, S.M.; Pinheiro, J.J.V.; Maia, C.; Crespo-Lopez, M.E.; Lima, R.R. Ethanol binge drinking during pregnancy and its effects on salivary glands of offspring rats: Oxidative stress, morphometric changes and salivary function impairments. Biomed. Pharmacother. = Biomed. Pharmacother. 2021, 133, 110979. [Google Scholar] [CrossRef]
- Jung, W.K.; Park, S.B.; Kim, H.R.; Ryu, H.Y.; Kim, Y.H.; Kim, J. Advanced Glycation End Products Increase Salivary Gland Hypofunction in d-Galactose-Induced Aging Rats and Its Prevention by Physical Exercise. Curr. Issues Mol. Biol. 2021, 43, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.D.; Oliveira, A.R.d. Oxygen free radicals and exercise: Mechanisms of synthesis and adaptation to the physical training. Rev. Bras. De Med. Do Esporte 2004, 10, 308–313. [Google Scholar] [CrossRef]
- Subudhi, A.W.; Davis, S.L.; Kipp, R.W.; Askew, E.W. Antioxidant status and oxidative stress in elite alpine ski racers. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 32–41. [Google Scholar] [CrossRef]
- Kröller-Schön, S.; Jansen, T.; Hauptmann, F.; Schüler, A.; Heeren, T.; Hausding, M.; Oelze, M.; Viollet, B.; Keaney, J.F., Jr.; Wenzel, P. α1AMP-activated protein kinase mediates vascular protective effects of exercise. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Child, R.B.; Wilkinson, D.M.; Fallowfield, J.L.; Donnelly, A.E. Elevated serum antioxidant capacity and plasma malondialdehyde concentration in response to a simulated half-marathon run. Med. Sci. Sport. Exerc. 1998, 30, 1603–1607. [Google Scholar] [CrossRef] [PubMed]
- Vetreno, R.P.; Crews, F.T. Binge ethanol exposure during adolescence leads to a persistent loss of neurogenesis in the dorsal and ventral hippocampus that is associated with impaired adult cognitive functioning. Front. Neurosci. 2015, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh Maleki, B.; Tartibian, B.; Mooren, F.C.; FitzGerald, L.Z.; Krüger, K.; Chehrazi, M.; Malandish, A. Low-to-moderate intensity aerobic exercise training modulates irritable bowel syndrome through antioxidative and inflammatory mechanisms in women: Results of a randomized controlled trial. Cytokine 2018, 102, 18–25. [Google Scholar] [CrossRef]
- Farzanegi, P.; Dana, A.; Ebrahimpoor, Z.; Asadi, M.; Azarbayjani, M.A. Mechanisms of beneficial effects of exercise training on non-alcoholic fatty liver disease (NAFLD): Roles of oxidative stress and inflammation. Eur. J. Sport Sci. 2019, 19, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cabrera, M.C.; Domenech, E.; Viña, J. Moderate exercise is an antioxidant: Upregulation of antioxidant genes by training. Free. Radic. Biol. Med. 2008, 44, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Tung, B.T.; Rodriguez-Bies, E.; Thanh, H.N.; Le-Thi-Thu, H.; Navas, P.; Sanchez, V.M.; López-Lluch, G. Organ and tissue-dependent effect of resveratrol and exercise on antioxidant defenses of old mice. Aging Clin. Exp. Res. 2015, 27, 775–783. [Google Scholar] [CrossRef]
- Lima, F.D.; Stamm, D.N.; Della-Pace, I.D.; Dobrachinski, F.; de Carvalho, N.R.; Royes, L.F.; Soares, F.A.; Rocha, J.B.; González-Gallego, J.; Bresciani, G. Swimming training induces liver mitochondrial adaptations to oxidative stress in rats submitted to repeated exhaustive swimming bouts. PLoS ONE 2013, 8, e55668. [Google Scholar] [CrossRef]
- Martins, R.R.; de Oliveira Macedo, U.B.; Leite, L.D.; Rezende, A.A.; Brandão-Neto, J.; Almeida, M. Lipoic acid and moderate swimming improves the estrous cycle and oxidative stress in Wistar rats. Appl. Physiol. Nutr. Metab. Physiol. Appl. Nutr. Metab. 2011, 36, 693–697. [Google Scholar] [CrossRef]
- González, D.; Marquina, R.; Rondón, N.; Rodriguez-Malaver, A.J.; Reyes, R. Effects of aerobic exercise on uric acid, total antioxidant activity, oxidative stress, and nitric oxide in human saliva. Res. Sport. Med. (Print) 2008, 16, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Buthet, L.R.; Maciel, M.E.; Quintans, L.N.; Rodríguez de Castro, C.; Costantini, M.H.; Fanelli, S.L.; Castro, J.A.; Castro, G.D. Acetaldehyde content and oxidative stress in the deleterious effects of alcohol drinking on rat uterine horn. J. Toxicol. 2013, 2013, 161496. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Ito, H.; Matsui, H.; Hyodo, I. Acetaldehyde is an oxidative stressor for gastric epithelial cells. J. Clin. Biochem. Nutr. 2014, 55, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Homann, N.; Tillonen, J.; Meurman, J.H.; Rintamäki, H.; Lindqvist, C.; Rautio, M.; Jousimies-Somer, H.; Salaspuro, M. Increased salivary acetaldehyde levels in heavy drinkers and smokers: A microbiological approach to oral cavity cancer. Carcinogenesis 2000, 21, 663–668. [Google Scholar] [CrossRef]
- King, S.E.; Waddell, J.T.; Corbin, W.R. A preliminary investigation of salivary α-amylase as a novel biomarker of subjective response to alcohol: A brief report. Exp. Clin. Psychopharmacol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.J.; Fortes, M.B.; Richardson, K.; Bilzon, J.L.; Walsh, N.P. The effects of post-exercise feeding on saliva anti-microbial proteins. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Yamane, A.; Suzuki, M.; Nakagawa, Y. Availability of saliva for the assessment of alterations in the autonomic nervous system caused by physical exercise training. Arch. Oral Biol. 2009, 54, 977–985. [Google Scholar] [CrossRef]
- Rosa, L.; Teixeira, A.; Lira, F.; Tufik, S.; Mello, M.; Santos, R. Moderate acute exercise (70% VO2 peak) induces TGF-β, α-amylase and IgA in saliva during recovery. Oral Dis. 2014, 20, 186–190. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peinado, B.R.R.; Aragão, W.A.B.; Pereira, C.d.S.; da Silva, D.C.B.; Ferreira, R.d.O.; Chaves-Neto, A.H.; Fernandes, L.M.P.; Lima, R.R. Physical Exercise Mitigates Salivary Gland and Saliva Damages in Rats Exposed to Binge-like Ethanol Pattern. Antioxidants 2023, 12, 1038. https://doi.org/10.3390/antiox12051038
Peinado BRR, Aragão WAB, Pereira CdS, da Silva DCB, Ferreira RdO, Chaves-Neto AH, Fernandes LMP, Lima RR. Physical Exercise Mitigates Salivary Gland and Saliva Damages in Rats Exposed to Binge-like Ethanol Pattern. Antioxidants. 2023; 12(5):1038. https://doi.org/10.3390/antiox12051038
Chicago/Turabian StylePeinado, Beatriz Rodrigues Risuenho, Walessa Alana Bragança Aragão, Cristian dos Santos Pereira, Diane Cleydes Baia da Silva, Railson de Oliveira Ferreira, Antônio Hernandes Chaves-Neto, Luanna Melo Pereira Fernandes, and Rafael Rodrigues Lima. 2023. "Physical Exercise Mitigates Salivary Gland and Saliva Damages in Rats Exposed to Binge-like Ethanol Pattern" Antioxidants 12, no. 5: 1038. https://doi.org/10.3390/antiox12051038