
Intraoperative Hemi-Diaphragm Electrical Stimulation Demonstrates Attenuated Mitochondrial Function without Change in Oxidative Stress in Cardiothoracic Surgery Patients
 ,
,  and
and
Abstract
:1. Background
2. Methods
2.1. Study Design
2.2. Screening Procedures
2.3. Phrenic Stimulation
2.4. Diaphragm Biopsies
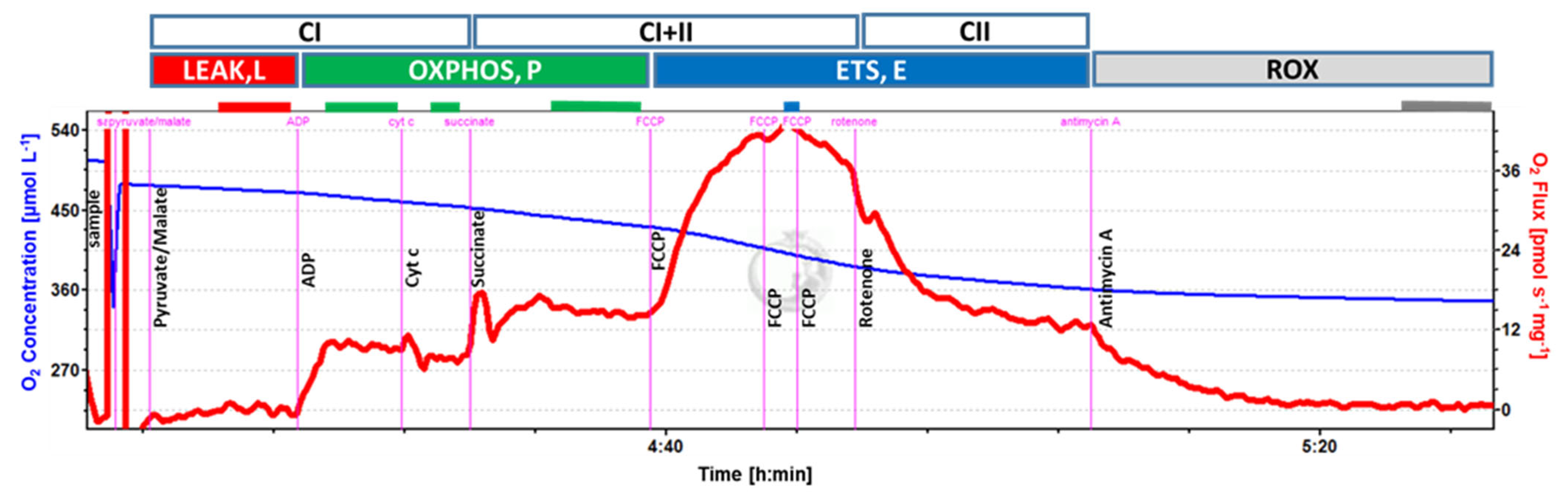
2.5. Mitochondrial Respiration
2.6. Enzymatic Activity
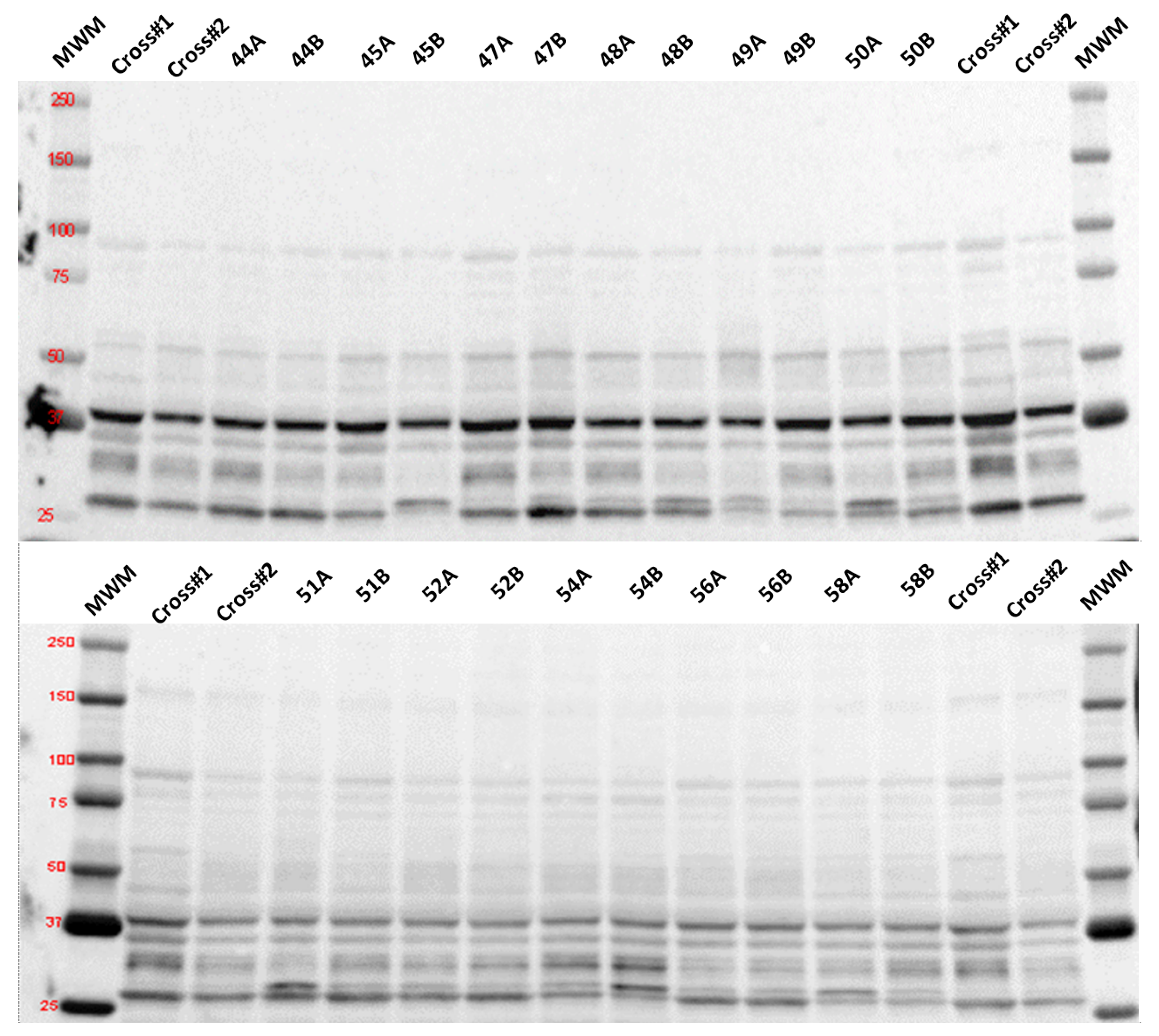
2.7. Determination of Selected Proteins by Immunodetection
2.8. Statistical Analysis
3. Results
3.1. Patient Characteristics and Tissue Acquisition
3.2. Mitochondrial Respiration
3.3. Oxidative Stress
3.4. Mitophagy
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Alexander, J.H.; Smith, P.K. Coronary-Artery Bypass Grafting. N. Engl. J. Med. 2016, 375, e22. [Google Scholar] [CrossRef] [PubMed]
- Goligher, E.C.; Dres, M.; Fan, E.; Rubenfeld, G.D.; Scales, D.C.; Herridge, M.S.; Vorona, S.; Sklar, M.C.; Rittayamai, N.; Lanys, A.; et al. Mechanical Ventilation-Induced Diaphragm Atrophy Strongly Impacts Clinical Outcomes. Am. J. Respir. Crit. Care Med. 2018, 197, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Horn, A.G.; Davis, R.T., 3rd; Baumfalk, D.R.; Kunkel, O.N.; Bruells, C.S.; McCullough, D.J.; Opoku-Acheampong, A.B.; Poole, D.C.; Behnke, B.J. Impaired diaphragm resistance vessel vasodilation with prolonged mechanical ventilation. J. Appl. Physiol. 2019, 127, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, H.W.; Ozdemir, M.; Yoshihara, T.; Nguyen, B.L.; Deminice, R.; Powers, S.K. Calpains play an essential role in mechanical ventilation-induced diaphragmatic weakness and mitochondrial dysfunction. Redox Biol. 2021, 38, 101802. [Google Scholar] [CrossRef]
- Sollanek, K.J.; Burniston, J.G.; Kavazis, A.N.; Morton, A.B.; Wiggs, M.P.; Ahn, B.; Smuder, A.J.; Powers, S.K. Global Proteome Changes in the Rat Diaphragm Induced by Endurance Exercise Training. PLoS ONE 2017, 12, e0171007. [Google Scholar] [CrossRef]
- Picard, M.; Azuelos, I.; Jung, B.; Giordano, C.; Matecki, S.; Hussain, S.; White, K.; Li, T.; Liang, F.; Benedetti, A.; et al. Mechanical ventilation triggers abnormal mitochondrial dynamics and morphology in the diaphragm. J. Appl. Physiol. 2015, 118, 1161–1171. [Google Scholar] [CrossRef]
- van den Berg, M.; Hooijman, P.E.; Beishuizen, A.; de Waard, M.C.; Paul, M.A.; Hartemink, K.J.; van Hees, H.W.H.; Lawlor, M.W.; Brocca, L.; Bottinelli, R.; et al. Diaphragm Atrophy and Weakness in the Absence of Mitochondrial Dysfunction in the Critically Ill. Am. J. Respir. Crit. Care Med. 2017, 196, 1544–1558. [Google Scholar] [CrossRef]
- Martin, A.D.; Joseph, A.M.; Beaver, T.M.; Smith, B.K.; Martin, T.D.; Berg, K.; Hess, P.J.; Deoghare, H.V.; Leeuwenburgh, C. Effect of intermittent phrenic nerve stimulation during cardiothoracic surgery on mitochondrial respiration in the human diaphragm. Crit. Care Med. 2014, 42, e152–e156. [Google Scholar] [CrossRef]
- Mankowski, R.T.; Ahmed, S.; Beaver, T.; Dirain, M.; Han, C.; Hess, P.; Martin, T.; Smith, B.K.; Someya, S.; Leeuwenburgh, C.; et al. Intraoperative hemidiaphragm electrical stimulation reduces oxidative stress and upregulates autophagy in surgery patients undergoing mechanical ventilation: Exploratory study. J. Transl. Med. 2016, 14, 305. [Google Scholar] [CrossRef]
- Graham, B.L.; Steenbruggen, I.; Miller, M.R.; Barjaktarevic, I.Z.; Cooper, B.G.; Hall, G.L.; Hallstrand, T.S.; Kaminsky, D.A.; McCarthy, K.; McCormack, M.C.; et al. Standardization of Spirometry 2019 Update. An Official American Thoracic Society and European Respiratory Society Technical Statement. Am. J. Respir. Crit. Care Med. 2019, 200, e70–e88. [Google Scholar] [CrossRef]
- Laveneziana, P.; Albuquerque, A.; Aliverti, A.; Babb, T.; Barreiro, E.; Dres, M.; Dube, B.P.; Fauroux, B.; Gea, J.; Guenette, J.A.; et al. ERS statement on respiratory muscle testing at rest and during exercise. Eur. Respir. J. 2019, 53, 1801214. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Veksler, V.; Gellerich, F.N.; Saks, V.; Margreiter, R.; Kunz, W.S. Analysis of mitochondrial function in situ in permeabilized muscle fibers, tissues and cells. Nat. Protoc. 2008, 3, 965–976. [Google Scholar] [CrossRef]
- Li, C.; White, S.H.; Warren, L.K.; Wohlgemuth, S.E. Effects of aging on mitochondrial function in skeletal muscle of American American Quarter Horses. J. Appl. Physiol. 2016, 121, 299–311. [Google Scholar] [CrossRef]
- Picca, A.; Saini, S.K.; Mankowski, R.T.; Kamenov, G.; Anton, S.D.; Manini, T.M.; Buford, T.W.; Wohlgemuth, S.E.; Xiao, R.; Calvani, R.; et al. Altered Expression of Mitoferrin and Frataxin, Larger Labile Iron Pool and Greater Mitochondrial DNA Damage in the Skeletal Muscle of Older Adults. Cells 2020, 9, 2579. [Google Scholar] [CrossRef] [PubMed]
- Amirbeigiarab, S.; Kiani, P.; Velazquez Sanchez, A.; Krisp, C.; Kazantsev, A.; Fester, L.; Schluter, H.; Ignatova, Z. Invariable stoichiometry of ribosomal proteins in mouse brain tissues with aging. Proc. Natl. Acad. Sci. USA 2019, 116, 22567–22572. [Google Scholar] [CrossRef] [PubMed]
- Fosang, A.J.; Colbran, R.J. Transparency Is the Key to Quality. J. Biol. Chem. 2015, 290, 29692–29694. [Google Scholar] [CrossRef] [PubMed]
- Thacker, J.S.; Yeung, D.H.; Staines, W.R.; Mielke, J.G. Total protein or high-abundance protein: Which offers the best loading control for Western blotting? Anal. Biochem. 2016, 496, 76–78. [Google Scholar] [CrossRef]
- Larsen, S.; Nielsen, J.; Hansen, C.N.; Nielsen, L.B.; Wibrand, F.; Stride, N.; Schroder, H.D.; Boushel, R.; Helge, J.W.; Dela, F.; et al. Biomarkers of mitochondrial content in skeletal muscle of healthy young human subjects. J. Physiol. 2012, 590, 3349–3360. [Google Scholar] [CrossRef]
- Bresciani, G.B.; Beaver, T.; Martin, A.D.; Van der Pijl, R.; Mankowski, R.; Leeuwenburgh, C.; Ottenheijm, C.A.C.; Martin, T.; Arnaoutakis, G.J.; Ahmed, S.; et al. Intraoperative phrenic stimulation offsets diaphragm fiber weakness during cardiothoracic surgery. medRxiv 2022. [Google Scholar] [CrossRef]
- Layec, G.; Blain, G.M.; Rossman, M.J.; Park, S.Y.; Hart, C.R.; Trinity, J.D.; Gifford, J.R.; Sidhu, S.K.; Weavil, J.C.; Hureau, T.J.; et al. Acute High-Intensity Exercise Impairs Skeletal Muscle Respiratory Capacity. Med. Sci. Sports Exerc. 2018, 50, 2409–2417. [Google Scholar] [CrossRef]
- Larsen, F.J.; Schiffer, T.A.; Ortenblad, N.; Zinner, C.; Morales-Alamo, D.; Willis, S.J.; Calbet, J.A.; Holmberg, H.C.; Boushel, R. High-intensity sprint training inhibits mitochondrial respiration through aconitase inactivation. FASEB J. 2016, 30, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, N.; Sato, S.; Shiba, K.; Okatsu, K.; Saisho, K.; Gautier, C.A.; Sou, Y.S.; Saiki, S.; Kawajiri, S.; Sato, F.; et al. PINK1 stabilized by mitochondrial depolarization recruits Parkin to damaged mitochondria and activates latent Parkin for mitophagy. J. Cell Biol. 2010, 189, 211–221. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, T.G.; Prescott, A.R.; Montava-Garriga, L.; Ball, G.; Singh, F.; Barini, E.; Muqit, M.M.K.; Brooks, S.P.; Ganley, I.G. Basal Mitophagy Occurs Independently of PINK1 in Mouse Tissues of High Metabolic Demand. Cell Metab. 2018, 27, 439–449.e5. [Google Scholar] [CrossRef] [PubMed]
- Yamano, K.; Matsuda, N.; Tanaka, K. The ubiquitin signal and autophagy: An orchestrated dance leading to mitochondrial degradation. EMBO Rep. 2016, 17, 300–316. [Google Scholar] [CrossRef]
- Schwalm, C.; Jamart, C.; Benoit, N.; Naslain, D.; Premont, C.; Prevet, J.; Van Thienen, R.; Deldicque, L.; Francaux, M. Activation of autophagy in human skeletal muscle is dependent on exercise intensity and AMPK activation. FASEB J. 2015, 29, 3515–3526. [Google Scholar] [CrossRef]
- Tardo-Dino, P.E.; Touron, J.; Bauge, S.; Bourdon, S.; Koulmann, N.; Malgoyre, A. The effect of a physiological increase in temperature on mitochondrial fatty acid oxidation in rat myofibers. J. Appl. Physiol. 2019, 127, 312–319. [Google Scholar] [CrossRef]
- Jarmuszkiewicz, W.; Woyda-Ploszczyca, A.; Koziel, A.; Majerczak, J.; Zoladz, J.A. Temperature controls oxidative phosphorylation and reactive oxygen species production through uncoupling in rat skeletal muscle mitochondria. Free. Radic. Biol. Med. 2015, 83, 12–20. [Google Scholar] [CrossRef]
- Ahn, B.; Beaver, T.; Martin, T.; Hess, P.; Brumback, B.A.; Ahmed, S.; Smith, B.K.; Leeuwenburgh, C.; Martin, A.D.; Ferreira, L.F. Phrenic nerve stimulation increases human diaphragm fiber force after cardiothoracic surgery. Am. J. Respir. Crit. Care Med. 2014, 190, 837–839. [Google Scholar] [CrossRef]
- Decker, S.T.; Kwon, O.S.; Zhao, J.; Hoidal, J.R.; Heuckstadt, T.; Richardson, R.S.; Sanders, K.A.; Layec, G. Skeletal muscle mitochondrial adaptations induced by long-term cigarette smoke exposure. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E80–E89. [Google Scholar] [CrossRef]
- Gifford, J.R.; Trinity, J.D.; Kwon, O.S.; Layec, G.; Garten, R.S.; Park, S.Y.; Nelson, A.D.; Richardson, R.S. Altered skeletal muscle mitochondrial phenotype in COPD: Disease vs. disuse. J. Appl. Physiol. 2018, 124, 1045–1053. [Google Scholar] [CrossRef]
- Puente-Maestu, L.; Lazaro, A.; Tejedor, A.; Camano, S.; Fuentes, M.; Cuervo, M.; Navarro, B.O.; Agusti, A. Effects of exercise on mitochondrial DNA content in skeletal muscle of patients with COPD. Thorax 2011, 66, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Gnoni, A.; Ballini, A.; Trentadue, R.; Taurino, F.; Santacroce, L.; Ferrara, P.; Massaro, F.; Brienza, N.; Massari, A.M.; Sardaro, N.; et al. Induction of mitochondrial dysfunction in patients under cardiopulmonary by-pass: Preliminary results. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8115–8123. [Google Scholar] [PubMed]
- Sirvinskas, E.; Kinderyte, A.; Trumbeckaite, S.; Lenkutis, T.; Raliene, L.; Giedraitis, S.; Macas, A.; Borutaite, V. Effects of sevoflurane vs. propofol on mitochondrial functional activity after ischemia-reperfusion injury and the influence on clinical parameters in patients undergoing CABG surgery with cardiopulmonary bypass. Perfusion 2015, 30, 590–595. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, F.; Castillo-Olivares, J.L.; Romero, G.; Eguaras, M.G.; Juffe, A.; Gosalvez, M.; Figuera, D. Anoxic cardiac arrest: Its effect on myocardial mitochondrial metabolism. J. Cardiovasc. Surg. 1975, 16, 493–499. [Google Scholar]
- Ragnarsdottir, M.; KristjAnsdottir, A.; Ingvarsdottir, I.; Hannesson, P.; Torfason, B.; Cahalin, L. Short-term changes in pulmonary function and respiratory movements after cardiac surgery via median sternotomy. Scand. Cardiovasc. J. 2004, 38, 46–52. [Google Scholar] [CrossRef]
- Hooijman, P.E.; Ottenheijm, C.A. Mitochondrial respiration and passive stretch of the diaphragm during unilateral phrenic nerve stimulation. Crit. Care Med. 2014, 42, e633–e634. [Google Scholar] [CrossRef]
- van der Pijl, R.; Strom, J.; Conijn, S.; Lindqvist, J.; Labeit, S.; Granzier, H.; Ottenheijm, C. Titin-based mechanosensing modulates muscle hypertrophy. J. Cachexia Sarcopenia Muscle 2018, 9, 947–961. [Google Scholar] [CrossRef]
- Glancy, B.; Willis, W.T.; Chess, D.J.; Balaban, R.S. Effect of calcium on the oxidative phosphorylation cascade in skeletal muscle mitochondria. Biochemistry 2013, 52, 2793–2809. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Patient # | Age (Yrs)/Sex | BMI (Kg/m2) | %FVC | %FEV1 | PI Max (cm H2O) | Intubation to Biopsy (Min) | Surgery Type |
|---|---|---|---|---|---|---|---|
| 1 | 61/F | 36.2 | 105 | 96 | 83 | 450 | Maze, MVR, AVR |
| 2 | 60/M | 29.6 | 85 | 90 | 85 | 289 | Ascending aorta graft, aortic root replacement, AVR |
| 3 | 71/F | 28.8 | 78 | 86 | 67 | 249 | CABG × 3 |
| 4 | 76/F | 29.6 | 79 | 83 | 79 | 232 | CABG × 3 |
| 5 | 74/F | 21.4 | 98 | 87 | 83 | 319 | CABG × 1, MVR |
| 6 | 62/F | 30.8 | 100 | 101 | 72 | 295 | CABG × 4 |
| 7 | 58/F | 34.6 | 73 | 71 | 61 | 293 | CABG × 3 |
| 8 | 62/M | 28.9 | - | - | - | 281 | Bilateral lung transplant |
| 9 | 55/M | 27.1 | - | - | - | 273 | Bilateral lung transplant |
| 10 | 69/M | 27.5 | 65 | 58 | 92 | 240 | AVR, Maze |
| 11 | 47/M | 32 | 101 | 97 | 120 | 203 | AVR |
| 12 | 55/M | 27.9 | 78 | 64 | 76 | 230 | Ascending aorta graft, aortic root replacement, AVR |
| 13 | 35/M | 34.5 | 90 | 89 | 155 | 358 | CABG × 3 |
| 14 | 72/M | 30.1 | 87 | 94 | 86 | 261 | CABG × 2 |
| 15 | 56/F | 31.8 | 81 | 79 | 83 | 233 | Mitral repair |
| 16 | 35/F | 32.9 | 95 | 96 | 64 | 235 | MVR |
| 17 | 51/M | 31.9 | 111 | 106 | 110 | 415 | AVR, CABG × 3 |
| 18 | 49/M | 22.6 | 78 | 68 | 93 | 269 | Ascending aorta graft, aortic root, AVR, MVR, Maze |
| 19 | 71/M | 24.3 | 83 | 78 | 124 | 195 | AVR, MVR |
| 20 | 57/F | 35.4 | 91 | 83 | 90 | 227 | AVR, ascending aorta graft |
| 21 | 63/M | 28.2 | 92 | 94 | 80 | 286 | CABG × 4 |
| Patient # | Intubation to First Stim (Min) | Cardio-Pulmonary Bypass (Min) | Average Stimulation (mA) | Number of Stims | Minimum Core Temp (C) | Core Temp at Biopsy (C) | Intubation to Biopsy (Min) |
|---|---|---|---|---|---|---|---|
| 1 | 104 | 341 | 6.8 | 12 | 30.3 | 37.4 | 450 |
| 2 | 112 | 152 | 10.0 | 6 | 33.5 | 37.0 | 289 |
| 3 | 71 | 91 | 12.0 | 6 | 33.2 | 36.9 | 249 |
| 4 | 71 | 72 | 10.0 | 6 | 36.2 | - | 232 |
| 5 | 68 | 204 | 17.3 | 8 | 36.8 | 35.3 | 319 |
| 6 | 102 | 128 | 19.0 | 7 | 34.9 | 36.9 | 295 |
| 7 | 77 | 115 | 19.3 | 7 | 34.2 | - | 293 |
| 8 | 83 | - | 16.0 | 7 | 36.5 | - | 281 |
| 9 | 140 | - | 23.0 | 5 | 36.9 | 36.9 | 273 |
| 10 | 57 | 173 | 12.0 | 6 | 36.8 | 35.0 | 240 |
| 11 | 91 | 90 | 20.0 | 4 | 33.1 | 35.0 | 203 |
| 12 | 73 | 125 | 20.2 | 5 | 32.3 | 35.1 | 230 |
| 13 | 108 | 135 | 25.0 | 9 | 34.2 | 37.2 | 358 |
| 14 | 112 | 66 | 25.0 | 5 | 34.5 | 36.5 | 261 |
| 15 | 111 | 118 | 20.8 | 5 | 32.0 | 37.2 | 233 |
| 16 | 76 | 102 | 13.2 | 5 | 31.3 | 37.3 | 235 |
| 17 | 210 | 202 | 23.3 | 7 | 28.3 | 35.4 | 415 |
| 18 | 94 | 165 | 20.0 | 6 | 32.2 | 35.7 | 269 |
| 19 | 93 | 64 | 25.0 | 4 | 33.9 | 36.0 | 195 |
| 20 | 111 | 108 | 20.2 | 4 | 32.5 | 36.0 | 227 |
| 21 | 83 | 149 | 20.7 | 6 | 29.8 | 36.9 | 286 |
| mean ± SD | 98 ± 33 | 137 ± 65 | 18.0 ± 5.4 | 6.2 ± 1.9 | 33.5 ± 2.4 | 36.5 ± 0.8 | 278 ± 65 |
| Corrected | Unstimulated Side (n) | Stimulated Side (n) | p-Value |
|---|---|---|---|
| Flux (pmol/s/mg wwt) | |||
| L | 3.5 ± 0.7 (17) | 2.9 ± 0.6 (16) | 0.135 |
| PCI | 9.6 ± 1.0 (19) | 9.1 ± 0.7 (19) | 0.155 |
| PCII | 8. 7 ± 0.9 (19) | 8.1 ± 0.9 (19) | 0.256 |
| PCI + II | 18.3 ± 1.5 (19) | 17.2 ± 1.4 (19) | 0.065 |
| ECI + II | 31.1 ± 3.5 (19) | 27.1 ± 2.1 (19) | 0.026 |
| ECII | 17.7 ± 2.2 (11) | 14.2 ± 1.5 (12) | 0.032 |
| Flux/unit CS activity (pmol/s/CS activity) | |||
| L | 46.3 ± 10.2 (14) | 35.9 ± 7.9 (15) | 0.052 |
| PCI | 123.4 ± 12.6 (15) | 110.3 ± 8.7 (18) | 0.042 |
| PCII | 113.1 ± 15.3 (15) | 100.4 ± 9.0 (18) | 0.504 |
| PCI + II | 236.5 ± 21.5 (15) | 210.7 ± 13.8 (18) | 0.192 |
| ECI + II | 415.6 ± 61.5 (15) | 339.3 ± 29.8 (18) | 0.083 |
| ECII | 231.1 ± 41.7 (9) | 179.0 ± 24.1 (11) | 0.083 |
| Flux Control Ratio (Flux relative to ECI + II) | |||
| L | 0.141 ± 0.038 (17) | 0.122 ± 0.035 (16) | 0.246 |
| PCI | 0.336 ± 0.030 (19) | 0.351 ± 0.026 (19) | 0.389 |
| PCII | 0.296 ± 0.029 (19) | 0.308 ± 0.029 (19) | 0.420 |
| PCI + II | 0.630 ± 0.032 (19) | 0.659 ± 0.036 (19) | 0.134 |
| ECII | 0.493 ± 0.051 (11) | 0.493 ± 0.046 (12) | 0.746 |
| ATP-linked Respiration (PCI) | 6.7 ± 1.1 (17) | 7.2 ± 0.9 (15) | 0.794 |
| ATP-linked Respiration (PCI + II) | 15.4 ± 1.8 (17) | 14.2 ± 1.5 (16) | 0.609 |
| ATP-linked Respiration (PCI/CS) | 82.2 ± 15.4 (14) | 80.3 ± 12.5 (15) | 0.537 |
| ATP-linked Respiration (PCI + II/CS) | 197.0 ± 25.9 (14) | 174.1 ± 17.7 (15) | 0.530 |
| Spare capacity (ECI + II−PCI + II) | 12.8 ± 2.5 (19) | 9.9 ± 1.6 (19) | 0.039 |
| Spare capacity (ECI + II/CS−PCI + II/CS) | 179.0 ± 44.5 (15) | 128.6 ± 22.9 (18) | 0.048 |
| Coupling efficiency (1-L/PCI + II) | 0.792 ± 0.05 (17) | 0.817 ± 0.05 (16) | 0.241 |
| CS activity/mg tissue (mU/mg wwt) | 0.084 ± 0.006 (18) | 0.082 ± 0.004 (20) | 0.626 |
| Mt-aconitase activity (mU/mg protein) | 0.196 ± 0.01 (18) | 0.175 ± 0.01 (20) | 0.060 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mankowski, R.T.; Wohlgemuth, S.E.; Bresciani, G.; Martin, A.D.; Arnaoutakis, G.; Martin, T.; Jeng, E.; Ferreira, L.; Machuca, T.; Rackauskas, M.; et al. Intraoperative Hemi-Diaphragm Electrical Stimulation Demonstrates Attenuated Mitochondrial Function without Change in Oxidative Stress in Cardiothoracic Surgery Patients. Antioxidants 2023, 12, 1009. https://doi.org/10.3390/antiox12051009
Mankowski RT, Wohlgemuth SE, Bresciani G, Martin AD, Arnaoutakis G, Martin T, Jeng E, Ferreira L, Machuca T, Rackauskas M, et al. Intraoperative Hemi-Diaphragm Electrical Stimulation Demonstrates Attenuated Mitochondrial Function without Change in Oxidative Stress in Cardiothoracic Surgery Patients. Antioxidants. 2023; 12(5):1009. https://doi.org/10.3390/antiox12051009
Chicago/Turabian StyleMankowski, Robert T., Stephanie E. Wohlgemuth, Guilherme Bresciani, A. Daniel Martin, George Arnaoutakis, Tomas Martin, Eric Jeng, Leonardo Ferreira, Tiago Machuca, Mindaugas Rackauskas, and et al. 2023. "Intraoperative Hemi-Diaphragm Electrical Stimulation Demonstrates Attenuated Mitochondrial Function without Change in Oxidative Stress in Cardiothoracic Surgery Patients" Antioxidants 12, no. 5: 1009. https://doi.org/10.3390/antiox12051009