
SXRF for Studying the Distribution of Trace Metals in the Pancreas and Liver

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Genotypes and Diet
2.2. Sample Preparation
2.3. Synchrotron X-ray Fluorescence (SXRF) Microscopy
3. Results
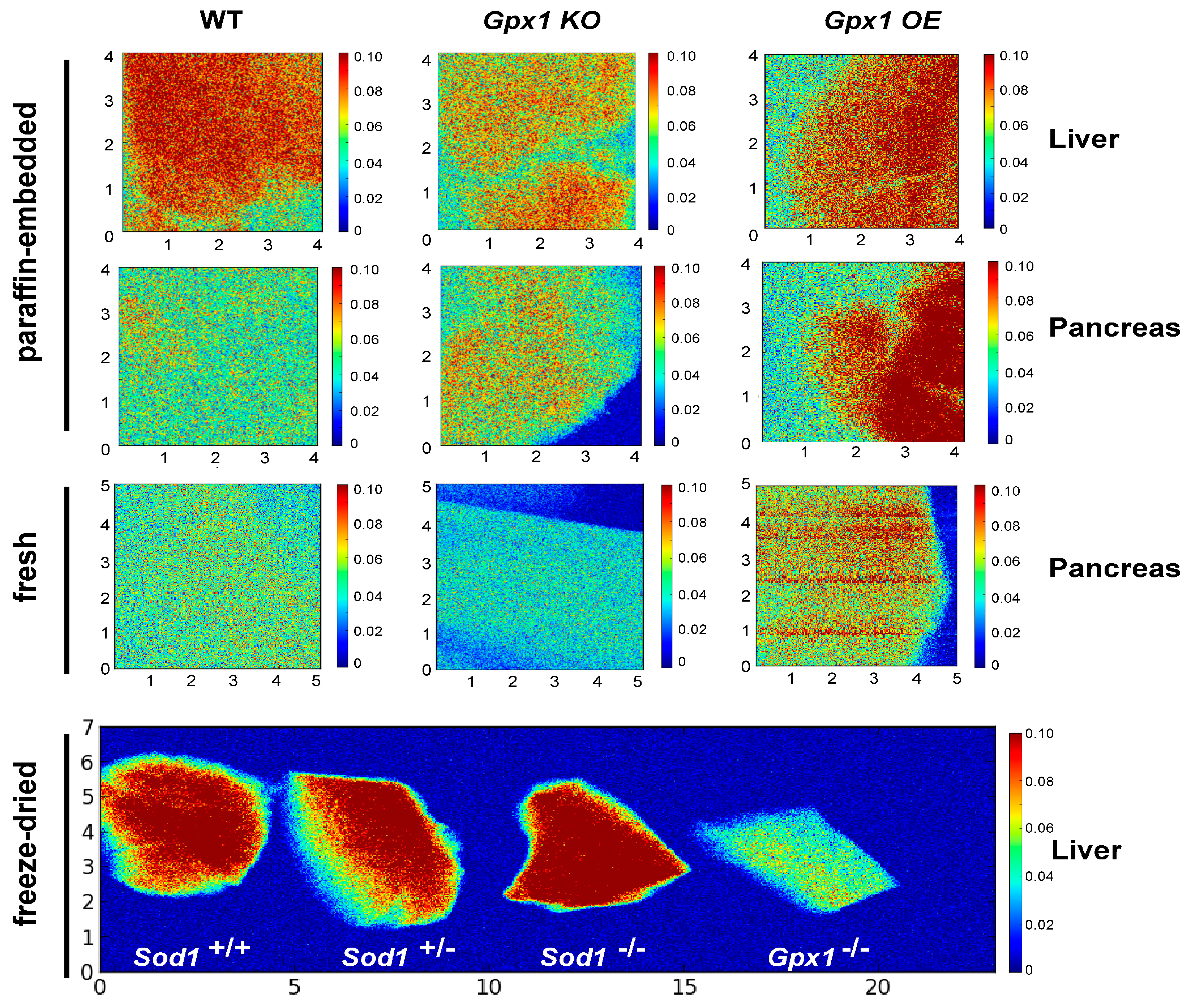
3.1. Selenium in Pancreas and Liver of Gpx1−/− and Gpx1-Overexpressing Mouse Models
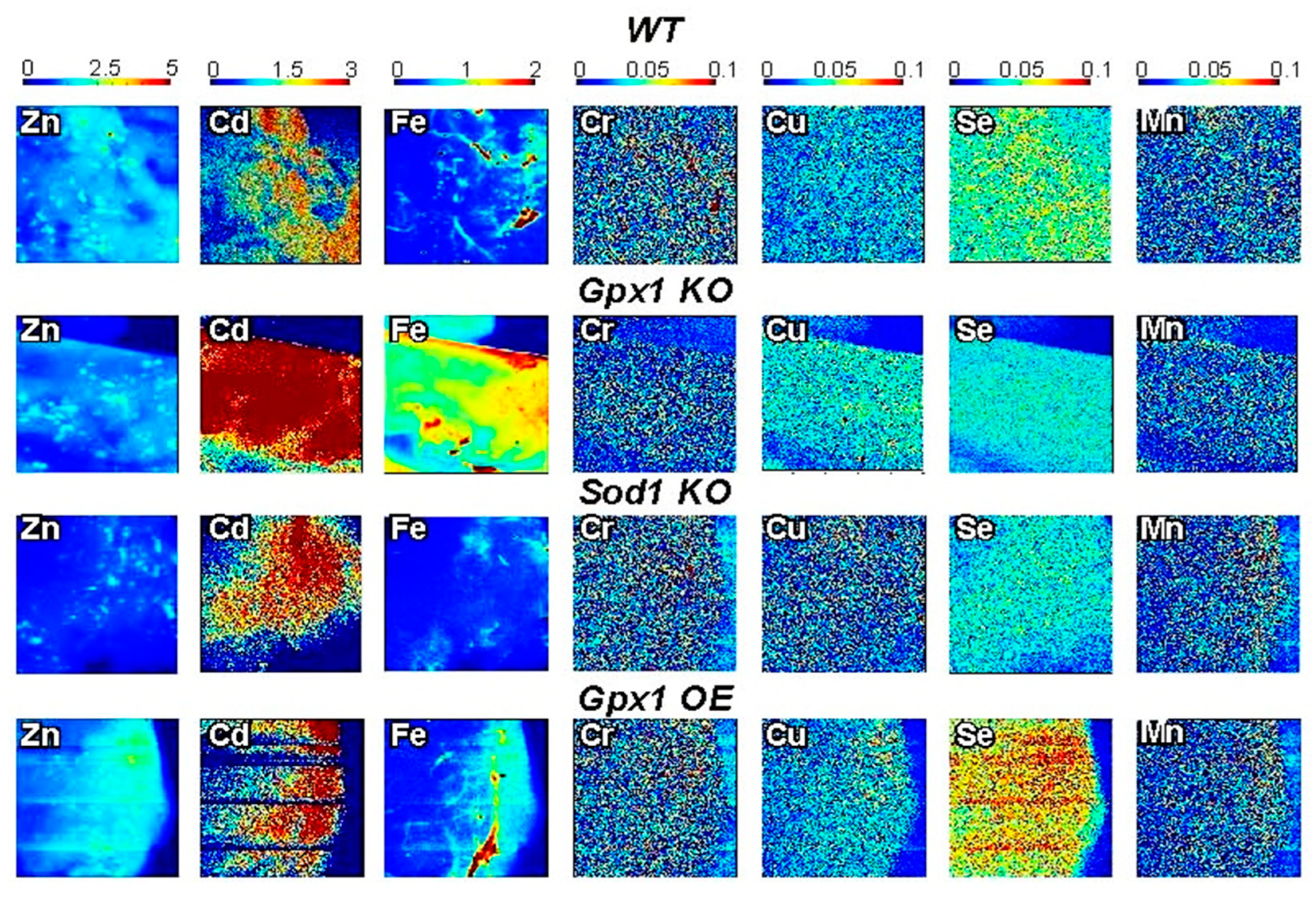
3.2. Micronutrients Visualization in Pancreases of Antioxidant Enzyme-Altered Mice Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McRae, R.; Bagchi, P.; Sumalekshmy, S.; Fahrni, C.J. In situ imaging of metals in cells and tissues. Chem. Rev. 2009, 109, 4780–4827. [Google Scholar] [CrossRef] [Green Version]
- Prashanth, L.; Kattapagari, K.; Chitturi, R.; Baddam, V.; Prasad, L. A review on role of essential trace elements in health and disease. J. Dr. NTR Univ. Health Sci. 2015, 4, 75–85. [Google Scholar] [CrossRef]
- Bongers, A.; van den Heuvel, E. Prebiotics and the Bioavailability of Minerals and Trace Elements. Food Rev. Int. 2003, 19, 397–422. [Google Scholar] [CrossRef]
- Muss, C.; Mosgoeller, W.; Endler, T. Neuroprotective impact of a vitamin trace element composition—A randomized, double blind, placebo controlled clinical trial with healthy volunteers. Neuro Endocrinol. Lett. 2015, 36, 31–40. [Google Scholar]
- Agarwal, A.; Khanna, P.; Baidya, D.K.; Arora, M.K. Trace Elements in Critical Illness. J. Endocrinol. Metab. 2011, 1, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Kaler, S.G. Inborn errors of copper metabolism. Handb. Clin. Neurol. 2013, 113, 1745–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, R.; Shi, Y.; Zhou, H.; Yue, A.; Zhang, L.; Sylvia, S.; Medina, A.; Rozelle, S. Micronutrient deficiencies and developmental delays among infants: Evidence from a cross-sectional survey in rural China. BMJ Open 2015, 5, e008400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, A.; Tamano, H.; Nishio, R.; Murakami, T. Behavioral Abnormality Induced by Enhanced Hypothalamo-Pituitary-Adrenocortical Axis Activity under Dietary Zinc Deficiency and Its Usefulness as a Model. Int. J. Mol. Sci. 2016, 17, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, E.; Kurian, M.A.; Hayflick, S.J. Neurodegeneration with Brain Iron Accumulation: Genetic Diversity and Pathophysiological Mechanisms. Annu. Rev. Genom. Hum. Genet. 2015, 16, 257–279. [Google Scholar] [CrossRef]
- Matschinsky, F.; Ellerman, J. Metabolism of glucose in the islets of Langerhans. J. Biol. Chem. 1968, 243, 2730–2736. [Google Scholar] [CrossRef]
- Jensen, M.V.; Joseph, J.W.; Ronnebaum, S.M.; Burgess, S.C.; Sherry, A.D.; Newgard, C.B. Metabolic cycling in control of glucose-stimulated insulin secretion. Am. J. Physiol. -Endocrinol. Metab. 2008, 295, E1287–E1297. [Google Scholar] [CrossRef] [Green Version]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M. Reactive oxygen species enhance insulin sensitivity. Cell Metab. 2009, 10, 260–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, J.; Thomas, K.A.; Rubin, B.H.; Richardson, D.C. Crystal structure of bovine Cu, Zn superoxide dismutase at 3 A resolution: Chain tracing and metal ligands. Proc. Natl. Acad. Sci. USA 1975, 72, 1349–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, G.C. Hemoglobin catabolism. I. Glutathione peroxidase, an erythrocyte enzyme which protects hemoglobin from oxidative breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [CrossRef]
- Mariotti, M.; Ridge, P.G.; Zhang, Y.; Lobanov, A.V.; Pringle, T.H.; Guigo, R.; Hatfield, D.L.; Gladyshev, V.N. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS ONE 2012, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Vatamaniuk, M.Z.; Roneker, C.A.; Pepper, M.P.; Hu, L.G.; Simmons, R.A.; Lei, X.G. Knockouts of SOD1 and GPX1 exert different impacts on murine islet function and pancreatic integrity. Antioxid. Redox Signal. 2011, 14, 391–401. [Google Scholar] [CrossRef] [Green Version]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, X.G.; Vatamaniuk, M.Z. Two tales of antioxidant enzymes on β cells and diabetes. Antioxid. Redox Signal. 2011, 14, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Vatamaniuk, M.; Wang, S.; Roneker, C.; Simmons, R.; Lei, X. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008, 51, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Flohe, L.; Günzler, W.; Schock, H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Newburger, P.E.; Cohen, H.J. Glutathione peroxidase protein. Absence in selenium deficiency states and correlation with enzymatic activity. J. Clin. Investig. 1986, 77, 1402–1404. [Google Scholar] [CrossRef] [PubMed]
- Weiss Sachdev, S.; Sunde, R. Selenium regulation of transcript abundance and translational efficiency of glutathione peroxidase-1 and -4 in rat liver. Biochem. J. 2001, 357, 851–858. [Google Scholar] [CrossRef]
- Moriarty, P.M.; Reddy, C.C.; Maquat, L.E. Selenium Deficiency Reduces the Abundance of mRNA for Se-Dependent Glutathione Peroxidase 1 by a UGA-Dependent Mechanism Likely To Be Nonsense Codon-Mediated Decay of Cytoplasmic mRNA. Mol. Cell. Biol. 1998, 18, 2932–2939. [Google Scholar] [CrossRef] [Green Version]
- Pepper, M.; Vatamaniuk, M.; Yan, X.; Roneker, C.; Lei, X. Impacts of dietary selenium deficiency on metabolic phenotypes of diet-restricted GPX1-overexpressing mice. Antioxid. Redox Signal. 2011, 14, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Pepper, M.P.; Vatamaniuk, M.Z.; Roneker, C.A.; Li, L.; Lei, X.G. Dietary selenium deficiency partially rescues type 2 diabetes-like phenotypes of glutathione peroxidase-1-overexpressing male mice. J. Nutr. 2012, 142, 1975–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.-S.; Magnenat, J.-L.; Bronson, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.P. Cryofixation of tissues for electron microscopy: A review of plunge cooling methods. Scanning Microsc. 1992, 6, 715–743. [Google Scholar]
- Solé, V.A.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A multiplatform code for the analysis of energy-dispersive X-ray fluorescence spectra. Spectrochim. Acta Part. B At. Spectrosc. 2007, 62, 63–68. [Google Scholar] [CrossRef]
- Khan, A.R.; Awan, F.R. Metals in the pathogenesis of type 2 diabetes. J. Diabetes Metab. Disord. 2014, 13, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiernsperger, N.; Rapin, J. Trace elements in glucometabolic disorders: An update. Diabetol. Metab. Syndr. 2010, 2, 70. [Google Scholar] [CrossRef] [Green Version]
- Flegal, A.R.; Smith, D.R. Measurements of environmental lead contamination and human exposure. Rev. Environ. Contam. Toxicol. 1995, 143, 1–45. [Google Scholar] [PubMed]
- Harrington, J.M.; Young, D.J.; Essader, A.S.; Sumner, S.J.; Levine, K.E. Analysis of Human Serum and Whole Blood for Mineral Content by ICP-MS and ICP-OES: Development of a Mineralomics Method. Biol. Trace Elem. Res. 2014, 160, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, K.P.; Young, A.M.; Palmer, A.E. Fluorescent sensors for measuring metal ions in living systems. Chem. Rev. 2014, 114, 4564–4601. [Google Scholar] [CrossRef] [PubMed]
- Ascone, I.; Strange, R. Biological X-ray absorption spectroscopy and metalloproteomics. J. Synchrotron Radiat. 2009, 16, 413–421. [Google Scholar] [CrossRef]
- Finney, L.; Mandava, S.; Ursos, L.; Zhang, W.; Rodi, D.; Vogt, S.; Legnini, D.; Maser, J.; Ikpatt, F.; Olopade, O.I.; et al. X-ray fluorescence microscopy reveals large-scale relocalization and extracellular translocation of cellular copper during angiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Finney, L.A.; O’Halloran, T.V. Transition Metal Speciation in the Cell: Insights from the Chemistry of Metal Ion Receptors. Science 2003, 300, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paunesku, T.; Vogt, S.; Maser, J.; Lai, B.; Woloschak, G. X-ray fluorescence microprobe imaging in biology and medicine. J. Cell. Biochem. 2006, 99, 1489–1502. [Google Scholar] [CrossRef]
- De Samber, B.; Bensellam, M.; Van Malderen, S.J.; Seiboth, F.; Brückner, D.; Garrevoet, J.; Falkenberg, G.; Jonas, J.-C.; Vincze, L. Proof-of-concept for 2D/CT element analysis of entire cryofrozen islets of Langerhans using a cryoloop synchrotron X-ray fluorescence setup. J. Anal. At. Spectrom. 2020, 35, 1368–1379. [Google Scholar] [CrossRef]
- Slepchenko, K.G.; Chen, S.; Corbin, K.L.; Colvin, R.A.; Nunemaker, C.S. The use of synchrotron X-ray fluorescent imaging to study distribution and content of elements in chemically fixed single cells: A case study using mouse pancreatic beta-cells. Metallomics 2023, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Bai, Y.; Zhang, Q.; Wong, V.; Floering, L.M.; Daniel, K.; Reece, J.M.; Deeney, J.T.; Andersen, M.E.; Corkey, B.E.; et al. Reactive Oxygen Species as a Signal in Glucose-Stimulated Insulin Secretion. Diabetes 2007, 56, 1783–1791. [Google Scholar] [CrossRef] [Green Version]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.J.; Mahadev, K.; Wu, X. Redox Paradox: Insulin Action Is Facilitated by Insulin-Stimulated Reactive Oxygen Species With Multiple Potential Signaling Targets. Diabetes 2005, 54, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Steinbrenner, H. Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism. Free Radic. Biol. Med. 2013, 65, 1538–1547. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Lee, B.C.; Handy, D.E.; Loscalzo, J.; Hatfield, D.L.; Gladyshev, V.N. Both Maximal Expression of Selenoproteins and Selenoprotein Deficiency Can Promote Development of Type 2 Diabetes-like Phenotype in Mice. Antioxid. Redox Signal. 2011, 14, 2327–2336. [Google Scholar] [CrossRef]
- Chimienti, F. Zinc, pancreatic islet cell function and diabetes: New insights into an old story. Nutr. Res. Rev. 2013, 26, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Seve, M.; Chimienti, F.; Favier, A. Role of intracellular zinc in programmed cell death. Pathol. Biol. 2002, 50, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Chang, I.; Cho, N.; Koh, J.Y.; Lee, M.S. Pyruvate inhibits zinc-mediated pancreatic islet cell death and diabetes. Diabetologia 2003, 46, 1220–1227. [Google Scholar] [CrossRef] [Green Version]
- Boden, M.J.; Brandon, A.E.; Tid-Ang, J.D.; Preston, E.; Wilks, D.; Stuart, E.; Cleasby, M.E.; Turner, N.; Cooney, G.J.; Kraegen, E.W. Overexpression of manganese superoxide dismutase ameliorates high-fat diet-induced insulin resistance in rat skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E798–E805. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Rodriguez, E.; Bermejo, L.M.; Lopez-Sobaler, A.M.; Ortega, R.M. An inadequate intake of manganese may favour insulin resistance in girls. Nutr. Hosp. 2011, 26, 965–970. [Google Scholar] [CrossRef]
- Li, M.; Wang, S.; Liu, X.; Sheng, Z.; Li, B.; Li, J.; Zhang, J.; Zhang, Z. Cadmium exposure decreases fasting blood glucose levels and exacerbates type-2 diabetes in a mouse model. Endocrine 2022, 76, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, R.; Olsen, A.; Nguyen, J.; Wong, W.; El Muayed, M.; Edwards, J. Pancreatic Islets Accumulate Cadmium in a Rodent Model of Cadmium-Induced Hyperglycemia. Int. J. Mol. Sci. 2020, 22, 360. [Google Scholar] [CrossRef] [PubMed]
- Simcox, J.A.; McClain, D.A. Iron and diabetes risk. Cell Metab. 2013, 17, 329–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havel, P.J. A scientific review: The role of chromium in insulin resistance. Diabetes Educ. 2004, 30 (Suppl. 3), 2–14. [Google Scholar]
- Ather Ali, N.D.; Ma, Y.; Reynolds, J.; Wise, J.P.; Inzucchi, S.E.; Katz, D.L. Chromium Effects on Glucose Tolerance and Insulin Sensitivity in Persons at Risk for Diabetes Mellitus. Endocr. Pract. 2011, 17, 16–25. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vatamaniuk, M.Z.; Huang, R.; Zhao, Z.; Lei, X.G. SXRF for Studying the Distribution of Trace Metals in the Pancreas and Liver. Antioxidants 2023, 12, 846. https://doi.org/10.3390/antiox12040846
Vatamaniuk MZ, Huang R, Zhao Z, Lei XG. SXRF for Studying the Distribution of Trace Metals in the Pancreas and Liver. Antioxidants. 2023; 12(4):846. https://doi.org/10.3390/antiox12040846
Chicago/Turabian StyleVatamaniuk, Marko Z., Rong Huang, Zeping Zhao, and Xin Gen Lei. 2023. "SXRF for Studying the Distribution of Trace Metals in the Pancreas and Liver" Antioxidants 12, no. 4: 846. https://doi.org/10.3390/antiox12040846