
Interdependencies of Gene Expression and Function between Two Redox Enzymes and REG Family Proteins in Murine Pancreatic Islets and Human Pancreatic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Tissue Sample Collection, and Islet Isolation
2.2. In Vitro Islets Treatment and Proliferation Assay
2.3. Cell Culture and Viability Assay
2.4. Quantitative Real-Time PCR and Enzyme Activity Assays
2.5. Statistical Analysis
3. Results
3.1. Effects of Knockouts of Gpx1 and/or Sod1 on Expression of Reg Family Genes in Mouse Islets
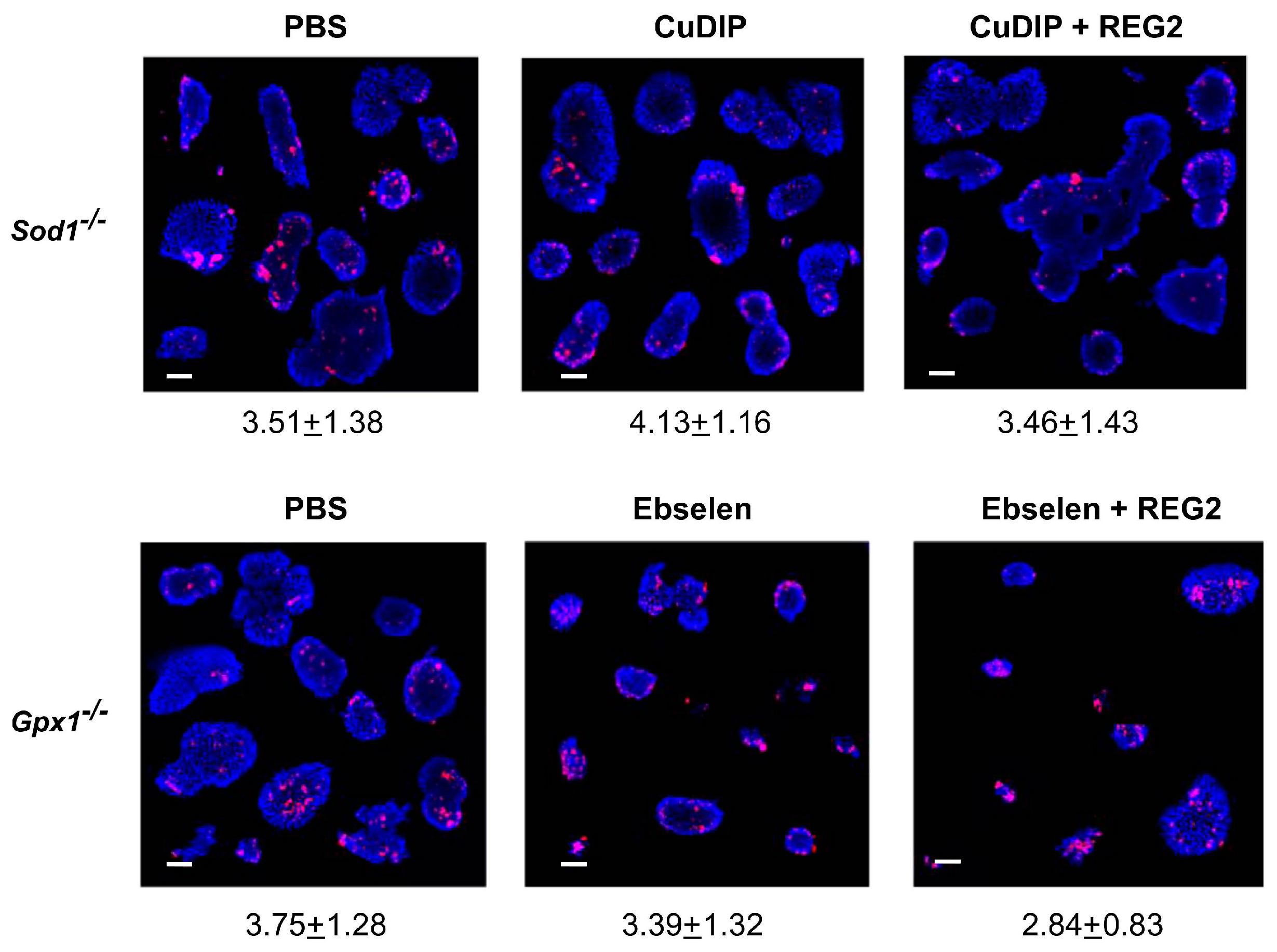
3.2. Impacts of GPX1 and SOD1 Status on Inhibitions of Islet Proliferation by Exogenous REG2 Protein
3.3. Effects of Murine REG2 Protein on REG Gene Expression and Antioxidant Enzyme Activities in PANC1 Cells
3.4. Effects of Murine REG2 Protein on Cell Viability and Its Dependence on Free Calcium in PANC1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Pi, J.; Bai, Y.; Zhang, Q.; Wong, V.; Floering, L.M.; Daniel, K.; Reece, J.M.; Deeney, J.T.; Andersen, M.E.; Corkey, B.E.; et al. Reactive oxygen species as a signal in glucose-stimulated insulin secretion. Diabetes 2007, 56, 1783–1791. [Google Scholar] [CrossRef] [Green Version]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Mills, G.C. Hemoglobin catabolism. I. Glutathione peroxidase, an erythrocyte enzyme which protects hemoglobin from oxidative breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [CrossRef]
- Flohe, L.; Gunzler, W.; Schock, H. Glutathione peroxidase: A selenoenzyme. FEBS Lett. 1973, 32, 132–134. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Superoxide radical and superoxide dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef]
- Cheng, W.H.; Ho, Y.S.; Ross, D.A.; Han, Y.; Combs, G.F., Jr.; Lei, X.G. Overexpression of cellular glutathione peroxidase does not affect expression of plasma glutathione peroxidase or phospholipid hydroperoxide glutathione peroxidase in mice offered diets adequate or deficient in selenium. J. Nutr. 1997, 127, 675–680. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.G.; Zhu, J.H.; McClung, J.P.; Aregullin, M.; Roneker, C.A. Mice deficient in Cu,Zn-superoxide dismutase are resistant to acetarninophen toxicity. Biochem. J. 2006, 399, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, X.G.; Vatamaniuk, M.Z. Two tales of antioxidant enzymes on β cells and diabetes. Antioxid. Redox Signal. 2011, 14, 489–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.D.; Vatamaniuk, M.Z.; Roneker, C.A.; Pepper, M.P.; Hu, L.G.; Simmons, R.A.; Lei, X.G. Knockouts of SOD1 and GPX1 exert different impacts on murine islet function and pancreatic integrity. Antioxid. Redox Signal. 2011, 14, 391–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.D.; Vatamaniuk, M.Z.; Wang, S.K.; Roneker, C.A.; Simmons, R.A.; Lei, X.G. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008, 51, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.W.; Zhao, Z.; Yan, X.; Vatamaniuk, M.Z.; Lei, X.G. Glutathione peroxidase-1 inhibits transcription of regenerating islet-derived protein-2 in pancreatic islets. Free Radic. Biol. Med. 2019, 134, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; List, E.O.; Kopchick, J.J. Differentially expressed proteins in the pancreas of diet-induced diabetic mice. Mol. Cell Proteom. 2005, 4, 1311–1318. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Cui, W. Which gene, Reg2 or Reg3beta, was targeted that affected liver regeneration? Hepatology 2007, 45, 1584–1585. [Google Scholar] [CrossRef]
- Narushima, Y.; Unno, M.; Nakagawara, K.; Mori, M.; Miyashita, H.; Suzuki, Y.; Noguchi, N.; Takasawa, S.; Kumagai, T.; Yonekura, H.; et al. Structure, chromosomal localization and expression of mouse genes encoding type III Reg, RegIII alpha, RegIII beta, RegIII gamma. Gene 1997, 185, 159–168. [Google Scholar] [CrossRef]
- Terazono, K.; Yamamoto, H.; Takasawa, S.; Shiga, K.; Yonemura, Y.; Tochino, Y.; Okamoto, H. A novel gene activated in regenerating islets. J. Biol. Chem. 1988, 263, 2111–2114. [Google Scholar] [CrossRef]
- Huszarik, K.; Wright, B.; Keller, C.; Nikoopour, E.; Krougly, O.; Lee-Chan, E.; Qin, H.Y.; Cameron, M.J.; Gurr, W.K.; Hill, D.J.; et al. Adjuvant immunotherapy increases beta cell regenerative factor Reg2 in the pancreas of diabetic mice. J. Immunol. 2010, 185, 5120–5129. [Google Scholar] [CrossRef] [Green Version]
- Lieu, H.T.; Simon, M.T.; Nguyen-Khoa, T.; Kebede, M.; Cortes, A.; Tebar, L.; Smith, A.J.; Bayne, R.; Hunt, S.P.; Bréchot, C.; et al. Reg2 inactivation increases sensitivity to Fas hepatotoxicity and delays liver regeneration post-hepatectomy in mice. Hepatology 2006, 44, 1452–1464. [Google Scholar] [CrossRef]
- Baeza, N.J.; Moriscot, C.I.; Renaud, W.P.; Okamoto, H.; Figarella, C.G.; Vialettes, B.H. Pancreatic regenerating gene overexpression in the nonobese diabetic mouse during active diabetogenesis. Diabetes 1996, 45, 67–70. [Google Scholar] [CrossRef]
- Cavard, C.; Terris, B.; Grimber, G.; Christa, L.; Audard, V.; Radenen-Bussiere, B.; Simon, M.T.; Renard, C.A.; Buendia, M.A.; Perret, C. Overexpression of regenerating islet-derived 1 alpha and 3 alpha genes in human primary liver tumors with beta-catenin mutations. Oncogene 2006, 25, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.C.; Ferrara, J.L.; Braun, T.M.; Holler, E.; Teshima, T.; Levine, J.E.; Choi, S.W.; Landfried, K.; Akashi, K.; Vander Lugt, M.; et al. Plasma biomarkers of lower gastrointestinal and liver acute GVHD. Blood 2012, 119, 2960–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurr, W.; Shaw, M.; Li, Y.; Sherwin, R. RegII is a beta-cell protein and autoantigen in diabetes of NOD mice. Diabetes 2007, 56, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, B.; Miao, X.; Ramgattie, C.; Gao, Z.H.; Liu, J.L. Reg2 expression is required for pancreatic islet compensation in response to aging and high-fat diet-induced obesity. Endocrinology 2017, 158, 1634–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duplan, L.; Michel, B.; Boucraut, J.; Barthellémy, S.; Desplat-Jego, S.; Marin, V.; Gambarelli, D.; Bernard, D.; Berthézène, P.; Alescio-Lautier, B.; et al. Lithostathine and pancreatitis-associated protein are involved in the very early stages of Alzheimer’s disease. Neurobiol. Aging 2001, 22, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Gargano, M.; Cao, J.; Bronson, R.T.; Heimler, I.; Hutz, R.J. Reduced fertility in female mice lacking copper-zinc superoxide dismutase. J. Biol. Chem. 1998, 273, 7765–7769. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.S.; Magnenat, J.L.; Bronson, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Zhao, Z.; Weaver, J.; Sun, T.; Yun, J.W.; Roneker, C.A.; Hu, F.; Doliba, N.M.; McCormick, C.C.W.; Vatamaniuk, M.Z.; et al. Role and mechanism of REG2 depletion in insulin secretion augmented by glutathione peroxidase-1 overproduction. Redox Biol. 2022, 56, 102457. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.R.; Nagar, B.; Young, N.M.; Hirama, T.; Rini, J.M. X-ray crystal structure of a galactose-specific C-type lectin possessing a novel decameric quaternary structure. Biochemistry 2004, 43, 3783–3792. [Google Scholar] [CrossRef]
- Sun, L.H.; Li, J.G.; Zhao, H.; Shi, J.; Huang, J.Q.; Wang, K.N.; Xia, X.J.; Li, L.; Lei, X.G. Porcine serum can be biofortified with selenium to inhibit proliferation of three types of human cancer cells. J. Nutr. 2013, 143, 1115–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, R.A.; Sunde, R.A.; Schwartz, G.L.; Hoekstra, W.G. Glutathione peroxidase activity in rat lens and other tissues in relation to dietary selenium intake. Exp. Eye Res. 1974, 18, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.M.; Buss, J.; Easley, R.; Yang, Z.; Korpe, P.S.; Niu, F.; Ma, J.Z.; Olortegui, M.P.; Haque, R.; Kosek, M.N.; et al. REG1B as a predictor of childhood stunting in Bangladesh and Peru. Am. J. Clin. Nutr. 2013, 97, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, A.; Itaya-Hironaka, A.; Sakuramoto-Tsuchida, S.; Takeda, M.; Yoshimoto, K.; Miyaoka, T.; Fujimura, T.; Tsujinaka, H.; Tsuchida, C.; Ota, H.; et al. Synergistic activations of REGIα and REGIβ promoters by IL-6 and glucocorticoids through JAK/STAT pathway in human pancreatic β cells. J. Diabetes Res. 2015, 2015, 173058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Beelen Granlund, A.; Østvik, A.E.; Brenna, Ø.; Torp, S.H.; Gustafsson, B.I.; Sandvik, A.K. REG gene expression in inflamed and healthy colon mucosa explored by in situ hybridisation. Cell Tissue Res. 2013, 352, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Lenzen, S.; Drinkgern, J.; Tiedge, M. Low antioxidant enzyme gene expression in pancreatic islets compared with various other mouse tissues. Free Radic. Biol. Med. 1996, 20, 463–466. [Google Scholar] [CrossRef]
- Abe, M.; Nata, K.; Akiyama, T.; Shervani, N.J.; Kobayashi, S.; Tomioka-Kumagai, T.; Ito, S.; Takasawa, S.; Okamoto, H. Identification of a novel Reg family gene, Reg IIIδ, and mapping of all three types of Reg family gene in a 75 kilobase mouse genomic region. Gene 2000, 246, 111–122. [Google Scholar] [CrossRef]
- Miyashita, H.; Nakagawara, K.; Mori, M.; Narushima, Y.; Noguchi, N.; Moriizumi, S.; Takasawa, S.; Yonekura, H.; Takeuchi, T.; Okamoto, H. Human REG family genes are tandemly ordered in a 95-kilobase region of chromosome 2p12. FEBS Lett. 1995, 377, 429–433. [Google Scholar]
- Hartupee, J.C.; Zhang, H.; Bonaldo, M.F.; Soares, M.B.; Dieckgraefe, B.K. Isolation and characterization of a cDNA encoding a novel member of the human regenerating protein family: Reg IV. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1518, 287–293. [Google Scholar] [CrossRef]
- De León, D.D.; Farzad, C.; Crutchlow, M.F.; Brestelli, J.; Tobias, J.; Kaestner, K.H.; Stoffers, D.A. Identification of transcriptional targets during pancreatic growth after partial pancreatectomy and exendin-4 treatment. Physiol. Genom. 2006, 24, 133–143. [Google Scholar] [CrossRef]
- Li, B.; Wang, X.; Liu, J.L. Pancreatic acinar-specific overexpression of Reg2 gene offered no protection against either experimental diabetes or pancreatitis in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G413–G421. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chowdhury, S.; Uppal, S.; Fang, X.; Liu, J.L.; Srikant, C.B. mReg2 inhibits nuclear entry of apoptosis-inducing factor in mouse insulinoma cells. Growth Factors 2015, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mirochnitchenko, O.; Palnitkar, U.; Philbert, M.; Inouye, M. Thermosensitive phenotype of transgenic mice overproducing human glutathione peroxidases. Proc. Natl. Acad. Sci. USA 1995, 92, 8120–8124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alenzi, F.Q. Links between apoptosis, proliferation and the cell cycle. Br. J. Biomed. Sci. 2004, 61, 99–102. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, X.; Li, B.; Chowdhury, S.; Lu, Y.; Srikant, C.B.; Ning, G.; Liu, J.L. Pancreatic islet-specific overexpression of Reg3beta protein induced the expression of pro-islet genes and protected the mice against streptozotocin-induced diabetes mellitus. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E669–E680. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, Y.; Xie, J.; Li, C.; Wang, X.; Shen, J.; Zhang, Y.; Wang, S.; Cheng, N. Regenerating gene 1B silencing inhibits colon cancer cell HCT116 proliferation and invasion. Int. J. Biol. Markers 2015, 30, 217–225. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, R.; Wang, L.; Shen, S.; Okamoto, H.; Sugawara, A.; Xia, L.; Wang, X.; Noguchi, N.; Yoshikawa, T.; et al. Upregulation of REG Ialpha accelerates tumor progression in pancreatic cancer with diabetes. Int. J. Cancer 2010, 127, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Gironella, M.; Calvo, C.; Fernández, A.; Closa, D.; Iovanna, J.L.; Rosello-Catafau, J.; Folch-Puy, E. Reg3beta deficiency impairs pancreatic tumor growth by skewing macrophage polarization. Cancer Res. 2013, 73, 5682–5694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wei, W.; Li, X.; Shen, P.; Ju, D.; Wang, Z.; Zhang, R.; Yang, F.; Chen, C.; Cao, K. BMI1 and MEL18 promote colitis-associated cancer in mice via REG3B and STAT3. Gastroenterology 2017, 153, 1607–1620. [Google Scholar] [CrossRef] [Green Version]
- Gironella, M.; Folch-Puy, E.; LeGoffic, A.; Garcia, S.; Christa, L.; Smith, A.; Tebar, L.; Hunt, S.P.; Bayne, R.; Smith, A.J. Experimental acute pancreatitis in PAP/HIP knock-out mice. Gut 2007, 56, 1091–1097. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Kandil, E.; Lin, Y.Y.; Levi, G.; Zenilman, M.E. Targeted inhibition of gene expression of pancreatitis-associated proteins exacerbates the severity of acute pancreatitis in rats. Scand. J. Gastroenterol. 2004, 39, 870–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Vatamaniuk, M.Z.; Zhao, Z.; Lei, X.G. Interdependencies of Gene Expression and Function between Two Redox Enzymes and REG Family Proteins in Murine Pancreatic Islets and Human Pancreatic Cells. Antioxidants 2023, 12, 849. https://doi.org/10.3390/antiox12040849
Wang H, Vatamaniuk MZ, Zhao Z, Lei XG. Interdependencies of Gene Expression and Function between Two Redox Enzymes and REG Family Proteins in Murine Pancreatic Islets and Human Pancreatic Cells. Antioxidants. 2023; 12(4):849. https://doi.org/10.3390/antiox12040849
Chicago/Turabian StyleWang, Hong, Marko Z. Vatamaniuk, Zeping Zhao, and Xin Gen Lei. 2023. "Interdependencies of Gene Expression and Function between Two Redox Enzymes and REG Family Proteins in Murine Pancreatic Islets and Human Pancreatic Cells" Antioxidants 12, no. 4: 849. https://doi.org/10.3390/antiox12040849