
Tissue-Specific Antioxidant Activities of Germinated Seeds in Lentil Cultivars during Thermal Processing

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Sample Processing
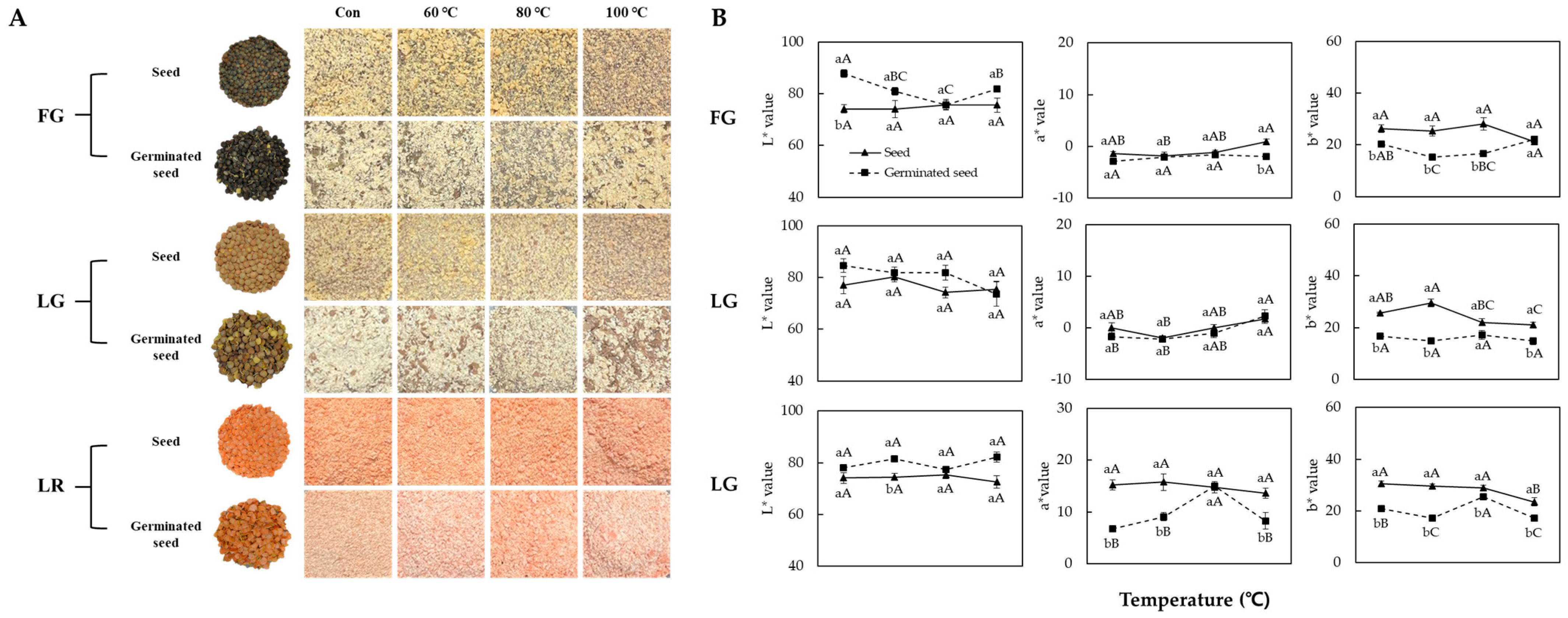
2.3. Measurement of Color Variation
2.4. Sample Extraction
2.5. Measurement of TPC and TFC
2.6. HPLC Analysis
2.7. Measurement of Antioxidant Activity
2.8. Statistical Analysis
3. Results and Discussion
3.1. Morphology and Color Variation
3.2. Changes in Total Phenolic Content
3.3. Changes in Flavonoid Composition and Content
3.3.1. Changes of Total Flavonoid Content
3.3.2. Changes in Major Flavonoid Contents
3.4. Changes in Antioxidant Activity
3.4.1. The Effects of Germination on Antioxidant Activities Variation in Three Different Cultivars of Lentils
3.4.2. The Effects of Thermal Processing on Antioxidant Activities Variation in Different Cultivar of Lentils
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NGS | Nongerminated Seeds |
| GS | Germinated Seeds |
| AA | Antioxidant Activity |
| FG | French Green |
| LG | Lentil Green |
| LR | Lentil Red |
| TPC | Total Phenolic Content |
| TFC | Total Flavonoid Content |
References
- Alkaltham, M.S.; Musa Özcan, M.; Uslu, N.; Salamatullah, A.M.; Hayat, K. Changes in antioxidant activity, phenolic compounds, fatty acids, and mineral contents of raw, germinated, and boiled lentil seeds. J. Food Sci. 2022, 87, 1639–1649. [Google Scholar] [CrossRef] [PubMed]
- Pal, R.S.; Bhartiya, A.; Yadav, P.; Kant, L.; Mishra, K.K.; Aditya, J.P.; Pattanayak, A. Effect of dehulling, germination and cooking on nutrients, anti-nutrients, fatty acid composition and antioxidant properties in lentil (Lens culinaris). J. Food Sci. Technol. 2017, 54, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Gallo, V.; Ferranti, P.; Masi, P. Lentil flour: Nutritional and technological properties, in vitro digestibility and perspectives for use in the food industry. Curr. Opin. Food Sci. 2021, 40, 157–167. [Google Scholar] [CrossRef]
- Hefnawy, T.H. Effect of processing methods on nutritional composition and anti-nutritional factors in lentils (Lens culinaris). Ann. Agric. Sci. 2011, 56, 57–61. [Google Scholar] [CrossRef]
- Nikmaram, N.; Leong, S.Y.; Koubaa, M.; Zhu, Z.; Barba, F.J.; Greiner, R.; Oey, I.; Roohinejad, S. Effect of extrusion on the anti-nutritional factors of food products: An overview. Food Control 2017, 79, 62–73. [Google Scholar] [CrossRef]
- Joshi, M.; Timilsena, Y.; Adhikari, B. Global production, processing and utilization of lentil: A review. J. Integr. Agric. 2017, 16, 2898–2913. [Google Scholar] [CrossRef]
- Vidal-Valverde, C.; Frias, J.; Sierra, I.; Blazquez, I.; Lambein, F.; Kuo, Y.H. New functional legume foods by germination: Effect on the nutritive value of beans, lentils and peas. Eur. Food Res. Technol. 2002, 215, 472–477. [Google Scholar] [CrossRef]
- Aguilera, Y.; Díaz, M.F.; Jiménez, T.; Benítez, V.; Herrera, T.; Cuadrado, C.; Martín-Pedrosa, M.; Martín-Cabrejas, M.A. Changes in Nonnutritional Factors and Antioxidant Activity during Germination of Nonconventional Legumes. J. Agric. Food Chem. 2013, 61, 8120–8125. [Google Scholar] [CrossRef]
- Lim, Y.J.; Jeong, H.Y.; Gil, C.S.; Kwon, S.J.; Na, J.K.; Lee, C.; Eom, S.H. Isoflavone accumulation and the metabolic gene expression in response to persistent UV-B irradiation in soybean sprouts. Food Chem. 2020, 303, 125376. [Google Scholar] [CrossRef]
- Lim, Y.J.; Lyu, J.I.; Kwon, S.-J.; Eom, S.H. Effects of UV-A radiation on organ-specific accumulation and gene expression of isoflavones and flavonols in soybean sprout. Food Chem. 2021, 339, 128080. [Google Scholar] [CrossRef]
- Nam, T.G.; Kim, D.O.; Eom, S.H. Effects of light sources on major flavonoids and antioxidant activity in common buckwheat sprouts. Food Sci. Biotechnol. 2018, 27, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.S.; Silva, B.; Valente, L.M.P.; Gruber, S.; Vasconcelos, M.W. The effect of sprouting in lentil (Lens culinaris) nutritional and microbiological profile. Food 2020, 9, 400. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.A.; Rehab, F.M. Effect of germination time on proximate analysis, bioactive compounds and antioxidant activity of lentil (Lens culinaris Medik.) sprouts. Acta Sci. Pol. Technol. Aliment. 2015, 14, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, B.; Estrella, I.; Hernandez, T. Changes in phenolic compounds in lentils (Lens culinaris) during germination and fermentation. Z. Für Lebensm. Und-Forsch. A 1997, 205, 290–294. [Google Scholar] [CrossRef]
- Aguilera, Y.; Liébana, R.; Herrera, T.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Benítez, V.; Martín-Cabrejas, M.A. Effect of Illumination on the Content of Melatonin, Phenolic Compounds, and Antioxidant Activity During Germination of Lentils (Lens culinaris L.) and Kidney Beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2014, 62, 10736–10743. [Google Scholar] [CrossRef]
- Avilés-Gaxiola, S.; Chuck-Hernández, C.; Serna Saldívar, S.O. Inactivation Methods of Trypsin Inhibitor in Legumes: A Review. J. Food Sci. 2018, 83, 17–29. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Kilonzi, S.M.; Shabbir, U.; Oh, D.H. Edible Plant Sprouts: Health Benefits, Trends, and Opportunities for Novel Exploration. Nutrients 2021, 13, 2882. [Google Scholar] [CrossRef]
- Razavi, S.N.; Hojjatoleslamy, M.; Molavi, H.; Boroujeni, L.S. The Effect of Germinated Lentil Flour on the Physicochemical and Organoleptic Characteristics of Sangak Bread. J. Culin. Sci. Technol. 2022, 20, 253–265. [Google Scholar] [CrossRef]
- Hernandez-Aguilar, C.; Dominguez-Pacheco, A.; Palma Tenango, M.; Valderrama-Bravo, C.; Soto Hernández, M.; Cruz-Orea, A.; Ordonez-Miranda, J. Lentil sprouts: A nutraceutical alternative for the elaboration of bread. J. Food Sci. Technol. 2020, 57, 1817–1829. [Google Scholar] [CrossRef]
- Atudorei, D.; Mironeasa, S.; Codină, G.G. Effects of Germinated Lentil Flour on Dough Rheological Behavior and Bread Quality. Foods 2022, 11, 2982. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Shah, W.H. Thermal heat processing effects on antinutrients, protein and starch digestibility of food legumes. Food Chem. 2005, 91, 327–331. [Google Scholar] [CrossRef]
- Udensi, E.A.; Ekwu, F.C.; Isinguzo, J.N. Antinutrient factors of vegetable cowpea (Sesquipedalis) seeds during thermal processing. Pak. J. Nutr. 2007, 6, 194–197. [Google Scholar]
- Atudorei, D.; Stroe, S.G.; Codină, G.G. Physical, physiological and minerals changes of different legumes types during the germination process. Food Technol. 2020, 9, 844–863. [Google Scholar] [CrossRef]
- Duan, S.; Kwon, S.J.; Gil, C.S.; Eom, S.H. Improving the Antioxidant Activity and Flavor of Faba (Vicia faba L.) Leaves by Domestic Cooking Methods. Antioxidants 2022, 11, 931. [Google Scholar] [CrossRef]
- Lim, Y.J.; Eom, S.H. The Different Contributors to Antioxidant Activity in Thermally Dried Flesh and Peel of Astringent Persimmon Fruit. Antioxidants 2022, 11, 597. [Google Scholar] [CrossRef]
- Lim, Y.J.; Kwon, S.-J.; Qu, S.; Kim, D.-G.; Eom, S.H. Antioxidant contributors in seed, seed coat, and cotyledon of γ-ray-induced soybean mutant lines with different seed coat colors. Antioxidants 2021, 10, 353. [Google Scholar] [CrossRef]
- Duan, S.; Liu, J.R.; Wang, X.; Sun, X.M.; Gong, H.S.; Jin, C.W.; Eom, S.H. Thermal control using far-infrared irradiation for Producing deglycosylated bioactive compounds from Korean ginseng leaves. Molecules 2022, 27, 4782. [Google Scholar] [CrossRef]
- Duan, S.C.; Kwon, S.J.; Eom, S.H. Effect of thermal processing on color, phenolic compounds, and antioxidant activity of faba bean (Vicia faba L.) leaves and seeds. Antioxidants 2021, 10, 1207. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Gałkowska, D.; Rożnowski, J.; Fortuna, T. Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef]
- Francis, H.; Debs, E.; Koubaa, M.; Alrayess, Z.; Maroun, R.G.; Louka, N. Sprouts Use as Functional Foods. Optimization of Germination of Wheat (Triticum aestivum L.), Alfalfa (Medicago sativa L.), and Radish (Raphanus sativus L.) Seeds Based on Their Nutritional Content Evolution. Foods 2022, 11, 1460. [Google Scholar] [CrossRef]
- Świeca, M.; Gawlik-Dziki, U.; Kowalczyk, D.; Złotek, U. Impact of germination time and type of illumination on the antioxidant compounds and antioxidant capacity of Lens culinaris sprouts. Sci. Hortic. 2012, 140, 87–95. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Rao, J.; Chen, B. Phenolic compounds in germinated cereal and pulse seeds: Classification, transformation, and metabolic process. Crit. Rev. Food Sci. Nutr. 2020, 60, 740–759. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Yeo, Y.-S.; Park, S.-Y.; Lee, S.-G.; Lee, S.-M.; Cho, H.-S.; Chung, N.-J.; Oh, S.-W. Compositional Analysis of Lentil (Lens culinaris) Cultivars Related to Colors and Their Antioxidative Activity. Plant Breed. Biotechnol. 2017, 5, 192–203. [Google Scholar] [CrossRef]
- Wang, H.; Gui, M.; Tian, X.; Xin, X.; Wang, T.; Li, J. Effects of UV-B on vitamin C, phenolics, flavonoids and their related enzyme activities in mung bean sprouts (Vigna radiata). Int. J. Food Sci. 2017, 52, 827–833. [Google Scholar] [CrossRef]
- Ren, S.C.; Sun, J.T. Changes in phenolic content, phenylalanine ammonia-lyase (PAL) activity, and antioxidant capacity of two buckwheat sprouts in relation to germination. J. Funct. Foods 2014, 7, 298–304. [Google Scholar] [CrossRef]
- Qu, S.; Kwon, S.J.; Duan, S.; Lim, Y.J.; Eom, S.H. Isoflavone changes in immature and mature soybeans by thermal processing. Molecules 2021, 26, 7471. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Hernández-Figueroa, K.; Sánchez, E. Antioxidant Capacity and Phytonutrient Content in the Seed Coat and Cotyledon of Common Beans (Phaseolus vulgaris L.) from Various Regions in Mexico. Antioxidants 2018, 8, 5. [Google Scholar] [CrossRef]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—Chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Eom, S.H.; Park, H.J.; Seo, D.W.; Kim, W.W.; Cho, D.H. Stimulating effects of far-infrared ray radiation on the release of antioxidative phenolics in grape berries. Food Sci. Biotechnol. 2009, 18, 362–366. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FG | LG | LR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Kaempferol | Luteolin | Myricetin | Kaempferol | Luteolin | Myricetin | Kaempferol | Luteolin | Myricetin | ||
| Whole | NGS | 406.96 ± 15.71 b | 107.11 ± 14.71 b | 131.14 ± 17.25 b | 446.73 ± 8.66 ab | N.D. | N.D. | 93.80 ± 3.00 a | N.D. | N.D. |
| GS | 357.50 ± 38.29 b | 66.20 ± 4.18 bc | 78.53 ± 11.15 c | 398.39 ± 47.51 b | N.D. | N.D. | 103.64 ± 13.73 a | N.D. | N.D. | |
| Endosperm | NGS | 354.97 ± 41.27 b | N.D. | N.D. | 367.20 ± 12.93 b | N.D. | N.D. | 93.80 ± 3.00 a | N.D. | N.D. |
| GS | 337.02 ± 8.53 c | 32.18 ± 1.04 cd | 31.67 ± 1.70 d | 423.20 ± 26.63 b | N.D. | N.D. | 103.64 ± 13.73 a | N.D. | N.D. | |
| Seed coat | NGS | 539.98 ± 17.31 a | 441.47 ± 27.86 a | 572.45 ± 6.55 a | 537.11 ± 10.10 a | 22.03 ± 5.14 a | 8.67 ± 0.20 a | N.D. | N.D. | N.D. |
| GS | 146.34 ± 3.04 c | 111.62 ± 7.04 b | 125.74 ± 9.33 b | 104.30 ± 6.69 c | 12.31 ± 1.84 a | 11.84 ± 3.39 a | N.D. | N.D. | N.D. | |
| Temperature | FG | LG | LR | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Kaempferol | Luteolin | Myricetin | Kaempferol | Luteolin | Myricetin | Kaempferol | Luteolin | Myricetin | ||
| 30 °C | NGS | 406.96 ± 15.71 ab | 107.11 ± 14.71 a | 131.14 ± 17.25 a | 446.73 ± 8.66 a | N.D. | N.D. | 93.80 ± 3.00 ab | N.D. | N.D. |
| GS | 357.50 ± 38.29 b | 66.20 ± 4.18 a | 78.53 ± 11.15 a | 398.39 ± 47.51 a | N.D. | N.D. | 103.64 ± 13.73 ab | N.D. | N.D. | |
| 60 °C | NGS | 524.81 ± 40.68 a | 123.32 ± 20.53 a | 128.39 ± 15.26 a | 375.78 ± 39.27 a | N.D. | N.D. | 84.67 ± 1.07 ab | N.D. | N.D. |
| GS | 401.13 ± 29.19 ab | 51.72 ± 8.01 a | 70.77 ± 7.50 a | 365.51 ± 20.28 a | N.D. | N.D. | 71.57 ± 1.90 ab | N.D. | N.D. | |
| 80 °C | NGS | 480.27 ± 17.73 ab | 120.84 ± 23.49 a | 132.16 ± 19.88 a | 482.87 ± 22.69 a | N.D. | N.D. | 153.35 ± 48.84 ab | N.D. | N.D. |
| GS | 453.68 ± 15.07 ab | 94.12 ± 2.01 a | 116.12 ± 9.37 a | 427.26 ± 7.73 a | N.D. | N.D. | 64.41 ± 5.58 b | N.D. | N.D. | |
| 100 °C | NGS | 444.16 ± 26.49 ab | 115.16 ± 30.45 a | 134.04 ± 32.73 a | 461.55 ± 28.74 a | N.D. | N.D. | 153.35 ± 48.84 ab | N.D. | N.D. |
| GS | 481.11 ± 29.50 ab | 66.83 ± 14.95 a | 74.37 ± 7.93 a | 430.85 ± 57.39 a | N.D. | N.D. | 163.81 ± 23.65 a | N.D. | N.D. | |
| TPC | TFC | Kaempferol | Luteolin | Myricetin | DPPH | ABTS | |
|---|---|---|---|---|---|---|---|
| TPC | 1 | ||||||
| TFC | 0.861 *** | 1 | |||||
| Kaempferol | 0.508 *** | 0.514 *** | 1 | ||||
| Luteolin | 0.507 *** | 0.498 *** | 0.348 * | 1 | |||
| Myricetin | 0.490 *** | 0.480 *** | 0.353 * | 0.993 *** | 1 | ||
| DPPH | 0.958 *** | 0.785 *** | 0.405 ** | 0.326 * | 0.306 * | 1 | |
| ABTS | 0.926 *** | 0.778 *** | 0.434 ** | 0.168 ns | 0.142 ns | 0.963 *** | 1 |
| TPC | TFC | Kaempferol | Luteolin | Myricetin | DPPH | ABTS | |
|---|---|---|---|---|---|---|---|
| TPC | 1 | ||||||
| TFC | 0.744 *** | 1 | |||||
| Kaempferol | 0.862 *** | 0.663 *** | 1 | ||||
| Luteolin | 0.587 *** | 0.135 ns | 0.501 *** | 1 | |||
| Myricetin | 0.589 *** | 0.149 ns | 0.502 *** | 0.991 *** | 1 | ||
| DPPH | 0.904 *** | 0.604 *** | 0.666 *** | 0.594 *** | 0.590 *** | 1 | |
| ABTS | 0.833 *** | 0.560 *** | 0.546 *** | 0.507 *** | 0.498 *** | 0.973 *** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Duan, S.; Park, Y.R.; Eom, S.H. Tissue-Specific Antioxidant Activities of Germinated Seeds in Lentil Cultivars during Thermal Processing. Antioxidants 2023, 12, 670. https://doi.org/10.3390/antiox12030670
Kim JH, Duan S, Park YR, Eom SH. Tissue-Specific Antioxidant Activities of Germinated Seeds in Lentil Cultivars during Thermal Processing. Antioxidants. 2023; 12(3):670. https://doi.org/10.3390/antiox12030670
Chicago/Turabian StyleKim, Ji Hye, Shucheng Duan, You Rang Park, and Seok Hyun Eom. 2023. "Tissue-Specific Antioxidant Activities of Germinated Seeds in Lentil Cultivars during Thermal Processing" Antioxidants 12, no. 3: 670. https://doi.org/10.3390/antiox12030670