
Brain Damage in Preterm and Full-Term Neonates: Serum Biomarkers for the Early Diagnosis and Intervention

 ,
,  , , , and
, , , and 
Abstract
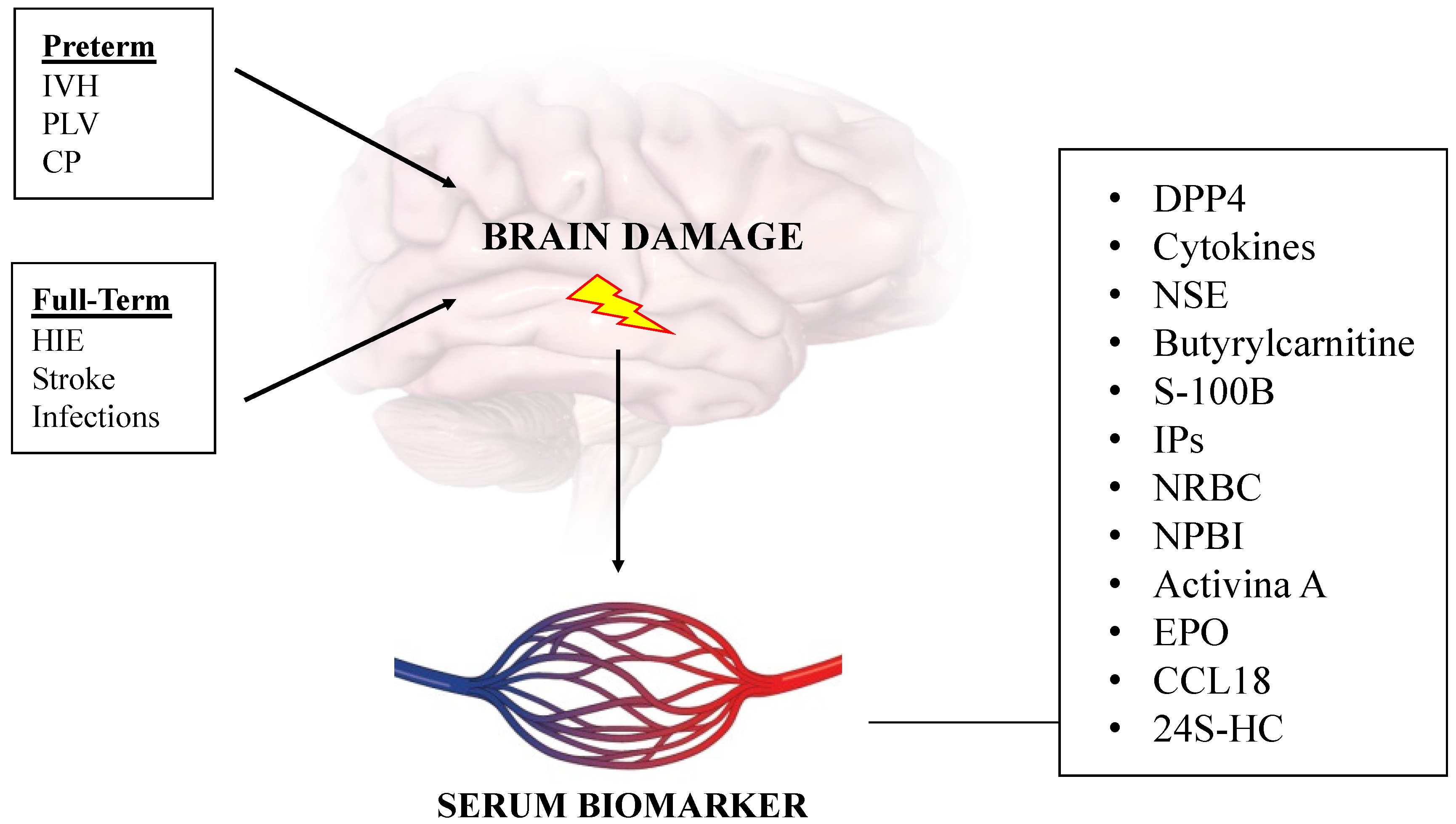
:1. Introduction
2. Brain Development
3. Brain Damage in Preterm Newborn
- Grade 1: hemorrhage apparently confined to germinal matrix, distended less than 10% of ventricular space;
- Grade 2: hemorrhage clearly localized in the ventricle but occupying 10–50% of ventricular space;
- Grade 3: large hemorrhage, occupying greater than 50% of ventricle with or without ventricular echo-densities.
4. Brain Damage in Full-Term Newborn
5. Serum Biomarkers
5.1. Dipeptidylpeptidase 4
5.2. Cytokines
5.3. Neuron-Specific Enolase
5.4. Butyrylcarnitine
5.5. Acidic Calcium-Binding Protein
5.6. F2-Isoprostanes
5.7. Nucleated Red Blood Cells
5.8. Non-Protein-Bound Protein
5.9. Activina A
5.10. Erythropoietin
5.11. Chemokine Ligand 18
5.12. 24S-Hydroxycholesterol
5.13. Neurotrophins
6. Placenta and Perinatal Brain Damage
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| OS | oxidative stress |
| FRs | free radicals |
| cUS | cerebral ultrasound |
| MRI | magnetic resonance imaging |
| IVH | intraventricular hemorrhage |
| PVHI | periventricular hemorrhagic infarction |
| PVL | periventricular leukomalacia |
| CP | cerebral palsy |
| HIE | hypoxic ischemic encephalopathy |
| HI | hypoxic ischemia |
| cCMV | congenital cytomegalovirus |
| DPP4 | dipeptidylpeptidase 4 |
| NO | nitric oxide |
| ROS | reactive oxygen species |
| NSE | neuron-specific enolase |
| SCAD | short-chain acyl-CoA dehydrogenase |
| S-100B | acidic calcium-binding protein |
| SGA | small for gestational age |
| AGA | adequate for gestational age |
| IPs | F2-Isoprostanes |
| WMI | white matter injury |
| OLs | preoligodendrocytes |
| NRBC | nucleated red blood cells |
| NPBI | non-protein-bound protein |
| EPO | erythropoietin |
| CCL18 | chemokine ligand 18 |
| 24S-HC | 24S-hydroxycholesterol |
| NGF | nerve growth factor |
| BDNF | Brain-derived neurotrophic factor |
| bFGF | basic fibroblast growth factor |
| CRH | corticotropin-releasing hormone |
References
- Tataranno, M.L.; Perrone, S.; Buonocore, G. Plasma Biomarkers of Oxidative Stress in Neonatal Brain Injury. Clin. Perinatol. 2015, 42, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Santacroce, A.; Picardi, A.; Buonocore, G. Fetal Programming and Early Identification of Newborns at High Risk of Free Radical-Mediated Diseases. WJCP 2016, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Czerska, M.; Mikołajewska, K.; Zieliński, M.; Gromadzińska, J.; Wąsowicz, W. Today’s Oxidative Stress Markers. Med. Pr. 2015, 66, 393–405. [Google Scholar] [CrossRef]
- Perrone, S.; Tataranno, L.M.; Stazzoni, G.; Ramenghi, L.; Buonocore, G. Brain Susceptibility to Oxidative Stress in the Perinatal Period. J. Matern.-Fetal Neonatal Med. 2015, 28, 2291–2295. [Google Scholar] [CrossRef]
- Lembo, C.; Buonocore, G.; Perrone, S. Oxidative Stress in Preterm Newborns. Antioxidants 2021, 10, 1672. [Google Scholar] [CrossRef] [PubMed]
- Bersani, I.; Pluchinotta, F.; Dotta, A.; Savarese, I.; Campi, F.; Auriti, C.; Chuklantseva, N.; Piersigilli, F.; Gazzolo, F.; Varrica, A.; et al. Early Predictors of Perinatal Brain Damage: The Role of Neurobiomarkers. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 471–486. [Google Scholar] [CrossRef] [Green Version]
- Volpe, J.J.; Volpe, J.J. (Eds.) Volpe’s Neurology of the Newborn, 6th ed.; Elsevier: Philadelphia, PA, USA, 2018; ISBN 978-0-323-42876-7. [Google Scholar]
- Volpe, J.J. Brain Injury in Premature Infants: A Complex Amalgam of Destructive and Developmental Disturbances. Lancet Neurol. 2009, 8, 110–124. [Google Scholar] [CrossRef] [Green Version]
- Girard, N.J.; Dory-Lautrec, P.; Koob, M.; Dediu, A.M. MRI Assessment of Neonatal Brain Maturation. Imaging Med. 2012, 4, 613–632. [Google Scholar] [CrossRef]
- Kostović, I.; Radoš, M.; Kostović-Srzentić, M.; Krsnik, Ž. Fundamentals of the Development of Connectivity in the Human Fetal Brain in Late Gestation: From 24 Weeks Gestational Age to Term. J. Neuropathol. Exp. Neurol. 2021, 80, 393–414. [Google Scholar] [CrossRef]
- Andersen, A.; Fisker, A.B.; Rodrigues, A.; Martins, C.; Ravn, H.; Lund, N.; Biering-Sørensen, S.; Benn, C.S.; Aaby, P. National Immunization Campaigns with Oral Polio Vaccine Reduce All-Cause Mortality: A Natural Experiment within Seven Randomized Trials. Front. Public Health 2018, 6, 13. [Google Scholar] [CrossRef]
- Boscarino, G.; Conti, M.G.; Gasparini, C.; Onestà, E.; Faccioli, F.; Dito, L.; Regoli, D.; Spalice, A.; Parisi, P.; Terrin, G. Neonatal Hyperglycemia Related to Parenteral Nutrition Affects Long-Term Neurodevelopment in Preterm Newborn: A Prospective Cohort Study. Nutrients 2021, 13, 1930. [Google Scholar] [CrossRef]
- Puia-Dumitrescu, M.; Comstock, B.A.; Li, S.; Heagerty, P.J.; Perez, K.M.; Law, J.B.; Wood, T.R.; Gogcu, S.; Mayock, D.E.; Juul, S.E.; et al. Assessment of 2-Year Neurodevelopmental Outcomes in Extremely Preterm Infants Receiving Opioids and Benzodiazepines. JAMA Netw. Open 2021, 4, e2115998. [Google Scholar] [CrossRef] [PubMed]
- Boscarino, G.; Di Chiara, M.; Cellitti, R.; De Nardo, M.C.; Conti, M.G.; Parisi, P.; Spalice, A.; Di Mario, C.; Ronchi, B.; Russo, A.; et al. Effects of Early Energy Intake on Neonatal Cerebral Growth of Preterm Newborn: An Observational Study. Sci. Rep. 2021, 11, 18457. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Kane, A.E.; Brown, S.E.; Tarver, T.; Dusing, S.C. Effect of Neonatal Therapy on the Motor, Cognitive, and Behavioral Development of Infants Born Preterm: A Systematic Review. Dev. Med. Child Neurol. 2020, 62, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Terrin, G.; Boscarino, G.; Gasparini, C.; Di Chiara, M.; Faccioli, F.; Onestà, E.; Parisi, P.; Spalice, A.; De Nardo, M.C.; Dito, L.; et al. Energy-Enhanced Parenteral Nutrition and Neurodevelopment of Preterm Newborns: A Cohort Study. Nutrition 2021, 89, 111219. [Google Scholar] [CrossRef] [PubMed]
- Terrin, G.; De Nardo, M.C.; Boscarino, G.; Di Chiara, M.; Cellitti, R.; Ciccarelli, S.; Gasparini, C.; Parisi, P.; Urna, M.; Ronchi, B.; et al. Early Protein Intake Influences Neonatal Brain Measurements in Preterms: An Observational Study. Front. Neurol. 2020, 11, 885. [Google Scholar] [CrossRef]
- Whitelaw, A. Intraventricular Haemorrhage and Posthaemorrhagic Hydrocephalus: Pathogenesis, Prevention and Future Interventions. Semin. Neonatol. 2001, 6, 135–146. [Google Scholar] [CrossRef]
- Deng, W.; Pleasure, J.; Pleasure, D. Progress in Periventricular Leukomalacia. Arch. Neurol. 2008, 65, 1291–1295. [Google Scholar] [CrossRef] [Green Version]
- Bracci, R.; Perrone, S.; Buonocore, G. The Timing of Neonatal Brain Damage. Neonatology 2006, 90, 145–155. [Google Scholar] [CrossRef]
- Perrone, S.; Lembo, C.; Gironi, F.; Petrolini, C.; Catalucci, T.; Corbo, G.; Buonocore, G.; Gitto, E.; Esposito, S.M.R. Erythropoietin as a Neuroprotective Drug for Newborn Infants: Ten Years after the First Use. Antioxidants 2022, 11, 652. [Google Scholar] [CrossRef]
- Kolnik, S.; Wood, T.R. Role of Vitamin E in Neonatal Neuroprotection: A Comprehensive Narrative Review. Life 2022, 12, 1083. [Google Scholar] [CrossRef] [PubMed]
- Abiramalatha, T.; Ramaswamy, V.V.; Ponnala, A.K.; Kallem, V.R.; Murkunde, Y.V.; Punnoose, A.M.; Vivekanandhan, A.; Pullattayil, A.K.; Amboiram, P. Emerging Neuroprotective Interventions in Periventricular Leukomalacia—A Systematic Review of Preclinical Studies. Expert Opin. Investig. Drugs 2022, 31, 305–330. [Google Scholar] [CrossRef] [PubMed]
- A Report: The Definition and Classification of Cerebral Palsy April 2006. Dev. Med. Child Neurol. 2007, 49, 8–14. [CrossRef]
- The Australian Cerebral Palsy Register. Australian Cerebral Palsy Register Bulletin: Birth Years 1995–2014; The Australian Cerebral Palsy Register: North Adelaide, Australia, 2020. [Google Scholar]
- MacLennan, A.H.; Thompson, S.C.; Gecz, J. Cerebral Palsy: Causes, Pathways, and the Role of Genetic Variants. Am. J. Obstet. Gynecol. 2015, 213, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Novak, I.; Morgan, C.; Adde, L.; Blackman, J.; Boyd, R.N.; Brunstrom-Hernandez, J.; Cioni, G.; Damiano, D.; Darrah, J.; Eliasson, A.-C.; et al. Early, Accurate Diagnosis and Early Intervention in Cerebral Palsy: Advances in Diagnosis and Treatment. JAMA Pediatr. 2017, 171, 897. [Google Scholar] [CrossRef]
- Doyle, L.W.; Crowther, C.A.; Middleton, P.; Marret, S.; Rouse, D. Magnesium Sulphate for Women at Risk of Preterm Birth for Neuroprotection of the Fetus. Cochrane Database Syst. Rev. 2009, 21, CD004661. [Google Scholar] [CrossRef]
- Schmidt, B.; Roberts, R.S.; Davis, P.; Doyle, L.W.; Barrington, K.J.; Ohlsson, A.; Solimano, A.; Tin, W. Long-Term Effects of Caffeine Therapy for Apnea of Prematurity. N. Engl. J. Med. 2007, 357, 1893–1902. [Google Scholar] [CrossRef] [Green Version]
- Spittle, A.J.; Morgan, C.; Olsen, J.E.; Novak, I.; Cheong, J.L.Y. Early Diagnosis and Treatment of Cerebral Palsy in Children with a History of Preterm Birth. Clin. Perinatol. 2018, 45, 409–420. [Google Scholar] [CrossRef]
- Hamilton, B.E.; Hoyert, D.L.; Martin, J.A.; Strobino, D.M.; Guyer, B. Annual Summary of Vital Statistics: 2010–2011. Pediatrics 2013, 131, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Yıldız, E.P.; Ekici, B.; Tatlı, B. Neonatal Hypoxic Ischemic Encephalopathy: An Update on Disease Pathogenesis and Treatment. Expert Rev. Neurother. 2017, 17, 449–459. [Google Scholar] [CrossRef]
- Shankaran, S.; Laptook, A.R.; Ehrenkranz, R.A.; Tyson, J.E.; McDonald, S.A.; Donovan, E.F.; Fanaroff, A.A.; Poole, W.K.; Wright, L.L.; Higgins, R.D.; et al. Whole-Body Hypothermia for Neonates with Hypoxic–Ischemic Encephalopathy. N. Engl. J. Med. 2005, 353, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Thoresen, M.; Tooley, J.; Liu, X.; Jary, S.; Fleming, P.; Luyt, K.; Jain, A.; Cairns, P.; Harding, D.; Sabir, H. Time Is Brain: Starting Therapeutic Hypothermia within Three Hours after Birth Improves Motor Outcome in Asphyxiated Newborns. Neonatology 2013, 104, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Azzopardi, D.V.; Strohm, B.; Edwards, A.D.; Dyet, L.; Halliday, H.L.; Juszczak, E.; Kapellou, O.; Levene, M.; Marlow, N.; Porter, E.; et al. Moderate Hypothermia to Treat Perinatal Asphyxial Encephalopathy. N. Engl. J. Med. 2009, 361, 1349–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferriero, D.M. Neonatal Brain Injury. N. Engl. J. Med. 2004, 351, 1985–1995. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Zitta, K.; Groenendaal, F.; van Bel, F.; Peeters-Scholte, C. Neuroprotective Strategies Following Perinatal Hypoxia-Ischemia: Taking Aim at NOS. Free. Radic. Biol. Med. 2019, 142, 123–131. [Google Scholar] [CrossRef]
- Bennet, L.; Roelfsema, V.; Pathipati, P.; Quaedackers, J.S.; Gunn, A.J. Relationship between Evolving Epileptiform Activity and Delayed Loss of Mitochondrial Activity after Asphyxia Measured by Near-Infrared Spectroscopy in Preterm Fetal Sheep: Post-Hypoxic Mitochondrial Failure in Preterm Fetal Sheep. J. Physiol. 2006, 572, 141–154. [Google Scholar] [CrossRef]
- Cross, J.L.; Meloni, B.P.; Bakker, A.J.; Lee, S.; Knuckey, N.W. Modes of Neuronal Calcium Entry and Homeostasis Following Cerebral Ischemia. Stroke Res. Treat. 2010, 2010, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cerio, F.; Lara-Celador, I.; Alvarez, A.; Hilario, E. Neuroprotective Therapies after Perinatal Hypoxic-Ischemic Brain Injury. Brain Sci. 2013, 3, 191–214. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.B.; Lynch, J.K. Stroke in Newborn Infants. Lancet Neurol. 2004, 3, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Laugesaar, R.; Kolk, A.; Tomberg, T.; Metsvaht, T.; Lintrop, M.; Varendi, H.; Talvik, T. Acutely and Retrospectively Diagnosed Perinatal Stroke: A Population-Based Study. Stroke 2007, 38, 2234–2240. [Google Scholar] [CrossRef]
- Lee, J. Maternal and Infant Characteristics Associated With Perinatal Arterial Stroke in the Infant. JAMA 2005, 293, 723. [Google Scholar] [CrossRef] [PubMed]
- Kirton, A.; deVeber, G. Life After Perinatal Stroke. Stroke 2013, 44, 3265–3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernson-Leung, M.E.; Rivkin, M.J. Stroke in Neonates and Children. Pediatr. Rev. 2016, 37, 463–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, K.; Jin, G.; Navaratna, D.; Lo, E.H. Brain Angiogenesis in Developmental and Pathological Processes: Neurovascular Injury and Angiogenic Recovery after Stroke: Neurovascular Responses in Stroke. FEBS J. 2009, 276, 4644–4652. [Google Scholar] [CrossRef] [Green Version]
- Kirton, A. Advancing Non-Invasive Neuromodulation Clinical Trials in Children: Lessons from Perinatal Stroke. Eur. J. Paediatr. Neurol. 2017, 21, 75–103. [Google Scholar] [CrossRef] [PubMed]
- Wanigasinghe, J.; Reid, S.M.; Mackay, M.T.; Reddihough, D.S.; Harvey, A.S.; Freeman, J.L. Epilepsy in Hemiplegic Cerebral Palsy Due to Perinatal Arterial Ischaemic Stroke: Epilepsy in CP Due to Stroke. Dev. Med. Child Neurol. 2010, 52, 1021–1027. [Google Scholar] [CrossRef]
- Boppana, S.B.; Rivera, L.B.; Fowler, K.B.; Mach, M.; Britt, W.J. Intrauterine Transmission of Cytomegalovirus to Infants of Women with Preconceptional Immunity. N. Engl. J. Med. 2001, 344, 1366–1371. [Google Scholar] [CrossRef] [PubMed]
- Pass, R.F.; Fowler, K.B.; Boppana, S.B.; Britt, W.J.; Stagno, S. Congenital Cytomegalovirus Infection Following First Trimester Maternal Infection: Symptoms at Birth and Outcome. J. Clin. Virol. 2006, 35, 216–220. [Google Scholar] [CrossRef]
- Natale, F.; De Curtis, M.; Bizzarri, B.; Orlando, M.P.; Ralli, M.; Liuzzi, G.; Caravale, B.; Franco, F.; Gaeta, A.; Giancotti, A.; et al. Isolated Auditory Neuropathy at Birth in Congenital Cytomegalovirus Infection. Ital. J. Pediatr. 2020, 46, 3. [Google Scholar] [CrossRef] [Green Version]
- Thigpen, J. Congenital Cytomegalovirus—History, Current Practice, and Future Opportunities. Neonatal Netw. 2020, 39, 293–298. [Google Scholar] [CrossRef]
- Johnson, J.; Anderson, B. Cytomegalovirus: Should We Screen Pregnant Women for Primary Infection? Am. J. Perinatol. 2013, 30, 121–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiopris, G.; Veronese, P.; Cusenza, F.; Procaccianti, M.; Perrone, S.; Daccò, V.; Colombo, C.; Esposito, S. Congenital Cytomegalovirus Infection: Update on Diagnosis and Treatment. Microorganisms 2020, 8, 1516. [Google Scholar] [CrossRef] [PubMed]
- Tomicic, M.T.; Bey, E.; Wutzler, P.; Thust, R.; Kaina, B. Comparative Analysis of DNA Breakage, Chromosomal Aberrations and Apoptosis Induced by the Anti-Herpes Purine Nucleoside Analogues Aciclovir, Ganciclovir and Penciclovir. Mutat. Res. /Fundam. Mol. Mech. Mutagen. 2002, 505, 1–11. [Google Scholar] [CrossRef]
- Yakovleva, A.A.; Zolotov, N.N.; Sokolov, O.Y.; Kost, N.V.; Kolyasnikova, K.N.; Micheeva, I.G. Dipeptidylpeptidase 4 (DPP4, CD26) Activity in the Blood Serum of Term and Preterm Neonates with Cerebral Ischemia. Neuropeptides 2015, 52, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Chaparro-Huerta, V.; Flores-Soto, M.E.; Merin Sigala, M.E.; Barrera de León, J.C.; Lemus-Varela, M.d.L.; Torres-Mendoza, B.M.d.G.; Beas-Zárate, C. Proinflammatory Cytokines, Enolase and S-100 as Early Biochemical Indicators of Hypoxic-Ischemic Encephalopathy Following Perinatal Asphyxia in Newborns. Pediatr. Neonatol. 2017, 58, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, C.; Pellegrini, G.; Panero, A.; De Luca, T.; Assumma, M.; Signore, F.; Pacifico, L. Umbilical Cord Interleukin-6 Levels Are Elevated in Term Neonates with Perinatal Asphyxia: Serum IL-6 after Perinatal Asphyxia. Eur. J. Clin. Investig. 2003, 33, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Fotopoulos, S.; Pavlou, K.; Skouteli, H.; Papassotiriou, I.; Lipsou, N.; Xanthou, M. Early Markers of Brain Damage in Premature Low-Birth-Weight Neonates Who Suffered from Perinatal Asphyxia and/or Infection. Biol. Neonate 2001, 79, 213–218. [Google Scholar] [CrossRef]
- Nagdyman, N.; Kömen, W.; Ko, H.K.; Müller, C.; Obladen, M. Early Biochemical Indicators of Hypoxic-Ischemic Encephalopathy after Birth Asphyxia. Pediatr. Res. 2001, 49, 502–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuseppe, D.; Sergio, C.; Pasqua, B.; Giovanni, L.V.; Salvatore, C.; Frigiola, A.; Petra, H.; Maurizio, A. Perinatal Asphyxia in Preterm Neonates Leads to Serum Changes in Protein S-100 and Neuron Specific Enolase. Curr. Neurovasc. Res. 2009, 6, 110–116. [Google Scholar] [CrossRef]
- López-Suárez, O.; Concheiro-Guisán, A.; Sánchez-Pintos, P.; Cocho, J.A.; Fernández Lorenzo, J.R.; Couce, M.L. Acylcarnitine Profile in Neonatal Hypoxic-Ischemic Encephalopathy: The Value of Butyrylcarnitine as a Prognostic Marker. Medicine 2019, 98, e15221. [Google Scholar] [CrossRef]
- Celtik, C.; Acunaş, B.; Oner, N.; Pala, O. Neuron-Specific Enolase as a Marker of the Severity and Outcome of Hypoxic Ischemic Encephalopathy. Brain Dev. 2004, 26, 398–402. [Google Scholar] [CrossRef]
- Gazzolo, D.; Di Iorio, R.; Marinoni, E.; Masetti, P.; Serra, G.; Giovannini, L.; Michetti, F. S100B Protein Is Increased in Asphyxiated Term Infants Developing Intraventricular Hemorrhage. Crit. Care Med. 2002, 30, 1356–1360. [Google Scholar] [CrossRef]
- Gazzolo, D.; Marinoni, E.; Di Iorio, R.; Lituania, M.; Bruschettini, P.L.; Michetti, F. Circulating S100β Protein Is Increased in Intrauterine Growth-Retarded Fetuses. Pediatr. Res. 2002, 51, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzolo, D.; Vinesi, P.; Bartocci, M.; Geloso, M.C.; Bonacci, W.; Serra, G.; Haglid, K.G.; Michetti, F. Elevated S100 Blood Level as an Early Indicator of Intraventricular Hemorrhage in Preterm Infants. Correlation with Cerebral Doppler Velocimetry. J. Neurol. Sci. 1999, 170, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Murabayashi, M.; Minato, M.; Okuhata, Y.; Makimoto, M.; Hosono, S.; Masaoka, N.; Okada, T.; Yamamoto, T.; Mugishima, H.; Takahashi, S.; et al. Kinetics of Serum S100B in Newborns with Intracranial Lesions. Pediatr. Int. 2008, 50, 17–22. [Google Scholar] [CrossRef]
- Strzalko, B.; Karowicz-Bilinska, A.; Wyka, K.; Krajewski, P.; Kesiak, M.; Kociszewska-Najman, B. Serum S100B Protein Concentrations in SGA/FGR Newborns. Ginekol. Pol. 2022, 93, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Velipaşaoğlu, M.; Yurdakök, M.; Özyüncü, Ö.; Portakal, O.; Deren, Ö. Neural Injury Markers to Predict Neonatal Complications in Intrauterine Growth Restriction. J. Obstet. Gynaecol. 2015, 35, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Thorngren-Jerneck, K.; Alling, C.; Herbst, A.; Amer-Wahlin, I.; Marsal, K. S100 Protein in Serum as a Prognostic Marker for Cerebral Injury in Term Newborn Infants with Hypoxic Ischemic Encephalopathy. Pediatr. Res. 2004, 55, 406–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, M.A.; Aschner, J.L.; Stark, A.R.; Moore, P.E.; Slaughter, J.C.; Steele, S.; Beller, A.; Milne, G.L.; Settles, O.; Chorna, O.; et al. Increasing F2-Isoprostanes in the First Month after Birth Predicts Poor Respiratory and Neurodevelopmental Outcomes in Very Preterm Infants. J. Perinatol. 2016, 36, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Coviello, C.; Perrone, S.; Buonocore, G.; Negro, S.; Longini, M.; Dani, C.; de Vries, L.S.; Groenendaal, F.; Vijlbrief, D.C.; Benders, M.J.N.L.; et al. Isoprostanes as Biomarker for White Matter Injury in Extremely Preterm Infants. Front. Pediatr. 2020, 8, 618622. [Google Scholar] [CrossRef]
- Coviello, C.; Perrone, S.; Buonocore, G.; Negro, S.; Longini, M.; Groenendaal, F.; Vijlbrief, D.C.; Dani, C.; Benders, M.J.N.L.; Tataranno, M.L. Oxidative Stress Biomarkers and Early Brain Activity in Extremely Preterm Infants: A Prospective Cohort Study. Children 2022, 9, 1376. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Hendon, B.; Mimouni, F.B. Nucleated Erythrocytes and Intraventricular Hemorrhage in Preterm Neonates. Pediatrics 1995, 96, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Florio, P.; Perrone, S.; Luisi, S.; Longini, M.; Tanganelli, D.; Petraglia, F.; Buonocore, G. Activin A Plasma Levels at Birth: An Index of Fetal Hypoxia in Preterm Newborn. Pediatr. Res. 2003, 54, 696–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buonocore, G.; Perrone, S.; Longini, M.; Paffetti, P.; Vezzosi, P.; Gatti, M.G.; Bracci, R. Non Protein Bound Iron as Early Predictive Marker of Neonatal Brain Damage. Brain 2003, 126, 1224–1230. [Google Scholar] [CrossRef] [Green Version]
- Florio, P.; Perrone, S.; Luisi, S.; Vezzosi, P.; Longini, M.; Marzocchi, B.; Petraglia, F.; Buonocore, G. Increased Plasma Concentrations of Activin A Predict Intraventricular Hemorrhage in Preterm Newborns. Clin. Chem. 2006, 52, 1516–1521. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, V.; Buhimschi, C.S.; Han, C.S.; Lee, S.Y.; Pettker, C.M.; Campbell, K.H.; Dulay, A.T.; Oliver, E.A.; Werner, E.F.; Buhimschi, I.A. Cord Blood Erythropoietin and Interleukin-6 for Prediction of Intraventricular Hemorrhage in the Preterm Neonate. J. Matern.-Fetal Neonatal Med. 2011, 24, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Kallankari, H.; Kaukola, T.; Ojaniemi, M.; Herva, R.; Perhomaa, M.; Vuolteenaho, R.; Kingsmore, S.F.; Hallman, M. Chemokine CCL18 Predicts Intraventricular Hemorrhage in Very Preterm Infants. Ann. Med. 2010, 42, 416–425. [Google Scholar] [CrossRef]
- Lambeir, A.-M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-Peptidase IV from Bench to Bedside: An Update on Structural Properties, Functions, and Clinical Aspects of the Enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef]
- Ansorge, S.; Reinhold, D. Immune Peptides Related to Dipeptidyl Aminopeptidase IV/CD26. In Handbook of Biologically Active Peptides; Elsevier: Amsterdam, The Netherlands, 2006; pp. 567–572. ISBN 978-0-12-369442-3. [Google Scholar]
- García-Horsman, J.A.; Männistö, P.T.; Venäläinen, J.I. On the Role of Prolyl Oligopeptidase in Health and Disease. Neuropeptides 2007, 41, 1–24. [Google Scholar] [CrossRef]
- Röhnert, P.; Schmidt, W.; Emmerlich, P.; Goihl, A.; Wrenger, S.; Bank, U.; Nordhoff, K.; Täger, M.; Ansorge, S.; Reinhold, D.; et al. Dipeptidyl Peptidase IV, Aminopeptidase N and DPIV/APN-like Proteases in Cerebral Ischemia. J. Neuroinflamm. 2012, 9, 44. [Google Scholar] [CrossRef]
- Lokshina, L.A. Plasma membrane proteinases from lymphoid cells and their biological functions. Bioorg. Khim. 1998, 24, 323–331. [Google Scholar] [PubMed]
- Malva, J.O.; Xapelli, S.; Baptista, S.; Valero, J.; Agasse, F.; Ferreira, R.; Silva, A.P. Multifaces of Neuropeptide Y in the Brain—Neuroprotection, Neurogenesis and Neuroinflammation. Neuropeptides 2012, 46, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Iwai, T.; Sawabe, T.; Tanimitsu, K.; Suzuki, M.; Sasaki-Hamada, S.; Oka, J.-I. Glucagon-like Peptide-1 Protects Synaptic and Learning Functions from Neuroinflammation in Rodents: GLP-1 Prevents Inflammation-Induced Dysfunction. J. Neurosci. Res. 2014, 92, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Escobar, M.; Weiss, M.D. Biomarkers of Hypoxic-Ischemic Encephalopathy in Newborns. Front. Neur. 2012, 3, 144. [Google Scholar] [CrossRef] [Green Version]
- Algra, S.O.; Groeneveld, K.M.; Schadenberg, A.W.; Haas, F.; Evens, F.C.; Meerding, J.; Koenderman, L.; Jansen, N.J.; Prakken, B.J. Cerebral Ischemia Initiates an Immediate Innate Immune Response in Neonates during Cardiac Surgery. J Neuroinflamm. 2013, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Cowell, R.M.; Xu, H.; Galasso, J.M.; Silverstein, F.S. Hypoxic-Ischemic Injury Induces Macrophage Inflammatory Protein-1α Expression in Immature Rat Brain. Stroke 2002, 33, 795–801. [Google Scholar] [CrossRef] [Green Version]
- Kaur, C.; Rathnasamy, G.; Ling, E.-A. Roles of Activated Microglia in Hypoxia Induced Neuroinflammation in the Developing Brain and the Retina. J. Neuroimmune Pharmacol. 2013, 8, 66–78. [Google Scholar] [CrossRef]
- Leviton, A. Preterm Birth and Cerebral Palsy: Is Tumor Necrosis Factor the Missing Link? Dev. Med. Child Neurol. 2008, 35, 553–558. [Google Scholar] [CrossRef]
- Isgrò, M.A.; Bottoni, P.; Scatena, R. Neuron-Specific Enolase as a Biomarker: Biochemical and Clinical Aspects. In Advances in Cancer Biomarkers; Scatena, R., Ed.; Advances in Experimental Medicine and Biology; Springer: Dordrecht, The Netherlands, 2015; Volume 867, pp. 125–143. ISBN 978-94-017-7214-3. [Google Scholar]
- Wolfe, L.; Jethva, R.; Oglesbee, D.; Vockley, J. Short-Chain Acyl-CoA Dehydrogenase Deficiency. In GeneReviews®; Adam, M.P., Everman, D.B., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; University of Washington Seattle: Seattle, WA, USA, 1993. [Google Scholar]
- Fernandez-Lainez, C.; Aguilar-Lemus, J.; Vela-Amieva, M.; Ibarra-Gonzalez, I. Tandem Mass Spectrometry Newborn Screening for Inborn Errors of Intermediary Metabolism: Abnormal Profile Interpretation. CMC 2012, 19, 4511–4522. [Google Scholar] [CrossRef]
- Millington, D.S.; Kodo, N.; Norwood, D.L.; Roe, C.R. Tandem Mass Spectrometry: A New Method for Acylcarnitine Profiling with Potential for Neonatal Screening for Inborn Errors of Metabolism. J Inher. Metab. Dise. 1990, 13, 321–324. [Google Scholar] [CrossRef]
- Beharier, O.; Kahn, J.; Shusterman, E.; Sheiner, E. S100B—A Potential Biomarker for Early Detection of Neonatal Brain Damage Following Asphyxia. J. Matern.-Fetal Neonatal Med. 2012, 25, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Michetti, F.; Corvino, V.; Geloso, M.C.; Lattanzi, W.; Bernardini, C.; Serpero, L.; Gazzolo, D. The S100B Protein in Biological Fluids: More than a Lifelong Biomarker of Brain Distress. J. Neurochem. 2012, 120, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Ikura, M. Calcium Binding and Conformational Response in EF-Hand Proteins. Trends Biochem. Sci. 1996, 21, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s Double Life: Intracellular Regulator and Extracellular Signal. Biochim. Biophys. Acta 2009, 1793, 1008–1022. [Google Scholar] [CrossRef] [Green Version]
- Van Eldik, L.J.; Wainwright, M.S. The Janus Face of Glial-Derived S100B: Beneficial and Detrimental Functions in the Brain. Restor. Neurol. Neurosci. 2003, 21, 97–108. [Google Scholar] [PubMed]
- Gerlach, R.; Demel, G.; König, H.-G.; Gross, U.; Prehn, J.H.M.; Raabe, A.; Seifert, V.; Kögel, D. Active Secretion of S100B from Astrocytes during Metabolic Stress. Neuroscience 2006, 141, 1697–1701. [Google Scholar] [CrossRef]
- Gazzolo, D.; Abella, R.; Marinoni, E.; di Iorio, R.; Li Volti, G.; Galvano, F.; Frigiola, A.; Temporini, F.; Moresco, L.; Colivicchi, M.; et al. New Markers of Neonatal Neurology. J. Matern Fetal Neonatal Med. 2009, 22 (Suppl. 3), 57–61. [Google Scholar] [CrossRef]
- Gazzolo, D.; Michetti, F. Perinatal S100B Protein Assessment in Human Unconventional Biological Fluids: A Minireview and New Perspectives. Cardiovasc. Psychiatry Neurol. 2010, 2010, 703563. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.G.; Koppal, T.; Akama, K.T.; Guo, L.; Craft, J.M.; Samy, B.; Schavocky, J.P.; Watterson, D.M.; Van Eldik, L.J. Mechanism of Glial Activation by S100B: Involvement of the Transcription Factor NFkappaB. Neurobiol. Aging 2001, 22, 765–772. [Google Scholar] [CrossRef]
- Florio, P. Biochemical Markers of Perinatal Brain Damage. Front. Biosci. 2010, S2, 47–72. [Google Scholar] [CrossRef]
- Gazzolo, D.; Abella, R.; Marinoni, E.; Di Iorio, R.; Li Volti, G.; Galvano, F.; Pongiglione, G.; Frigiola, A.; Bertino, E.; Florio, P. Circulating Biochemical Markers of Brain Damage in Infants Complicated by Ischemia Reperfusion Injury. Cardiovasc. Hematol. Agents Med. Chem. 2009, 7, 108–126. [Google Scholar] [CrossRef] [PubMed]
- Gazzolo, D.; Bruschettini, M.; Lituania, M.; Serra, G.; Bonacci, W.; Michetti, F. Increased Urinary S100B Protein as an Early Indicator of Intraventricular Hemorrhage in Preterm Infants: Correlation with the Grade of Hemorrhage. Clin. Chem. 2001, 47, 1836–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzolo, D.; Marinoni, E.; Di Iorio, R.; Bruschettini, M.; Kornacka, M.; Lituania, M.; Majewska, U.; Serra, G.; Michetti, F. Measurement of Urinary S100B Protein Concentrations for the Early Identification of Brain Damage in Asphyxiated Full-Term Infants. Arch. Pediatr. Adolesc. Med. 2003, 157, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Gazzolo, D.; Frigiola, A.; Bashir, M.; Iskander, I.; Mufeed, H.; Aboulgar, H.; Venturini, P.; Marras, M.; Serra, G.; Frulio, R.; et al. Diagnostic Accuracy of S100B Urinary Testing at Birth in Full-Term Asphyxiated Newborns to Predict Neonatal Death. PLoS ONE 2009, 4, e4298. [Google Scholar] [CrossRef]
- Gazzolo, D.; Marinoni, E.; Di Iorio, R.; Bruschettini, M.; Kornacka, M.; Lituania, M.; Majewska, U.; Serra, G.; Michetti, F. Urinary S100B Protein Measurements: A Tool for the Early Identification of Hypoxic-Ischemic Encephalopathy in Asphyxiated Full-Term Infants. Crit. Care Med. 2004, 32, 131–136. [Google Scholar] [CrossRef]
- Tskitishvili, E.; Komoto, Y.; Temma-Asano, K.; Hayashi, S.; Kinugasa, Y.; Tsubouchi, H.; Song, M.; Kanagawa, T.; Shimoya, K.; Murata, Y. S100B Protein Expression in the Amnion and Amniotic Fluid in Pregnancies Complicated by Pre-Eclampsia. Mol. Hum. Reprod. 2006, 12, 755–761. [Google Scholar] [CrossRef] [Green Version]
- Gazzolo, D.; Marinoni, E.; Di Iorio, R.; Lituania, M.; Marras, M.; Bruschettini, M.; Bruschettini, P.; Frulio, R.; Michetti, F.; Petraglia, F.; et al. High Maternal Blood S100B Concentrations in Pregnancies Complicated by Intrauterine Growth Restriction and Intraventricular Hemorrhage. Clin. Chem. 2006, 52, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Milne, G.L.; Musiek, E.S.; Morrow, J.D. F2-Isoprostanes as Markers of Oxidative Stress in Vivo: An Overview. Biomarkers 2005, 10, 10–23. [Google Scholar] [CrossRef]
- Ferriero, D.M.; Miller, S.P. Imaging Selective Vulnerability in the Developing Nervous System: Imaging Selective Vulnerability. J. Anat. 2010, 217, 429–435. [Google Scholar] [CrossRef]
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Levine, J.M.; Volpe, J.J.; Kinney, H.C. Late Oligodendrocyte Progenitors Coincide with the Developmental Window of Vulnerability for Human Perinatal White Matter Injury. J. Neurosci. 2001, 21, 1302–1312. [Google Scholar] [CrossRef]
- Back, S.A.; Luo, N.L.; Mallinson, R.A.; O’Malley, J.P.; Wallen, L.D.; Frei, B.; Morrow, J.D.; Petito, C.K.; Roberts, C.T.; Murdoch, G.H.; et al. Selective Vulnerability of Preterm White Matter to Oxidative Damage Defined by F2-Isoprostanes. Ann. Neurol. 2005, 58, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Paffetti, P.; Perrone, S.; Longini, M.; Ferrari, A.; Tanganelli, D.; Marzocchi, B.; Buonocore, G. Non-Protein-Bound Iron Detection in Small Samples of Biological Fluids and Tissues. BTER 2006, 112, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, G.; Pantopoulos, K. Iron Metabolism and Toxicity. Toxicol. Appl. Pharmacol. 2005, 202, 199–211. [Google Scholar] [CrossRef]
- Buonocore, G.; Perrone, S.; Bracci, R. Free Radicals and Brain Damage in the Newborn. Biol. Neonate 2001, 79, 180–186. [Google Scholar] [CrossRef]
- Florio, P.; Gazzolo, D.; Luisi, S.; Petraglia, F. Activin A In Brain Injury. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2007; Volume 43, pp. 117–130. ISBN 978-0-12-373703-8. [Google Scholar]
- He, J.-T.; Mang, J.; Mei, C.-L.; Yang, L.; Wang, J.-Q.; Xing, Y.; Yang, H.; Xu, Z.-X. Neuroprotective Effects of Exogenous Activin A on Oxygen-Glucose Deprivation in PC12 Cells. Molecules 2011, 17, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Mukerji, S.S.; Katsman, E.A.; Wilber, C.; Haner, N.A.; Selman, W.R.; Hall, A.K. Activin Is a Neuronal Survival Factor That Is Rapidly Increased after Transient Cerebral Ischemia and Hypoxia in Mice. J. Cereb. Blood Flow Metab. 2007, 27, 1161–1172. [Google Scholar] [CrossRef] [Green Version]
- Florio, P.; Luisi, S.; Bruschettini, M.; Grutzfeld, D.; Dobrzanska, A.; Bruschettini, P.; Petraglia, F.; Gazzolo, D. Cerebrospinal Fluid Activin A Measurement in Asphyxiated Full-Term Newborns Predicts Hypoxic Ischemic Encephalopathy. Clin. Chem. 2004, 50, 2386–2389. [Google Scholar] [CrossRef]
- Davis, L.E.; Widness, J.A.; Brace, R.A. Renal and Placental Secretion of Erythropoietin during Anemia or Hypoxia in the Ovine Fetus. Am. J. Obstet. Gynecol. 2003, 189, 1764–1770. [Google Scholar] [CrossRef]
- Teramo, K.A.; Widness, J.A. Increased Fetal Plasma and Amniotic Fluid Erythropoietin Concentrations: Markers of Intrauterine Hypoxia. Neonatology 2009, 95, 105–116. [Google Scholar] [CrossRef]
- Marti, H.H. Erythropoietin and the Hypoxic Brain. J. Exp. Biol. 2004, 207, 3233–3242. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O.; Nomiyama, H. The Chemokine and Chemokine Receptor Superfamilies and Their Molecular Evolution. Genome Biol. 2006, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Schutyser, E. Involvement of CC Chemokine Ligand 18 (CCL18) in Normal and Pathological Processes. J. Leukoc. Biol. 2005, 78, 14–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-Y.; Lee, Y.-H.; Leu, S.-J.; Wang, C.-Y.; Wei, C.-P.; Hung, K.-S.; Pai, M.-H.; Tsai, M.-D.; Wu, C.-H. CC-Chemokine Ligand 18/Pulmonary Activation-Regulated Chemokine Expression in the CNS with Special Reference to Traumatic Brain Injuries and Neoplastic Disorders. Neuroscience 2010, 165, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Björkhem, I.; Lütjohann, D.; Diczfalusy, U.; Ståhle, L.; Ahlborg, G.; Wahren, J. Cholesterol Homeostasis in Human Brain: Turnover of 24S-Hydroxycholesterol and Evidence for a Cerebral Origin of Most of This Oxysterol in the Circulation. J. Lipid Res. 1998, 39, 1594–1600. [Google Scholar] [CrossRef]
- Cartagena, C.M.; Burns, M.P.; Rebeck, G.W. 24S-Hydroxycholesterol Effects on Lipid Metabolism Genes Are Modeled in Traumatic Brain Injury. Brain Res. 2010, 1319, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lu, F.; Zhu, J.; Guo, S.; Wong, B.J.; Chehab, F.F.; Ferriero, D.M.; Jiang, X. Upregulation of Cholesterol 24-Hydroxylase Following Hypoxia-Ischemia in Neonatal Mouse Brain. Pediatr. Res. 2018, 83, 1218–1227. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Li, S.; Lv, S.H.; Piao, H.; Zhang, Y.H.; Zhang, Y.M.; Ma, H.; Zhang, J.; Sun, C.K.; Li, A.P. Hypoxia-Ischemia Brain Damage Disrupts Brain Cholesterol Homeostasis in Neonatal Rats. Neuropediatrics 2009, 40, 179–185. [Google Scholar] [CrossRef]
- Dave, A.M.; Peeples, E.S. Cholesterol Metabolism and Brain Injury in Neonatal Encephalopathy. Pediatr. Res. 2021, 90, 37–44. [Google Scholar] [CrossRef]
- Mast, N.; Anderson, K.W.; Johnson, K.M.; Phan, T.T.N.; Guengerich, F.P.; Pikuleva, I.A. In Vitro Cytochrome P450 46A1 (CYP46A1) Activation by Neuroactive Compounds. J. Biol. Chem. 2017, 292, 12934–12946. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, Y.; Meaney, S.; Heverin, M.; Ekström, L.; Brafman, A.; Shafir, M.; Andersson, U.; Olin, M.; Eggertsen, G.; Diczfalusy, U.; et al. Studies on the Transcriptional Regulation of Cholesterol 24-Hydroxylase (CYP46A1): Marked Insensitivity toward Different Regulatory Axes. J. Biol. Chem. 2006, 281, 3810–3820. [Google Scholar] [CrossRef]
- Sodero, A.O.; Weissmann, C.; Ledesma, M.D.; Dotti, C.G. Cellular Stress from Excitatory Neurotransmission Contributes to Cholesterol Loss in Hippocampal Neurons Aging in Vitro. Neurobiol. Aging 2011, 32, 1043–1053. [Google Scholar] [CrossRef]
- Lu, F.; Fan, S.; Romo, A.R.; Xu, D.; Ferriero, D.M.; Jiang, X. Serum 24S-Hydroxycholesterol Predicts Long-Term Brain Structural and Functional Outcomes after Hypoxia-Ischemia in Neonatal Mice. J. Cereb. Blood Flow Metab. 2021, 41, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. Neurotrophic Factors: An Overview. In Neurotrophic Factors; Skaper, S.D., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; Volume 1727, pp. 1–17. ISBN 978-1-4939-7570-9. [Google Scholar]
- Sun, Z.; Hu, W.; Yin, S.; Lu, X.; Zuo, W.; Ge, S.; Xu, Y. NGF Protects against Oxygen and Glucose Deprivation-Induced Oxidative Stress and Apoptosis by up-Regulation of HO-1 through MEK/ERK Pathway. Neurosci. Lett. 2017, 641, 8–14. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Kirschner, P.B.; Jenkins, B.G.; Schulz, J.B.; Finkelstein, S.P.; Matthews, R.T.; Rosen, B.R.; Flint Beal, M. NGF, BDNF and NT-5, but Not NT-3 Protect against MPP+ Toxicity and Oxidative Stress in Neonatal Animals. Brain Res. 1996, 713, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, M.; Liu, A.; Di, W.; Zhao, F.; Tian, Y.; Jia, J. Changes of Inflammatory Cytokines and Neurotrophins Emphasized Their Roles in Hypoxic–Ischemic Brain Damage. Int. J. Neurosci. 2013, 123, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Aisa, M.C.; Barbati, A.; Cappuccini, B.; De Rosa, F.; Gerli, S.; Clerici, G.; Kaptilnyy, V.A.; Ishenko, A.I.; Di Renzo, G.C. Urinary Nerve Growth Factor in Full-Term, Preterm and Intra Uterine Growth Restriction Neonates: Association with Brain Growth at 30–40 Days of Postnatal Period and with Neuro-Development Outcome at Two Years. A Pilot Study. Neurosci. Lett. 2021, 741, 135459. [Google Scholar] [CrossRef] [PubMed]
- Perrone, S.; Tataranno, M.L.; Negro, S.; Longini, M.; Toti, M.S.; Alagna, M.G.; Proietti, F.; Bazzini, F.; Toti, P.; Buonocore, G. Placental Histological Examination and the Relationship with Oxidative Stress in Preterm Infants. Placenta 2016, 46, 72–78. [Google Scholar] [CrossRef]
- Yanni, D.; Korzeniewski, S.J.; Allred, E.N.; Fichorova, R.N.; O’Shea, T.M.; Kuban, K.; Dammann, O.; Leviton, A. for the ELGAN Study Investigators; Both Antenatal and Postnatal Inflammation Contribute Information about the Risk of Brain Damage in Extremely Preterm Newborns. Pediatr. Res. 2017, 82, 691–696. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, H.; Mallard, C.; Ferriero, D.M.; Vannucci, S.J.; Levison, S.W.; Vexler, Z.S.; Gressens, P. The Role of Inflammation in Perinatal Brain Injury. Nat. Rev. Neurol. 2015, 11, 192–208. [Google Scholar] [CrossRef]
- Dammann, O.; Leviton, A. Intermittent or Sustained Systemic Inflammation and the Preterm Brain. Pediatr. Res. 2014, 75, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Vohr, B.R.; Wright, L.L.; Poole, W.K.; McDonald, S.A.; for the NICHD Neonatal Research Network Follow-up Study. Neurodevelopmental Outcomes of Extremely Low Birth Weight Infants <32 Weeks’ Gestation Between 1993 and 1998. Pediatrics 2005, 116, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Leviton, A.; Allred, E.N.; Kuban, K.C.K.; O’Shea, T.M.; Paneth, N.; Majzoub, J.; ELGAN Study Investigators. Brain Disorders Associated with Corticotropin-Releasing Hormone Expression in the Placenta among Children Born before the 28th Week of Gestation. Acta Paediatr. 2016, 105, e7–e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notarangelo, F.M.; Schwarcz, R. Restraint Stress during Pregnancy Rapidly Raises Kynurenic Acid Levels in Mouse Placenta and Fetal Brain. Dev. Neurosci. 2016, 38, 458–468. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Biomarker | Reference | Population | Sample Source (i.e., Cord or Peripheral Blood) | Brain Damage Patterns |
|---|---|---|---|---|
| DPP4 | Yakovleva et al. [56] | 39 full-term and 40 preterm | Cord and peripheral blood (3–5 days of life and before discharge) | ↑ cerebral ischemia |
| Cytokines | Chaparro-Huerta et al. [57] | 62 full-term newborns (32 control vs. 30 Asphyxiated) | Plasma samples in the first 24 h of life | TNF-α, IL-1β and IL-6 ↑ in HIE |
| Chiesa et al. [58] | Full-term: 50 case and 113 controls | Cord blood | IL-6 ↑ in HIE and worse neurodevelopmental outcome | |
| Fotopoulos et al. [59] | 57 LBW newborns | Peripheral blood | TNF-α, IL-1β and IL-6 ↑ Asphyxiated/infected | |
| NSE | Chaparro-Huerta et al. [57] | 62 full-term newborns (32 control vs. 30 Asphyxiated) | Plasma samples in the first 24 h of life | ↑ in HIE |
| Nagdyman et al. [60] | 29 asphyxiate and 20 control infants | cord blood and plasma samples at 2, 6, 12, and 24 h after birth | No difference | |
| Giuseppe et al. [61] | 30 neonates with perinatal asphyxia and 10 control | Capillary blood | ↑ in Perinatal Asphyxia | |
| López-Suárez et al. [62] | 67 full-term newborns with HIE, 827 full-term control | Serum samples collected in the first 6 h of life and on the 2nd and 3rd day of life | ↑ in HIE | |
| Celtik et al. [63] | 91 full-term newborns | Peripheral blood | ↑ in HIE | |
| Butyrylcarnitine | López-Suárez et al. [62] | 67 full-term newborns with HIE, 827 full-term control | Serum samples collected in the first 6 h of life and on the 2nd and 3rd day of life | ↑ in HIE |
| S-100B | Nagdyman et al. [60] | 29 asphyxiated and 20 control infants | Cord blood and plasma samples at 2, 6, 12, and 24 h after birth | ↑ in HIE |
| Giuseppe et al. [61] | 30 neonates with perinatal asphyxia and 10 control | Capillary blood | ↑ in Perinatal Asphyxia | |
| Gazzolo et al. [64] | 29 full-term newborns with IVH, 20 asphyxiated infants without IVH, and 80 normal newborns. | Plasma sample at 12 h of life | ↑ in IVH | |
| Gazzolo et al. [65] | 10 IUGR with normal and 10 with abnormal umbilical artery doppler findings. 40 uncomplicated pregnancies | Cord blood | ↑ in IUGR, positive correlation with middle cerebral artery pulsatility index and with umbilical artery pulsatility index to middle cerebral artery pulsatility index ratio. | |
| Gazzolo et al. [66] | 11 IVH and 14 controls | Plasma samples in the first 72 h of life | ↑ in IVH, correlated with the grade of hemorrhage. Positive correlation with the middle cerebral artery pulsatility index and S-100B. | |
| Murabayashi et al. [67] | 22 normal and 40 newborns with brain diseases | Serum samples on day 1, 2 and 6 | ↑ in HIE and Asphyxia | |
| Chaparro-Huerta et al. [57] | 62 full-term newborns (32 control vs. 30 Asphyxiated) | Plasma samples in the first 24 h of life | ↑ in HIE | |
| Strzalko et al. [68] | 88 SGA vs. 80 AGA | Cord blood | ↑ in SGA | |
| Velipaşaoğlu et al. [69] | 32 IUGR vs. 29 controls | Cord blood | ↑ in IUGR | |
| Thorngren-Jerneck et al. [70] | 62 full-term infants with birth asphyxia | Serum samples in the first 2–3 days of life | ↑ in relation with the grade of HIE and ↑ in CP | |
| IPs | Matthews et al. [71] | 136 preterm ≤ 28 wks | Plasma samples on days 14 and 28 of life | ↑ in worse developmental outcomes at 12 months |
| Coviello et al. [72] | 44 preterm < 28 wks | Cord and peripheral blood (24–48 h of life) | ↑ in WMI | |
| Coviello et al. [73] | 39 preterm < 28 wks | Cord and peripheral blood (24–48 h of life) | ↑ in poor EEG activity | |
| NRBC | Green et al. [74] | 149 preterm | Peripheral blood first 6 days of life | ↑ IVH |
| Fotopoulos et al. [59] | 57 LBW newborns | Peripheral blood | ↑ Asphyxiated/infected | |
| Florio et al. [75] | 50 preterm | Cord blood | ↑ Hypoxic | |
| NPBI | Buonocore et al. [76] | 384 newborns (225 >36 weeks, 90 from 32 to 36 weeks and 69 <32 wks). | Cord blood | ↑ in worse neurodevelopmental outcome |
| Activina A | Florio et al. [75] | 50 preterm | Cord blood | ↑ Hypoxic |
| Florio et al. [77] | 53 preterm | Arterial blood in the 1st h of life | ↑ IVH | |
| EPO | Bhandari et al. [78] | 116 preterm | Cord blood | ↑ IVH |
| CCL18 | Kallankari et al. [79] | 163 preterm <32 wks. | Cord blood | ↓ IVH |
| 24S-HC | No study in human babies, only animals. | |||
| NGF | Aisa et al. | 43 preterm >32 wks and full-term neonates. | No plasma or serum, only urine. | ↓ in worse developmental outcomes at 24 months |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrone, S.; Grassi, F.; Caporilli, C.; Boscarino, G.; Carbone, G.; Petrolini, C.; Gambini, L.M.; Di Peri, A.; Moretti, S.; Buonocore, G.; et al. Brain Damage in Preterm and Full-Term Neonates: Serum Biomarkers for the Early Diagnosis and Intervention. Antioxidants 2023, 12, 309. https://doi.org/10.3390/antiox12020309
Perrone S, Grassi F, Caporilli C, Boscarino G, Carbone G, Petrolini C, Gambini LM, Di Peri A, Moretti S, Buonocore G, et al. Brain Damage in Preterm and Full-Term Neonates: Serum Biomarkers for the Early Diagnosis and Intervention. Antioxidants. 2023; 12(2):309. https://doi.org/10.3390/antiox12020309
Chicago/Turabian StylePerrone, Serafina, Federica Grassi, Chiara Caporilli, Giovanni Boscarino, Giulia Carbone, Chiara Petrolini, Lucia Maria Gambini, Antonio Di Peri, Sabrina Moretti, Giuseppe Buonocore, and et al. 2023. "Brain Damage in Preterm and Full-Term Neonates: Serum Biomarkers for the Early Diagnosis and Intervention" Antioxidants 12, no. 2: 309. https://doi.org/10.3390/antiox12020309