
A Novel Therapy for Cisplatin-Induced Allodynia and Dysfunctional and Emotional Impairments in Male and Female Mice



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Drug Administration
2.4. Allodynia
2.5. Forelimb Grip Strength Test
2.6. Anxiety and Depressive Conducts
2.7. Western Blot
2.8. Statistical Analyses
3. Results
3.1. The Prophylactic Treatment with HRW Inhibited the Mechanical and Cold Allodynia Generated by CIS in Male and Female Mice
3.2. The Effects of the Prophylactic Treatment with HRW on the Grip Strength Deficits and Body Weight Loss Produced by CIS in Male and Female Mice
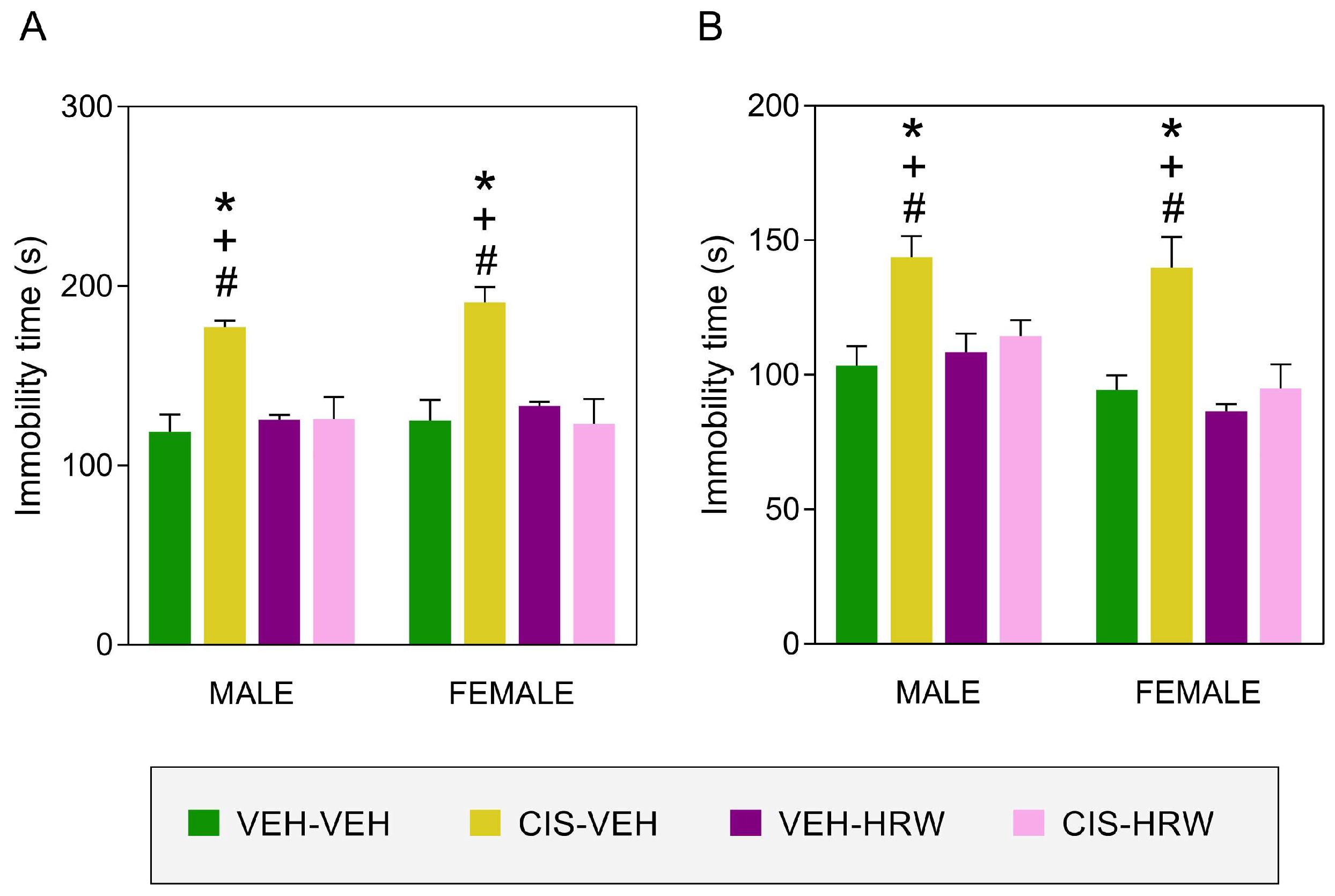
3.3. Treatment with HRW Inhibited the Emotional Disorders Causes by CIS
3.4. Effects of HRW on the Inflammatory and Oxidative Reactions Provoked by CIS in the DRG and PFC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hung, H.W.; Liu, C.Y.; Chen, H.F.; Chang, C.C.; Chen, S.C. Impact of Chemotherapy-Induced Peripheral Neuropathy on Quality of Life in Patients with Advanced Lung Cancer Receiving Platinum-Based Chemotherapy. Int. J. Environ. Res. Public Health 2021, 18, 5677. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, X.; Zhong, T.; Zhou, M.; Gao, L.; Chen, L. Prevalence and associated factors of chemotherapy-related cognitive impairment in older breast cancer survivors. J. Adv. Nurs. 2023, 10, 1111. [Google Scholar] [CrossRef] [PubMed]
- Miltenburg, N.C.; Booger, W. Chemotherapy-induced neuropathy: A comprehensive survey. Cancer Treat. Rev. 2014, 40, 872–882. [Google Scholar] [CrossRef]
- Zhang, J.; Junigan, J.M.; Trinh, R.; Kavelaars, A.; Heijnen, C.J.; Grace, P.M. HDAC6 Inhibition Reverses Cisplatin-Induced Mechanical Hypersensitivity via Tonic Delta Opioid Receptor Signaling. J. Neurosci. 2022, 42, 7862–7874. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, B.; VanCamp, L.; Trosko, J.E.; Mansour, V.H. Platinum compounds: A new class of potent antitumour agents. Nature 1969, 222, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.; Dizon, D.S. New-generation platinum agents for solid tumors. Future Oncol. 2009, 5, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Florea, A.M.; Büsselberg, D. Cisplatin as an anti-tumor drug: Cellular mechanisms of activity, drug resistance and induced side effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Bonhof, C.S.; van de Poll-Franse, L.V.; Vissers, P.A.J.; Wasowicz, D.K.; Wegdam, J.A.; Révész, D.; Vreugdenhil, G.; Mols, F. Anxiety and depression mediate the association between chemotherapy-induced peripheral neuropathy and fatigue: Results from the population-based PROFILES registry. Psychooncology 2019, 28, 1926–1933. [Google Scholar] [CrossRef]
- Albany, C.; Dockter, T.; Wolfe, E.; Le-Rademacher, J.; Wagner-Johnston, N.; Einhorn, L.; Lafky, J.M.; Smith, E.; Pachman, D.; Staff, N.; et al. Cisplatin-associated neuropathy characteristics compared with those associated with other neurotoxic chemotherapy agents (Alliance A151724). Support. Care Cancer 2021, 29, 833–840. [Google Scholar] [CrossRef]
- Conte, E.; Bresciani, E.; Rizzi, L.; Cappellari, O.; De Luca, A.; Torsello, A.; Liantonio, A. Cisplatin-Induced Skeletal Muscle Dysfunction: Mechanisms and Counteracting Therapeutic Strategies. Int. J. Mol. Sci. 2020, 21, 1242. [Google Scholar] [CrossRef]
- Mogil, J.S.; Bailey, A.L. Sex and gender differences in pain and analgesia. Prog. Brain Res. 2010, 186, 141–157. [Google Scholar] [PubMed]
- Rosen, S.; Ham, B.; Mogil, J.S. Sex differences in neuroimmunity and pain. J. Neurosci. Res. 2017, 95, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Lacroix-Fralish, M.L.; Tawfik, V.L.; Nutile-McMenemy, N.; DeLeo, J.A. Progesterone mediates gonadal hormone differences in tactile and thermal hypersensitivity following L5 nerve root ligation in female rats. Neuroscience 2006, 138, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Naji-Esfahani, H.; Vaseghi, G.; Safaeian, L.; Pilehvarian, A.A.; Abed, A.; Rafieian-Kopaei, M. Gender differences in a mouse model of chemotherapy-induced neuropathic pain. Lab Anim. 2016, 50, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Woller, S.A.; Corr, M.; Yaksh, T.L. Differences in cisplatin-induced mechanical allodynia in male and female mice. Eur. J. Pain 2015, 19, 1476–1485. [Google Scholar] [CrossRef]
- Basu, S.; Sodhi, A. Increased release of interleukin-1 and tumour necrosis factor by interleukin-2-induced lymphokine-activated killer cells in the presence of cisplatin and FK-565. Immunol. Cell Biol. 1992, 70, 15–24. [Google Scholar] [CrossRef]
- Wang, X.M.; Lehky, T.J.; Brell, J.M.; Dorsey, S.G. Discovering cytokines as targets for chemotherapy-induced painful peripheral neuropathy. Cytokine 2012, 59, 3–9. [Google Scholar] [CrossRef]
- Thoma, A.; Lightfoot, A.P. NF-kB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 267–279. [Google Scholar]
- Furtado, M.; Katzman, M.A. Examining the role of neuroinflammation in major depression. Psychiatry Res. 2015, 229, 27–36. [Google Scholar] [CrossRef]
- Chen, C.; Smith, M.T. The NLRP3 inflammasome: Role in the pathobiology of chronic pain. Inflammopharmacology 2023, 31, 1589–1603. [Google Scholar] [CrossRef]
- Jin, C.; Flavell, R.A. Molecular mechanism of NLRP3 inflammasome activation. J. Clin. Immunol. 2010, 30, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Monteleone, M.; Adolphe, C.; Batoon, L.; Sandrock, C.J.; Tay, B.; Deuis, J.R.; Smith, A.V.; Mueller, A.; Nadar, E.I.; et al. Vincristine-induced peripheral neuropathy is driven by canonical NLRP3 activation and IL-1β release. J. Exp. Med. 2021, 218, e20201452. [Google Scholar] [CrossRef]
- Lin, T.; Hu, L.; Hu, F.; Li, K.; Wang, C.Y.; Zong, L.J.; Zhao, Y.Q.; Zhang, X.; Li, Y.; Yang, Y.; et al. NET-Triggered NLRP3 Activation and IL18 Release Drive Oxaliplatin-Induced Peripheral Neuropathy. Cancer Immunol. Res. 2022, 10, 1542–1558. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Wu, C.; Gao, F.; Xiang, H.; Sun, N.; Peng, P.; Li, J.; Yuan, X.; Li, H.; Meng, X.; et al. Activation of NLRP3 inflammasome in peripheral nerve contributes to paclitaxel-induced neuropathic pain. Mol. Pain 2017, 13, 1744806917719804. [Google Scholar] [CrossRef] [PubMed]
- Spera, M.C.; Cesta, M.C.; Zippoli, M.; Varrassi, G.; Allegretti, M. Emerging Approaches for the Management of Chemotherapy-Induced Peripheral Neuropathy (CIPN): Therapeutic Potential of the C5a/C5aR Axis. Pain Ther. 2022, 11, 1113–1136. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Makkar, T.K.; Goel, L.; Pahuja, M. Role of inflammation and oxidative stress in chemotherapy-induced neurotoxicity. Immunol. Res. 2022, 70, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Khasabova, I.A.; Khasabov, S.G.; Olson, J.K.; Uhelski, M.L.; Kim, A.H.; Albi-no-Ramírez, A.M.; Wagner, C.L.; Seybold, V.S.; Simone, D.A. Pioglitazone, a PPARγ agonist, reduces cisplatin-evoked neuropathic pain by protecting against oxidative stress. Pain 2019, 160, 688. [Google Scholar] [CrossRef]
- Vukovic, R.; Selakovic, D.; Stankovic, J.S.K.; Kumburovic, I.; Jovicic, N.; Rosic, G. Alteration of Oxidative stress and apoptotic markers alterations in the rat prefrontal cortex influence behavioral response induced by cisplatin and N-acetylcysteine in the tail suspension test. J. Integr. Neurosci. 2021, 20, 711–718. [Google Scholar]
- Moreira-Pais, A.; Ferreira, R.; Gil da Costa, R. Platinum-induced muscle wasting in cancer chemotherapy: Mechanisms and potential targets for therapeutic intervention. Life Sci. 2018, 208, 1–9. [Google Scholar] [CrossRef]
- Shim, H.S.; Bae, C.; Wang, J.; Lee, K.H.; Hankerd, K.M.; Kim, H.K.; Chung, J.M.; La, J.H. Peripheral and central oxidative stress in chemotherapy-induced neuropathic pain. Mol. Pain 2019, 15, 1744806919840098. [Google Scholar] [CrossRef]
- Chen, J.A.; Splenser, A.; Guillory, B.; Luo, J.; Mendiratta, M.; Belinova, B.; Halder, T.; Zhang, G.; Li, Y.P.; Garcia, J.M. Ghrelin prevents tumour- and cisplatin-induced muscle wasting: Characterization of multiple mechanisms involved. J. Cachexia Sarcopenia Muscle 2015, 6, 132–143. [Google Scholar] [CrossRef]
- Saadati, H.; Noroozzadeh, S.; Esmaeili, H.; Amirshahrokhi, K.; Shadman, J.; Niapour, A. The Neuroprotective Effect of Mesna on Cisplatin-Induced Neurotoxicity: Behavioral, Electrophysiological, and Molecular Studies. Neurotox Res. 2021, 39, 826–840. [Google Scholar] [CrossRef]
- Hong, M.; Han, I.H.; Choi, I.; Cha, N.; Kim, W.; Kim, S.K.; Bae, H. Magnoliae Cortex Alleviates Muscle Wasting by Modulating M2 Macrophages in a Cisplatin-Induced Sarcopenia Mouse Model. Int. J. Mol. Sci. 2021, 22, 3188. [Google Scholar] [CrossRef]
- Lian, N.; Shen, M.; Zhang, K.; Pan, J.; Jiang, Y.; Yu, Y.; Yu, Y. Drinking Hydrogen-Rich Water Alleviates Chemotherapy-Induced Neuropathic Pain Through the Regulation of Gut Microbiota. J. Pain Res. 2021, 14, 681–691. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, H.T.; Qin, S.C. Neuroprotective Effects of Molecular Hydrogen: A Critical Review. Neurosci. Bull. 2021, 37, 389–404. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Martínez-Martel, I.; Bai, X.; Batallé, G.; Pol, O. New Treatment for the Cognitive and Emotional Deficits Linked with Paclitaxel-Induced Peripheral Neuropathy in Mice. Antioxidants 2022, 11, 2387. [Google Scholar] [CrossRef]
- Martínez-Serrat, M.; Martínez-Martel, I.; Coral-Pérez, S.; Bai, X.; Batallé, G.; Pol, O. Hydrogen-Rich Water as a Novel Therapeutic Strategy for the Affective Disorders Linked with Chronic Neuropathic Pain in Mice. Antioxidants 2022, 11, 1826. [Google Scholar] [CrossRef]
- Coral-Pérez, S.; Martínez-Martel, I.; Martínez-Serrat, M.; Batallé, G.; Bai, X.; Leite-Panissi, C.R.A.; Pol, O. Treatment with Hydrogen-Rich Water Improves the Nociceptive and Anxio-Depressive-like Behaviors Associated with Chronic Inflammatory Pain in Mice. Antioxidants 2022, 11, 2153. [Google Scholar] [CrossRef]
- Satoh, Y.; Araki, Y.; Kashitani, M.; Nishii, K.; Kobayashi, Y.; Fujita, M.; Suzuki, S.; Morimoto, Y.; Tokuno, S.; Tsumatori, G.; et al. Molecular Hydrogen Prevents Social Deficits and Depression-Like Behaviors Induced by Low-Intensity Blast in Mice. J. Neuropathol. Exp. Neurol. 2018, 77, 827–836. [Google Scholar] [CrossRef]
- Gao, Q.; Song, H.; Wang, X.T.; Liang, Y.; Xi, Y.J.; Gao, Y.; Guo, Q.J.; LeBaron, T.; Luo, Y.X.; Li, S.C.; et al. Molecular hydrogen increases resilience to stress in mice. Sci. Rep. 2017, 7, 9625. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Sasaki, A.T.; Ebisu, K.; Tajima, K.; Kajimoto, O.; Nojima, J.; Kuratsune, H.; Hori, H.; Watanabe, Y. Hydrogen-rich water for improvements of mood, anxiety, and autonomic nerve function in daily life. Med. Gas Res. 2018, 7, 247–255. [Google Scholar] [PubMed]
- Bushnell, M.C.; Ceko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.G. Normalization of Neuroinflammation: A New Strategy for Treatment of Persistent Pain and Memory/Emotional Deficits in Chronic Pain. J. Inflamm. Res. 2022, 15, 5201–5233. [Google Scholar] [CrossRef]
- Krukowski, K.; Ma, J.; Golonzhka, O.; Laumet, G.O.; Gutti, T.; van Duzer, J.H.; Mazitschek, R.; Jarpe, M.B.; Heijnen, C.J.; Kavelaars, A. HDAC6 inhibition effectively reverses chemotherapy-induced peripheral neuropathy. Pain 2017, 158, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, A.; Yoo, K.H.; Rashid, M.A.; Corujo-Ramirez, A.; Hur, B.; Sung, J.; Liu, Y.; Hawse, J.R.; Choi, D.S.; Boison, D.; et al. Adenosine A2A receptor blockade prevents cisplatin-induced impairments in neurogenesis and cognitive function. Proc. Natl. Acad. Sci. USA 2022, 119, e2206415119. [Google Scholar] [CrossRef] [PubMed]
- Boehmerle, W.; Huehnchen, P.; Peruzzaro, S.; Balkaya, M.; Endres, M. Electrophysiological, behavioral and histological characterization of paclitaxel, cisplatin, vincristine and bortezomib-induced neuropathy in C57Bl/6 mice. Sci. Rep. 2014, 4, 6370. [Google Scholar] [CrossRef]
- Cheng, S.; Peng, L.; Xu, B.; Chen, W.; Chen, Y.; Gu, Y. Protective Effects of Hydrogen-Rich Water Against Cartilage Damage in a Rat Model of Osteoarthritis by Inhibiting Oxidative Stress, Matrix Catabolism, and Apoptosis. Med. Sci. Monit. 2020, 26, e92021. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Batallé, G.; Bai, X.; Pol, O. The Interaction between Carbon Monoxide and Hydrogen Sulfide during Chronic Joint Pain in Young Female Mice. Antioxidants 2022, 11, 1271. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef]
- Mao-Ying, Q.L.; Kavelaars, A.; Krukowski, K.; Huo, X.J.; Zhou, W.; Price, T.J.; Cleeland, C.; Heijnen, C.J. The anti-diabetic drug metformin protects against chemotherapy-induced peripheral neuropathy in a mouse model. PLoS ONE 2014, 9, e100701. [Google Scholar] [CrossRef]
- Becker, G.; Fialho, M.F.P.; Brusco, I.; Oliveira, S.M. Kinin B1 and B2 Receptors Contribute to Cisplatin-Induced Painful Peripheral Neuropathy in Male Mice. Pharmaceutics 2023, 15, 852. [Google Scholar] [CrossRef]
- Maj, M.A.; Ma, J.; Krukowski, K.N.; Kavelaars, A.; Heijnen, C.J. Inhibition of Mitochondrial p53 Accumulation by PFT-μ Prevents Cisplatin-Induced Peripheral Neuropathy. Front. Mol. Neurosci. 2017, 10, 108. [Google Scholar] [CrossRef]
- Singh, A.K.; Mahalingam, R.; Squillace, S.; Jacobson, K.A.; Tosh, D.K.; Dharmaraj, S.; Farr, S.A.; Kavelaars, A.; Salvemini, D.; Heijnen, C.J. Targeting the A3 adenosine receptor to prevent and reverse chemotherapy-induced neurotoxicities in mice. Acta Neuropathol. Commun. 2022, 10, 11. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, P.; Zhou, S.; Yan, X.; Zhang, J.; Sun, X.; Yuan, H.; Yu, W. Hydrogen-rich saline attenuated neuropathic pain by reducing oxidative stress. Can. J. Neurol. Sci. 2013, 40, 857–863. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Satoh, Y.; Otsubo, Y.; Kazama, T. Molecular hydrogen attenuates neuropathic pain in mice. PLoS ONE 2014, 9, e100352. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, H.; Xie, K.; Liu, L.; Li, Y.; Yu, Y.; Wang, G. H2 Treatment Attenuated Pain Behavior and Cytokine Release Through the HO-1/CO Pathway in a Rat Model of Neuropathic Pain. Inflammation 2015, 38, 1835–1846. [Google Scholar] [CrossRef]
- Wang, H.; Huo, X.; Chen, H.; Li, B.; Liu, J.; Ma, W.; Wang, X.; Xie, K.; Yu, Y.; Shi, K. Hydrogen-Rich Saline Activated Autophagy via HIF-1 α Pathways in Neuropathic Pain Model. Biomed. Res. Int. 2018, 2018, 4670834. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzadeh, M.; Panahpour, H.; Ghaheri, S.; Saadati, H. Calcitriol supplementation attenuates cisplatin-induced behavioral and cognitive impairments through up-regulation of BDNF in male rats. Brain Res. Bull. 2022, 181, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Chiang, Y.F.; Huang, T.C.; Chen, H.Y.; Lin, P.H.; Ali, M.; Hsia, S.M. Capsaicin alleviates cisplatin-induced muscle loss and atrophy in vitro and in vivo. J. Cachexia Sarcopenia Muscle 2023, 14, 182–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Yang, T.T.; Cheng, T.S.; Hsiao, T.F.; Chang, P.M.; Leu, J.Y.; Wang, F.S.; Hsu, S.L.; Huang, C.F.; Lai, J.M. Modified Sijunzi decoction can alleviate cisplatin-induced toxicity and prolong the survival time of cachectic mice by recovering muscle atrophy. J. Ethnopharmacol. 2019, 233, 47–55. [Google Scholar] [CrossRef]
- Wen, D.; Zhao, P.; Hui, R.; Wang, J.; Shen, Q.; Gong, M.; Guo, H.; Cong, B.; Ma, C. Hydrogen-rich saline attenuates anxiety-like behaviors in morphine-withdrawn mice. Neuropharmacology 2017, 118, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.P.; Panikkar, R. Sarcopenia associated with chemotherapy and targeted agents for cancer therapy. Ann. Palliat. Med. 2019, 8, 86–101. [Google Scholar] [CrossRef]
- Akbar, S.; Subhan, F.; Shahid, M.; Wadood, A.; Shahbaz, N.; Farooq, U.; Ayaz, M.; Raziq, N. Methoxyflavanone abates cisplatin-induced neuropathic pain apropos anti-inflammatory mechanisms: A behavioral and molecular simulation study. Eur. J. Pharmacol. 2020, 872, 172972. [Google Scholar] [CrossRef]
- Suárez-Rojas, I.; Pérez-Fernández, M.; Bai, X.; Martínez-Martel, I.; Intagliata, S.; Pittalà, V.; Salerno, L.; Pol, O. The Inhibition of Neuropathic Pain Incited by Nerve Injury and Accompanying Mood Disorders by New Heme Oxygenase-1 Inducers: Mechanisms Implicated. Antioxidants 2023, 12, 1859. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, X.; Ding, Y.; Zhou, W.; Tao, L.; Lu, P.; Wang, Y.; Hu, R. Nuclear Factor E2-Related Factor-2 Negatively Regulates NLRP3 Inflammasome Activity by Inhibiting Reactive Oxygen Species-Induced NLRP3 Priming. Antioxid. Redox Signal. 2017, 26, 28–43. [Google Scholar] [CrossRef]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef]
- Abdelkader, N.F.; Saad, M.A.; Abdelsalam, R.M. Neuroprotective effect of nebivolol against cisplatin-associated depressive-like behavior in rats. J. Neurochem. 2017, 141, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, W.J.; Chen, Y.; Wu, T.Y.; Gong, H.; Shen, X.L.; Wang, Y.X.; Sun, X.J.; Jiang, C.L. Effects of hydrogen-rich water on depressive-like behavior in mice. Sci. Rep. 2016, 6, 23742. [Google Scholar] [CrossRef] [PubMed]
- Aydin, D.; Peker, E.G.; Karakurt, M.D.; Gurel, A.; Ayyildiz, M.; Cevher, Ş.C.; Agar, E.; Dane, S. Effects of Ginkgo biloba extract on brain oxidative condition after cisplatin exposure. Clin. Investig. Med. 2016, 39, 27511. [Google Scholar] [CrossRef]
- Kumburovic, I.; Selakovic, D.; Juric, T.; Jovicic, N.; Mihailovic, V.; Stankovic, J.K.; Sreckovic, N.; Kumburovic, D.; Jakovljevic, V.; Rosic, G. Antioxidant Effects of Satureja hortensis L. Attenuate the Anxiogenic Effect of Cisplatin in Rats. Oxid. Med. Cell. Longev. 2019, 2019, 8307196. [Google Scholar] [CrossRef] [PubMed]
- Vukovic, R.; Kumburovic, I.; Joksimovic Jovic, J.; Jovicic, N.; Katanic Stankovic, J.S.; Mihailovic, V.; Djuric, M.; Velickovic, S.; Arnaut, A.; Selakovic, D.; et al. N-Acetylcysteine Protects against the Anxiogenic Response to Cisplatin in Rats. Biomolecules 2019, 9, 892. [Google Scholar] [CrossRef]
- Hou, C.; Peng, Y.; Qin, C.; Fan, F.; Liu, J.; Long, J. Hydrogen-rich water improves cognitive impairment gender-dependently in APP/PS1 mice without affecting Aβ clearance. Free Radic Res. 2018, 52, 1311–1322. [Google Scholar] [CrossRef] [PubMed]
- Shao, A.; Wu, H.; Hong, Y.; Tu, S.; Sun, X.; Wu, Q.; Zhao, Q.; Zhang, J.; Sheng, J. Hydrogen-Rich Saline Attenuated Subarachnoid Hemorrhage-Induced Early Brain Injury in Rats by Suppressing Inflammatory Response: Possible Involvement of NF-κB Pathway and NLRP3 Inflammasome. Mol. Neurobiol. 2016, 53, 3462–3476. [Google Scholar] [CrossRef]
- Jangra, A.; Kwatra, M.; Singh, T.; Pant, R.; Kushwah, P.; Ahmed, S.; Dwivedi, D.; Saroha, B.; Lahkar, M. Edaravone alleviates cisplatin-induced neurobehavioral deficits via modulation of oxidative stress and inflammatory mediators in the rat hippocampus. Eur. J. Pharmacol. 2016, 791, 51–61. [Google Scholar] [CrossRef]
- Lin, C.P.; Chuang, W.C.; Lu, F.J.; Chen, C.Y. Anti-oxidant and anti-inflammatory effects of hydrogen-rich water alleviate ethanol-induced fatty liver in mice. World J. Gastroenterol. 2017, 23, 4920–4934. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution | Purchased |
|---|---|---|
| NLRP3 | 1:200 | Adipogen Life Sciences, Epalinges, Switzerland |
| 4-HNE | 1:200 | Abcam, Cambridge, UK |
| HO-1 | 1:100 | Enzo Life Science, New York, NY, USA |
| SOD-1 | 1:150 | Novus Biologic, Littleton, CO, USA |
| GAPDH | 1:5000 | Merck, Billerica, MA, USA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Martel, I.; Pol, O. A Novel Therapy for Cisplatin-Induced Allodynia and Dysfunctional and Emotional Impairments in Male and Female Mice. Antioxidants 2023, 12, 2063. https://doi.org/10.3390/antiox12122063
Martínez-Martel I, Pol O. A Novel Therapy for Cisplatin-Induced Allodynia and Dysfunctional and Emotional Impairments in Male and Female Mice. Antioxidants. 2023; 12(12):2063. https://doi.org/10.3390/antiox12122063
Chicago/Turabian StyleMartínez-Martel, Ignacio, and Olga Pol. 2023. "A Novel Therapy for Cisplatin-Induced Allodynia and Dysfunctional and Emotional Impairments in Male and Female Mice" Antioxidants 12, no. 12: 2063. https://doi.org/10.3390/antiox12122063