

Endothelial Notch Signaling Regulates the Function of the Retinal Pigment Epithelial Barrier via EC Angiocrine Signaling
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Laser-Induced CNV (Choroidal Neovascularization) Model in Mice
2.3. Sodium Iodate Model in Mice
2.4. Cell Culture and Treatment
2.5. RPE and HUVEC Cells’ Coculture
2.6. RPE Polarization and TER Measurements
2.7. Immunofluorescence Assays
2.8. HBEGF Secretion Assays
2.9. MMP-9 Enzyme Activity Assay
2.10. RT- qPCR Assays
2.11. Western Blot Assays
2.12. Statistics
3. Results
3.1. Inhibiting Notch Signaling in Choroid Endothelial Cells (CECs) Affects the RPE Barrier Formation and Aggravates the RPE Barrier Disruption Induced by Sodium Iodate
3.2. Endothelial Notch Signaling Promotes RPE Barrier Formation through Angiocrine Function In Vitro
3.3. Endothelial Notch Signaling Promotes ECM Deposition in the Formation of the RPE Barrier
3.4. Endothelial Notch Signaling Activation Alleviates RPE Barrier Dysfunction in Hypoxia through Angiocrine Signaling
3.5. Endothelial Notch Signaling Affects RPE Barrier Function by Regulating the HBEGF Paracrine of ECs
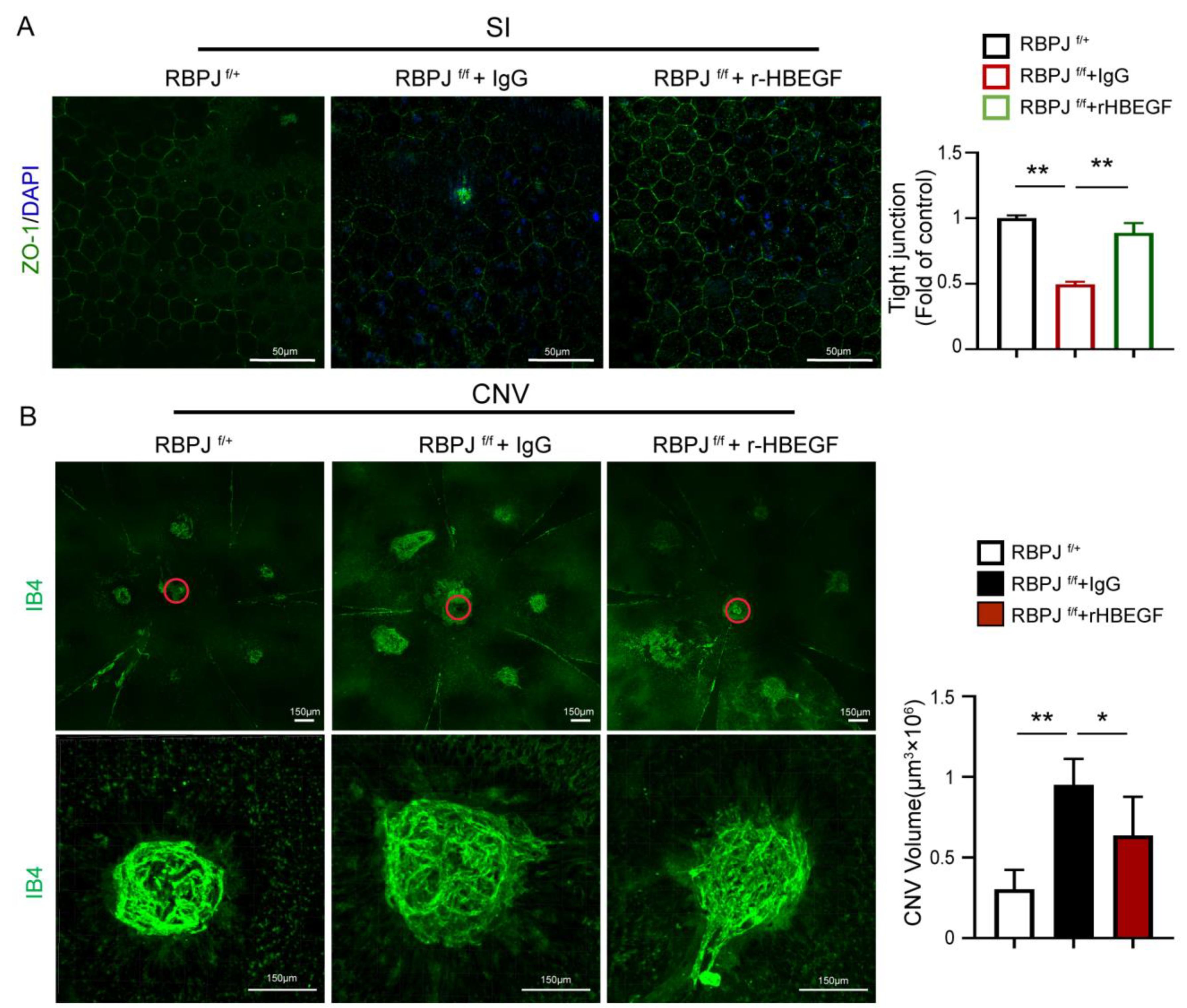
3.6. r-HBEGF Ameliorated the RPE Barrier Dysfunction of EC-Specific Notch Signaling Deletion Mice
3.7. Notch Signaling-Driven Endothelial HBEGF Improved RPE Barrier Function by Regulating MMP-9 Expression in RPE Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tisi, A.; Feligioni, M.; Passacantando, M.; Ciancaglini, M.; Maccarone, R. The Impact of Oxidative Stress on Blood-Retinal Barrier Physiology in Age-Related Macular Degeneration. Cells 2021, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, F.; Campbell, M. The blood-retina barrier in health and disease. FEBS J. 2023, 290, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [PubMed]
- Hudson, N.; Celkova, L.; Hopkins, A.; Greene, C.; Storti, F.; Ozaki, E.; Fahey, E.; Theodoropoulou, S.; Kenna, P.F.; Humphries, M.M.; et al. Dysregulated claudin-5 cycling in the inner retina causes retinal pigment epithelial cell atrophy. J. Clin. Investig. 2019, 4, e130273. [Google Scholar] [CrossRef]
- Liu, W.; Liu, S.; Li, P.; Yao, K. Retinitis Pigmentosa: Progress in Molecular Pathology and Biotherapeutical Strategies. Int. J. Mol. Sci. 2022, 23, 4883. [Google Scholar] [CrossRef]
- Henning, Y.; Blind, U.S.; Larafa, S.; Matschke, J.; Fandrey, J. Hypoxia aggravates ferroptosis in RPE cells by promoting the Fenton reaction. Cell Death Dis. 2022, 13, 662. [Google Scholar] [CrossRef]
- McMenamin, P.G.; Saban, D.R.; Dando, S.J. Immune cells in the retina and choroid: Two different tissue environments that require different defenses and surveillance. Prog. Retin. Eye Res. 2019, 70, 85–98. [Google Scholar] [CrossRef]
- Marazita, M.C.; Dugour, A.; Marquioni-Ramella, M.D.; Figueroa, J.M.; Suburo, A.M. Oxidative stress-induced premature senescence dysregulates VEGF and CFH expression in retinal pigment epithelial cells: Implications for Age-related Macular Degeneration. Redox Biol. 2016, 7, 78–87. [Google Scholar] [CrossRef]
- Chen, Q.; Tang, L.; Zhang, Y.; Wan, C.; Yu, X.; Dong, Y.; Chen, X.; Wang, X.; Li, N.; Xin, G.; et al. STING up-regulates VEGF expression in oxidative stress-induced senescence of retinal pigment epithelium via NF-kappaB/HIF-1alpha pathway. Life Sci. 2022, 293, 120089. [Google Scholar] [CrossRef]
- Eamegdool, S.S.; Sitiwin, E.I.; Cioanca, A.V.; Madigan, M.C. Extracellular matrix and oxidative stress regulate human retinal pigment epithelium growth. Free Radic. Biol. Med. 2020, 146, 357–371. [Google Scholar] [CrossRef]
- Naylor, A.; Hopkins, A.; Hudson, N.; Campbell, M. Tight Junctions of the Outer Blood Retina Barrier. Int. J. Mol. Sci. 2019, 21, 211. [Google Scholar] [CrossRef] [PubMed]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Vaz, J.; Bernardes, R.; Lobo, C. Blood-retinal barrier. Eur. J. Ophthalmol. 2011, 21 (Suppl. 6), S3–S9. [Google Scholar] [CrossRef]
- Fields, M.A.; Del Priore, L.V.; Adelman, R.A.; Rizzolo, L.J. Interactions of the choroid, Bruch’s membrane, retinal pigment epithelium, and neurosensory retina collaborate to form the outer blood-retinal-barrier. Prog. Retin. Eye Res. 2020, 76, 100803. [Google Scholar] [CrossRef] [PubMed]
- Chirco, K.R.; Sohn, E.H.; Stone, E.M.; Tucker, B.A.; Mullins, R.F. Structural and molecular changes in the aging choroid: Implications for age-related macular degeneration. Eye 2017, 31, 10–25. [Google Scholar] [CrossRef]
- Rafii, S.; Butler, J.M.; Ding, B.S. Angiocrine functions of organ-specific endothelial cells. Nature 2016, 529, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.S.; Nolan, D.J.; Butler, J.M.; James, D.; Babazadeh, A.O.; Rosenwaks, Z.; Mittal, V.; Kobayashi, H.; Shido, K.; Lyden, D.; et al. Inductive angiocrine signals from sinusoidal endothelium are required for liver regeneration. Nature 2010, 468, 310–315. [Google Scholar] [CrossRef]
- Ding, B.S.; Nolan, D.J.; Guo, P.; Babazadeh, A.O.; Cao, Z.; Rosenwaks, Z.; Crystal, R.G.; Simons, M.; Sato, T.N.; Worgall, S.; et al. Endothelial-derived angiocrine signals induce and sustain regenerative lung alveolarization. Cell 2011, 147, 539–553. [Google Scholar] [CrossRef]
- Benedicto, I.; Lehmann, G.L.; Ginsberg, M.; Nolan, D.J.; Bareja, R.; Elemento, O.; Salfati, Z.; Alam, N.M.; Prusky, G.T.; Llanos, P.; et al. Concerted regulation of retinal pigment epithelium basement membrane and barrier function by angiocrine factors. Nat. Commun. 2017, 8, 15374. [Google Scholar] [CrossRef]
- Spencer, C.; Abend, S.; McHugh, K.J.; Saint-Geniez, M. Identification of a synergistic interaction between endothelial cells and retinal pigment epithelium. J. Cell Mol. Med. 2017, 21, 2542–2552. [Google Scholar] [CrossRef]
- Hori, K.; Sen, A.; Artavanis-Tsakonas, S. Notch signaling at a glance. J. Cell Sci. 2013, 126, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.K.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 2014, 507, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.L.; Ruan, B.; Song, P.; Fang, Z.Q.; Yue, Z.S.; Liu, J.J.; Dou, G.R.; Han, H.; Wang, L. Shear stress-induced cellular senescence blunts liver regeneration through Notch-sirtuin 1-P21/P16 axis. Hepatology 2022, 75, 584–599. [Google Scholar] [CrossRef]
- Augustin, H.G.; Koh, G.Y. Organotypic vasculature: From descriptive heterogeneity to functional pathophysiology. Science 2017, 357, eaal2379. [Google Scholar] [CrossRef] [PubMed]
- Dou, G.R.; Wang, Y.C.; Hu, X.B.; Hou, L.H.; Wang, C.M.; Xu, J.F.; Wang, Y.S.; Liang, Y.M.; Yao, L.B.; Yang, A.G.; et al. RBP-J, the transcription factor downstream of Notch receptors, is essential for the maintenance of vascular homeostasis in adult mice. FASEB J. 2008, 22, 1606–1617. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, Z.; Chen, Y.; Li, N.; Wang, L.; Dou, G.; Liu, Y.; Duan, J.; Feng, L.; Deng, S.; et al. Endothelial cells-targeted soluble human Delta-like 4 suppresses both physiological and pathological ocular angiogenesis. Sci. China Life Sci. 2015, 58, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.C.; Dou, G.R.; Wang, L.; Liang, L.; Tian, D.M.; Cao, X.L.; Qin, H.Y.; Wang, C.M.; Zhang, P.; Han, H. Inhibition of tumor angiogenesis and tumor growth by the DSL domain of human Delta-like 1 targeted to vascular endothelial cells. Neoplasia 2013, 15, 815–825. [Google Scholar] [CrossRef]
- Taylor, S.R.; Markesbery, M.G.; Harding, P.A. Heparin-binding epidermal growth factor-like growth factor (HB-EGF) and proteolytic processing by a disintegrin and metalloproteinases (ADAM): A regulator of several pathways. Semin. Cell Dev. Biol. 2014, 28, 22–30. [Google Scholar] [CrossRef]
- Ongusaha, P.P.; Kwak, J.C.; Zwible, A.J.; Macip, S.; Higashiyama, S.; Taniguchi, N.; Fang, L.; Lee, S.W. HB-EGF is a potent inducer of tumor growth and angiogenesis. Cancer Res. 2004, 64, 5283–5290. [Google Scholar] [CrossRef]
- Hollborn, M.; Iandiev, I.; Seifert, M.; Schnurrbusch, U.E.; Wolf, S.; Wiedemann, P.; Bringmann, A.; Kohen, L. Expression of HB-EGF by retinal pigment epithelial cells in vitreoretinal proliferative disease. Curr. Eye Res. 2006, 31, 863–874. [Google Scholar] [CrossRef]
- Kushwaha, R.; Mishra, J.; Tripathi, S.; Raza, W.; Mandrah, K.; Roy, S.K.; Bandyopadhyay, S. Arsenic Attenuates Heparin-Binding EGF-like Growth Factor/EGFR Signaling That Promotes Matrix Metalloprotease 9-Dependent Astrocyte Damage in the Developing Rat Brain. Toxicol. Sci. 2018, 162, 406–428. [Google Scholar] [CrossRef] [PubMed]
- Ji Cho, M.; Yoon, S.J.; Kim, W.; Park, J.; Lee, J.; Park, J.G.; Cho, Y.L.; Hun Kim, J.; Jang, H.; Park, Y.J.; et al. Oxidative stress-mediated TXNIP loss causes RPE dysfunction. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.S.; Fischer, A. Notch Signaling in the Vasculature: Angiogenesis and Angiocrine Functions. Cold Spring Harb. Perspect. Med. 2023, 13, a041166. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, H.; Nielsen, C.M.; Simonetto, D.A.; Ferrell, L.; Shah, V.H.; Wang, R.A. Endothelial notch signaling is essential to prevent hepatic vascular malformations in mice. Hepatology 2016, 64, 1302–1316. [Google Scholar] [CrossRef]
- Stegen, S.; Carmeliet, G. The skeletal vascular system—Breathing life into bone tissue. Bone 2018, 115, 50–58. [Google Scholar] [CrossRef]
- Duan, J.L.; Ruan, B.; Yan, X.C.; Liang, L.; Song, P.; Yang, Z.Y.; Liu, Y.; Dou, K.F.; Han, H.; Wang, L. Endothelial Notch activation reshapes the angiocrine of sinusoidal endothelia to aggravate liver fibrosis and blunt regeneration in mice. Hepatology 2018, 68, 677–690. [Google Scholar] [CrossRef]
- Yang, J.M.; Park, C.S.; Kim, S.H.; Noh, T.W.; Kim, J.H.; Park, S.; Lee, J.; Park, J.R.; Yoo, D.; Jung, H.H.; et al. Dll4 Suppresses Transcytosis for Arterial Blood-Retinal Barrier Homeostasis. Circ. Res. 2020, 126, 767–783. [Google Scholar] [CrossRef]
- Mammadzada, P.; Corredoira, P.M.; Andre, H. The role of hypoxia-inducible factors in neovascular age-related macular degeneration: A gene therapy perspective. Cell Mol. Life Sci. 2020, 77, 819–833. [Google Scholar] [CrossRef]
- Kaur, C.; Foulds, W.S.; Ling, E.A. Blood-retinal barrier in hypoxic ischaemic conditions: Basic concepts, clinical features and management. Prog. Retin. Eye Res. 2008, 27, 622–647. [Google Scholar] [CrossRef]
- Wang, S.; Du, S.; Wu, Q.; Hu, J.; Li, T. Decorin Prevents Retinal Pigment Epithelial Barrier Breakdown under Diabetic Conditions by Suppressing p38 MAPK Activation. Investig. Opthalmol. Vis. Sci. 2015, 56, 2971–2979. [Google Scholar] [CrossRef]
- Skottman, H.; Muranen, J.; Lahdekorpi, H.; Pajula, E.; Makela, K.; Koivusalo, L.; Koistinen, A.; Uusitalo, H.; Kaarniranta, K.; Juuti-Uusitalo, K. Contacting co-culture of human retinal microvascular endothelial cells alters barrier function of human embryonic stem cell derived retinal pigment epithelial cells. Exp. Cell Res. 2017, 359, 101–111. [Google Scholar] [CrossRef]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma 2014, 23, S20–S23. [Google Scholar] [CrossRef] [PubMed]
- Hubmacher, D.; Apte, S.S. The biology of the extracellular matrix: Novel insights. Curr. Opin. Rheumatol. 2013, 25, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Al-Ubaidi, M.R.; Naash, M.I.; Conley, S.M. A perspective on the role of the extracellular matrix in progressive retinal degenerative disorders. Investig. Ophthalmol. Vis. Sci. 2013, 54, 8119–8124. [Google Scholar] [CrossRef] [PubMed]
- Reinhard, J.; Joachim, S.C.; Faissner, A. Extracellular matrix remodeling during retinal development. Exp. Eye Res. 2015, 133, 132–140. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Concentration | Catalogue Number | Company |
|---|---|---|---|
| ZO-1 | 1:200 | ab221547 | Abcam |
| Fibronectin | 1:200 | ab268020 | Abcam |
| CollagenIV | 1:500 | sc-59814 | Santa Cruz |
| isolectin-B4 | 1:100 | FL-1201 | Vectorlabs |
| Gene Name | Species | Gene Accession Number | Primers | Primer Sequence 5′→3′ | Product Length (bp) |
|---|---|---|---|---|---|
| β-actin | human | NM_001101.5 | Forward | CTCCATCCTGGCCTCGCTGT | 268 |
| Reverse | GCTGTCACCTTCACCGTTCC | ||||
| ZO-1 | human | NM_001301025.3 | Forward | ACCAGTAAGTCGTCCTGATCC | 128 |
| Reverse | TCGGCCAAATCTTCTCACTCC | ||||
| Hey1 | human | NM_001040708.2 | Forward | GAGTGCGGACGAGAATGGAA | 115 |
| Reverse | TCGTCGGCGCTTCTCAATTA | ||||
| Hes1 | human | NM_005524.4 | Forward | GGAAATGACAGTGAAGCACCTCC | 130 |
| Reverse | GAAGCGGGTCACCTCGTTCATG | ||||
| Collagen IV | human | NM_000092.5 | Forward | AGAGATTGCTCTGTTTGCCAC | 143 |
| Reverse | CGGTCCCCTCTCATTCCTT | ||||
| MMP-2 | human | NM_001127891.3 | Forward | CTTCCAAGTCTGGAGCGATGT | 119 |
| Reverse | TACCGTCAAAGGGGTATCCAT | ||||
| DCN | human | NM_001920.5 | Forward | TCACAGAGCAGCACCTACCC | 139 |
| Reverse | TTCACAACCAGGGAACCTTTTAAT | ||||
| Notch1 | human | NM_017617.5 | Forward | GGTGAACTGCTCTGAGGAGATC | 150 |
| Reverse | GGATTGCAGTCGTCCACGTTGA | ||||
| DLL4 | human | NM_019074.4 | Forward | CTGCGAGAAGAAAGTGGACAGG | 139 |
| Reverse | ACAGTCGCTGACGTGGAGTTCA | ||||
| HBEGF | human | NM_001945.3 | Forward | CATCGTGGGGCTTCTCATGT | 126 |
| Reverse | CCAGCCGATTCCTTGAGCA | ||||
| MMP-9 | human | NM_004994.3 | Forward | TGTACCGCTATGGTTACACTCG | 97 |
| Reverse | GGCAGGGACAGTTGCTTCT | ||||
| β-actin | mouse | NM_007393.5 | Forward | TATAAAACCCGGCGGCGCA | 117 |
| Reverse | TCATCCATGGCGAACTGGTG | ||||
| Fibronectin | mouse | NM_010233.2 | Forward | CCCTATCTCTGATACCGTTGTCC | 145 |
| Reverse | TGCCGCAACTACTGTGATTCGG | ||||
| Collagen IV | mouse | NM_007735.2 | Forward | GAACCTGGAAGAAAGGGAGAGG | 123 |
| Reverse | GGAAGTGACTGCTTCTCCTGCA | ||||
| LOX | mouse | NM_010728.4 | Forward | TGCACACACACAGGGATTGA | 185 |
| Reverse | TGTAGCGAATGTCACAGCGT | ||||
| DCN | mouse | NM_007833.6 | Forward | ACTCTCCAGGAACTTCGTGTCC | 146 |
| Reverse | AGTCCCTGGAAGGCTCCGTTTT |
| Antibody | Concentration | Catalog Number | Company |
|---|---|---|---|
| ZO-1 | 1:1000 | 13663S | CST |
| HBEGF | 1:1000 | ab185555 | Abcam |
| MMP-9 | 1:1000 | ab76003 | Abcam |
| β-tublin | 1:1000 | 2148 | CST |
| β-actin | 1:1000 | ab5316 | Abcam |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Xi, Y.; Jing, Y.; Zhou, Z.; Sun, X.; Zhang, G.; Yuan, T.; Chang, T.; Dou, G. Endothelial Notch Signaling Regulates the Function of the Retinal Pigment Epithelial Barrier via EC Angiocrine Signaling. Antioxidants 2023, 12, 1979. https://doi.org/10.3390/antiox12111979
Niu Y, Xi Y, Jing Y, Zhou Z, Sun X, Zhang G, Yuan T, Chang T, Dou G. Endothelial Notch Signaling Regulates the Function of the Retinal Pigment Epithelial Barrier via EC Angiocrine Signaling. Antioxidants. 2023; 12(11):1979. https://doi.org/10.3390/antiox12111979
Chicago/Turabian StyleNiu, Yali, Yixuan Xi, Yutong Jing, Ziyi Zhou, Xiaojia Sun, Guoheng Zhang, Tianhao Yuan, Tianfang Chang, and Guorui Dou. 2023. "Endothelial Notch Signaling Regulates the Function of the Retinal Pigment Epithelial Barrier via EC Angiocrine Signaling" Antioxidants 12, no. 11: 1979. https://doi.org/10.3390/antiox12111979