
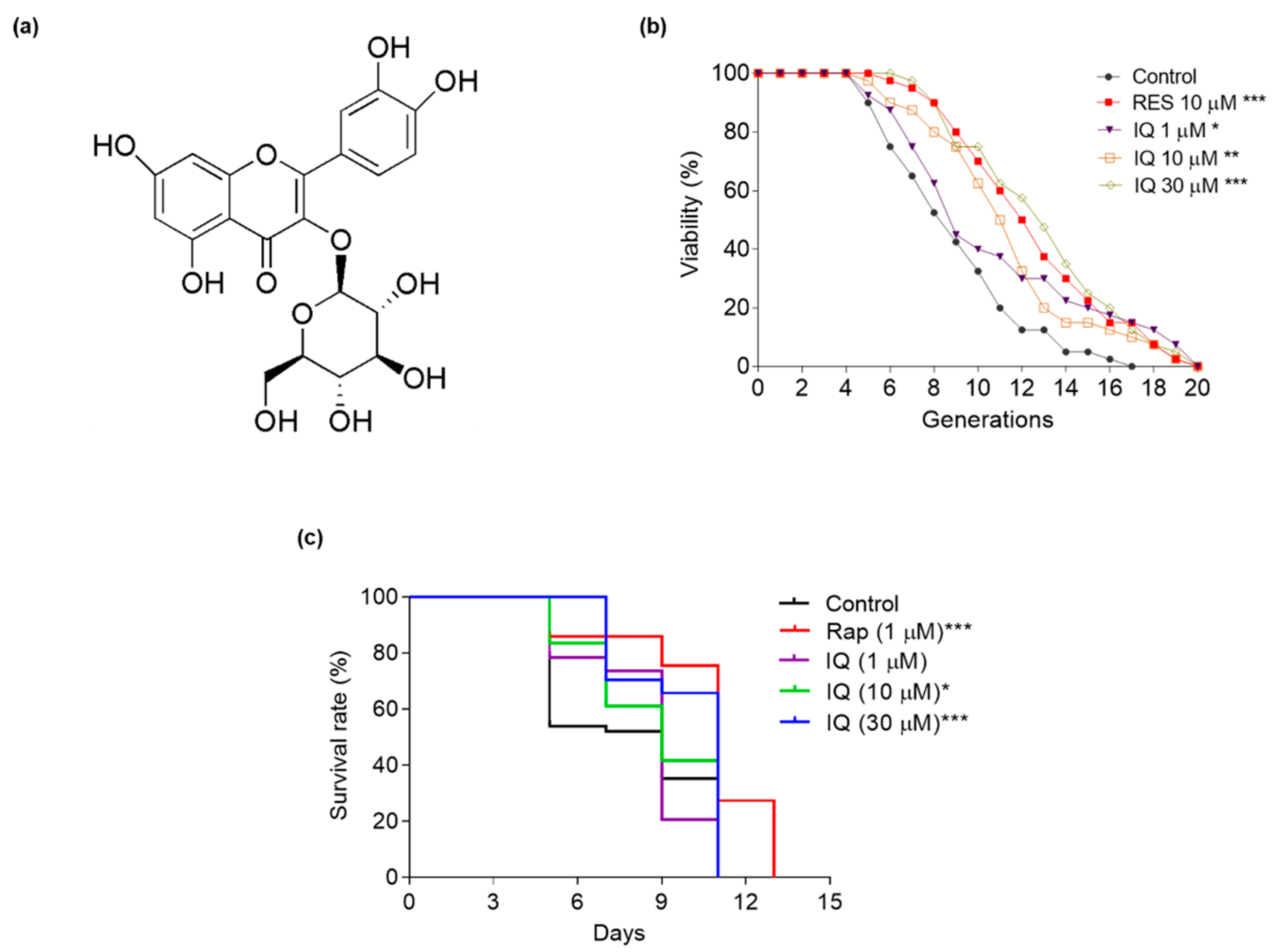
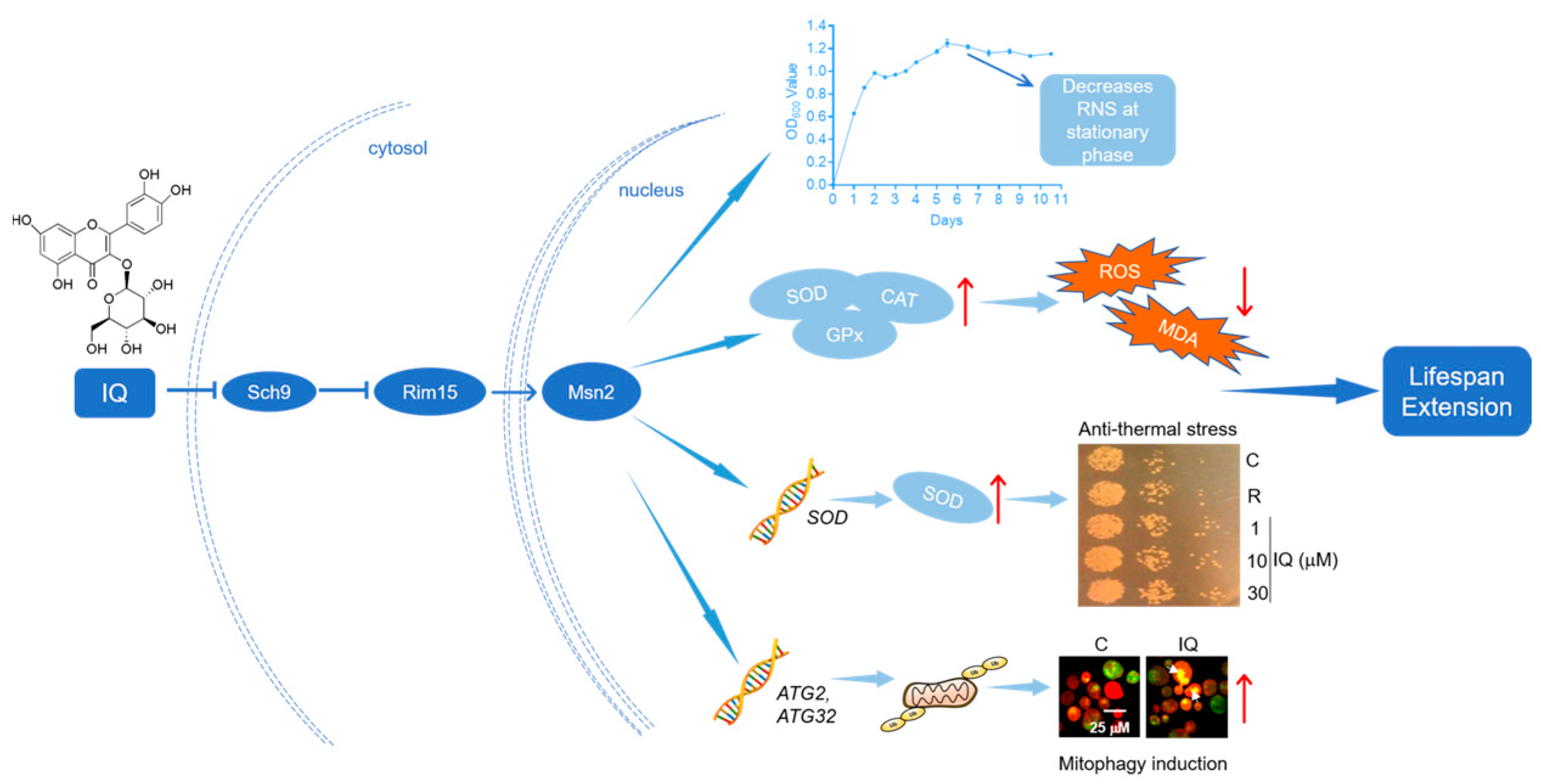
Isoquercitrin from Apocynum venetum L. Exerts Antiaging Effects on Yeasts via Stress Resistance Improvement and Mitophagy Induction through the Sch9/Rim15/Msn Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Isolation and Purification of Isoquercitrin
2.3. Yeast Strains, Culture Medium, and Lifespan Assay
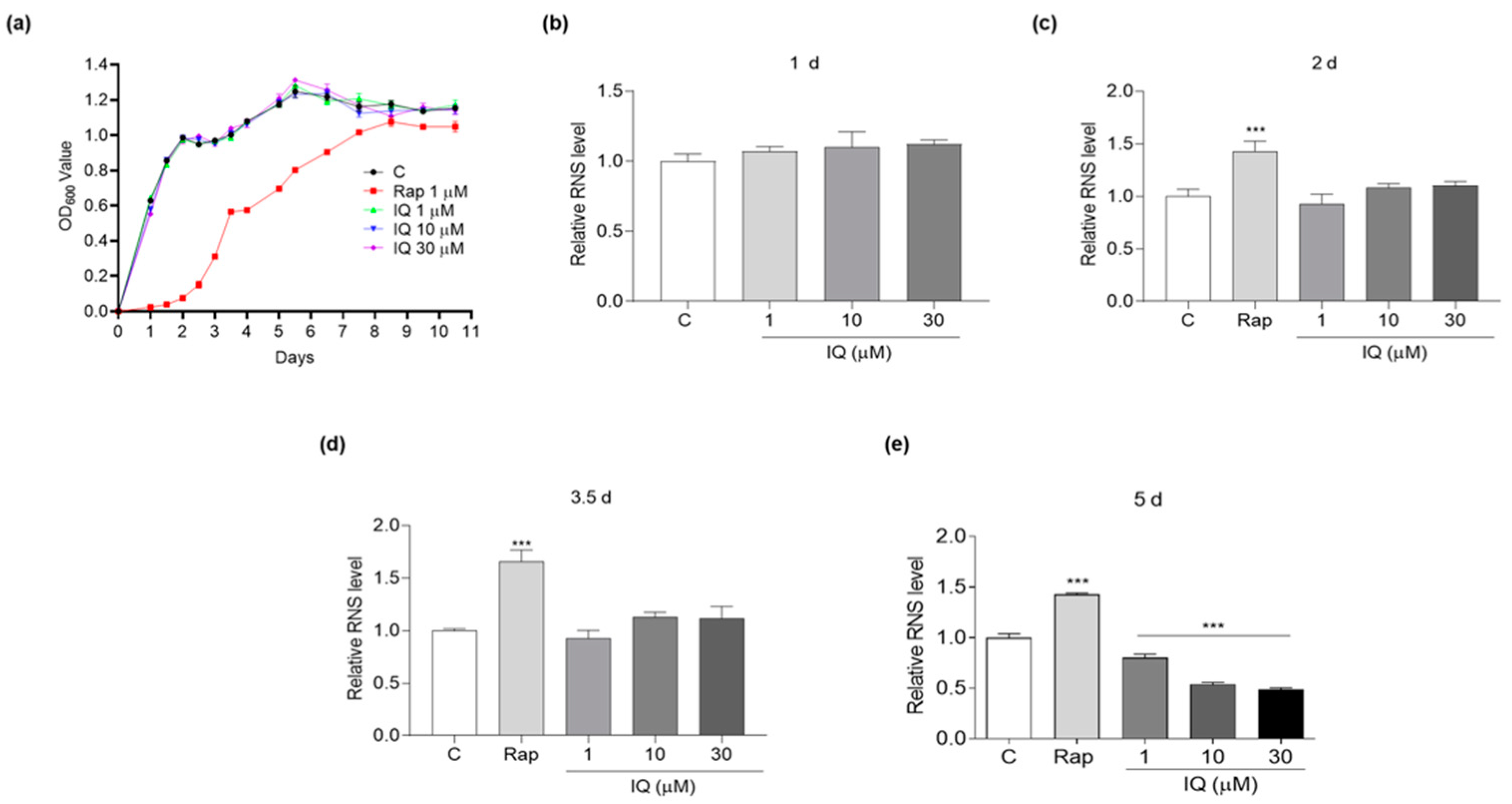
2.4. Growth Curve Determination
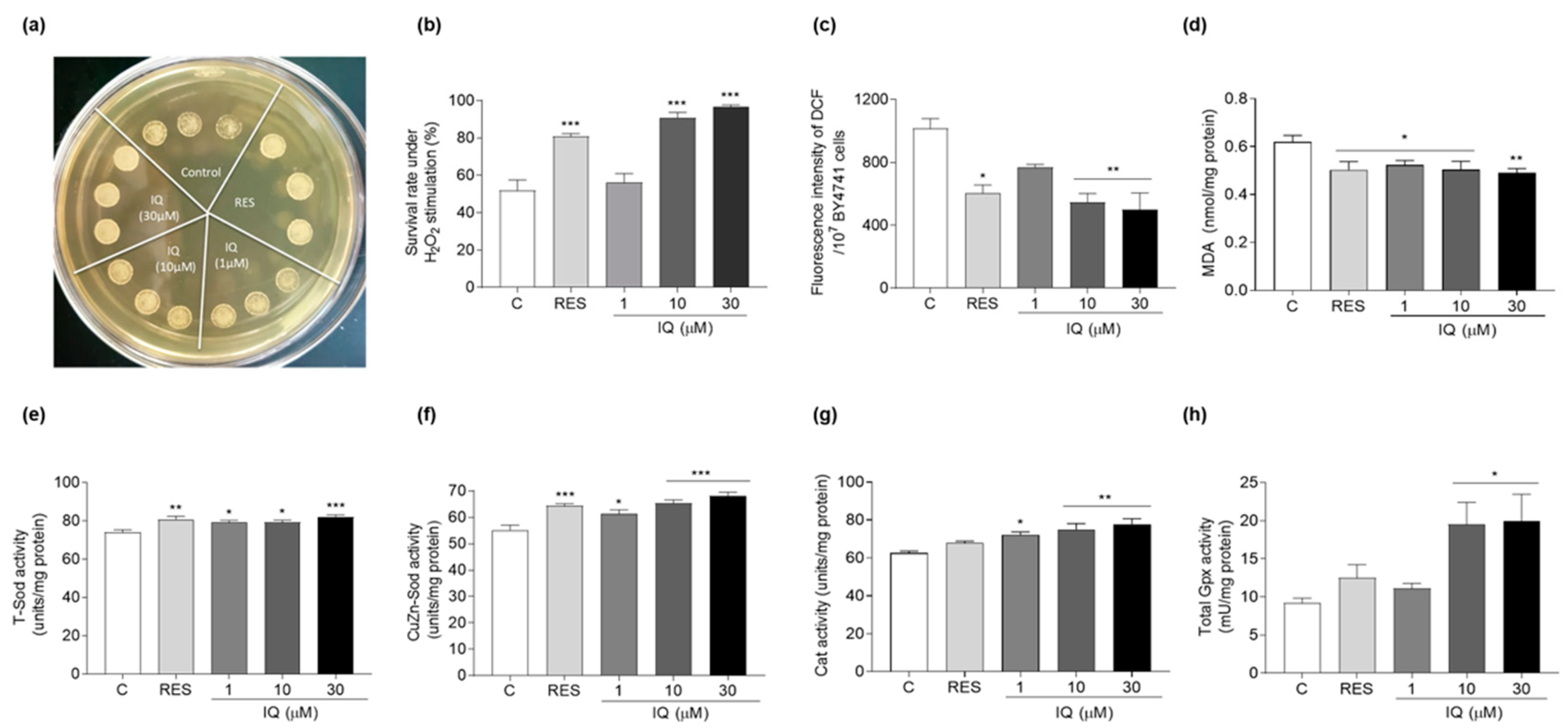
2.5. Determination of Yeast Survival Ability under H2O2 Stimulation
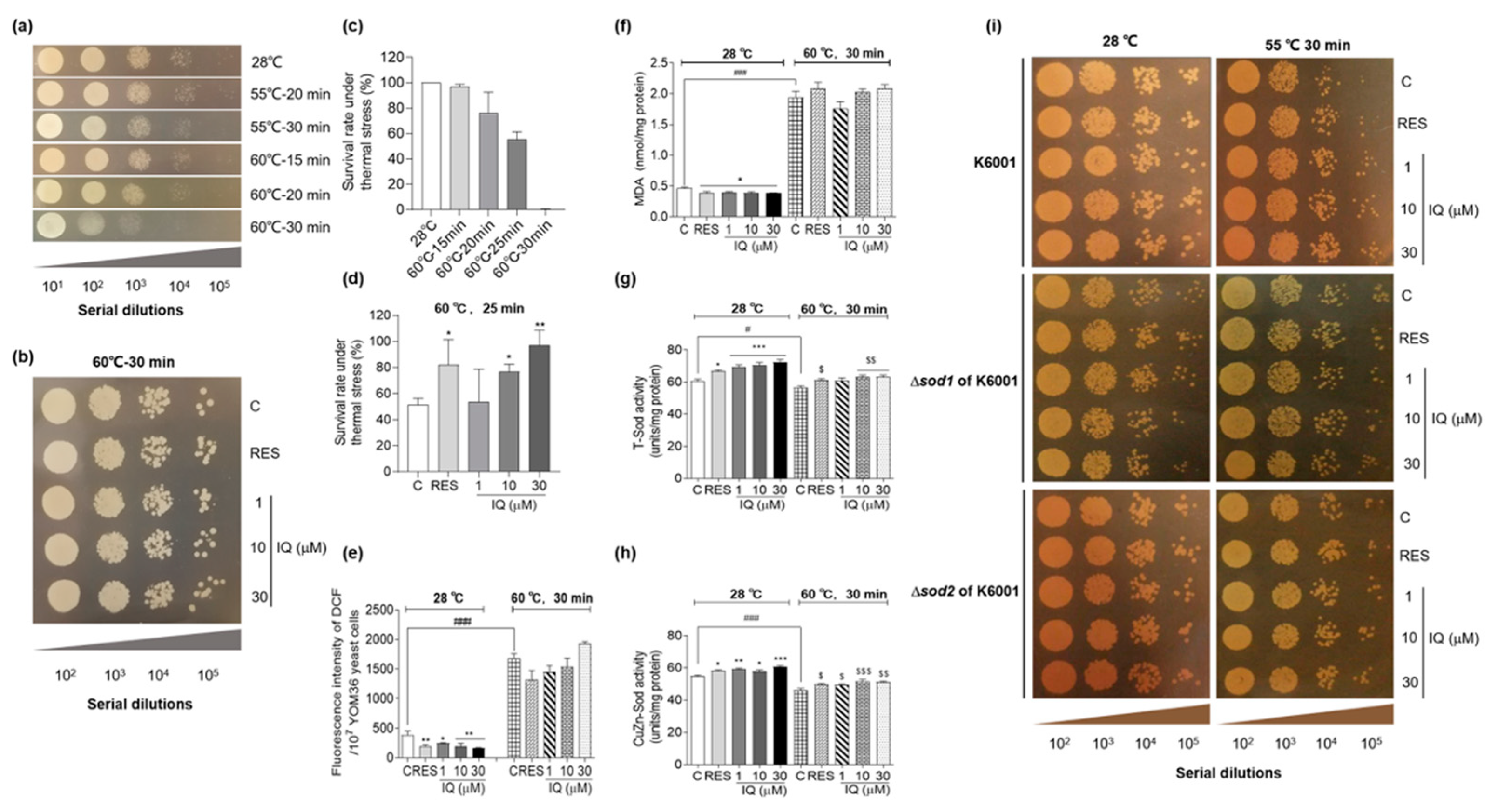
2.6. Thermal-Stress-Resistance Assay
2.7. Measurement of ROS, RNS, and MDA Levels of Yeasts
2.8. Antioxidant Enzyme Activity Determination
2.9. Real-Time Polymerase Chain Reaction (RT-PCR) Analysis
2.10. Observation of Autophagy and Mitophagy in Yeasts
2.11. Observation of the Nuclear Translocation of GFP-Rim15 and GFP-Msn2 in Yeasts
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
3.1. IQ Extends the Lifespans of Yeasts
3.2. The Effect of IQ on RNS of Yeasts during Chronological Aging
3.3. IQ Increases the Antioxidative-Stress Activity of Yeasts
3.4. IQ Confers Thermotolerance on Yeasts via SOD
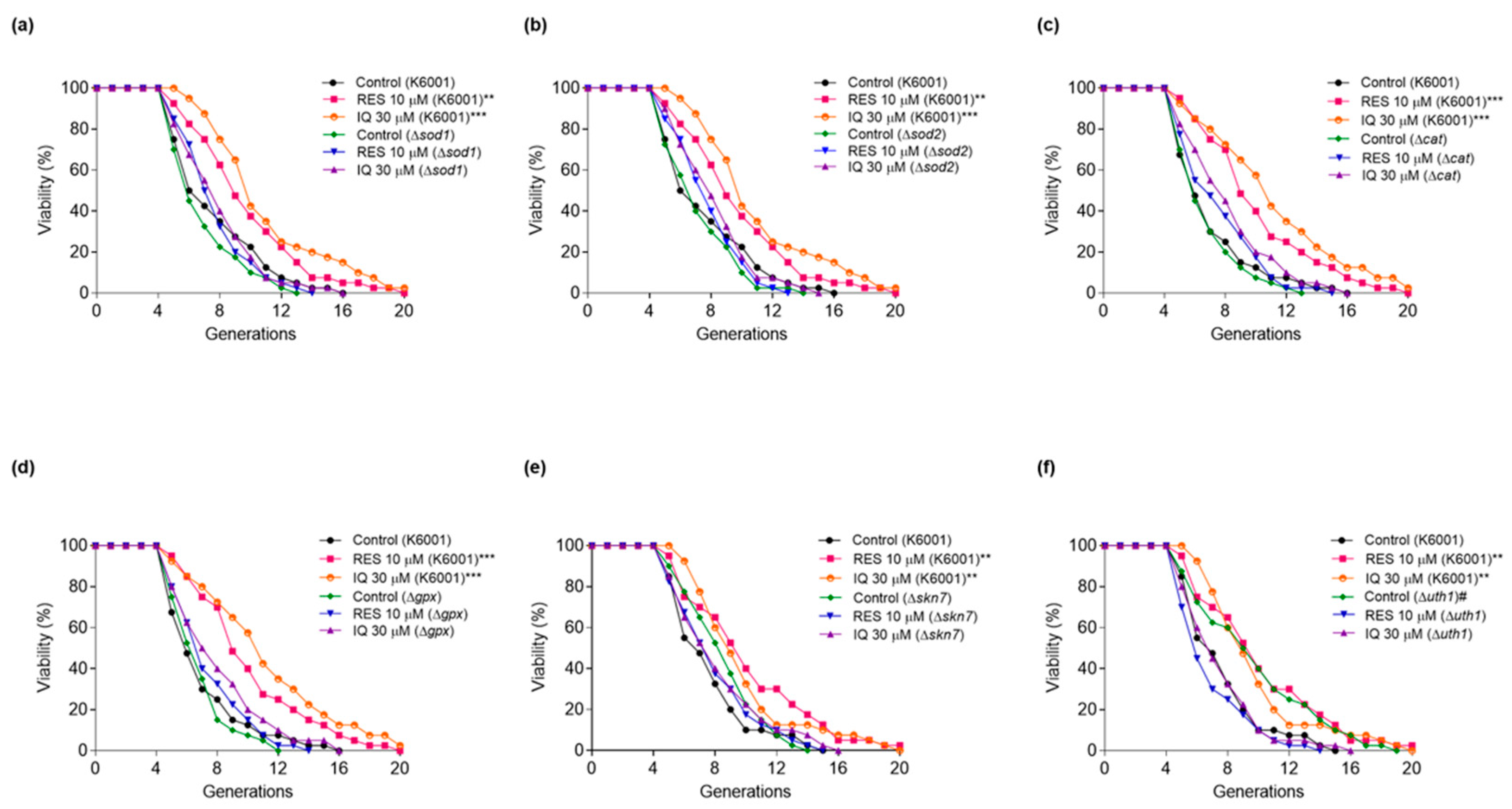
3.5. SOD1, SOD2, CAT, GPx, SKN7, and UTH1 Are Involved in the Antiaging Effect of IQ
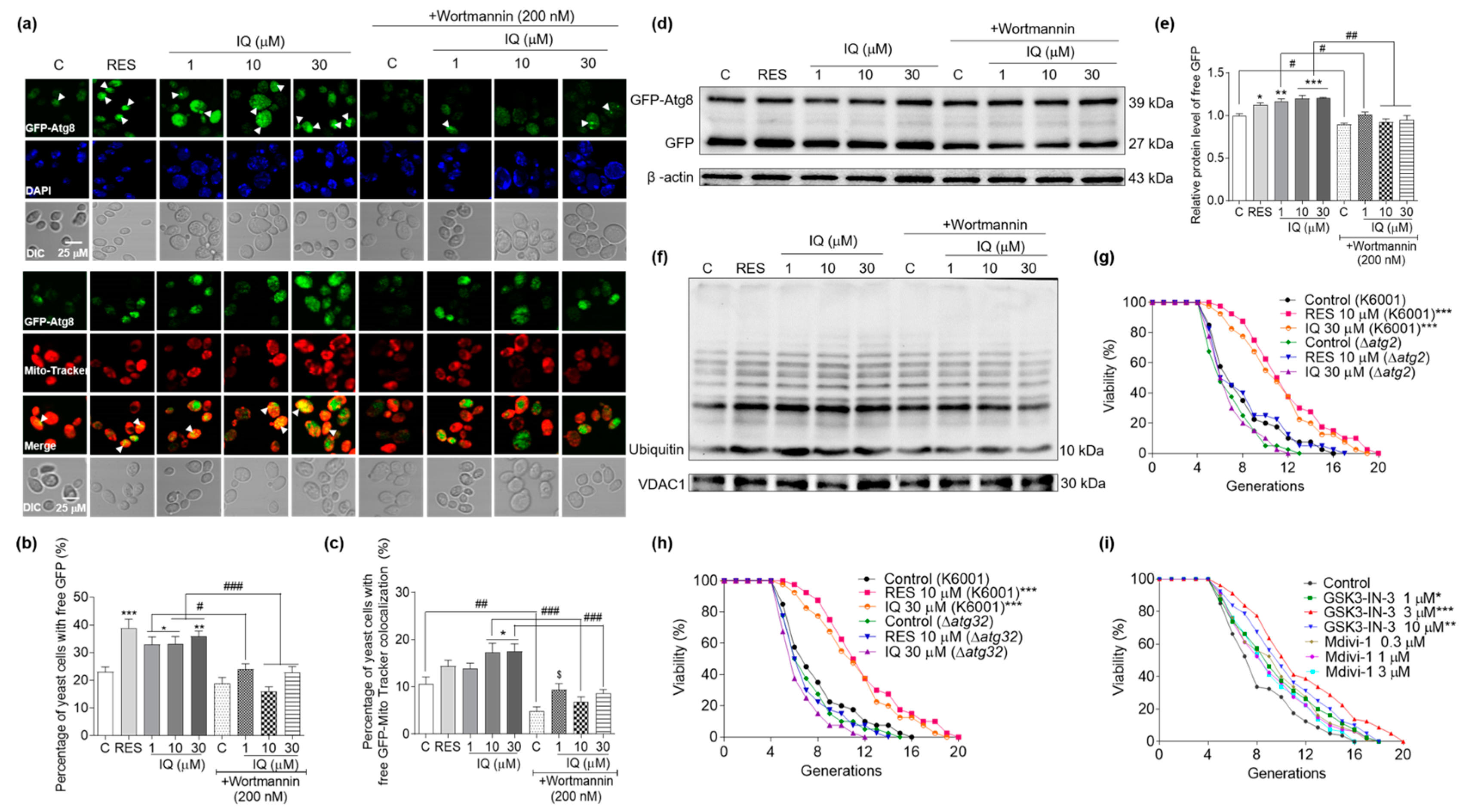
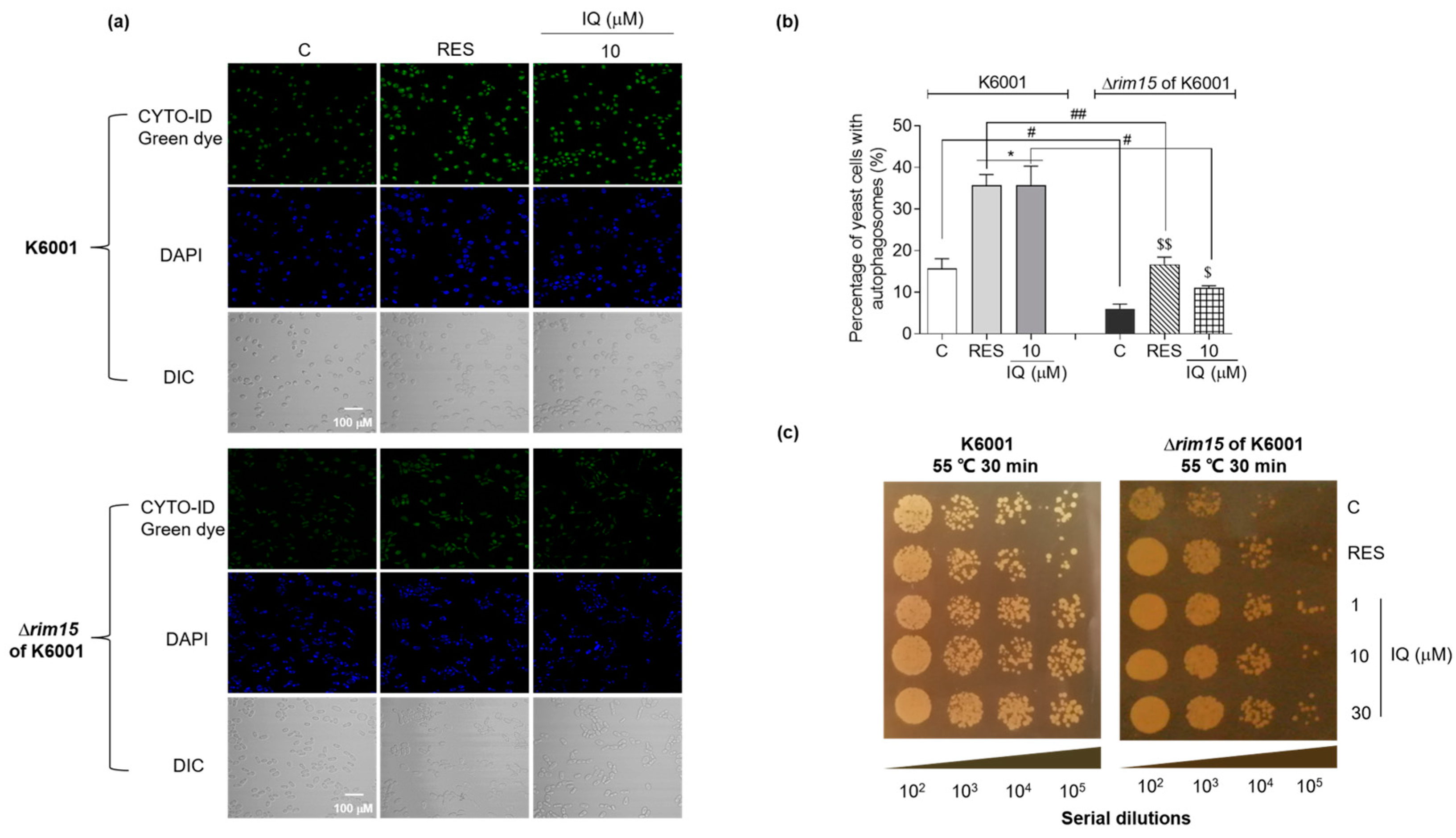
3.6. Effects of IQ on Autophagy and Mitophagy in YOM38 Yeasts
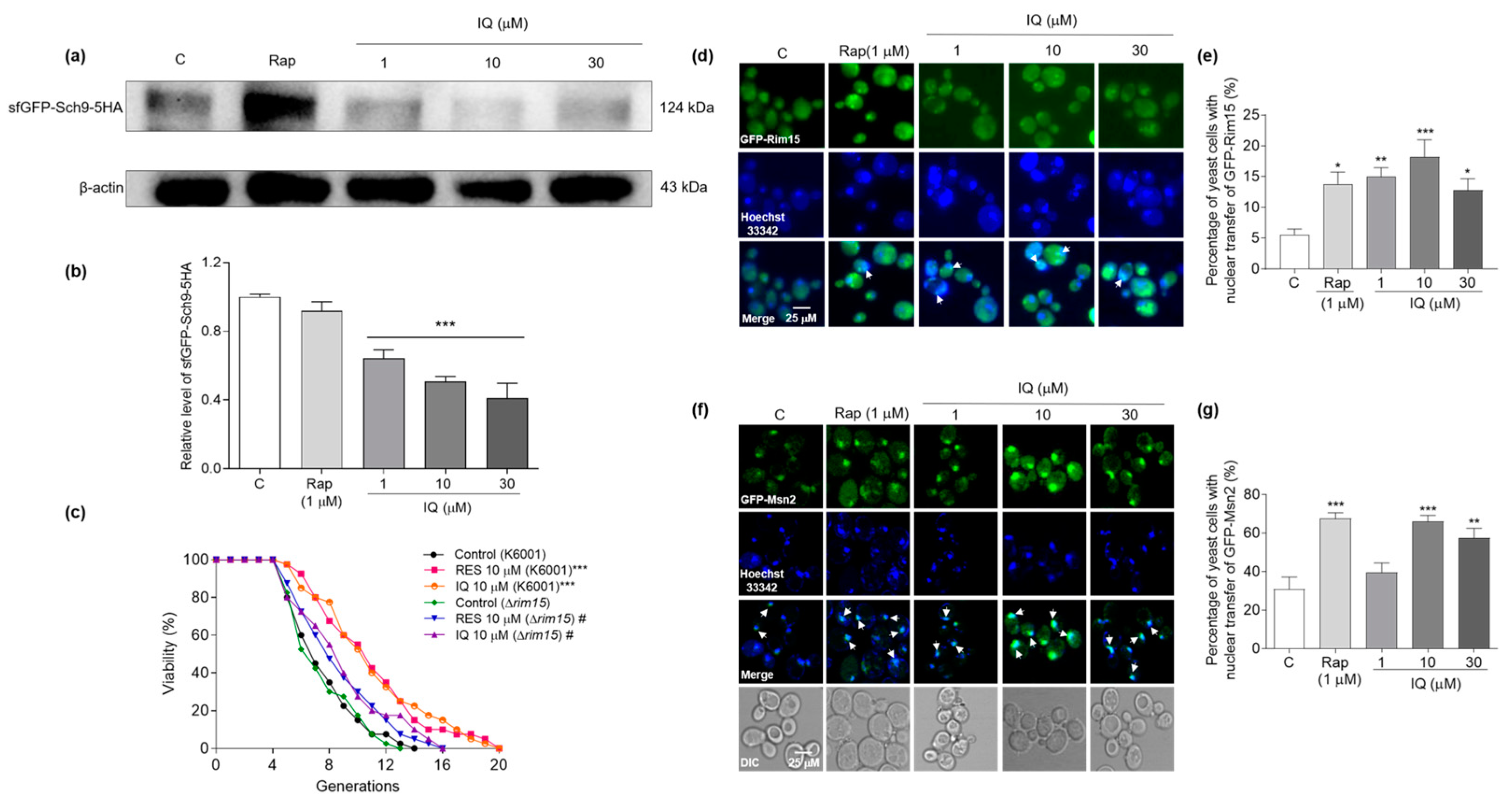
3.7. IQ Inhibits the Expression of Sch9 and Promotes GFP-Rim15 and GFP-Msn2 Transfer into the Nucleus in Yeasts
3.8. IQ Exerts Autophagy-Induction and Thermal-Stress-Resistance Effects via Rim15
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| MDA | Malondialdehyde |
| Sod | Superoxide dismutase |
| Cat | Catalase |
| Gpx | Glutathione peroxidase |
| GFP | Green fluorescent protein |
| RES | Resveratrol |
| YPD | Yeast peptone dextrose |
| SD | Synthetic defined |
References
- Dogra, S.; Dunstan, D.W.; Sugiyama, T.; Stathi, A.; Gardiner, P.A.; Owen, N. Active aging and public health: Evidence, implications, and opportunities. Annu. Rev. Public Health 2022, 43, 439–459. [Google Scholar] [PubMed]
- Saul, D.; Kosinsky, R.L. Epigenetics of aging and aging-associated diseases. Int. J. Mol. Sci. 2021, 22, 401. [Google Scholar] [PubMed]
- Dutta, N.; Garcia, G.; Higuchi-Sanabria, R. Hijacking cellular stress responses to promote lifespan. Front. Aging 2022, 3, 860404. [Google Scholar] [PubMed]
- Farooqui, T.; Farooqui, A.A. Aging: An important factor for the pathogenesis of neurodegenerative diseases. Mech. Ageing Dev. 2009, 130, 203–215. [Google Scholar] [PubMed]
- Burstein, M.T.; Kyryakov, P.; Beach, A.; Richard, V.R.; Koupaki, O.; Gomez-Perez, A.; Leonov, A.; Levy, S.; Noohi, F.; Titorenko, V.I. Lithocholic acid extends longevity of chronologically aging yeast only if added at certain critical periods of their lifespan. Cell Cycle 2012, 11, 3443–3462. [Google Scholar]
- Zeng, L.; Sun, C.; Pei, Z.; Yun, T.; Fan, S.; Long, S.; Wu, T.; Chen, Z.; Yang, Z.; Xu, F. Liangyi Gao extends lifespan and exerts an antiaging effect in Caenorhabditis elegans by modulating DAF-16/FOXO. Biogerontology 2019, 20, 665–676. [Google Scholar]
- Zhang, H.; Gong, W.; Wu, S.; Perrett, S. Hsp70 in redox homeostasis. Cells 2022, 11, 829. [Google Scholar]
- Ichimiya, T.; Yamakawa, T.; Hirano, T.; Yokoyama, Y.; Hayashi, Y.; Hirayama, D.; Wagatsuma, K.; Itoi, T.; Nakase, H. Autophagy and autophagy-related diseases: A Review. Int. J. Mol. Sci. 2020, 21, 8974. [Google Scholar]
- Yu, Z.; Sun, L.; Jiang, Z.; Liu, X.; Zhao, D.; Wang, H.; He, W.; Dong, M.; Du, L. Atg38-Atg8 interaction in fission yeast establishes a positive feedback loop to promote autophagy. Autophagy 2020, 16, 2036–2051. [Google Scholar] [CrossRef]
- Okamoto, K.; Kondo-Okamoto, N.; Ohsumi, Y. Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy. Dev. Cell 2009, 17, 87–97. [Google Scholar]
- Xu, P.; Damschroder, D.; Zhang, M.; Ryall, K.A.; Adler, P.N.; Saucerman, J.; Wessells, R.J.; Yan, Z. Atg2, Atg9 and Atg18 in mitochondrial integrity, cardiac function and healthspan in Drosophila. J. Mol. Cell Cardiol. 2019, 127, 116–124. [Google Scholar] [PubMed]
- Liu, Y.; Shi, Y.; Han, R.; Liu, C.; Qin, X.; Li, P.; Gu, R. Signaling pathways of oxidative stress response: The potential therapeutic targets in gastric cancer. Front. Immunol. 2023, 14, 1139589. [Google Scholar] [PubMed]
- Deprez, M.A.; Eskes, E.; Winderickx, J.; Wilms, T. The TORC1-Sch9 pathway as a crucial mediator of chronological lifespan in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, foy048. [Google Scholar]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab. 2012, 16, 18–31. [Google Scholar]
- Wei, M.; Fabrizio, P.; Hu, J.; Ge, H.; Cheng, C.; Li, L.; Longo, V.D. Life span extension by calorie restriction depends on Rim15 and transcription factors downstream of Ras/PKA, Tor, and Sch9. PLoS Genet. 2008, 4, e13. [Google Scholar] [CrossRef]
- Yorimitsu, T.; Zaman, S.; Broach, J.R.; Klionsky, D.J. Protein kinase A and Sch9 cooperatively regulate induction of autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell. 2007, 18, 4180–4189. [Google Scholar] [CrossRef]
- Oliveira, A.V.; Vilaça, R.; Santos, C.N.; Costa, V.; Menezes, R. Exploring the power of yeast to model aging and age-related neurodegenerative disorders. Biogerontology 2017, 18, 3–34. [Google Scholar] [PubMed]
- Jarolim, S.; Millen, J.; Heeren, G.; Laun, P.; Goldfarb, D.S.; Breitenbach, M. A novel assay for replicative lifespan in Saccharomyces cerevisiae. FEMS Yeast Res. 2004, 5, 169–177. [Google Scholar] [CrossRef]
- Maruyama, Y.; Ito, T.; Kodama, H.; Matsuura, A. Availability of amino acids extends chronological lifespan by suppressing hyper-acidification of the environment in Saccharomyces cerevisiae. PLoS ONE 2016, 11, e0151894. [Google Scholar]
- Xie, W.; Zhang, X.; Wang, T.; Hu, J. Botany, traditional uses, phytochemistry and pharmacology of Apocynum venetum L. (Luobuma): A review. J. Ethnopharmacol. 2012, 141, 1–8. [Google Scholar]
- Shen, Y.; Zhang, Q.; Huang, Z.; Zhu, J.; Qiu, J.; Ma, W.; Yang, X.; Ding, F.; Sun, H. Isoquercitrin delays denervated soleus muscle atrophy by inhibiting oxidative stress and inflammation. Front. Physiol. 2020, 11, 988. [Google Scholar] [CrossRef] [PubMed]
- Shui, L.; Wang, W.; Xie, M.; Ye, B.; Li, X.; Liu, Y.; Zheng, M. Isoquercitrin induces apoptosis and autophagy in hepatocellular carcinoma cells via AMPK/mTOR/p70S6K signaling pathway. Aging 2020, 12, 24318–24332. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.; Muroi, M.; Ogawa, N.; Kobayashi, H.; Nishimura, H.; Chen, D.; Fasina, O.B.; Wang, J.; Osada, H.; Yoshida, M.; et al. Isoquercitrin from Apocynum venetum L. produces an anti-obesity effect on obese mice by targeting C-1-tetrahydrofolate synthase, carbonyl reductase, and glutathione S-transferase P and modification of the AMPK/SREBP-1c/FAS/CD36 signaling pathway in mice in vivo. Food Funct. 2022, 13, 10923–10936. [Google Scholar]
- Lee, S.; Lee, J.; Lee, H.; Sung, J. Relative protective activities of quercetin, quercetin-3-glucoside, and rutin in alcohol-induced liver injury. J. Food Biochem. 2019, 43, e13002. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Jiang, Y.; Zhang, X.; Chen, X.; Liu, Z.; Tian, X. Isoquercetin ameliorates myocardial infarction through anti-inflammation and anti-apoptosis factor and regulating TLR4-NF-κB signal pathway. Mol. Med. Rep. 2018, 17, 6675–6680. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Q.; Chen, D.; Matsuura, A.; Xiang, L.; Qi, J. Inokosterone from Gentiana rigescens Franch extends the longevity of yeast and mammalian cells via antioxidative stress and mitophagy induction. Antioxidants 2022, 11, 214. [Google Scholar] [CrossRef]
- Disasa, D.; Cheng, L.; Manzoor, M.; Liu, Q.; Wang, Y.; Xiang, L.; Qi, J. Amarogentin from Gentiana rigescens Franch exhibits antiaging and neuroprotective effects through antioxidative stress. Oxid. Med. Cell Longev. 2020, 2020, 3184019. [Google Scholar] [CrossRef]
- Dakik, P.; Rodriguez, M.E.L.; Junio, J.A.B.; Mitrofanova, D.; Medkour, Y.; Tafakori, T.; Taifour, T.; Lutchman, V.; Samson, E.; Arlia-Ciommo, A.; et al. Discovery of fifteen new geroprotective plant extracts and identification of cellular processes they affect to prolong the chronological lifespan of budding yeast. Oncotarget 2020, 11, 2182–2203. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Zhang, L.; Dai, Y.; Yue, L.; Chen, G.; Yuan, L.; Zhang, S.; Li, F.; Zhang, H.; Li, G.; Zhu, S.; et al. Heat stress response in Chinese cabbage (Brassica rapa L.) revealed by transcriptome and physiological analysis. PeerJ 2022, 10, e13427. [Google Scholar] [CrossRef]
- Singh, A.K.; Kashyap, M.P.; Tripathi, V.K.; Singh, S.; Garg, G.; Rizvi, S.I. Neuroprotection through rapamycin-induced activation of autophagy and PI3K/Akt1/mTOR/CREB signaling against amyloid-β-induced oxidative stress, synaptic/neurotransmission dysfunction, and neurodegeneration in adult rats. Mol. Neurobiol. 2017, 54, 5815–5828. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Ordureau, A.; Heo, J.M. Building and decoding ubiquitin chains for mitophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, M.; Zhong, Y.; Ma, X.; Sun, S.; Xu, C.; Peng, L.; Li, G.; Zhang, L.; Liu, Z.; et al. A Glb1-2A-mCherry reporter monitors systemic aging and predicts lifespan in middle-aged mice. Nat. Commun. 2022, 13, 7028. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, C.; Lei, H.; Chen, C.; Cao, Z.; Song, Y.; Chen, G.; Wu, F.; Zhou, J.; Lu, Y.; et al. Structural insights into amelioration effects of quercetin and its glycoside derivatives on NAFLD in mice by modulating the gut microbiota and host metabolism. J. Agric. Food Chem. 2022, 70, 14732–14743. [Google Scholar] [CrossRef]
- Zheng, M.; Chen, S.; Liu, Y.; He, Y. α-Glucosidase inhibitory activities of constituents from Psidium guajava leaves. Nat. Prod. Res. 2023, 28, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Zheng, D.; Pi, W.; Qiu, Y.; Xia, K.; Guo, W. Isoquercitrin restrains the proliferation and promotes apoptosis of human osteosarcoma cells by inhibiting the Wnt/β-catenin pathway. J. Bone Oncol. 2023, 3, 100468. [Google Scholar] [CrossRef]
- Liu, J.; Hui, A.; Wang, J.; Hu, Q.; Li, S.; Chen, Y.; Wu, Z.; Zhang, W. Discovery of acylated isoquercitrin derivatives as potent anti-neuroinflammatory agents in vitro and in vivo. Chem. Biol. Interact. 2023, 383, 110675. [Google Scholar] [CrossRef]
- Chen, C.; Yi, C.; Chen, C.; Tian, L.; Li, L.; Yang, L.; Li, Z.; He, L. Inhibitory effect of PPARδ agonist GW501516 on proliferation of hypoxia-induced pulmonary arterial smooth muscle cells by regulating the mTOR pathway. Curr. Med. Sci. 2023, 43, 979–987. [Google Scholar] [CrossRef]
- Song, H.; Yao, P.; Zhang, S.; Jia, H.; Yang, Y.; Liu, L. A non-specific lipid transfer protein, NtLTPI.38, positively mediates heat tolerance by regulating photosynthetic ability and antioxidant capacity in tobacco. Plant Physiol. Biochem. 2023, 200, 107791. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Shen, L.; Matsuura, A.; Xiang, L.; Qi, J. Isoquercitrin from Apocynum venetum L. Exerts Antiaging Effects on Yeasts via Stress Resistance Improvement and Mitophagy Induction through the Sch9/Rim15/Msn Signaling Pathway. Antioxidants 2023, 12, 1939. https://doi.org/10.3390/antiox12111939
Liu Y, Shen L, Matsuura A, Xiang L, Qi J. Isoquercitrin from Apocynum venetum L. Exerts Antiaging Effects on Yeasts via Stress Resistance Improvement and Mitophagy Induction through the Sch9/Rim15/Msn Signaling Pathway. Antioxidants. 2023; 12(11):1939. https://doi.org/10.3390/antiox12111939
Chicago/Turabian StyleLiu, Yanan, Le Shen, Akira Matsuura, Lan Xiang, and Jianhua Qi. 2023. "Isoquercitrin from Apocynum venetum L. Exerts Antiaging Effects on Yeasts via Stress Resistance Improvement and Mitophagy Induction through the Sch9/Rim15/Msn Signaling Pathway" Antioxidants 12, no. 11: 1939. https://doi.org/10.3390/antiox12111939