

Multi-Organ Nutrigenomic Effects of Dietary Grapes in a Mouse Model
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Diet
2.3. Tissue Collection
2.4. RNA Extraction and RNA Sequencing
2.5. Pathway and GO Term Enrichment Analyses
2.6. Heat Map Generation
2.7. Statistical Analyses
3. Results and Discussion
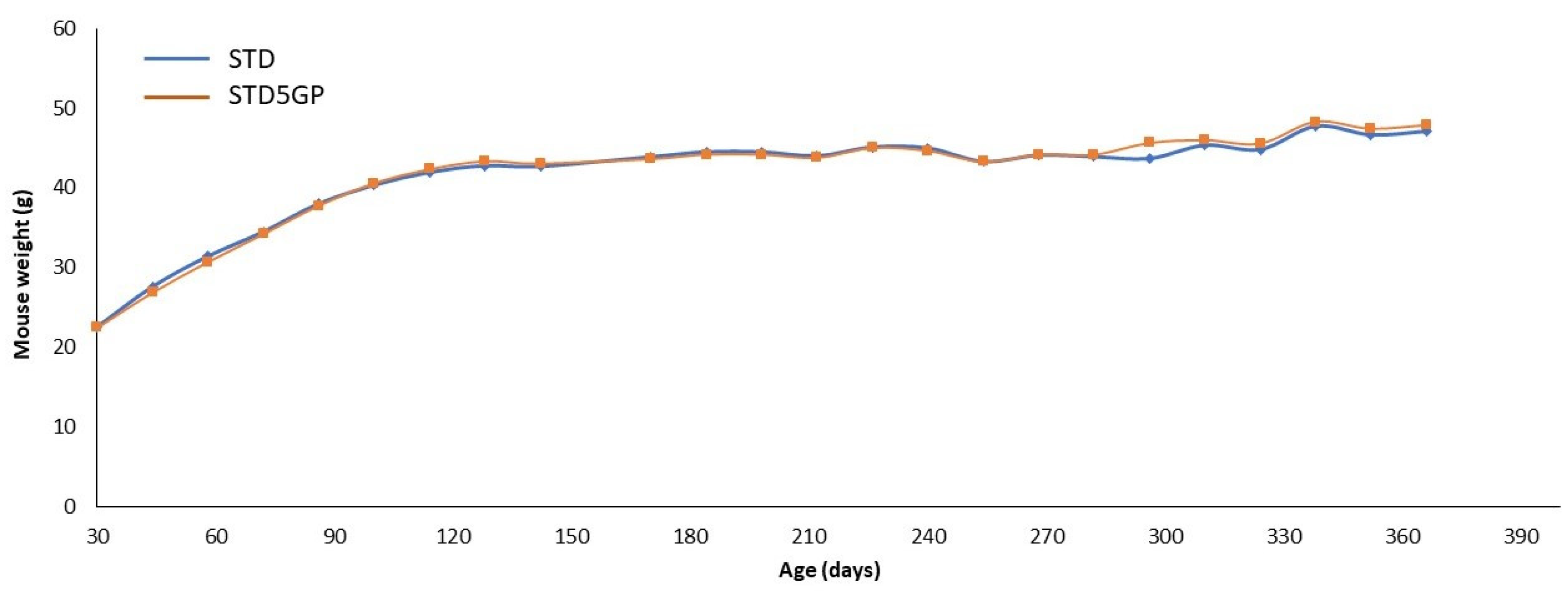
3.1. Body Weight and Dose Selection
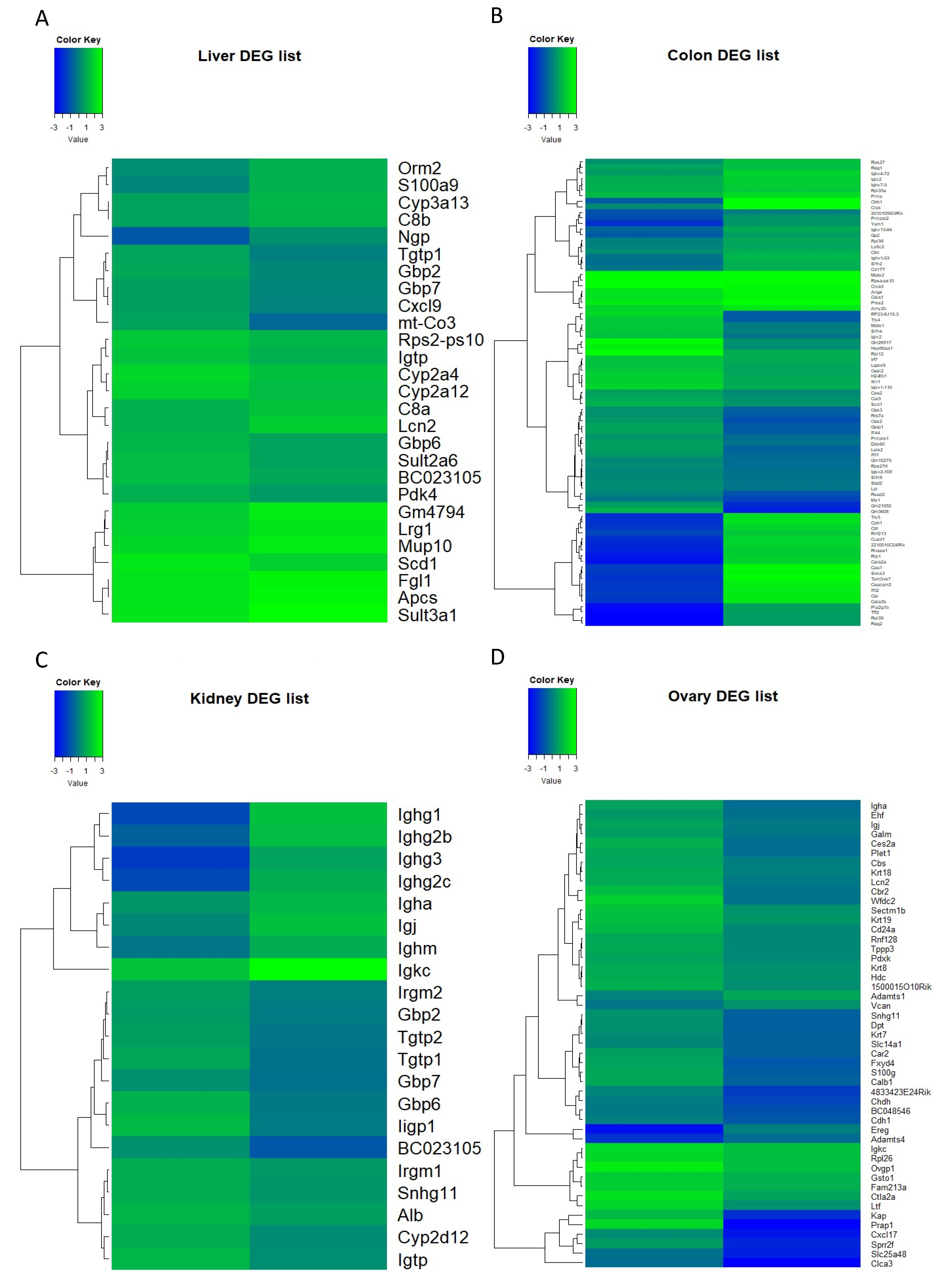
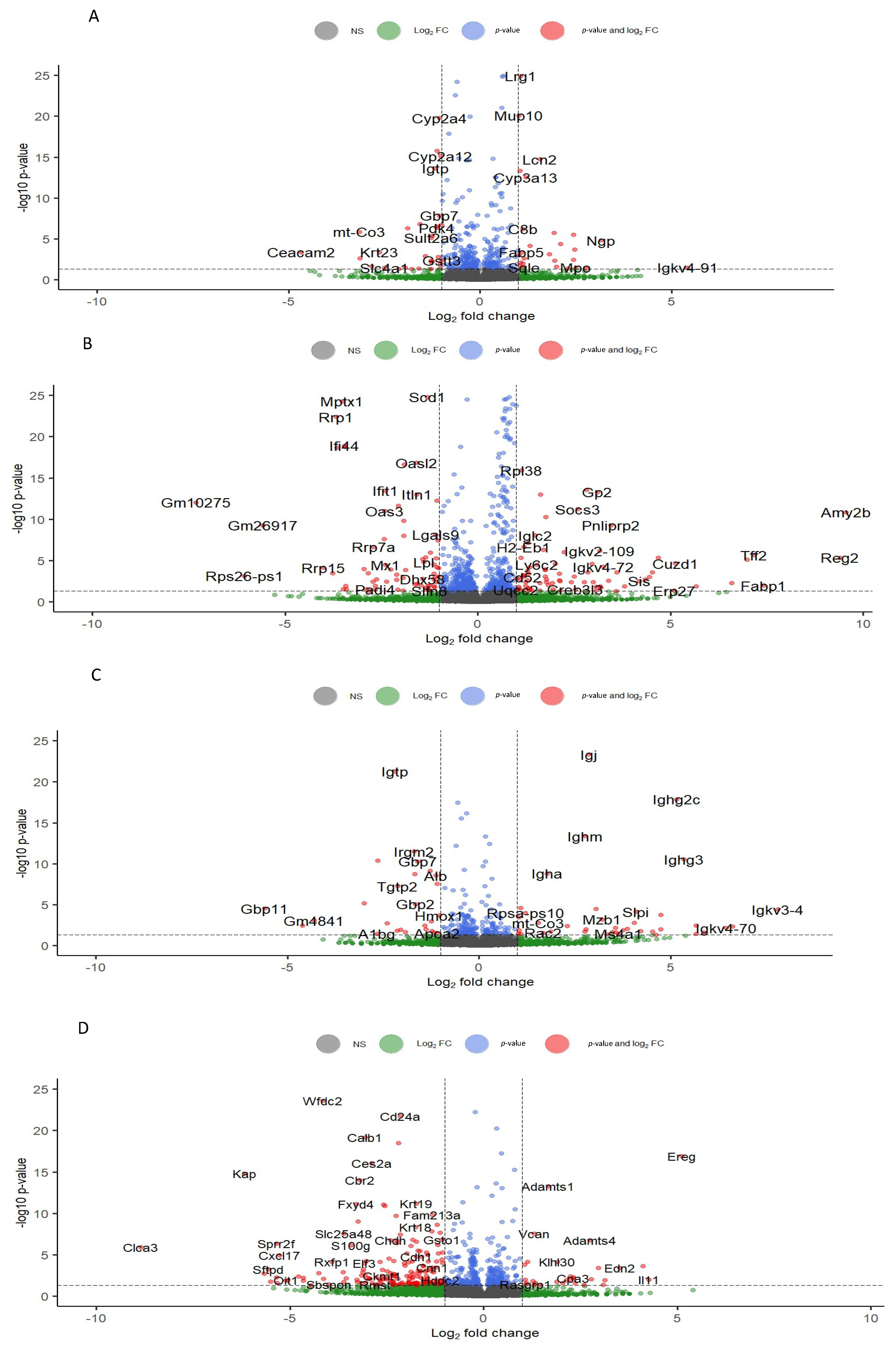
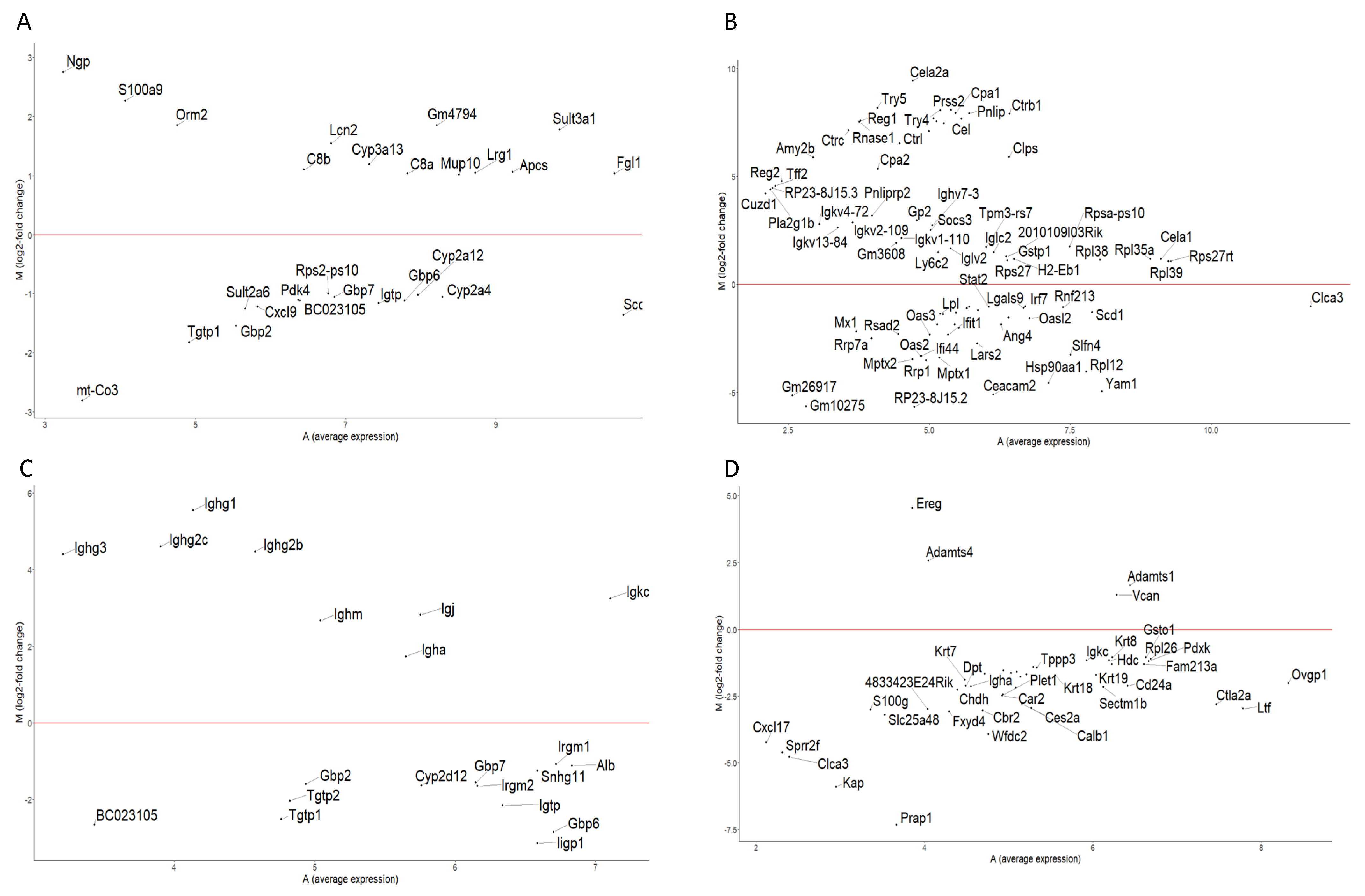
3.2. Differential Gene Expression
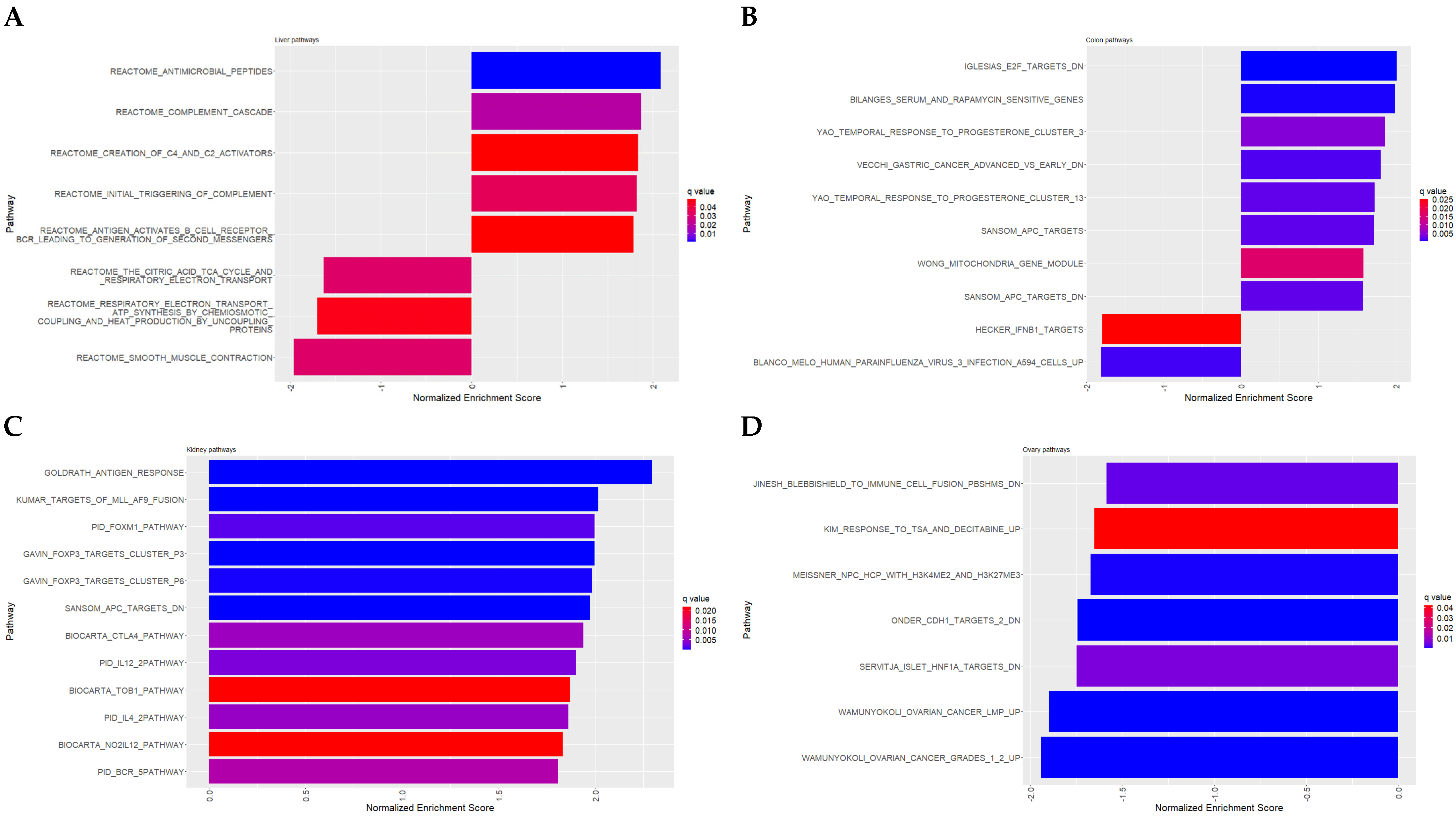
3.3. Pathway Analysis and Gene Enrichment
3.3.1. Liver
- Pathway analysis
- Genes enriched
3.3.2. Colon
- Pathway analysis
- Genes enriched
3.3.3. Kidney
- Pathway analysis
- Genes enriched
3.3.4. Ovary
- Pathway analysis
- Genes enriched
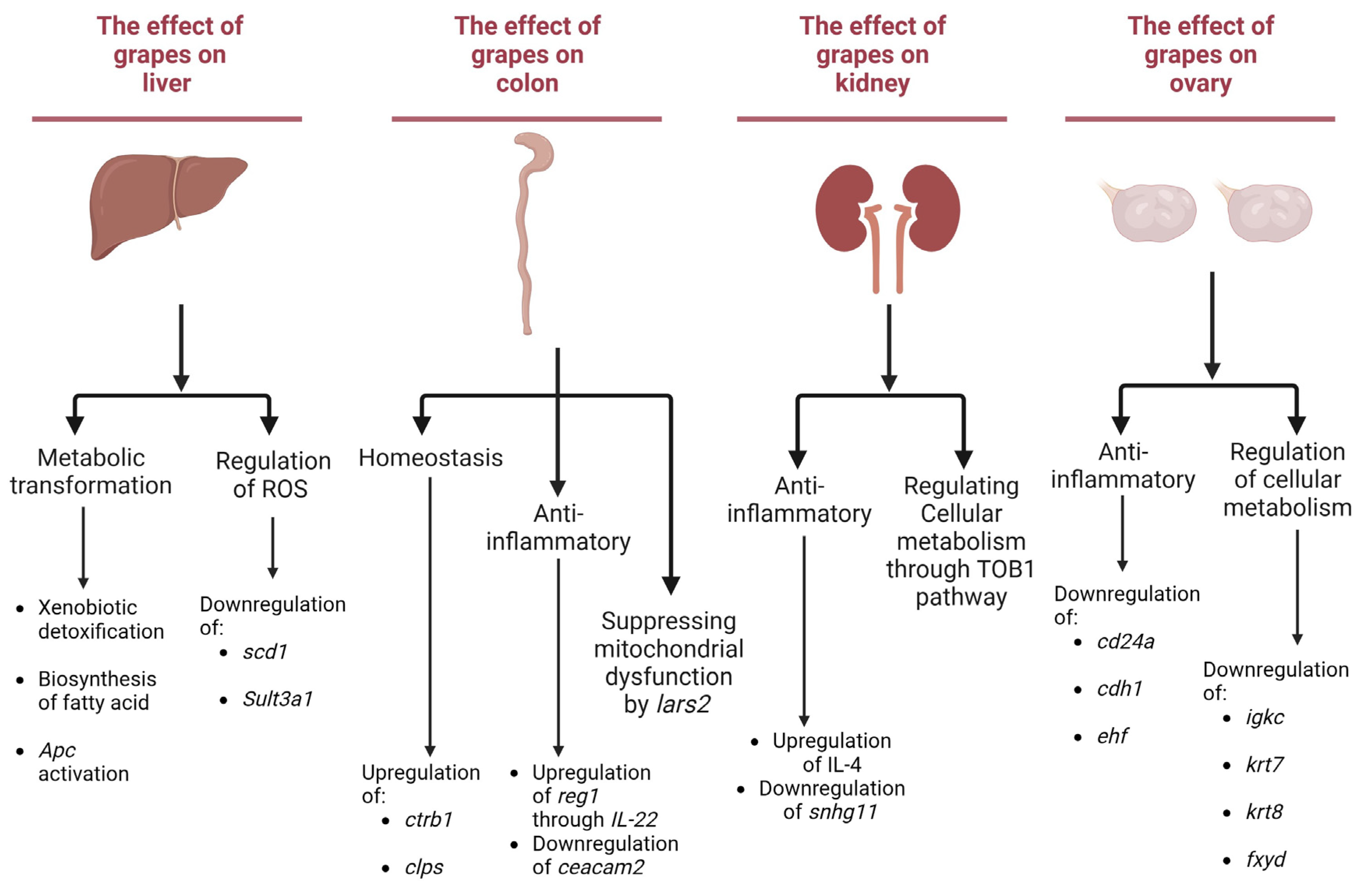
3.4. Potential Influence of Grape Consumption on Metabolic Pathways Associated with Health
3.4.1. Liver-Associated Effects
3.4.2. Colon-Associated Effects
3.4.3. Kidney-Associated Effects
3.4.4. Ovary-Associated Effects
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Pezzuto, J.M. Resveratrol: Twenty Years of Growth, Development and Controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, J.M. Grapes and Human Health: A Perspective. J. Agric. Food Chem. 2008, 56, 6777–6784. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Wright, B.; Li, Y.; Nosal, D.; Burton, T. Standardized Grape Powder for Basic and Clinical Research. In Grapes and Health; Pezzuto, J.M., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 17–26. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Grapes and Health; Springer: Heidelberg, Germany, 2016. [Google Scholar]
- Pezzuto, J.M.; Dave, A.; Park, E.-J.; Beyoğlu, D.; Idle, J.R. Short-Term Grape Consumption Diminishes UV-Induced Skin Erythema. Antioxidants 2022, 11, 2372. [Google Scholar] [CrossRef]
- Dave, A.; Beyoğlu, D.; Park, E.-J.; Idle, J.R.; Pezzuto, J.M. Influence of grape consumption on the human microbiome. Sci. Rep. 2023, 13, 7706. [Google Scholar] [CrossRef] [PubMed]
- Kanner, J.; Frankel, E.; Granit, R.; German, B.; Kinsella, J.E. Natural antioxidants in grapes and wines. J. Agric. Food Chem. 1994, 42, 64–69. [Google Scholar] [CrossRef]
- Li, X.; Yang, X.; Cai, Y.; Qin, H.; Wang, L.; Wang, Y.; Huang, Y.; Wang, X.; Yan, S.; Wang, L.; et al. Proanthocyanidins from Grape Seeds Modulate the NF-κB Signal Transduction Pathways in Rats with TNBS-Induced Ulcerative Colitis. Molecules 2011, 16, 6721–6731. [Google Scholar] [CrossRef]
- Yang, J.; Xiao, Y.-Y. Grape Phytochemicals and Associated Health Benefits. Crit. Rev. Food Sci. Nutr. 2013, 53, 1202–1225. [Google Scholar] [CrossRef]
- Milella, R.A.; Gasparro, M.; Alagna, F.; Cardone, M.F.; Rotunno, S.; Ammollo, C.T.; Semeraro, F.; Tullo, A.; Marzano, F.; Catalano, D.; et al. Gene Expression Signature Induced by Grape Intake in Healthy Subjects Reveals Wide-Spread Beneficial Effects on Peripheral Blood Mononuclear Cells. J. Funct. Foods 2020, 64, 103705. [Google Scholar] [CrossRef]
- Dave, A.; Park, E.-J.; Kumar, A.; Parande, F.; Beyoğlu, D.; Idle, J.R.; Pezzuto, J.M. Consumption of Grapes Modulates Gene Expression, Reduces Non-Alcoholic Fatty Liver Disease, and Extends Longevity in Female C57BL/6J Mice Provided with a High-Fat Western-Pattern Diet. Foods 2022, 11, 1984. [Google Scholar] [CrossRef]
- Parande, F.; Dave, A.; Park, E.-J.; McAllister, C.; Pezzuto, J.M. Effect of Dietary Grapes on Female C57BL6/J Mice Consuming a High-Fat Diet: Behavioral and Genetic Changes. Antioxidants 2022, 11, 414. [Google Scholar] [CrossRef]
- Beyoğlu, D.; Park, E.-J.; Quiñones-Lombraña, A.; Dave, A.; Parande, F.; Pezzuto, J.M.; Idle, J.R. Addition of Grapes to Both a Standard and a High-Fat Western Pattern Diet Modifies Hepatic and Urinary Metabolite Profiles in the Mouse. Food Funct. 2022, 13, 8489–8499. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Dunkelberger, J.R.; Song, W.-C. Complement and its Role in Innate and Adaptive Immune Responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Suiko, M.; Kurogi, K.; Hashiguchi, T.; Sakakibara, Y.; Liu, M.-C. Updated Perspectives on the Cytosolic Sulfotransferases (SULTs) and SULT-mediated Sulfation. Biosci. Biotechnol. Biochem. 2017, 81, 63–72. [Google Scholar] [CrossRef]
- French, J.E.; Gatti, D.M.; Morgan, D.L.; Kissling, G.E.; Shockley, K.R.; Knudsen, G.A.; Shepard, K.G.; Price, H.C.; King, D.; Witt, K.L.; et al. Diversity Outbred Mice Identify Population-Based Exposure Thresholds and Genetic Factors that Influence Benzene-Induced Genotoxicity. Environ. Health Perspect. 2015, 123, 237–245. [Google Scholar] [CrossRef]
- Liu, X.-H.; Qi, L.-W.; Alolga, R.N.; Liu, Q. Implication of the Hepatokine, Fibrinogen-like Protein 1 in Liver Diseases, Metabolic Disorders and Cancer: The Need to Harness its Full Potential. Int. J. Biol. Sci. 2022, 18, 292–300. [Google Scholar] [CrossRef]
- Benhamouche, S.; Decaens, T.; Godard, C.; Chambrey, R.; Rickman, D.S.; Moinard, C.; Vasseur-Cognet, M.; Kuo, C.J.; Kahn, A.; Perret, C.; et al. Apc Tumor Suppressor Gene Is the “Zonation-Keeper” of Mouse Liver. Dev. Cell 2006, 10, 759–770. [Google Scholar] [CrossRef]
- Schartz, N.D.; Tenner, A.J. The Good, the Bad, and the Opportunities of the Complement System in Neurodegenerative Disease. J. Neuroinflamm. 2020, 17, 354. [Google Scholar] [CrossRef]
- Aljohani, A.; Khan, M.I.; Bonneville, A.; Guo, C.; Jeffery, J.; O’Neill, L.; Syed, D.N.; Lewis, S.A.; Burhans, M.; Mukhtar, H.; et al. Hepatic Stearoyl CoA Desaturase 1 Deficiency Increases Glucose Uptake in Adipose Tissue Partially through the PGC-1α–FGF21 Axis in Mice. J. Biol. Chem. 2019, 294, 19475–19485. [Google Scholar] [CrossRef]
- Guerbette, T.; Boudry, G.; Lan, A. Mitochondrial Function in Intestinal Epithelium Homeostasis and Modulation in Diet-Induced Obesity. Mol. Metab. 2022, 63, 101546. [Google Scholar] [CrossRef]
- Németh, B.Z.; Demcsák, A.; Micsonai, A.; Kiss, B.; Schlosser, G.; Geisz, A.; Hegyi, E.; Sahin-Tóth, M.; Pál, G. Arg236 in Human Chymotrypsin B2 (CTRB2) is a Key Determinant of High Enzyme Activity, Trypsinogen Degradation Capacity, and Protection against Pancreatitis. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2022, 1870, 140831. [Google Scholar] [CrossRef] [PubMed]
- Sæle, Ø.; Rød, K.E.L.; Quinlivan, V.H.; Li, S.; Farber, S.A. A Novel System to Quantify Intestinal Lipid Digestion and Transport. Biochim. Biophys. Acta 2018, 1863, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, R.; Contreras, H.; Weidemann, S.; Gorbokon, N.; Menz, A.; Büscheck, F.; Luebke, A.M.; Kluth, M.; Hube-Magg, C.; Hinsch, A.; et al. Carboxypeptidase A1 (CPA1) Immunohistochemistry is Highly Sensitive and Specific for Acinar Cell Carcinoma (ACC) of the Pancreas. Am. J. Surg. Pathol. 2022, 46, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-L.; Lin, X.; Yu, Y.-L.; Chen, L.; Hu, Q.-X.; Chen, M.; Cao, N.; Zhao, C.; Wang, C.-Y.; Huang, C.-W.; et al. Genome-wide Profiling in Colorectal Cancer Identifies PHF19 and TBC1D16 as Oncogenic Super Enhancers. Nat. Commun. 2021, 12, 6407. [Google Scholar] [CrossRef]
- Zhu, G.; Fang, Q.; Zhu, F.; Huang, D.; Yang, C. Structure and Function of Pancreatic Lipase-Related Protein 2 and Its Relationship with Pathological States. Front. Genet. 2021, 12, 693538. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.M.; Guo, X.; Elkahloun, A.G.; Mondal, D.; Bardhan, P.K.; Sugawara, A.; Duggal, P.; Haque, R.; Petri, W.A. The Expression of REG 1A and REG 1B is increased during Acute Amebic Colitis. Parasitol. Int. 2011, 60, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Gonzalo, G.; Hanrahan, N.; Rossini, V.; Singh, R.; Ahern, M.; Kelleher, M.; Hill, S.; O’sullivan, R.; Fanning, A.; Walsh, P.T.; et al. Regulation of CEACAM Family Members by IBD-Associated Triggers in Intestinal Epithelial Cells, Their Correlation to Inflammation and Relevance to IBD Pathogenesis. Front. Immunol. 2021, 12, 655960. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, Z.; Lin, S.; Xia, J.; Zhong, Z.; Xie, Z.; Xing, Y.; Qie, J.; Jiao, M.; Li, Y.; et al. Leucine-tRNA-synthetase-2-expressing B cells Contribute to Colorectal Cancer Immunoevasion. Immunity 2022, 55, 1067–1081.e8. [Google Scholar] [CrossRef]
- Pan, X.; Wang, Q.; Xu, C.; Yan, L.; Pang, S.; Gan, J. Prognostic Value of Chloride Channel Accessory mRNA Expression in Colon Cancer. Oncol. Lett. 2019, 18, 2967–2976. [Google Scholar] [CrossRef]
- Zhang, M.-Z.; Wang, X.; Wang, Y.; Niu, A.; Wang, S.; Zou, C.; Harris, R.C. IL-4/IL-13–Mediated Polarization of Renal Macrophages/Dendritic Cells to an M2a Phenotype is Essential for Recovery from Acute Kidney Injury. Kidney Int. 2017, 91, 375–386. [Google Scholar] [CrossRef]
- Chang-Panesso, M.; Kadyrov, F.F.; Lalli, M.; Wu, H.; Ikeda, S.; Kefaloyianni, E.; Abdelmageed, M.M.; Herrlich, A.; Kobayashi, A.; Humphreys, B.D. FOXM1 Drives Proximal Tubule Proliferation during Repair from Acute Ischemic Kidney Injury. J. Clin. Investig. 2019, 129, 5501–5517. [Google Scholar] [CrossRef]
- Jacobson, N.G.; Szabo, S.J.; Weber-Nordt, R.M.; Zhong, Z.; Schreiber, R.D.; Darnell, J.E., Jr.; Murphy, K.M. Interleukin 12 Signaling in T helper Type 1 (Th1) Cells Involves Tyrosine Phosphorylation of Signal Transducer and Activator of Transcription (Stat)3 and Stat4. J. Exp. Med. 1995, 181, 1755–1762. [Google Scholar] [CrossRef]
- Hamza, T.; Barnett, J.B.; Li, B. Interleukin 12 a Key Immunoregulatory Cytokine in Infection Applications. Int. J. Mol. Sci. 2010, 11, 789–806. [Google Scholar] [CrossRef]
- Lee, H.S.; Kundu, J.; Kim, R.N.; Shin, Y.K. Transducer of ERBB2.1 (TOB1) as a Tumor Suppressor: A Mechanistic Perspective. Int. J. Mol. Sci. 2015, 16, 29815–29828. [Google Scholar] [CrossRef] [PubMed]
- Diefenbach, A.; Schindler, H.; Röllinghoff, M.; Yokoyama, W.M.; Bogdan, C. Requirement for Type 2 NO Synthase for IL-12 Signaling in Innate Immunity. Science 1999, 284, 951–955. [Google Scholar] [CrossRef] [PubMed]
- Grywalska, E.; Smarz-Widelska, I.; Mertowski, S.; Gosik, K.; Mielnik, M.; Podgajna, M.; Abramiuk, M.; Drop, B.; Roliński, J.; Załuska, W. CTLA-4 Expression Inversely Correlates with Kidney Function and Serum Immunoglobulin Concentration in Patients with Primary Glomerulonephritides. Arch. Immunol. Ther. Exp. 2019, 67, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Rudman-Melnick, V.; Adam, M.; Stowers, K.; Potter, A.; Ma, Q.; Chokshi, S.M.; Vanhoutte, D.; Valiente-Alandi, I.; Lindquist, D.M.; Nieman, M.L.; et al. Single-cell Transcriptomic Profiling of Kidney Fibrosis Identifies a Novel Specific Fibroblast Marker and Putative Disease Target. bioRxiv 2022. bioRxiv:2022.09.13.507855. [Google Scholar] [CrossRef]
- Cruz-Rodriguez, N.; Combita, A.L.; Enciso, L.J.; Raney, L.F.; Pinzon, P.L.; Lozano, O.C.; Campos, A.M.; Peñaloza, N.; Solano, J.; Herrera, M.V.; et al. Prognostic Stratification Improvement by Integrating ID1/ID3/IGJ Gene Expression Signature and Immunophenotypic Profile in Adult Patients with B-ALL. J. Exp. Clin. Cancer Res. 2017, 36, 37. [Google Scholar] [CrossRef]
- Woof, J.M.; Kerr, M.A. The Function of Immunoglobulin A in Immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef]
- Bilal, S.; Etayo, A.; Hordvik, I. Immunoglobulins in Teleosts. Immunogenetics 2021, 73, 65–77. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, K.; Li, L.; Ma, K.; Hong, B.; Gong, Y.; Gong, K. Discovery and Validation of the Prognostic Value of the lncRNAs Encoding snoRNAs in Patients with Clear Cell Renal Cell Carcinoma. Aging 2020, 12, 4424–4444. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Hoffman, R.M.; Song, J.; Miao, S.; Zhang, J.; Ding, D.; Wang, D. Guanylate-binding Protein 2 Expression is Associated with Poor Survival and Malignancy in Clear-cell Renal Cell Carcinoma. Anticancer. Res. 2022, 42, 2341–2354. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, R.; Liao, C.; Liu, J.; Zhao, H.; Li, Z.; Liu, W.; Chen, L.; Wu, C.; Tan, H.; et al. High Expression of Immunity-Related GTPase Family M Protein in Glioma Promotes Cell Proliferation and Autophagy Protein Expression. Pathol. Res. Pract. 2019, 215, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Hoter, A.; Naim, H.Y. Heat Shock Proteins and Ovarian Cancer: Important Roles and Therapeutic Opportunities. Cancers 2019, 11, 1389. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Powers, M.A. Nuclear Pore Proteins and Cancer. Semin. Cell Dev. Biol. 2009, 20, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Skapenko, A.; Leipe, J.; Lipsky, P.E.; Schulze-Koops, H. The Role of the T cell in Autoimmune Inflammation. Arthritis Res. Ther. 2005, 7, S4–S14. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Tufekci, K.K.; Uslu, S.; Öner, J. Expression of ADAMTS 1-4-8 and Placental Growth Factor in Ovary and Oviduct during Pregnancy in the First Trimester. Anat. Histol. Embryol. 2023, 52, 619–626. [Google Scholar] [CrossRef]
- Russell, D.L.; Ochsner, S.A.; Hsieh, M.; Mulders, S.; Richards, J.S. Hormone-Regulated Expression and Localization of Versican in the Rodent Ovary. Endocrinology 2003, 144, 1020–1031. [Google Scholar] [CrossRef]
- Maines-Bandiera, S.; Woo, M.M.; Borugian, M.; Molday, L.L.; Hii, T.; Gilks, B.; Leung, P.C.; Molday, R.S.; Auersperg, N. Oviductal Glycoprotein (OVGP1, MUC9): A Differentiation-Based Mucin Present in Serum of Women with Ovarian Cancer. Int. J. Gynecol. Cancer 2010, 20, 16–22. [Google Scholar] [CrossRef]
- Tarhriz, V.; Bandehpour, M.; Dastmalchi, S.; Ouladsahebmadarek, E.; Zarredar, H.; Eyvazi, S. Overview of CD24 as a New Molecular Marker in Ovarian Cancer. J. Cell. Physiol. 2019, 234, 2134–2142. [Google Scholar] [CrossRef]
- MacGregor, H.L.; Garcia-Batres, C.; Sayad, A.; Elia, A.; Berman, H.K.; Toker, A.; Katz, S.R.; Shaw, P.A.; Clarke, B.A.; Crome, S.Q.; et al. Tumor Cell Expression of B7-H4 Correlates with Higher Frequencies of Tumor-Infiltrating APCs and Higher CXCL17 Expression in Human Epithelial Ovarian Cancer. OncoImmunology 2019, 8, e1665460. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, B.; Zhang, Y.-M.; Wang, W. The Association between CDH1 Promoter Methylation and Patients with Ovarian Cancer: A Systematic Meta-analysis. J. Ovarian Res. 2016, 9, 23. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, W.; Cui, S.; Mei, H.; Wu, X.; Zhuo, Z. Establishment of an Ovarian Cancer Omentum Metastasis-related Prognostic Model by Integrated Analysis of scRNA-seq and Bulk RNA-seq. J. Ovarian Res. 2022, 15, 123. [Google Scholar] [CrossRef] [PubMed]
- An, Q.; Liu, T.; Wang, M.; Yang, Y.; Zhang, Z.; Liu, Z.; Yang, B. KRT7 Promotes Epithelial-mesenchymal Transition in Ovarian Cancer via the TGF-β/Smad2/3 Signaling Pathway. Oncol. Rep. 2021, 45, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Brenne, K.; Nymoen, D.A.; Hetland, T.E.; Trope’, C.G.; Davidson, B. Expression of the Ets Transcription Factor EHF in Serous Ovarian Carcinoma Effusions is a Marker of Poor Survival. Hum. Pathol. 2012, 43, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Umezaki, Y.; Ito, M.; Nakashima, M.; Mihara, Y.; Kurohama, H.; Yatsunami, N.; Yasuhi, I. Low Expression of S100P Is Associated with Poor Prognosis in Patients with Clear Cell Adenocarcinoma of the Ovary. Int. J. Gynecol. Cancer 2015, 25, 1582–1586. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, A.L.; et al. Cystathionine Beta-synthase (CBS) Contributes to Advanced Ovarian Cancer Progression and Drug Resistance. PLoS ONE 2013, 8, e79167. [Google Scholar] [CrossRef]
- Hellström, I.; Raycraft, J.; Hayden-Ledbetter, M.; Ledbetter, J.A.; Schummer, M.; McIntosh, M.; Drescher, C.; Urban, N.; Hellström, K.E. The HE4 (WFDC2) Protein is a Biomarker for Ovarian Carcinoma. Cancer Res. 2003, 63, 3695–3700. [Google Scholar] [PubMed]
- Hou, L.; Wang, W.; Zhai, J.; Zhao, H. Circular RNA pyridoxal kinase (circPDXK) involves in the progression of ovarian cancer and glycolysis via regulating miR-654-3p and hexokinase II. Appl. Biol. Chem. 2022, 65, 81. [Google Scholar] [CrossRef]
- Jin, M.; Zhang, H.; Yang, J.; Zheng, Z.; Liu, K. Expression Mode and Prognostic Value of FXYD Family Members in Colon Cancer. Aging 2021, 13, 18404–18422. [Google Scholar] [CrossRef]
- Wu, Y. SNHG11: A New Budding Star in Tumors and Inflammatory Diseases. Mini-Rev. Med. Chem. 2023, 23, 14. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wang, Y.; Liang, M.; Pan, M. CALB1 Enhances the Interaction between p53 and MDM2, and Inhibits the Senescence of Ovarian Cancer Cells. Mol. Med. Rep. 2019, 19, 5097–5104. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, T.; Du, Y.; Qin, L.; Ma, F.; Wu, Z.; Ling, H.; Yang, L.; Wang, Z.; Zhou, Q.; et al. High Fat Diet Significantly Changed the Global Gene Expression Profile Involved in Hepatic Drug Metabolism and Pharmacokinetic System in Mice. Nutr. Metab. 2020, 17, 37. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Z.; Paonessa, J.D.; Meinl, W.; Bhattacharya, A.; Glatt, H.; Vouros, P.; Zhang, Y. Strong Impact of Sulfotransferases on DNA Adduct Formation by 4-Aminobiphenyl in Bladder and Liver in Mice. Cancer Med. 2018, 7, 5604–5610. [Google Scholar] [CrossRef]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D. Loss of Stearoyl–CoA Desaturase-1 Function Protects Mice against Adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [PubMed]
- Sen, U.; Coleman, C.; Sen, T. Stearoyl Coenzyme A Desaturase-1: Multitasker in Cancer, Metabolism, and Ferroptosis. Trends Cancer 2023, 9, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, F.; Yin, H.; Huang, Z.; Lin, Z.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, Present and Future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef]
- Colnot, S.; Decaens, T.; Niwa-Kawakita, M.; Godard, C.; Hamard, G.; Kahn, A.; Giovannini, M.; Perret, C. Liver-targeted Disruption of Apc in Mice Activates β-Catenin Signaling and Leads to Hepatocellular Carcinomas. Proc. Natl. Acad. Sci. USA 2004, 101, 17216–17221. [Google Scholar] [CrossRef]
- Bayrer, J.R.; Wang, H.; Nattiv, R.; Suzawa, M.; Escusa, H.S.; Fletterick, R.J.; Klein, O.D.; Moore, D.D.; Ingraham, H.A. LRH-1 Mitigates Intestinal Inflammatory Disease by Maintaining Epithelial Homeostasis and Cell Survival. Nat. Commun. 2018, 9, 4055. [Google Scholar] [CrossRef]
- Edwards, J.-A.; Tan, N.; Toussaint, N.; Ou, P.; Mueller, C.; Stanek, A.; Zinsou, V.; Roudnitsky, S.; Sagal, M.; Dresner, L.; et al. Role of Regenerating Islet-Derived Proteins in Inflammatory Bowel Disease. World J. Gastroenterol. 2020, 26, 2702–2714. [Google Scholar] [CrossRef]
- Kelleher, M.; Singh, R.; O’driscoll, C.M.; Melgar, S. Carcinoembryonic Antigen (CEACAM) Family Members and Inflammatory Bowel Disease. Cytokine Growth Factor Rev. 2019, 47, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Guan, M.-X. Human Mitochondrial Leucyl-tRNA Synthetase Corrects Mitochondrial Dysfunctions Due to the tRNALeu(UUR) A3243G Mutation, Associated with Mitochondrial Encephalomyopathy, Lactic acidosis, and Stroke-like Symptoms and Diabetes. Mol. Cell. Biol. 2010, 30, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Peruchetti, D.B.; Silva-Filho, J.L.; Silva-Aguiar, R.P.; Teixeira, D.E.; Takiya, C.M.; Souza, M.C.; Henriques, M.d.G.; Pinheiro, A.A.S.; Caruso-Neves, C. IL-4 Receptor α Chain Protects the Kidney Against Tubule-Interstitial Injury Induced by Albumin Overload. Front. Physiol. 2020, 11, 172. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.-K.; Harris, R.C. Deletion of the Epidermal Growth Factor Receptor in Renal Proximal Tubule Epithelial Cells Delays Recovery from Acute Kidney Injury. Kidney Int. 2012, 82, 45–52. [Google Scholar] [CrossRef]
- Stoll, S.W.; Stuart, P.E.; Swindell, W.R.; Tsoi, L.C.; Li, B.; Gandarillas, A.; Lambert, S.; Johnston, A.; Nair, R.P.; Elder, J.T. The EGF Receptor Ligand Amphiregulin Controls Cell Division via FoxM1. Oncogene 2015, 35, 2075–2086. [Google Scholar] [CrossRef]
- Ye, S.; Li, S.; Qin, L.; Zheng, W.; Liu, B.; Li, X.; Ren, Z.; Zhao, H.; Hu, X.; Ye, N.; et al. GBP2 Promotes Clear Cell Renal Cell Carcinoma Progression through Immune Infiltration and Regulation of PD-L1 Expression via STAT1 Signaling. Oncol. Rep. 2023, 49, 49. [Google Scholar] [CrossRef]
- Xu, L.; Huan, L.; Guo, T.; Wu, Y.; Liu, Y.; Wang, Q.; Huang, S.; Xu, Y.; Liang, L.; He, X. LncRNA SNHG11 Facilitates Tumor Metastasis by Interacting with and Stabilizing HIF-1α. Oncogene 2020, 39, 7005–7018. [Google Scholar] [CrossRef]
- Wu, Q.; Ma, J.; Wei, J.; Meng, W.; Wang, Y.; Shi, M. lncRNA SNHG11 Promotes Gastric Cancer Progression by Activating the Wnt/β-Catenin Pathway and Oncogenic Autophagy. Mol. Ther. 2021, 29, 1258–1278. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.-G. The Interplay between TGF-β Signaling and Cell Metabolism. Front. Cell Dev. Biol. 2022, 10, 846723. [Google Scholar] [CrossRef]
- Barkal, A.A.; Brewer, R.E.; Markovic, M.; Kowarsky, M.; Barkal, S.A.; Zaro, B.W.; Krishnan, V.; Hatakeyama, J.; Dorigo, O.; Barkal, L.J.; et al. CD24 Signalling through Macrophage Siglec-10 is a Target for Cancer Immunotherapy. Nature 2019, 572, 392–396. [Google Scholar] [CrossRef]
- Bai, Y.; Li, L.; Li, J.; Chen, R.; Yu, H.; Sun, H.; Wang, J.; Lu, X. A FXYD5/TGF-β/SMAD Positive Feedback Loop Drives Epithelial-to-mesenchymal Transition and Promotes Tumor Growth and Metastasis in Ovarian Cancer. Int. J. Oncol. 2020, 56, 301–314. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (STD) | Control + 5% Grape Powder (STD5GP) | ||
|---|---|---|---|
| TD.160157 | TD.160158 | ||
| Ingredient | (g/kg) | (g/kg) | |
| 1 | Casein | 195.0 | 192.94 |
| 2 | DL-Methionine | 3.0 | 3.0 |
| 3 | Sucrose | 191.0 | 191.0 |
| 4 | Dextrose, anhydrous | 66.45 | 44.3 |
| 5 | Fructose | 66.45 | 44.3 |
| 6 | Corn starch | 235.03 | 232.88 |
| 7 | Maltodextrin | 100.0 | 100.0 |
| 8 | Anhydrous milkfat | 30.0 | 29.85 |
| 9 | Soybean oil | 10.0 | 10.0 |
| 10 | Cellulose | 50.0 | 50.0 |
| 11 | Mineral mix, AIN-76 | 35.0 | 35.0 |
| 12 | Calcium carbonate | 4.0 | 4.0 |
| 13 | Potassium citrate, monohydrate | 4.03 | 2.69 |
| 14 | Vitamin mix, Teklad | 10.0 | 10.0 |
| 15 | Ethoxyquin, antioxidant | 0.04 | 0.04 |
| 16 | Grape powder, freeze-dried | 0.0 | 50.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dave, A.; Park, E.-J.; Pezzuto, J.M. Multi-Organ Nutrigenomic Effects of Dietary Grapes in a Mouse Model. Antioxidants 2023, 12, 1821. https://doi.org/10.3390/antiox12101821
Dave A, Park E-J, Pezzuto JM. Multi-Organ Nutrigenomic Effects of Dietary Grapes in a Mouse Model. Antioxidants. 2023; 12(10):1821. https://doi.org/10.3390/antiox12101821
Chicago/Turabian StyleDave, Asim, Eun-Jung Park, and John M. Pezzuto. 2023. "Multi-Organ Nutrigenomic Effects of Dietary Grapes in a Mouse Model" Antioxidants 12, no. 10: 1821. https://doi.org/10.3390/antiox12101821