
FaAKR23 Modulates Ascorbic Acid and Anthocyanin Accumulation in Strawberry (Fragaria × ananassa) Fruits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Phylogenetic Analysis and Sequence Alignment
2.3. Protein Subcellular Localization
2.4. Transient Expression of FaAKR23 in Strawberry Fruits
2.5. Anthocyanin and AsA Analysis
2.6. RNA Extraction and qRT-PCR Analysis
2.7. Transcriptome Deep Sequencing (RNA-Seq) Analysis
2.8. Statistical Analysis
3. Results
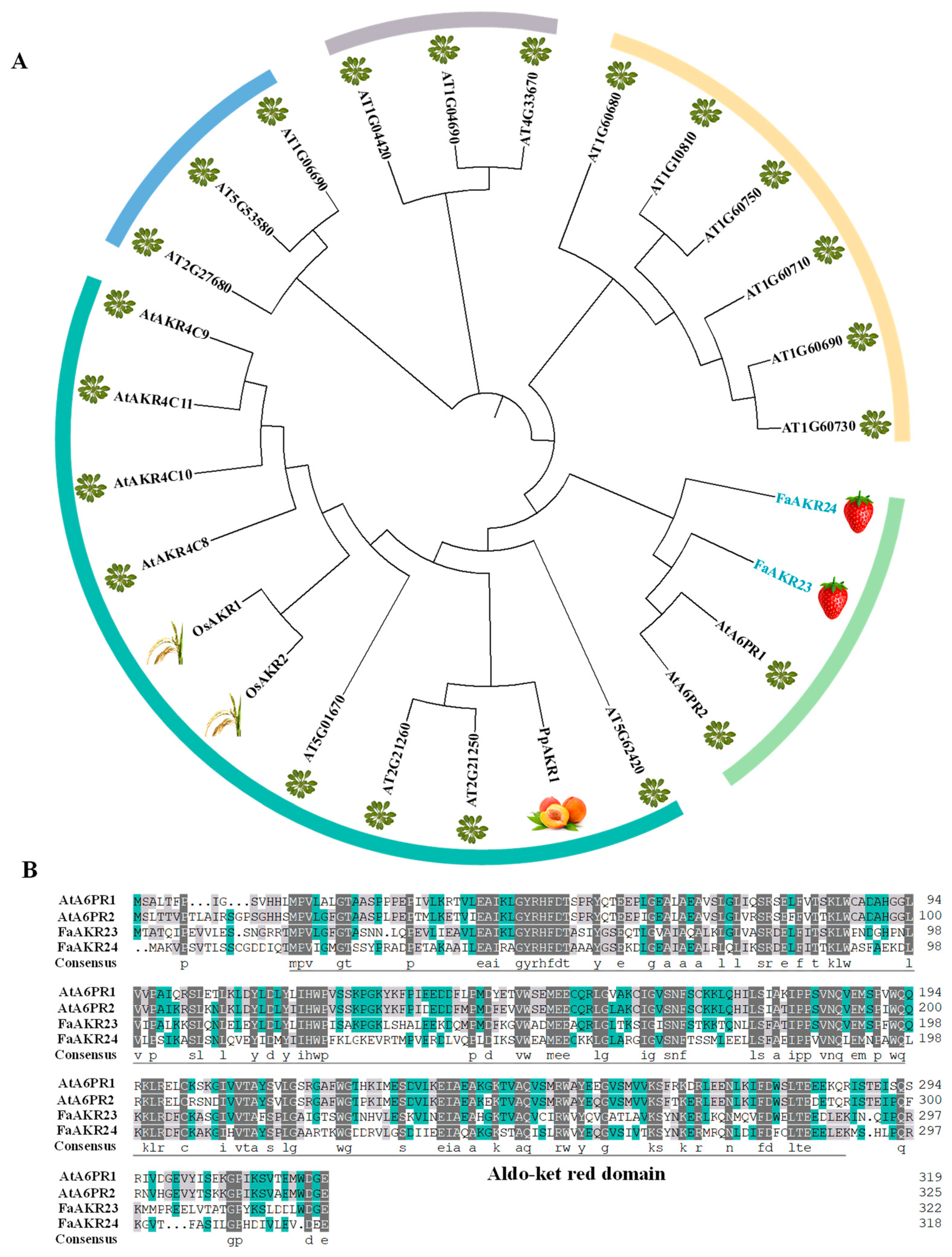
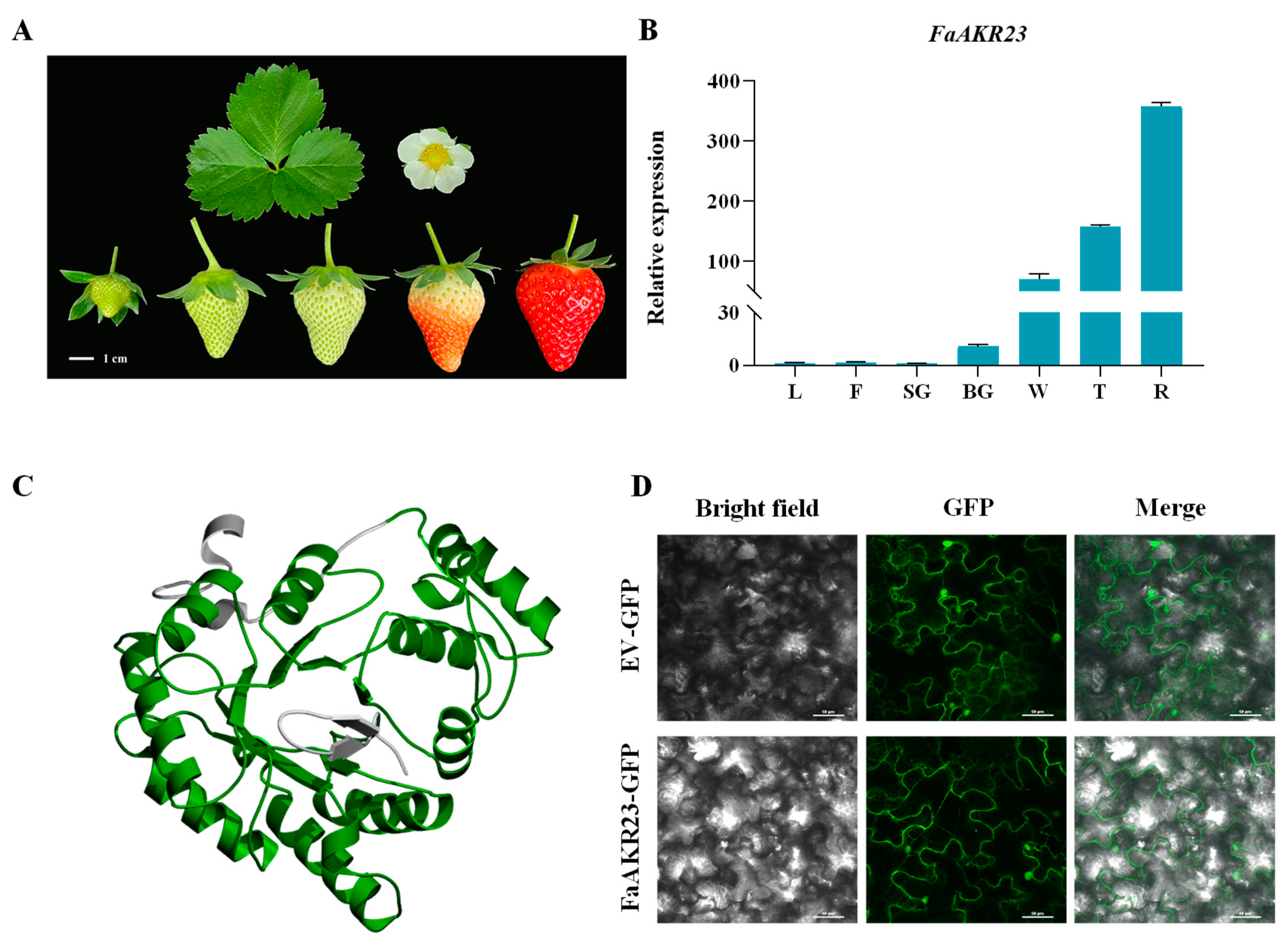
3.1. Identification and Characterization of FaAKR23 in Cultivated Strawberry
3.2. Subcellular Localization of FaAKR23
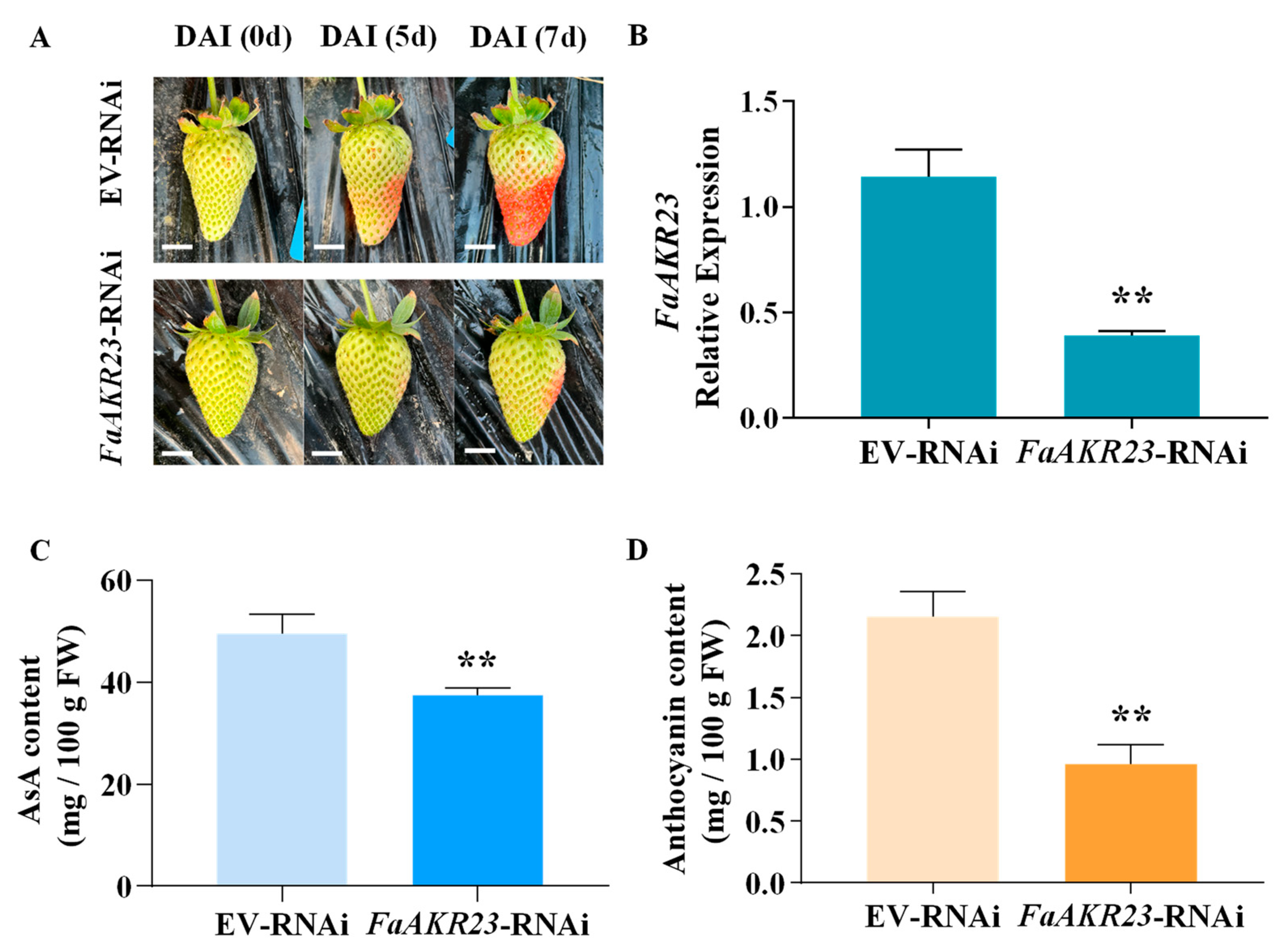
3.3. FaAKR23 Regulates Anthocyanin and AsA Accumulation in Strawberry Fruits
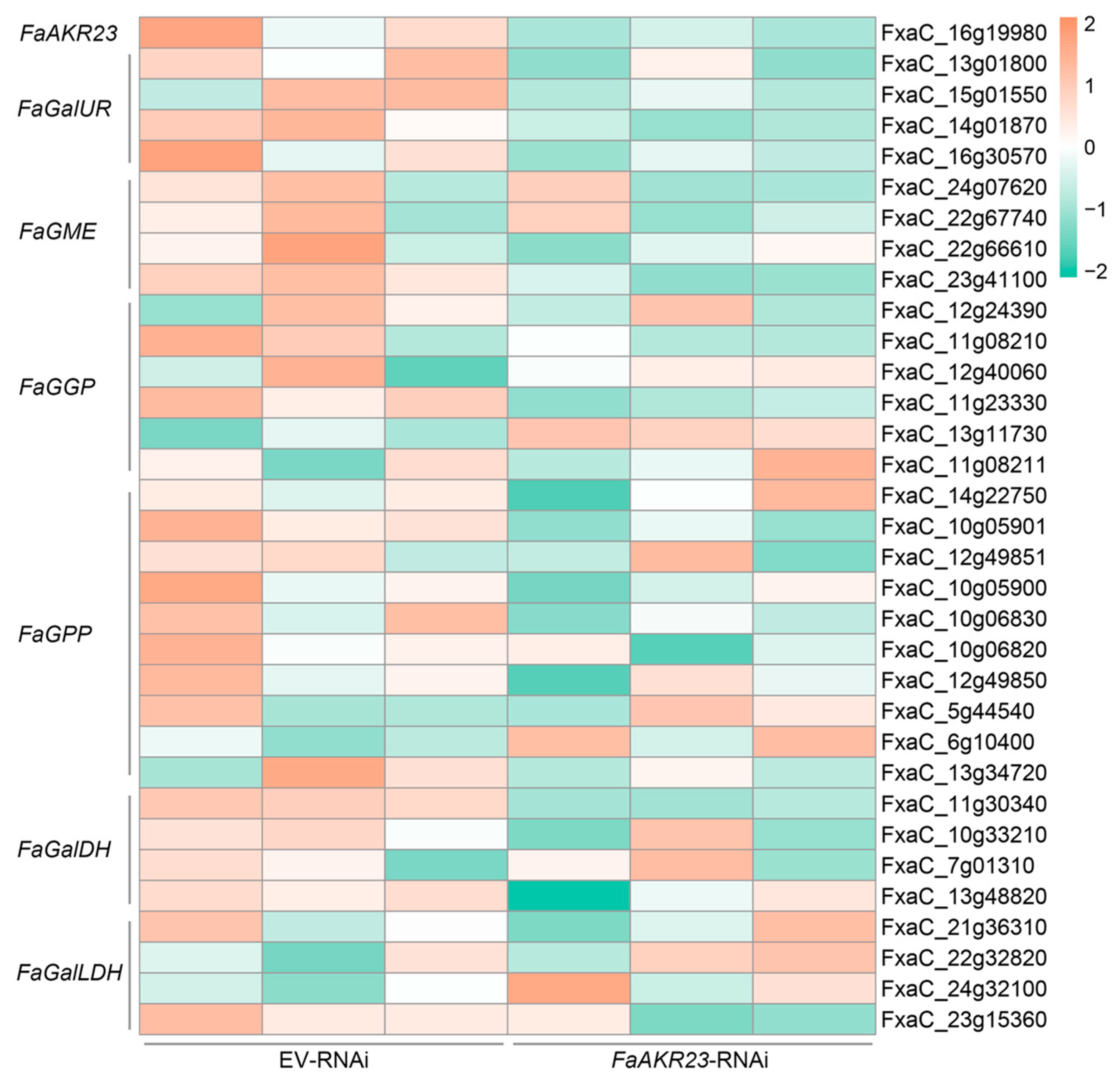
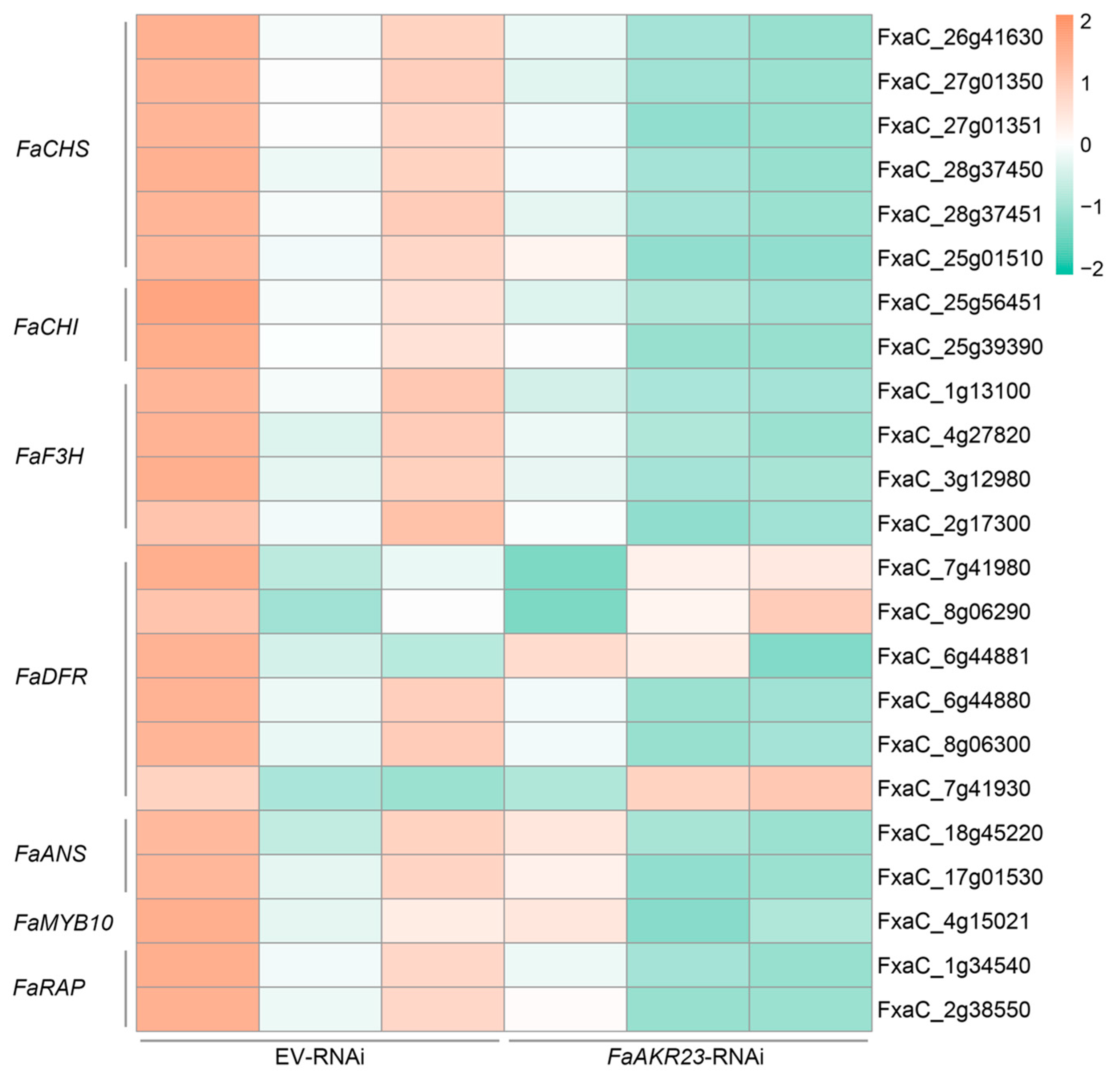
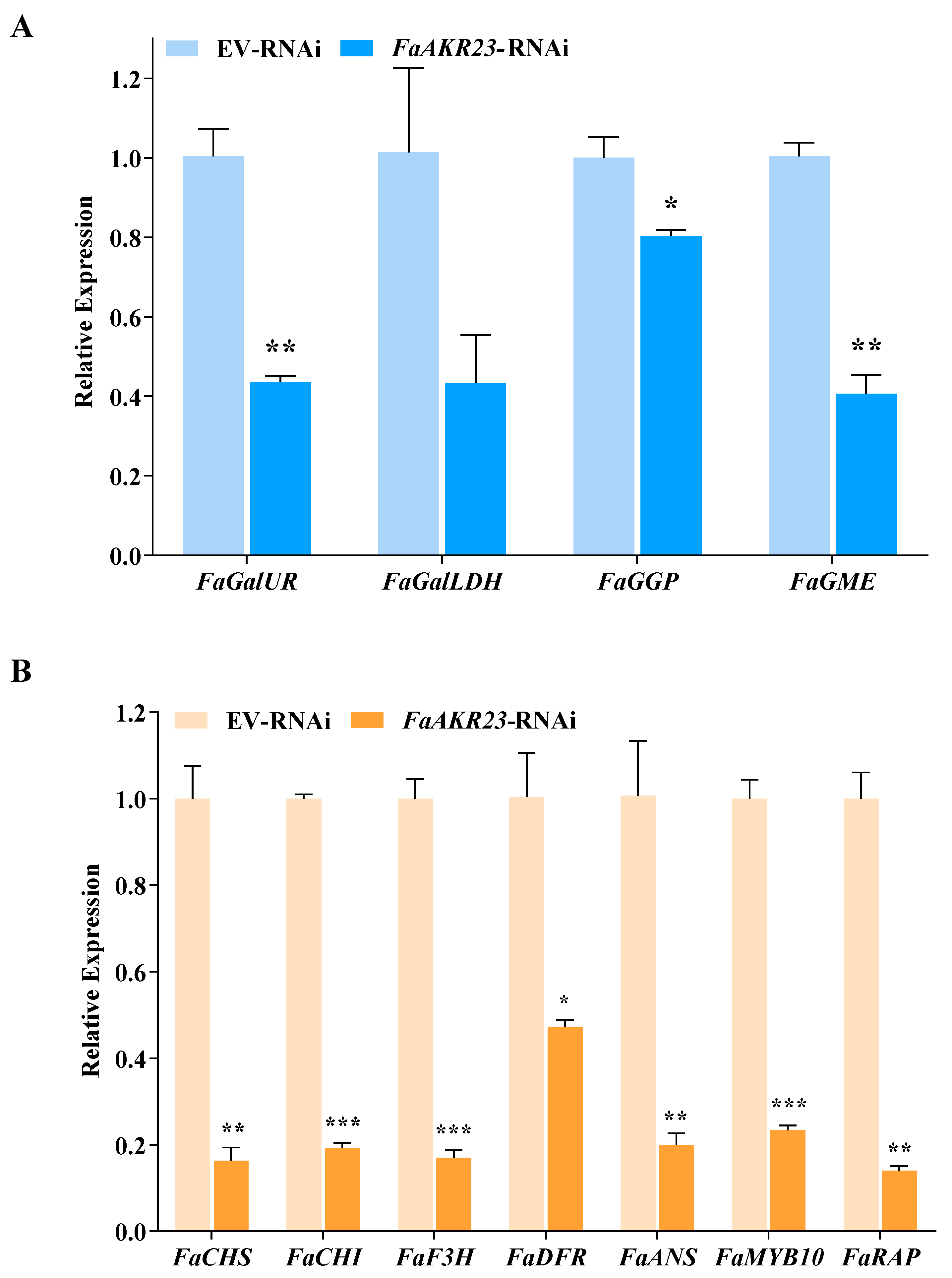
3.4. Expression Patterns of AsA and Anthocyanin Metabolism-Related Genes in EV-RNAi and FaAKR23-RNAi Fruits
3.5. FaAKR23 Involved in Plant–Pathogen Interaction and Starch and Sucrose Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Whitaker, V.M.; Knapp, S.J.; Hardigan, M.A.; Edger, P.P.; Slovin, J.P.; Bassil, N.V.; Hytönen, T.; Mackenzie, K.K.; Lee, S.; Jung, S.; et al. A roadmap for research in octoploid strawberry. Hortic. Res. 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, A.K.; Johnston, C.S. Vitamin C: Overview and update. J. Evid. Based Complementary Altern. Med. 2011, 16, 49–57. [Google Scholar] [CrossRef]
- Wheeler, G.L.; Jones, M.A.; Smirnoff, N. The biosynthetic pathway of vitamin C in higher plants. Nature. 1998, 393, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Dowdle, J.; Ishikawa, T.; Gatzek, S.; Rolinski, S.; Smirnoff, N. Two genes in Arabidopsis thaliana encoding GDP-l-galactose phosphorylase are required for ascorbate biosynthesis and seedling viability. Plant J. 2007, 52, 673–689. [Google Scholar] [CrossRef]
- Ishikawa, T.; Maruta, T.; Yoshimura, K.; Smirnoff, N. Biosynthesis and regulation of ascorbic acid in plants. In Antioxidants and Antioxidant Enzymes in Higher Plants, 2nd ed.; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; Volume 8, pp. 163–179. [Google Scholar]
- Agius, F.; González-Lamothe, R.; Caballero, J.L.; Munoz-Blanco, J.; Botella, M.A.; Valpuesta, V. Engineering increased vitamin C levels in plants by overexpression of a D-galacturonic acid reductase. Nat. Biotechnol. 2003, 21, 177–181. [Google Scholar] [CrossRef]
- Hemavathi; Upadhyaya, C.P.; Young, K.E.; Akula, N.; Kim, H.S.; Heung, J.J.; Oh, O.M.; Aswath, C.R.; Chun, S.C.; Kim, D.H.; et al. Over-expression of strawberry D-galacturonic acid reductase in potato leads to accumulation of vitamin C with enhanced abiotic stress tolerance. Plant Sci. 2009, 177, 659–667. [Google Scholar] [CrossRef]
- Cai, X.F.; Zhang, C.J.; Ye, J.; Hu, T.X.; Ye, Z.B.; Li, H.X.; Zhang, Y.Y. Ectopic expression of FaGalUR leads to ascorbate accumulation with enhanced oxidative stress, cold, and salt tolerance in tomato. Plant Growth Regul. 2015, 76, 187–197. [Google Scholar] [CrossRef]
- Broad, R.; Bonneau, J.P.; Hellens, R.P.; Johnson, A.A. Manipulation of ascorbate biosynthetic, recycling, and regulatory pathways for improved abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 1790. [Google Scholar] [CrossRef]
- Cruz-Rus, E.; Amaya, I.; Sánchez-Sevilla, J.F.; Botella, M.A.; Valpuesta, V. Regulation of L-ascorbic acid content in strawberry fruits. J. Exp. Bot. 2011, 62, 4191–4201. [Google Scholar] [CrossRef]
- Chen, J.X.; Mao, L.C.; Mi, H.B.; Lu, W.J.; Ying, T.J.; Luo, Z.S. Involvement of abscisic acid in postharvest water-deficit stress associated with the accumulation of anthocyanins in strawberry fruit. Postharvest Biol. Technol. 2016, 111, 99–105. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Yang, Y.; Li, M.F.; Zhang, Y.T.; Liu, J.S.; Dong, J.; Li, J.; Butelli, E.; Xue, Z.; et al. The control of red colour by a family of MYB transcription factors in octoploid strawberry (Fragaria × ananassa) fruits. Plant Biotechnol. J. 2020, 18, 1169–1184. [Google Scholar] [CrossRef]
- Schaart, J.G.; Dubos, C.; Fuente, I.R.D.L.; Houwelingen, A.M.M.L.V.; Vos, R.C.H.D.; Jonker, H.H.; Xu, W.J.; Routaboul, J.M.; Lepiniec, L.; Bovy, A.G. Identification and characterization of MYB-bHLH-WD40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (Fragaria × ananassa) fruits. New Phytol. 2013, 197, 454–467. [Google Scholar] [CrossRef]
- Lin-Wang, K.; McGhie, T.K.; Wang, M.; Liu, Y.H.; Warren, B.; Storey, R.; Espley, R.V.; Allan, A.C. Engineering the anthocyanin regulatory complex of strawberry (Fragaria vesca). Front. Plant Sci. 2014, 5, 651. [Google Scholar] [CrossRef] [PubMed]
- Vallarino, J.G.; Osorio, S.; Bombarely, A.; Casañal, A.; Cruz-Rus, E.; Sanchez-Sevilla, J.F.; Amaya, I.; Giavalisco, P.; Fernie, A.R.; Botella, M.A.; et al. Central role of FaGAMYB in transition of strawberry receptacle from development to ripening. New Phytol. 2015, 208, 482–496. [Google Scholar] [CrossRef]
- Lu, W.J.; Chen, J.X.; Ren, X.C.; Yuan, J.J.; Han, X.Y.; Mao, L.C.; Ying, T.J.; Luo, Z.S. One novel strawberry MADS-box transcription factor FaMADS1a acts as a negative regulator in fruit ripening. Sci. Hortic. 2018, 227, 124–131. [Google Scholar] [CrossRef]
- Vallarino, J.G.; Merchante, C.; Sánchez-Sevilla, J.F.; Balaguer, M.A.D.L.; Pott, D.M.; Ariza, M.T.; Casanal, A.; Pose, D.; Vioque, A.; Amaya, I.; et al. Characterizing the involvement of FaMADS9 in the regulation of strawberry fruit receptacle development. Plant Biotechnol. J. 2020, 18, 929–943. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Luo, H.F.; Li, Y.P.; Liu, Z.C.; Kang, C.Y. Genetic modulation of RAP alters fruit coloration in both wild and cultivated strawberry. Plant Biotechnol. J. 2020, 18, 1550–1561. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Shi, Y.N.; Ma, Y.C.; Yang, X.F.; Yin, X.R.; Zhang, Y.Y.; Xiao, Y.W.; Liu, W.L.; Li, Y.D.; Li, S.J.; et al. The strawberry transcription factor FaRAV1 positively regulates anthocyanin accumulation by activation of FaMYB10 and anthocyanin pathway genes. Plant Biotechnol. J. 2020, 18, 2267–2279. [Google Scholar] [PubMed]
- Martín-Pizarro, C.; Vallarino, J.G.; Osorio, S.; Meco, V.; Urrutia, M.; Pillet, J.; Casañal, A.; Merchante, C.; Amaya, I.; Willmitzer, L.; et al. The NAC transcription factor FaRIF controls fruit ripening in strawberry. Plant Cell. 2021, 33, 1574–1593. [Google Scholar] [CrossRef]
- Castillejo, C.; Waurich, V.; Wagner, H.; Ramos, R.; Oiza, N.; Muñoz, P.; Triviño, J.C.; Caruana, J.; Liu, Z.C.; Cobo, N.; et al. Allelic variation of MYB10 is the major force controlling natural variation in skin and flesh color in strawberry (Fragaria spp.) fruit. Plant Cell. 2020, 32, 3723–3749. [Google Scholar] [CrossRef]
- Jia, H.F.; Lu, D.; Sun, J.H.; Li, C.L.; Xing, Y.; Qin, L.; Shen, Y.Y. Type 2C protein phosphatase ABI1 is a negative regulator of strawberry fruit ripening. J. Exp. Bot. 2013, 64, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Dang, R.H.; Li, J.X.; Jiang, J.Z.; Zhang, N.; Jia, M.R.; Wei, L.Z.; Li, Z.Q.; Li, B.B.; Jia, W.S. SUCROSE NONFERMENTING1-RELATED PROTEIN KINASE2.6, an Ortholog of OPEN STOMATA1, is a negative regulator of strawberry fruit development and ripening. Plant Physiol. 2015, 167, 915–930. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.R.; Ding, N.; Zhang, Q.; Xing, S.N.; Wei, L.Z.; Zhao, Y.Y.; Du, P.; Mao, W.W.; Li, J.Z.; Li, B.B.; et al. A FERONIA-like receptor kinase regulates strawberry (Fragaria × ananassa) fruit ripening and quality formation. Front. Plant Sci. 2017, 8, 1099. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.W.; Han, Y.; Chen, Y.T.; Sun, M.Z.; Feng, Q.Q.; Li, L.; Liu, L.P.; Zhang, K.K.; Wei, L.Z.; Han, Z.H.; et al. Low temperature inhibits anthocyanin accumulation in strawberry fruit by activating FvMAPK3-induced phosphorylation of FvMYB10 and degradation of Chalcone Synthase 1. Plant Cell. 2022, 34, 1226–1249. [Google Scholar] [CrossRef]
- Jin, Y.; Penning, T.M. Aldo-keto reductases and bioactivation/detoxication. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 263. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Miyata, S.; Fujii, J.; Inai, Y.; Ueyama, S.; Araki, M.; Soga, T.; Fujinawa, R.; Nishitani, C.; Ariki, S.; et al. In vivo role of aldehyde reductase. Biochim. Biophys. Acta. 2012, 1820, 1787–1796. [Google Scholar] [CrossRef]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef]
- Oberschall, A.; Deak, M.; Torok, K.; Sass, L.; Vass, I.; Kovacs, I.; Feher, A.; Dudits, D.; Horvath, D.V. A novel aldose/aldehyde reductase protects transgenic plants against lipid peroxidation under chemical and drought stresses. Plant J. 2000, 24, 437–446. [Google Scholar] [CrossRef]
- Vemanna, R.S.; Vennapusa, A.R.; Easwaran, M.; Chandrashekar, B.K.; Rao, H.; Ghanti, K.; Sudhakar, C.; Mysore, K.S.; Makarla, U. Aldo-keto reductase enzymes detoxify glyphosate and improve herbicide resistance in plants. Plant Biotechnol. J. 2017, 15, 794–804. [Google Scholar] [CrossRef]
- Vemanna, R.S.; Babitha, K.C.; Solanki, J.K.; Reddy, V.A.; Sarangi, S.K.; Udayakumar, M. Aldo-keto reductase- 1 (AKR1) protect cellular enzymes from salt stress by detoxifying reactive cytotoxic compounds. Plant Physiol. Biochem. 2017, 113, 177–186. [Google Scholar] [CrossRef]
- Kanayama, Y.; Mizutani, R.; Yaguchi, S.; Hojo, A.; Ikeda, H.; Nishiyama, M.; Kanahama, K. Characterization of an uncharacterized aldo-keto reductase gene from peach and its role in abiotic stress tolerance. Phytochemistry. 2014, 104, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.B.; Wei, L.Z.; Ni, Y.; Chang, L.L.; Dong, J.; Zhong, C.F.; Sun, R.; Li, S.T.; Xiong, R.; Wang, G.X.; et al. Genome-wide analysis of ascorbic acid metabolism related genes in Fragaria × ananassa and its expression pattern analysis in strawberry fruits. Front. Plant Sci. 2022, 13, 2314. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Gilot, C.; Persiau, G.; Østergaard, J.; Han, Y.; Bauw, G.C.; Montagu, M.C.V. Ascorbate biosynthesis in Arabidopsis cell suspension culture. Plant Physiol. 1996, 121, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Clancy, M.A.; Rosli, H.G.; Chamala, S.; Barbazuk, W.B.; Civello, P.M.; Folta, K.M. Validation of reference transcripts in strawberry (Fragaria spp.). Mol. Genet. Genomics. 2013, 288, 671–681. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Turoczy, Z.; Kis, P.; Torok, K.; Cserhati, M.; Lendvai, A.; Dudits, D.; Horvath, G.V. Overproduction of a rice aldo-keto reductase increases oxidative and heat stress tolerance by malondialdehyde and methylglyoxal detoxification. Plant Mol. Biol. 2011, 75, 399–412. [Google Scholar] [CrossRef]
- Simpson, P.J.; Tantitadapitak, C.; Reed, A.M.; Mather, O.C.; Bunce, C.M.; White, S.A.; Ride, J.P. Characterization of two novel aldo–keto reductases from Arabidopsis: Expression patterns, broad substrate specificity, and an open active-site structure suggest a role in toxicant metabolism following stress. J. Mol. Biol. 2009, 392, 465–480. [Google Scholar] [CrossRef]
- Rojas, B.; Wurman, J.; Zamudio, M.S.; Donoso, A.; Cabedo, P.; Díaz, F.; Stange, C.; Handford, M. AtA6PR1 and AtA6PR2 encode putative aldose 6-phosphate reductases that are cytosolically localized and respond differentially to cold and salt stress in Arabidopsis thaliana. J. Plant Biochem. Biotechnol. 2019, 28, 114–119. [Google Scholar] [CrossRef]
- Jez, J.M.; Bennett, M.J.; Schlegel, B.P.; Lewis, M.; Penning, T.M. Comparative anatomy of the aldo-keto reductase superfamily. Biochem. J. 1997, 326, 625–636. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Wang, J.; Zhang, R.X.; Huang, R.F. The ethylene response factor AtERF98 enhances tolerance to salt through the transcriptional activation of ascorbic acid synthesis in Arabidopsis. Plant J. 2012, 71, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Kakan, X.; Yu, Y.W.; Li, S.H.; Li, X.Y.; Huang, R.F.; Wang, J. Ascorbic acid modulation by ABI4 transcriptional repression of VTC2 in the salt tolerance of Arabidopsis. BMC Plant Biol. 2021, 21, 112. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Li, W.F.; Ai, G.; Li, C.X.; Liu, G.Z.; Chen, W.F.; Wang, B.; Wang, W.Q.; Lu, Y.E.; Zhang, J.H.; et al. Genome-wide association analysis identifies a natural variation in basic helix-loop-helix transcription factor regulating ascorbate biosynthesis via d-mannose/l-galactose pathway in tomato. PLoS Genet. 2019, 15, e1008149. [Google Scholar] [CrossRef]
- Hu, T.X.; Ye, J.; Tao, P.W.; Li, H.X.; Zhang, J.H.; Zhang, Y.Y.; Ye, Z.B. The tomato HD-Zip I transcription factor SlHZ24 modulates ascorbate accumulation through positive regulation of the d-mannose/l-galactose pathway. Plant J. 2016, 85, 16–29. [Google Scholar] [CrossRef]
- Chen, W.F.; Hu, T.X.; Ye, J.; Wang, B.; Liu, G.Z.; Wang, Y.; Yuan, L.; Li, J.M.; Li, F.M.; Ye, Z.B.; et al. A CCAAT-binding factor, SlNFYA10, negatively regulates ascorbate accumulation by modulating the d-mannose/l-galactose pathway in tomato. Hort. Res. 2020, 7, 200. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shu, P.; Wang, R.C.; Du, X.F.; Xie, Y.; Du, K.; Deng, H.; Li, M.Z.; Zhang, Y.; Grierson, D.; et al. Ethylene response factor AcERF91 affects ascorbate metabolism via regulation of GDP-galactose phosphorylase encoding gene (AcGGP3) in kiwifruit. Plant Sci. 2021, 313, 111063. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Wu, R.M.; Bulley, S.M.; Zhong, C.H.; Li, D.W. Kiwifruit MYBS1-like and GBF3 transcription factors influence L-ascorbic acid biosynthesis by activating transcription of GDP-l-galactose phosphorylase 3. New Phytol. 2022, 234, 1782–1800. [Google Scholar] [CrossRef]
- Bulley, S.; Wright, M.; Rommens, C.; Yan, H.; Rassam, M.; Lin-Wang, K.; Andre, C.; Brewster, D.; Karunairetnam, S.; Allan, A.C.; et al. Enhancing ascorbate in fruits and tubers through over-expression of the l-galactose pathway gene GDP-l-galactose phosphorylase. Plant Biotechnol. J. 2012, 10, 390–397. [Google Scholar] [CrossRef]
- Amaya, I.; Osorio, S.; Martinez-Ferri, E.; Lima-Silva, V.; Doblas, V.G.; Fernández-Muñoz, R.; Fernie, A.R.; Botella, M.A.; Valpuesta, V. Increased antioxidant capacity in tomato by ectopic expression of the strawberry D-galacturonate reductase gene. Biotech. J. 2015, 10, 490–500. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Li, Y.X.; Zhang, Q.; Ren, S.Y.; Shen, Y.Y.; Qin, L.; Xing, Y. Genome-wide analysis of the expression of WRKY family genes in different developmental stages of wild strawberry (Fragaria vesca) fruit. PLoS ONE 2016, 11, e0154312. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Z.; Wang, Z.Q.; Yokosho, K.; Ding, B.; Fan, W.; Gong, Q.Q.; Li, G.X.; Wu, Y.R.; Yang, J.L.; Ma, J.F.; et al. Transcription factor WRKY22 promotes aluminum tolerance via activation of OsFRDL4 expression and enhancement of citrate secretion in rice (Oryza sativa). New Phytol. 2018, 219, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Zhang, D.W.; Lv, K.W.; Zhang, X.M.; Cheng, Z.H.; Li, R.H.; Zhou, B.R.; Jiang, T.B. Functional characterization of poplar WRKY75 in salt and osmotic tolerance. Plant Sci. 2019, 289, 110259. [Google Scholar] [CrossRef] [PubMed]
- Encinas-Villarejo, S.; Maldonado, A.M.; Amil-Ruiz, F.; Santos, B.D.L.; Romero, F.; Pliego-Alfaro, F.; Muñoz-Blanco, J.; Caballero, J.L. Evidence for a positive regulatory role of strawberry (Fragaria x ananassa) FaWRKY1 and Arabidopsis AtWRKY75 proteins in resistance. J. Exp. Bot. 2009, 60, 3043–3065. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Zhao, S.Q.; Gu, S.; Cao, X.Y.; Zhang, Y.; Niu, J.F.; Liu, L.; Li, A.R.; Jia, W.S.; Qi, B.X.; et al. FvWRKY48 binds to the pectate lyase FvPLA promoter to control fruit softening in Fragaria vesca. Plant physiol. 2022, 189, 1037–1049. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, L.; Liu, H.; Ni, Y.; Dong, J.; Zhong, C.; Sun, R.; Li, S.; Xiong, R.; Wang, G.; Sun, J.; et al. FaAKR23 Modulates Ascorbic Acid and Anthocyanin Accumulation in Strawberry (Fragaria × ananassa) Fruits. Antioxidants 2022, 11, 1828. https://doi.org/10.3390/antiox11091828
Wei L, Liu H, Ni Y, Dong J, Zhong C, Sun R, Li S, Xiong R, Wang G, Sun J, et al. FaAKR23 Modulates Ascorbic Acid and Anthocyanin Accumulation in Strawberry (Fragaria × ananassa) Fruits. Antioxidants. 2022; 11(9):1828. https://doi.org/10.3390/antiox11091828
Chicago/Turabian StyleWei, Lingzhi, Huabo Liu, Yang Ni, Jing Dong, Chuanfei Zhong, Rui Sun, Shuangtao Li, Rong Xiong, Guixia Wang, Jian Sun, and et al. 2022. "FaAKR23 Modulates Ascorbic Acid and Anthocyanin Accumulation in Strawberry (Fragaria × ananassa) Fruits" Antioxidants 11, no. 9: 1828. https://doi.org/10.3390/antiox11091828