
Novel Insights into Understanding the Molecular Dialogues between Bipolaroxin and the Gα and Gβ Subunits of the Wheat Heterotrimeric G-Protein during Host–Pathogen Interaction
 ,
,  , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Acquisition and Molecular Modeling of G-Protein Alpha Subunit
2.2. Molecular Docking of ‘Bipolaroxin’
2.3. MD Simulations of G-Alpha and -Beta Subunit-‘Bipolaroxin’ Complex
2.4. Clustering and Principal Component Analysis
2.5. MM/PBSA Binding Free Energy
2.6. In Planta Study
Fungal Strains and Growth Conditions
2.7. Plant Material and Creation of Artificial Epiphytotic Conditions
2.8. Extraction and Purification of Toxins
2.9. Experimental Setup
2.10. Expression Study
2.11. Histopathological Study
2.12. Development of Disease Symptom and Lignin Production
2.13. Statistical Analyses
3. Results
3.1. This Three-Dimensionl Structure of G-Alpha Subunit from Wheat
3.2. Validation of G-Alpha Subunit Modeled Structure
3.3. Docking Analysis of ‘Bipolaroxin’
3.4. MD Simulation
3.5. Essential Dynamics
3.6. Clustering Analysis
3.7. MM/PBSA Binding Free Energy
3.8. Probable Mode of Binding of ‘Bipolaroxin’
3.9. In Planta Assay
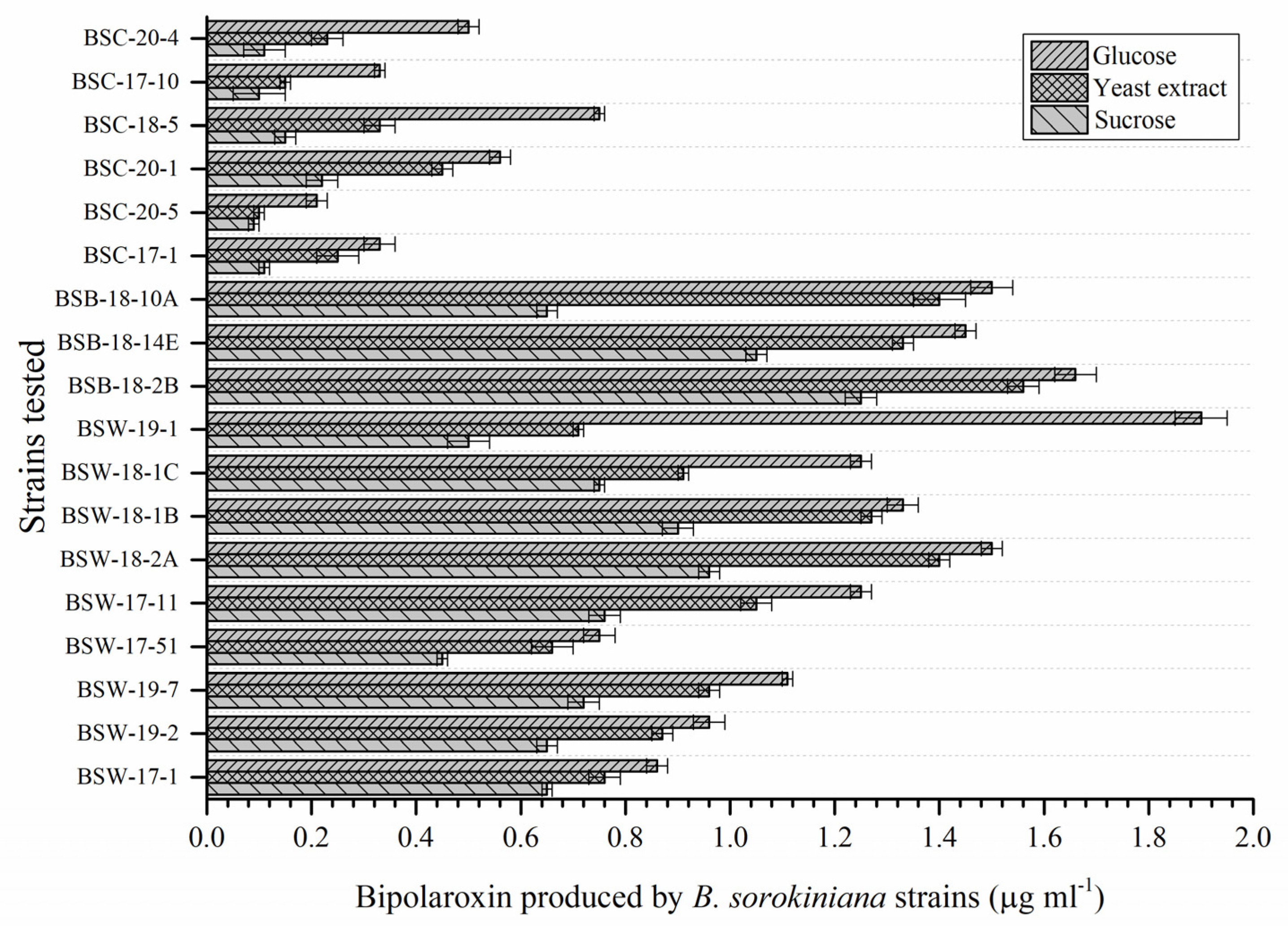
3.10. Selection of Pathogenic Strain
3.11. Expression Profile of Genes Conferring Plant Defense
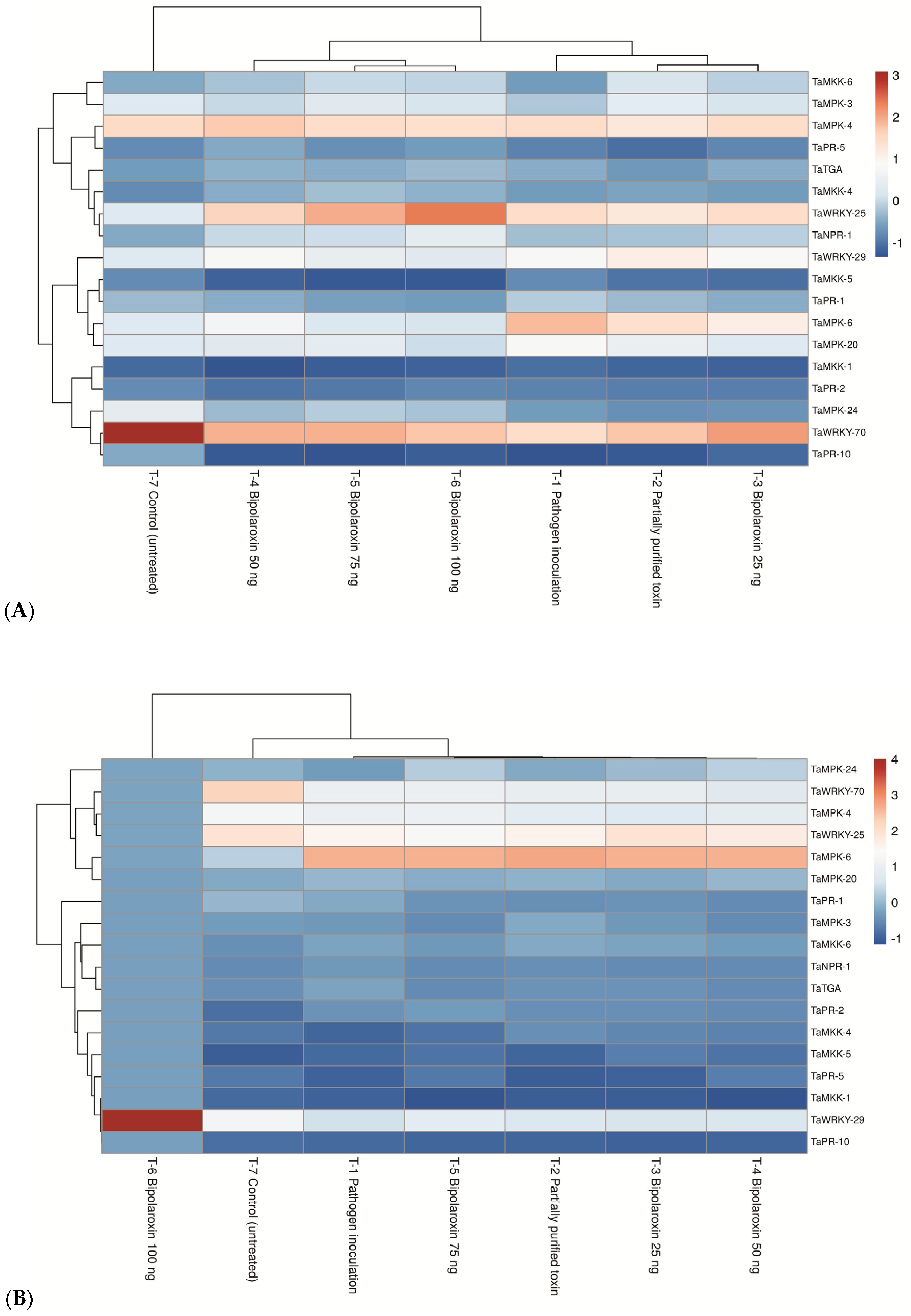
3.11.1. Effects on Expression Profile of Gα- and Gβ Subunits of Heteromeric G Protein
3.11.2. Effects on Expression Profile of Key Genes of Phenylpropanoid Pathway
3.11.3. Effects on Expression Profile of Key Genes of MKK/MAPK Pathways
3.11.4. Effects on Expression Profile of Antioxidant Genes
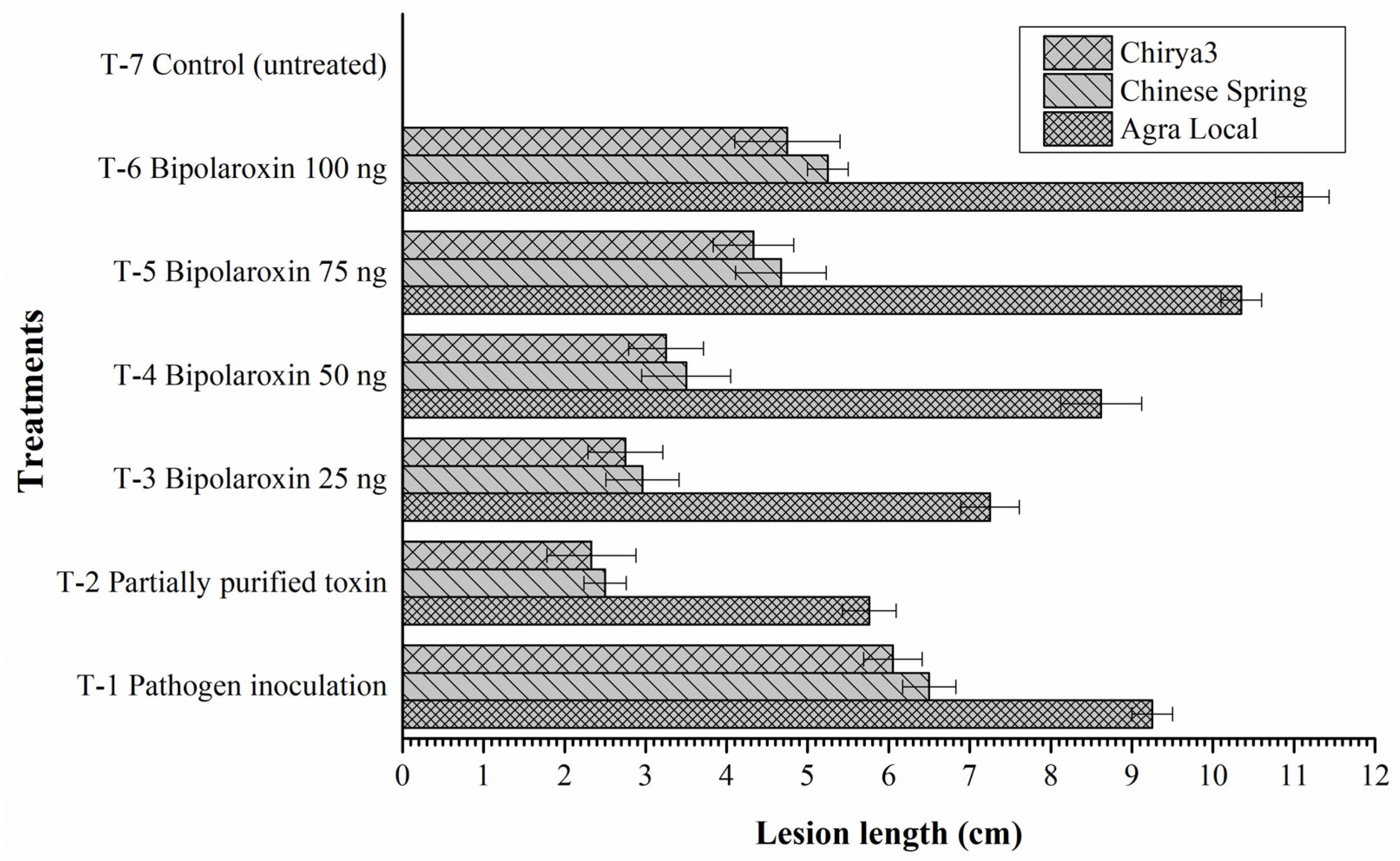
3.11.5. Effects on Symptom Development
3.12. Histopathology
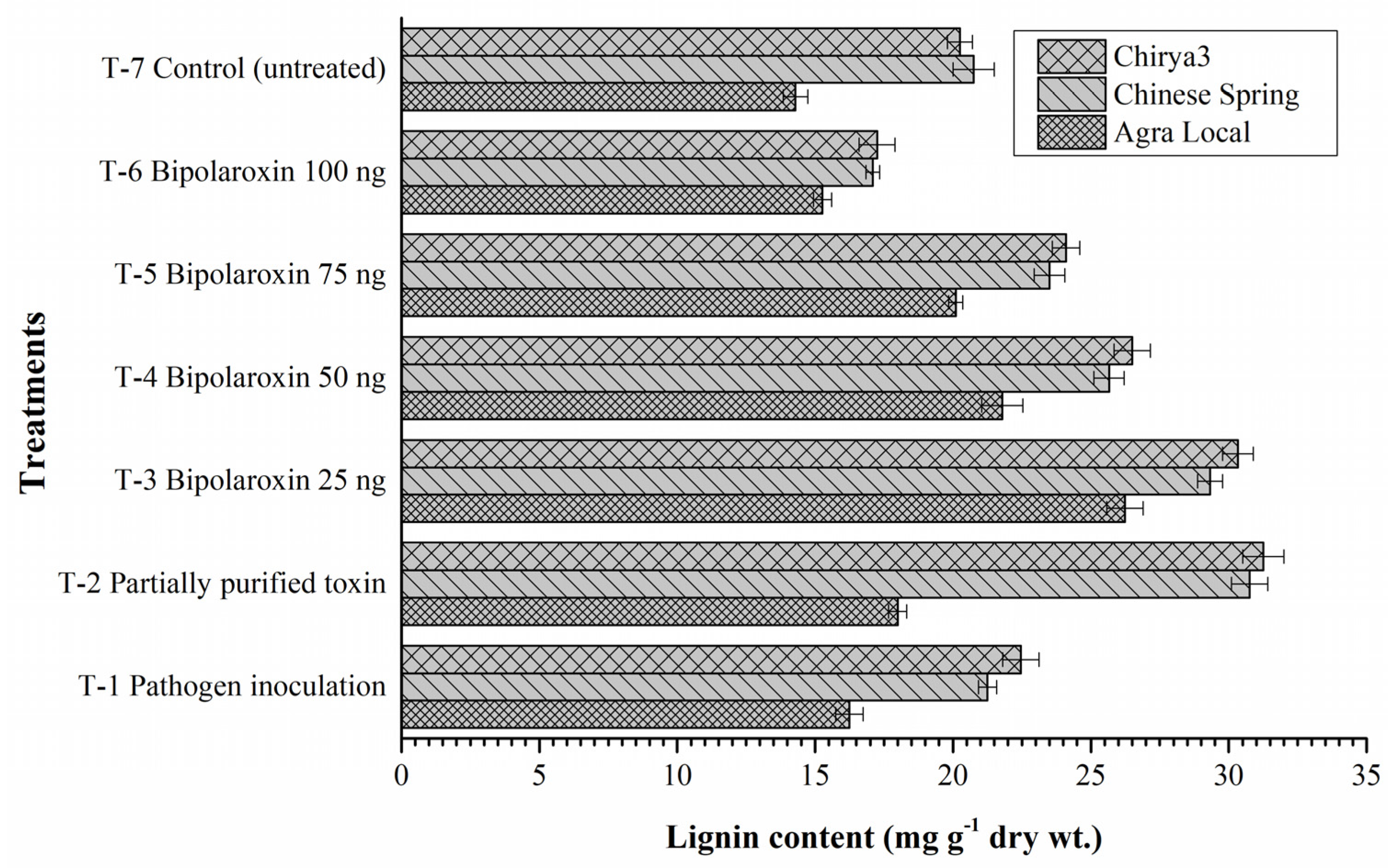
Effects on Lignin Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and methyl jasmonate-induced resistance to Bipolaris sorokiniana through enhanced phenylpropanoid activities in bread wheat (Triticum aestivum L.). Front. Microbiol. 2019, 10, 1697. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.B.; Singh, S.; Malviya, D.; Karthikeyan, N.; Imran, M.; Chaurasia, R.; Alam, M.; Singh, P.; Sarma, B.K.; Rai, J.P.; et al. Integration of anti-penetrant tricyclazole, signaling molecule salicylic acid and root associated Pseudomonas fluorescens enhances suppression of Bipolaris sorokiniana in bread wheat (Triticum aestivum L.). J. Plant Pathol. 2019, 101, 943–954. [Google Scholar] [CrossRef]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ortiz-Ferrara, G.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Kulkarni, G.S. Report of the work done in plant pathology section during the year 1922–23. Ann. Rept. Deptt. Agric. Bombay Pres. 1924, 23, 167–171. [Google Scholar]
- Van-Ginkel, M.; Rajaram, S. Breeding for resistance to spot blotch in wheat: Global perspective. In Helminthosporium Blights of Wheat: Spot Blotch and Tan Spot; Duveiller, E., Dubin, H.J., Reeves, J., McNab, A., Eds.; CIMMYT: Mexico City, Mexico, 1998; pp. 162–170. [Google Scholar]
- Dubin, H.J.; van-Ginkel, M. The status of wheat diseases and disease research in the warmer areas. In Wheat for the Nontraditional Warm Areas: A Proceedings of the International Conference July 29–August 3 1990, Foz do Iguaçu, Brazil; Saunder, D.A., Hettel, G.P., Eds.; CIMMYT: Foz do Iguaçu, Brazil, 1991. [Google Scholar]
- Jahani, M.; Aggarwal, R.; Gupta, S.; Sharma, S.; Dureja, P. Purification and characterization of a novel toxin from Bipolaris sorokiniana, causing spot blotch of wheat and analysis of variability in the pathogen. Cereal Res. Commun. 2014, 42, 252–261. [Google Scholar] [CrossRef]
- Qader, M.M.; Kumar, N.S.; Jayasinghe, L.; Araya, H.; Fujimoto, Y. Bioactive sesquiterpenes from an endophytic fungus Bipolaris sorokiniana isolated from a popular medicinal plant Costus speciosus. Mycology 2017, 8, 17–20. [Google Scholar] [CrossRef]
- Jahani, M.; Aggarwal, R.; Dureja, P.; Srivastava, K.D. Toxin production by Bipolaris sorokiniana and its role in determining resistance in wheat genotypes. Indian Phytopathol. 2006, 59, 340–344. [Google Scholar]
- Trusov, Y.; Jorda, L.; Molina, A.; Botella, J.R. G Proteins and Plant Innate Immunity. In Integrated G-Protein Signaling in Plants; Yalovsky, S., František, B., Alan, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 221–250. [Google Scholar]
- Llorente, F.; Alonso-Blanco, C.; Sanchez-Rodriguez, C.; Jorda, L.; Molina, A. ERECTA receptor-like kinase and heterotrimeric G protein from Arabidopsis are required for resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant J. 2005, 43, 165–180. [Google Scholar] [CrossRef]
- Trusov, Y.; Rookes, J.E.; Chakravorty, D.; Armour, D.; Schenk, P.M.; Botella, J.R. Heterotrimeric G proteins facilitate Arabidopsis resistance to necrotrophic pathogens and are involved in jasmonate signaling. Plant Physiol. 2006, 140, 210–220. [Google Scholar] [CrossRef]
- Trusov, Y.; Rookes, J.E.; Tilbrook, K.; Chakravorty, D.; Mason, M.G.; Anderson, D.; Chen, J.G.; Jones, A.M.; Botella, J.R. Heterotrimeric G Protein gamma subunits provide functional selectivity in Gβγ dimer signaling in Arabidopsis. Plant Cell 2007, 19, 1235–1250. [Google Scholar] [CrossRef]
- Patel, J.S.; Sarma, B.K.; Singh, H.B.; Upadhyay, R.S.; Kharwar, R.N.; Ahmed, M. Pseudomonas fluorescens and Trichoderma asperellum Enhance Expression of Gα Subunits of the Pea Heterotrimeric G-protein during Erysiphe pisi Infection. Front. Plant Sci. 2016, 6, 1206. [Google Scholar] [CrossRef]
- Jones, A.M.; Assmann, S.M. Plants: The latest model system for G-protein research. EMBO Rep. 2004, 5, 572–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H. GTP-binding proteins in plants: New members of an old family. Plant Mol. Biol. 1994, 26, 1611–1636. [Google Scholar] [CrossRef]
- Ma, H.; Yanofsky, M.F.; Meyerowitz, E.M. Molecular cloning and characterization of GPA1, a G protein alpha subunit gene from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1990, 87, 3821–3825. [Google Scholar] [CrossRef] [PubMed]
- Kato, C.; Mizutani, T.; Tamaki, H.; Kumagai, H.; Kamiya, T.; Hirobe, A.; Fujisawa, Y.; Kato, H.; Iwasaki, Y. Characterization of heterotrimeric G protein complexes in rice plasma membrane. Plant J. 2004, 38, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. Heterotrimeric and unconventional GTP binding proteins in plant cell signaling. Plant Cell 2002, 14 (Suppl. S1), S355–S373. [Google Scholar] [CrossRef]
- Perfus-Barbeoch, L.; Jones, A.M.; Assmann, S.M. Plant heterotrimeric G protein function: Insights from Arabidopsis and rice mutants. Curr. Opin. Plant Biol. 2004, 7, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.S.; Selvaraj, V.; Gunupuru, L.R.; Kharwar, R.N.; Sarma, B.K. Plant G-protein signaling cascade and host defense. 3 Biotech 2020, 10, 219. [Google Scholar] [CrossRef]
- Trusov, Y.; Sewelam, N.; Rookes, J.E.; Kunkel, M.; Nowak, E.; Schenk, P.M.; Botella, J. Heterotrimeric G proteins-mediated resistance to necrotrophic pathogens includes mechanisms independent of salicylic acid-, jasmonic acid/ethylene- and abscisic acid-mediated defense signaling. Plant J. 2009, 58, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Suharsono, U.; Fujisawa, Y.; Kawasaki, T.; Iwasaki, Y.; Satoh, H.; Shimamoto, K. The heterotrimeric G protein alpha subunit acts upstream of the small GTPase Rac in disease resistance of rice. Proc. Natl. Acad. Sci. USA 2002, 99, 13307–13312. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Imran, M.; Pathak, N.; Alam, M.; Rai, J.P.; Singh, R.K.; Sarma, B.K.; Sharma, P.K.; et al. Compatible salt-tolerant rhizosphere microbe-mediated induction of phenylpropanoid cascade and induced systemic responses against Bipolaris sorokiniana (Sacc.) Shoemaker causing spot blotch disease in wheat (Triticum aestivum L.). Appl. Soil Ecol. 2016, 108, 300–306. [Google Scholar] [CrossRef]
- Dehury, B.; Sahu, M.; Patra, M.C.; Sarma, K.; Sahu, J.; Sen, P.; Modi, M.K.; Choudhury, M.D.; Barooah, M. Insights into the structure–function relationship of disease resistance protein HCTR in maize (Zea mays L.): A computational structural biology approach. J. Mol. Graph. Model. 2013, 45, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Dehury, B.; Patra, M.C.; Maharana, J.; Sahu, J.; Sen, P.; Modi, M.K.; Choudhury, M.D.; Barooah, M. Structure-based computational study of two disease resistance gene homologues (Hm1 and Hm2) in maize (Zea mays L.) with implications in plant-pathogen interactions. PLoS ONE 2014, 9, e97852. [Google Scholar]
- Kumar, S.; Ahmad, K.; Tandon, G.; Singh, U.B.; Jha, Y.; Nagrale, D.T.; Singh, M.K.; Girdhar, K.; Mondal, P. Novel insight into the molecular interaction of catalase and sucrose: A combination of in silico and in planta assays study. Comput. Electron. Agric. 2018, 151, 258–263. [Google Scholar] [CrossRef]
- Kumar, S.; Ahmad, K.; Behera, S.K.; Nagrale, D.T.; Chaurasia, A.; Yadav, M.K.; Murmu, S.; Jha, Y.; Rajawat, M.V.S.; Malviya, D.; et al. Biocomputational Assessment of Natural Compounds as a Potent Inhibitor to Quorum Sensors in Ralstonia solanacearum. Molecules 2022, 27, 3034. [Google Scholar] [CrossRef] [PubMed]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative protein structure modeling using modeller. Curr. Protoc. Bioinform. 2016, 54, 5–6. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. Galaxy Refine: Protein structure refinement driven by side-chain repacking. Nucl. Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucl. Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Elofsson, A. Can correct protein models be identified? Protein Sci. 2003, 12, 1073–1086. [Google Scholar] [CrossRef]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Du, L. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell. Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Zaman, A.B.; Kamranfar, P.; Domeniconi, C.; Shehu, A. Reducing Ensembles of Protein Tertiary Structures Generated De Novo via Clustering. Molecules 2020, 25, 2228. [Google Scholar] [CrossRef]
- Girdhar, K.; Dehury, B.; Singh, M.K.; Daniel, V.P.; Choubey, A.; Dogra, S.; Mondal, P. Novel insights into the dynamics behavior of glucagon-like peptide-1 receptor with its small molecule agonists. J. Biomol. Struct. Dyn. 2019, 37, 3976–3986. [Google Scholar] [CrossRef]
- Daura, X.; Gademann, K.; Jaun, B.; Seebach, D.; Van Gunsteren, W.F.; Mark, A.E. Peptide folding: When simulation meets experiment. Angew. Chem. Int. Ed. 1999, 38, 236–240. [Google Scholar] [CrossRef]
- Dehury, B.; Behera, S.K.; Mahapatra, N. Structural dynamics of Casein Kinase I (CKI) from malarial parasite Plasmodium falciparum (Isolate 3D7): Insights from theoretical modelling and molecular simulations. J. Mol. Graph. Model. 2017, 71, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Praharaj, A.B.; Dehury, B.; Mahapatra, N.; Kar, S.K.; Behera, S.K. Molecular dynamics insights into the structure, function, and substrate binding mechanism of mucin desulfating sulfatase of gut microbe Bacteroides fragilis. J. Cell Biochem. 2018, 119, 3618–3631. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Chand, R.; Prasad, L.C.; Joshi, A.K. A new technique for monoconidial culture of the most aggressive isolate in a given population of Bipolaris sorokiniana, cause of foliar spot blotch in wheat and barley. World J. Microbiol. Biotechnol. 2007, 23, 1647–1651. [Google Scholar] [CrossRef]
- Bashyal, B.M.; Chand, R.; Kushwaha, C.; Sen, D.; Prasad, L.C.; Joshi, A.K. Association of melanin content with conidiogenesis in Bipolaris sorokiniana of barley (Hordeum vulgare L.). World J. Microbiol. Biotechnol. 2010, 26, 309–316. [Google Scholar] [CrossRef]
- Singh, S.; Singh, U.B.; Trivedi, M.; Malviya, D.; Sahu, P.K.; Roy, M.; Sharma, P.K.; Singh, H.V.; Manna, M.C.; Saxena, A.K. Restructuring the cellular responses: Connecting microbial intervention with ecological fitness and adaptiveness to the maize (Zea mays L.) grown in saline-sodic soil. Front. Microbiol. 2021, 11, 568325. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fredrik, A.; Elisabeth, S. Protocol for Round Robin Test of Lignin Content in Lignin Samples. COST FP0901. Version 3 (2010-12-14). 2011, pp. 1–3. Available online: http://web.abo.fi/fak/tkf/spk/costfp0901/Round_robin/COST_FP0901-Protocol_for_round_robin_test_of_lignin_content-version_3.pdf (accessed on 30 June 2022).
- Urano, D.; Chen, J.G.; Botella, J.R.; Jones, A.M. Heterotrimeric G protein signalling in the plant kingdom. Open Biol. 2013, 3, 120186. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Jones, J.C.; Wang, H.; Matthews, M.; Bradford, W.; Bennetzen, J.L.; Jones, A.M. G protein activation without a GEF in the plant kingdom. PLoS Genet. 2012, 8, e1002756. [Google Scholar] [CrossRef] [PubMed]
- Legendre, L.; Yueh, Y.G.; Crain, R.; Haddock, N.; Heinstein, P.F.; Low, P.S. Phospholipase C activation during elicitation of the oxidative burst in cultured plant cells. J. Biol. Chem. 1993, 268, 24559–24563. [Google Scholar] [CrossRef]
- Torres, M.A.; Morales, J.; Sánchez-Rodríguez, C.; Molina, A.; Dangl, J.L. Functional interplay between Arabidopsis NADPH oxidases and heterotrimeric G protein. Mol. Plant-Microbe. Interact. 2013, 26, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.N.; Kelliher, J.L.; Párraga-Solórzano, P.K.; Kehl-Fie, T.E. The Two-Component System ArlRS and Alterations in Metabolism Enable Staphylococcus aureus to Resist Calprotectin Induced Manganese Starvation. PloS Pathog. 2016, 12, e1006040. [Google Scholar] [CrossRef] [PubMed]
- Sahu, R.; Sharaff, M.; Pradhan, M.; Sethi, A.; Bandyopadhyay, T.; Mishra, V.K.; Pandey, S.P. Elucidation of defense-related signaling responses to spot blotch infection in bread wheat (Triticum aestivum L.). Plant J. 2016, 86, 35–49. [Google Scholar] [CrossRef]
- Yusuf, C.S.; Chand, R.; Mishra, V.K.; Joshi, A.K. The association between leaf malondialdehyde and lignin content and resistance to spot blotch in wheat. J. Phytopathol. 2016, 164, 896–903. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Pradhan, J.K.; Singh, B.P.; Roy, M.; Imram, M.; Pathak, N.; Baisyal, B.M.; Rai, J.P.; et al. Bio-protective microbial agents from rhizosphere eco-systems trigger plant defense responses provide protection against sheath blight disease in rice (Oryza sativa L.). Microbiol. Res. 2016, 192, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Marigoudra, R.M.; Desai, S.A.; Lohithaswa, H.C.; Kalappanavar, I.K.; Hanchinal, R.R. Biochemical and physiological basis of resistance to spot blotch in emmer wheat. Karnataka J. Agric. Sci. 2012, 25, 309–312. [Google Scholar]
- Eisa, M.; Chand, R.; Joshi, A.K. Biochemical and histochemical parameters associated with slow blighting of spot blotch (Bipolaris sorokiniana (Sacc.) Shoem.) in wheat (Triticum spp.). Zemdirbyste Agric. 2013, 100, 191–198. [Google Scholar] [CrossRef]
- Kumar, U.; Joshi, A.K.; Kumar, S.; Chand, R.; Röder, M.S. Quantitative trait loci for resistance to spot blotch caused by Bipolaris sorokiniana in wheat (T. aestivum L.) lines ‘Ning 8201′ and ‘Chirya 3′. Mol. Breed. 2010, 26, 477–491. [Google Scholar] [CrossRef]
- Kumar, U.; Joshi, A.K.; Kumar, S.; Chand, R.; Roder, M.S.; Genet, T.A. Mapping of resistance to spot blotch disease caused by Bipolaris sorokiniana in spring wheat. Theor. Appl. Genet. 2009, 118, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic, a virulent plant symbionts. Nat. Rev. Microbial. 2004, 2, 43. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Ann. Rev. Cell. Dev. Boil. 2012, 28, 489–521. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van-Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Ann. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liao, Y.; Kan, J.; Han, L.; Zheng, Y. Response of direct or priming defense against Botrytis cinerea to methyl jasmonate treatment at different concentrations in grape berries. Int. J. Food Microbiol. 2015, 194, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- McDonald, M.C.; Ahren, D.; Simpfendorfer, S.; Milgate, A.; Solomon, P.S. The discovery of the virulence gene ToxA in the wheat and barley pathogen Bipolaris sorokiniana. Mol. Plant Pathol. 2017, 19, 432–439. [Google Scholar] [CrossRef] [Green Version]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Validation Servers | Model Quality Parameters | Validation Scores |
|---|---|---|
| Procheck (Ramachandran plot) | Most favored regions (%) Additional allowed regions (%) Generously allowed regions (%) Disallowed regions (%) | 94.0 5.1 0.9 0.0 |
| Verify 3D | Averaged 3D-1D score >= 0.2(%) | 86.76 |
| ERRAT | Overall quality (%) | 82.22 |
| ProSA | Z score | −8.05 |
| ProQ | LG score Max Sub | 5.347 0.509 |
| Prove | Z score mean | −0.041 |
| METAMQAP-II | GDT_TS | 51.486 |
| Target | Binding Energy (kcal/mol) | No. of H-bonds | H-bond Forming Residues | Average H-bond Distance (Å) | Hydrophobic Contacts |
|---|---|---|---|---|---|
| G-Alpha subunit | −8.19 | 6 | Glu29, Ser30, Lys32, Ala177 | 2.75 | Gly28, Gly31, Ser33, Thr34, Arg178, Val179, Thr181, Gly209, |
| G-Beta subunit | −7.47 | 7 | Lys256, Phe306, Leu352 | 2.62 | Trp74, Ala305, Phe306, Ile308, Leu350, Gly351 and Ser354 |
| Protein–Ligand System | Van Der Waal Energy (kJ/mol) | Electrostatic Energy (kJ/mol) | Polar Solvation Energy (kJ/mol) | SASA Energy (kJ/mol) | Binding Energy (kJ/mol) |
|---|---|---|---|---|---|
| G-Alpha | −114.91 ± 0.85 | −51.89 ± 1.43 | 143.51 ± 1.72 | −12.25 ± 0.05 | −35.54 ± 1.07 |
| G-Beta | −134.61 ± 0.78 | −73.12 ± 1.65 | 185.54 ± 2.53 | −12.45 ± 0.06 | −34.68 ± 1.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malviya, D.; Singh, U.B.; Dehury, B.; Singh, P.; Kumar, M.; Singh, S.; Chaurasia, A.; Yadav, M.K.; Shankar, R.; Roy, M.; et al. Novel Insights into Understanding the Molecular Dialogues between Bipolaroxin and the Gα and Gβ Subunits of the Wheat Heterotrimeric G-Protein during Host–Pathogen Interaction. Antioxidants 2022, 11, 1754. https://doi.org/10.3390/antiox11091754
Malviya D, Singh UB, Dehury B, Singh P, Kumar M, Singh S, Chaurasia A, Yadav MK, Shankar R, Roy M, et al. Novel Insights into Understanding the Molecular Dialogues between Bipolaroxin and the Gα and Gβ Subunits of the Wheat Heterotrimeric G-Protein during Host–Pathogen Interaction. Antioxidants. 2022; 11(9):1754. https://doi.org/10.3390/antiox11091754
Chicago/Turabian StyleMalviya, Deepti, Udai B. Singh, Budheswar Dehury, Prakash Singh, Manoj Kumar, Shailendra Singh, Anurag Chaurasia, Manoj Kumar Yadav, Raja Shankar, Manish Roy, and et al. 2022. "Novel Insights into Understanding the Molecular Dialogues between Bipolaroxin and the Gα and Gβ Subunits of the Wheat Heterotrimeric G-Protein during Host–Pathogen Interaction" Antioxidants 11, no. 9: 1754. https://doi.org/10.3390/antiox11091754