
Cynanoside F Controls Skin Inflammation by Suppressing Mitogen-Activated Protein Kinase Activation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Western Blot Analysis
2.5. Reporter Gene Assay
2.6. Real-Time Quantitative PCR (qRT-PCR)
2.7. Oxazolone-Induced Atopic Dermatitis (AD) Mouse Model
2.8. Histological Analysis
2.9. ELISA
2.10. Statistical Analysis
3. Results
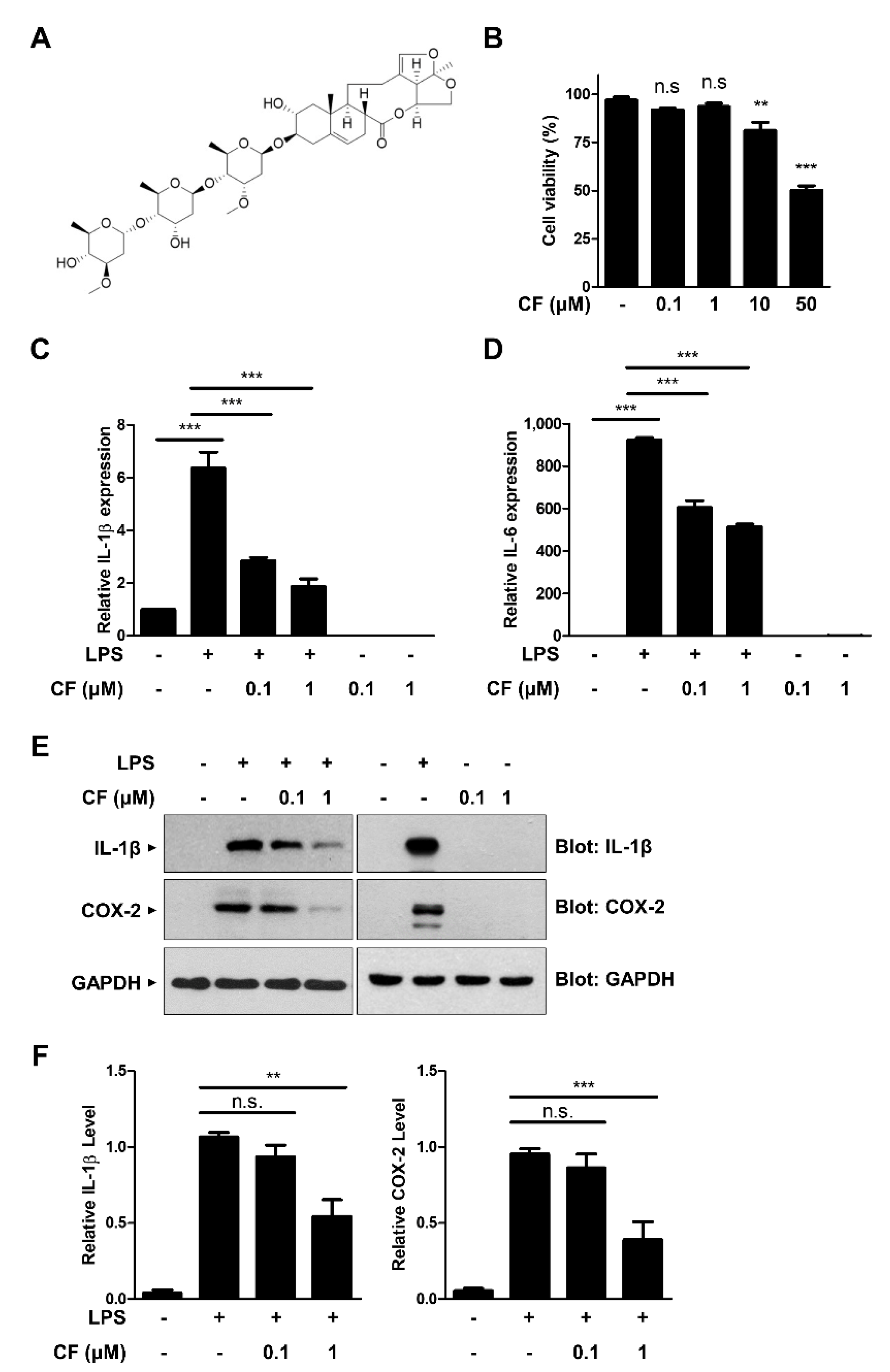
3.1. CF Has an Inhibitory Effect on Proinflammatory Cytokine Expression in RAW264.7 Macrophages
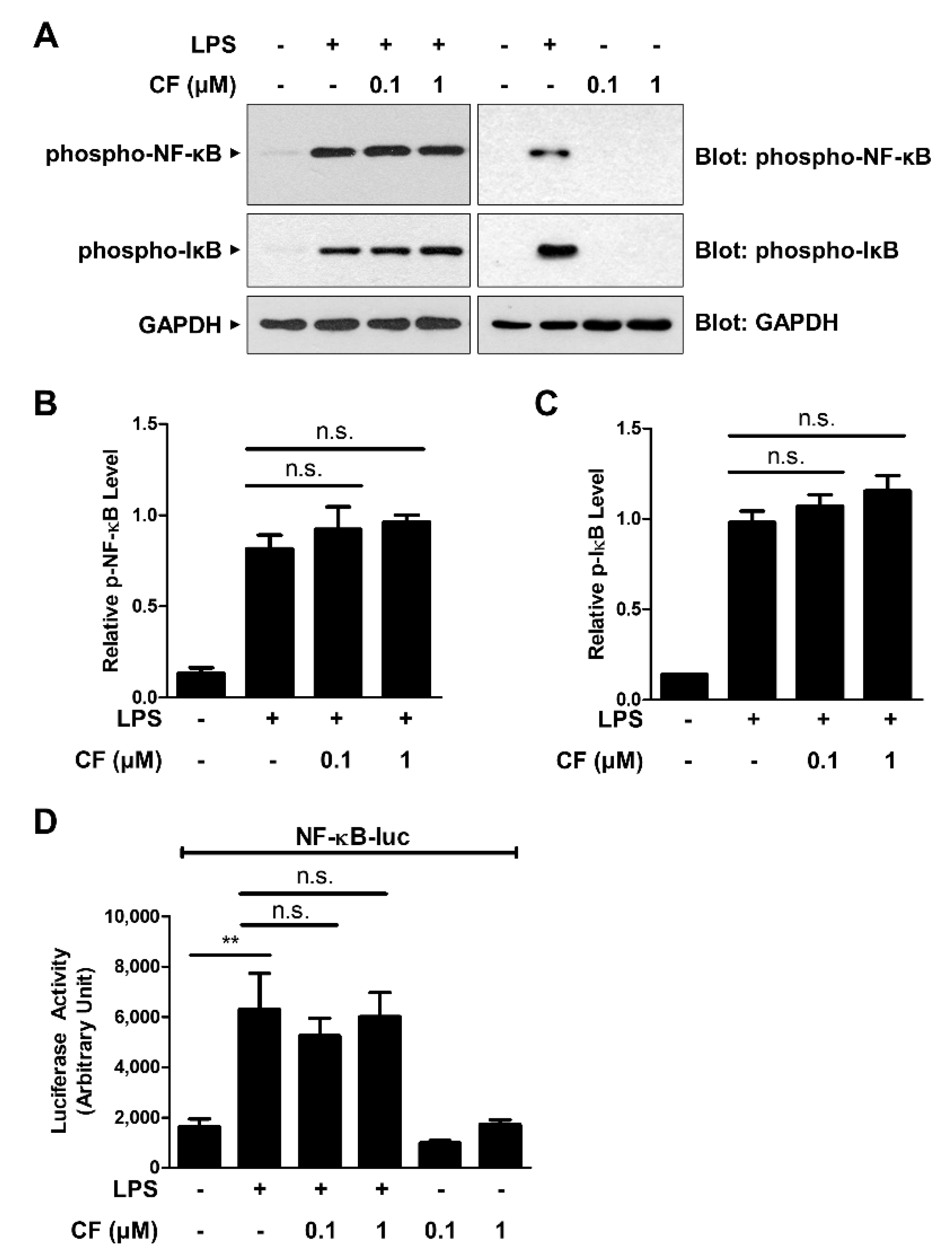
3.2. CF Does Not Inhibit NF-κB Signaling Pathways
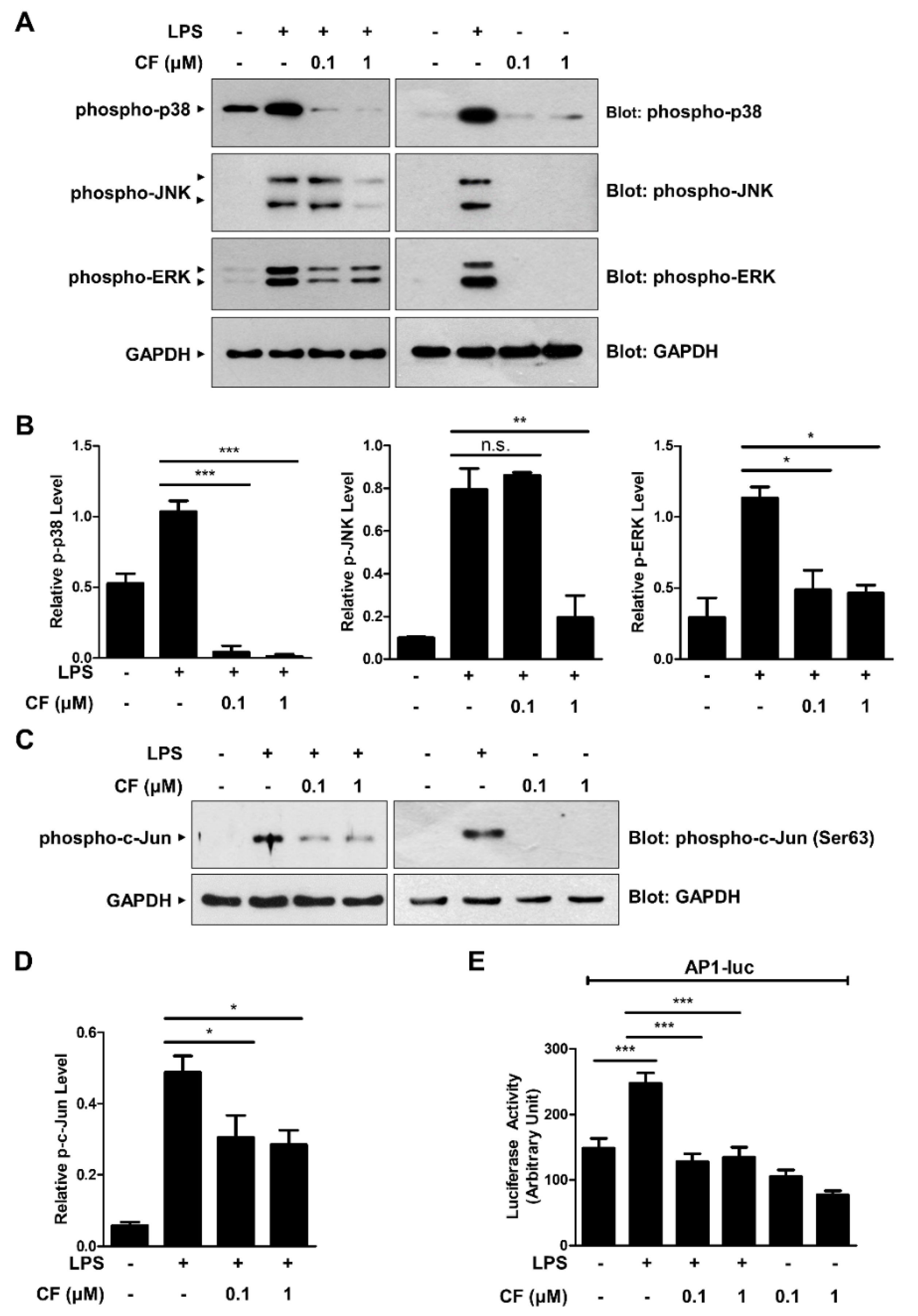
3.3. CF Inhibits the MAPK/AP-1 Signaling Pathway
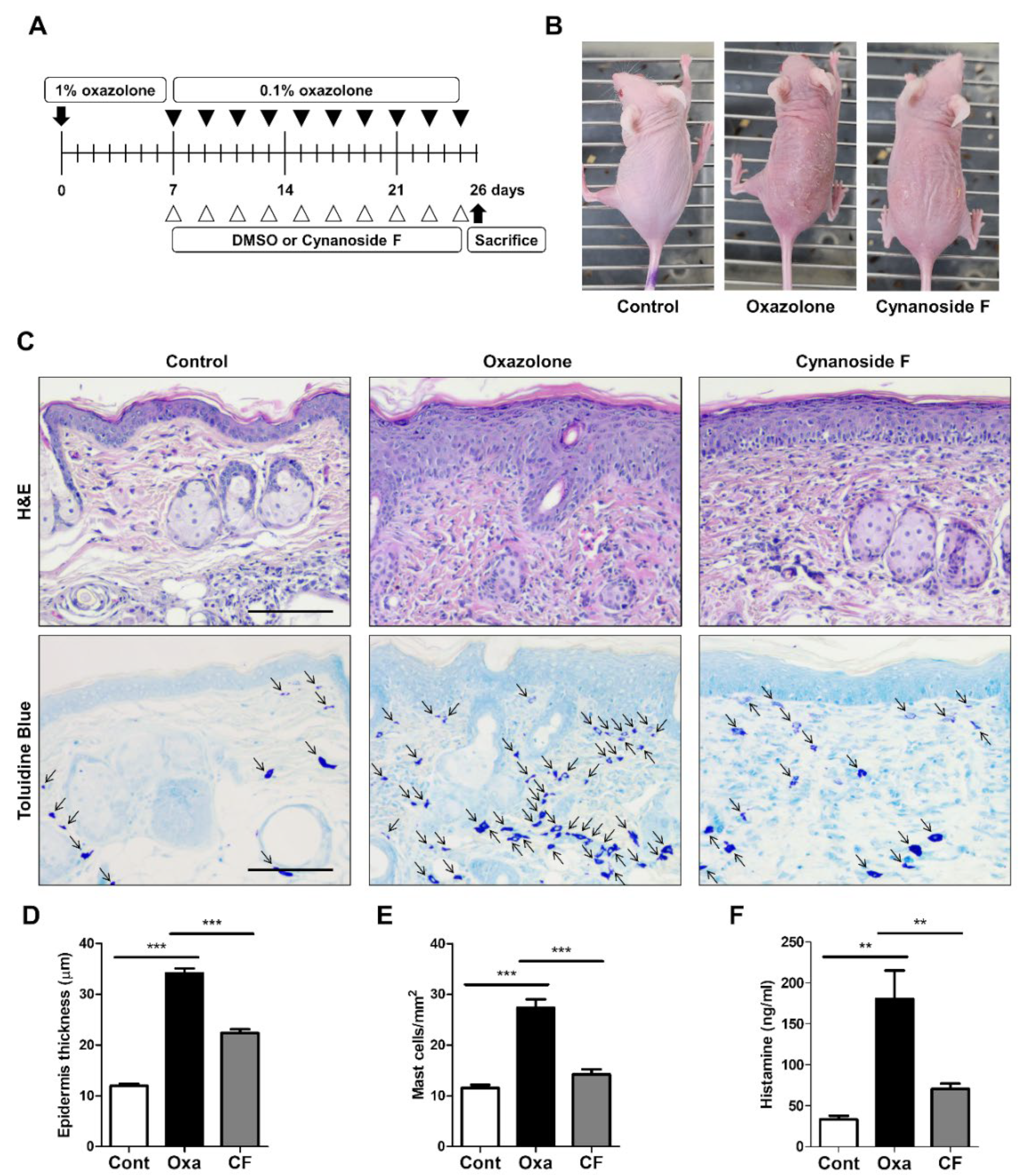
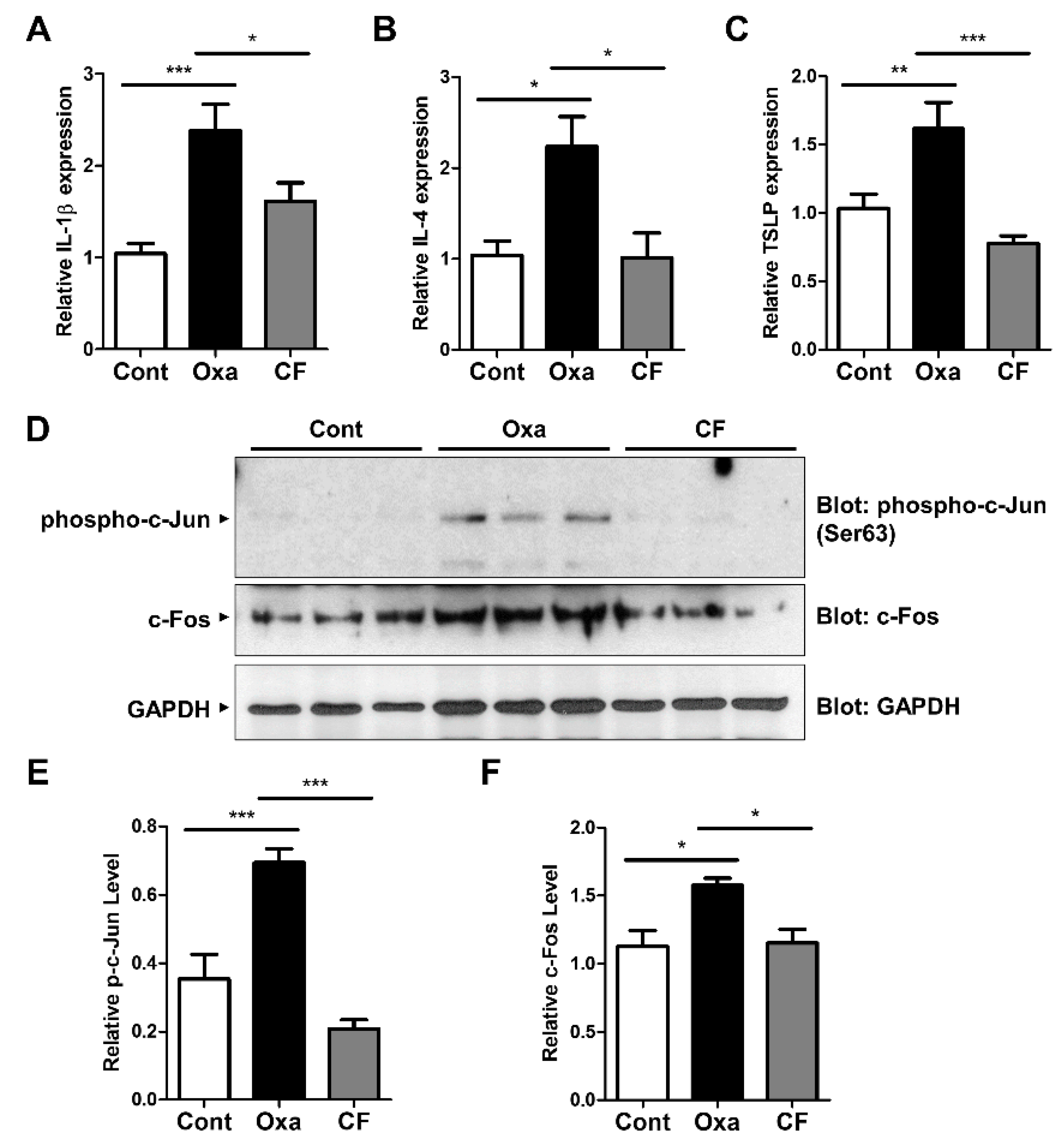
3.4. CF Suppresses Skin Inflammation in an AD Model
3.5. CF Suppresses Proinflammatory Cytokine Expression via AP-1 Inhibition in AD Mouse Tissue
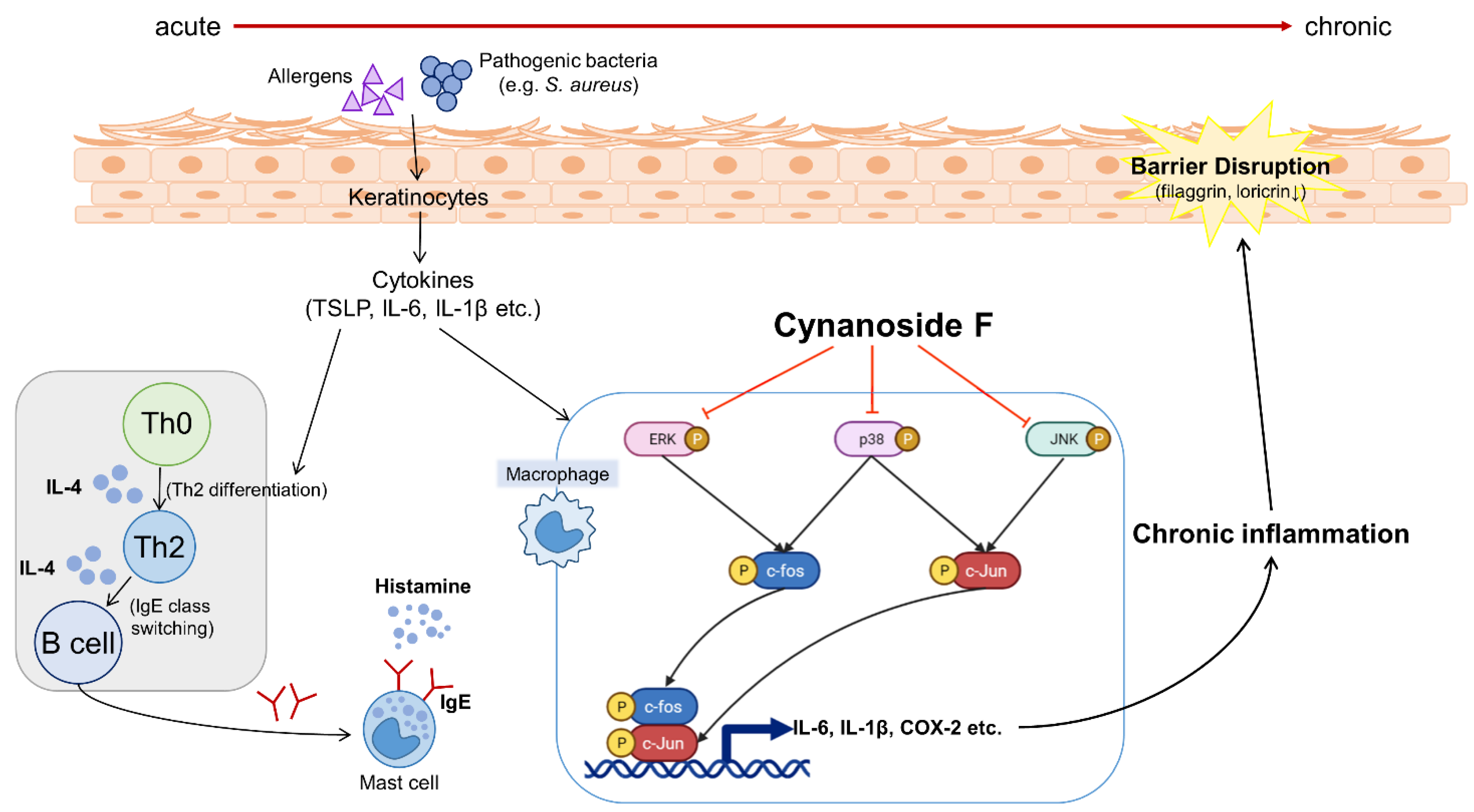
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lynde, C.W.; Bergman, J.; Fiorillo, L.; Guenther, L.; Keddy-Grant, J.; Landells, I.; Marcoux, D.; Ramien, M.; Rehmus, W. Clinical Insights About Topical Treatment of Mild-to-Moderate Pediatric and Adult Atopic Dermatitis. J. Cutan. Med. Surg. 2019, 23, 3S–13S. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Spergel, J.M.; Paller, A.S. Atopic dermatitis and the atopic march. J. Allergy Clin. Immunol. 2003, 112, S118–S127. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Park, J.K.; Choi, J.; Jang, H.; Seol, J.W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef] [PubMed]
- Umehara, Y.; Kiatsurayanon, C.; Trujillo-Paez, J.V.; Chieosilapatham, P.; Peng, G.; Yue, H.; Nguyen, H.L.T.; Song, P.; Okumura, K.; Ogawa, H.; et al. Intractable Itch in Atopic Dermatitis: Causes and Treatments. Biomedicines 2021, 9, 229. [Google Scholar] [CrossRef]
- Eliopoulos, A.G.; Dumitru, C.D.; Wang, C.C.; Cho, J.; Tsichlis, P.N. Induction of COX-2 by LPS in macrophages is regulated by Tpl2-dependent CREB activation signals. EMBO J. 2002, 21, 4831–4840. [Google Scholar] [CrossRef]
- Tang, T.; Scambler, T.E.; Smallie, T.; Cunliffe, H.E.; Ross, E.A.; Rosner, D.R.; O’Neil, J.D.; Clark, A.R. Macrophage responses to lipopolysaccharide are modulated by a feedback loop involving prostaglandin E2, dual specificity phosphatase 1 and tristetraprolin. Sci. Rep. 2017, 7, 4350. [Google Scholar] [CrossRef]
- Choi, H.E.; Kwak, H.J.; Kim, S.K.; Cheon, H.G. Foenumoside B isolated from Lysimachia foenum-graecum extract suppresses LPS-induced inflammatory response via NF-kappaB/AP-1 inactivation in murine macrophages and in endotoxin-induced shock model. Eur. J. Pharmacol. 2018, 832, 120–128. [Google Scholar] [CrossRef]
- Lin, Y.H.; Lin, Y.J.; Chang, T.H.; Chang, Y.H.; Lim, Y.P.; Chung, J.G.; Hsieh, W.T. Pipoxolan suppresses the inflammatory factors of NF-kappaB, AP-1, and STATs, but activates the antioxidative factor Nrf2 in LPS-stimulated RAW 264.7 murine macrophage cells. Environ. Toxicol. 2020, 35, 1352–1363. [Google Scholar] [CrossRef]
- Frazier, W.; Bhardwaj, N. Atopic Dermatitis: Diagnosis and Treatment. Am. Fam. Physician 2020, 101, 590–598. [Google Scholar]
- Dong, H.; Feng, C.; Cai, X.; Hao, Y.; Gu, X.; Cai, L.; Wu, S.; Chen, J.; Liu, Z.; Xie, W.; et al. 7-Methoxyisoflavone ameliorates atopic dermatitis symptoms by regulating multiple signaling pathways and reducing chemokine production. Sci. Rep. 2022, 12, 8760. [Google Scholar] [CrossRef] [PubMed]
- Dorjsembe, B.; Nho, C.W.; Choi, Y.; Kim, J.C. Extract from Black Soybean Cultivar A63 Extract Ameliorates Atopic Dermatitis-like Skin Inflammation in an Oxazolone-Induced Murine Model. Molecules 2022, 27, 2751. [Google Scholar] [CrossRef] [PubMed]
- Zuberbier, T.; Orlow, S.J.; Paller, A.S.; Taieb, A.; Allen, R.; Hernanz-Hermosa, J.M.; Ocampo-Candiani, J.; Cox, M.; Langeraar, J.; Simon, J.C. Patient perspectives on the management of atopic dermatitis. J. Allergy Clin. Immunol. 2006, 118, 226–232. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Zhou, J.; Hayashi, K.; Mitsuhashi, H. Studies on the constituents of Asclepiadaceae plants. LVIII. The structures of five glycosides, cynatratoside-A, -B, -C, -D, and -E, from the Chinese drug “pai-wei”, Cynanchum atratum Bunge. Chem. Pharm. Bull. 1985, 33, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Yoon, J.S.; Kim, E.S.; Kang, S.Y.; Kim, Y.C. Anti-acetylcholinesterase and anti-amnesic activities of a pregnane glycoside, cynatroside B, from Cynanchum atratum. Planta Med. 2005, 71, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Day, S.H.; Wang, J.P.; Won, S.J.; Lin, C.N. Bioactive constituents of the roots of Cynanchum atratum. J. Nat. Prod. 2001, 64, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Kim, N.K.; Seo, S.R. Cynanchi atrati and Its Phenolic Constituent Sinapic Acid Target Regulator of Calcineurin 1 (RCAN1) to Control Skin Inflammation. Antioxidants 2022, 11, 205. [Google Scholar] [CrossRef]
- Bai, H.; Li, W.; Koike, K. Pregnane glycosides from Cynanchum atratum. Steroids 2008, 73, 96–103. [Google Scholar] [CrossRef]
- Bai, H.; Li, W.; Asada, Y.; Satou, T.; Wang, Y.; Koike, K. Twelve pregnane glycosides from Cynanchum atratum. Steroids 2009, 74, 198–207. [Google Scholar] [CrossRef]
- Kim, S.S.; Seo, S.R. The regulator of calcineurin 1 (RCAN1/DSCR1) activates the cAMP response element-binding protein (CREB) pathway. J. Biol. Chem. 2011, 286, 37841–37848. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Christian, F.; Smith, E.L.; Carmody, R.J. The Regulation of NF-kappaB Subunits by Phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Benavides, F.; Oberyszyn, T.M.; VanBuskirk, A.M.; Reeve, V.E.; Kusewitt, D.F. The hairless mouse in skin research. J. Dermatol. Sci. 2009, 53, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Kader, H.A.; Azeem, M.; Jwayed, S.A.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.A.; Hetta, H.F.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Brunner, P.M.; Guttman-Yassky, E.; Leung, D.Y. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J. Allergy Clin. Immunol. 2017, 139, S65–S76. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, A.; Agnihothri, R.; McGirt, L.Y.; Bankova, L.G.; Beck, L.A. Atopic dermatitis: A disease caused by innate immune defects? J. Investig. Dermatol. 2009, 129, 14–30. [Google Scholar] [CrossRef]
- Hussain, Z.; Thu, H.E.; Shuid, A.N.; Kesharwani, P.; Khan, S.; Hussain, F. Phytotherapeutic potential of natural herbal medicines for the treatment of mild-to-severe atopic dermatitis: A review of human clinical studies. Biomed. Pharmacother. 2017, 93, 596–608. [Google Scholar] [CrossRef]
- Wu, S.; Pang, Y.; He, Y.; Zhang, X.; Peng, L.; Guo, J.; Zeng, J. A comprehensive review of natural products against atopic dermatitis: Flavonoids, alkaloids, terpenes, glycosides and other compounds. Biomed. Pharmacother. 2021, 140, 111741. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Ding, M.L.; Tao, L.J.; Zhang, M.; Xu, X.H.; Zhang, C.F. Immunosuppressive C(2)(1) steroidal glycosides from the root of Cynanchum atratum. Fitoterapia 2015, 105, 194–201. [Google Scholar] [CrossRef]
- Han, L.; Zhou, X.; Yang, M.; Zhou, L.; Deng, X.; Wei, S.; Wang, W.; Wang, Z.; Qiao, X.; Bai, C. Ethnobotany, Phytochemistry and Pharmacological Effects of Plants in Genus Cynanchum Linn. (Asclepiadaceae). Molecules 2018, 23, 1194. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.J.; Zhao, X.S.; Tan, Z.B.; Liu, B.; Xu, H.L.; Wu, Y.T.; Xie, L.P.; Bi, Y.M.; Lai, Y.G.; Liang, H.F.; et al. Effects and mechanism of action of Huang-Lian-Jie-Du-Tang in atopic dermatitis-like skin dysfunction in vivo and in vitro. J. Ethnopharmacol. 2019, 240, 111937. [Google Scholar] [CrossRef] [PubMed]
- Pohl, F.; Kong Thoo Lin, P. The Potential Use of Plant Natural Products and Plant Extracts with Antioxidant Properties for the Prevention/Treatment of Neurodegenerative Diseases: In Vitro, In Vivo and Clinical Trials. Molecules 2018, 23, 3283. [Google Scholar] [CrossRef]
- Luo, L.; Yang, J.; Wang, C.; Wu, J.; Li, Y.; Zhang, X.; Li, H.; Zhang, H.; Zhou, Y.; Lu, A.; et al. Natural products for infectious microbes and diseases: An overview of sources, compounds, and chemical diversities. Sci. China Life Sci. 2022, 65, 1123–1145. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kobayashi, T.; Nagao, K. Research Techniques Made Simple: Mouse Models of Atopic Dermatitis. J. Investig. Dermatol. 2019, 139, 984–990.E1. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Size (bp) |
|---|---|---|
| IL-1β | Forward: 5′-CCTCACAAGCAGAGCACAAG-3′ Reverse: 5′-TGTCTTGGCCGAGGACTAAG-3′ | 203 |
| IL-6 | Forward: 5′-TACCACTTCACAAGTCGGAGGC-3′ Reverse: 5′-CTGCAAGTGCATCATCGTTGTTC-3′ | 116 |
| IL-4 | Forward: 5′-ATCATCGGCATTTTGAACGAGGTC-3′ Reverse: 5′-ACCTTGGAAGCCCTACAGACGA-3′ | 125 |
| TSLP | Forward: 5′-CCCTTCACTCCCCGACAAAA-3′ Reverse: 5′-GCAGTGGTCATTGAGGGCTT-3′ | 61 |
| β-actin | Forward: 5′-AGAGGGAAATCGTGCGTGAC-3′ Reverse: 5′-CGATAGTGATGACCTGACCGT-3′ | 138 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleitas, M.M.D.; Kim, S.S.; Kim, N.K.; Seo, S.R. Cynanoside F Controls Skin Inflammation by Suppressing Mitogen-Activated Protein Kinase Activation. Antioxidants 2022, 11, 1740. https://doi.org/10.3390/antiox11091740
Fleitas MMD, Kim SS, Kim NK, Seo SR. Cynanoside F Controls Skin Inflammation by Suppressing Mitogen-Activated Protein Kinase Activation. Antioxidants. 2022; 11(9):1740. https://doi.org/10.3390/antiox11091740
Chicago/Turabian StyleFleitas, Mara Melissa Duarte, Seon Sook Kim, Nam Kyoung Kim, and Su Ryeon Seo. 2022. "Cynanoside F Controls Skin Inflammation by Suppressing Mitogen-Activated Protein Kinase Activation" Antioxidants 11, no. 9: 1740. https://doi.org/10.3390/antiox11091740