
Bacterial Metabolite Reuterin Attenuated LPS-Induced Oxidative Stress and Inflammation Response in HD11 Macrophages

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Phagocytosis Assays
2.5. LPS-Stimulated Inflammatory Model
2.6. Detection of Cellular ROS
2.7. Measurement of Nitric Oxide
2.8. Analysis of Oxidative Stress Indices
2.9. Real-Time Quantitative PCR Assay
2.10. Immunofluorescence Staining
2.11. Western Blotting Analysis
2.12. Statistical Analysis
3. Results
3.1. Effect of Reuterin on Cell Viability of HD11 Cell Line
3.2. Effects of Reuterin on the Expressional Levels of Polarization-Associated Genes of HD11 Cells
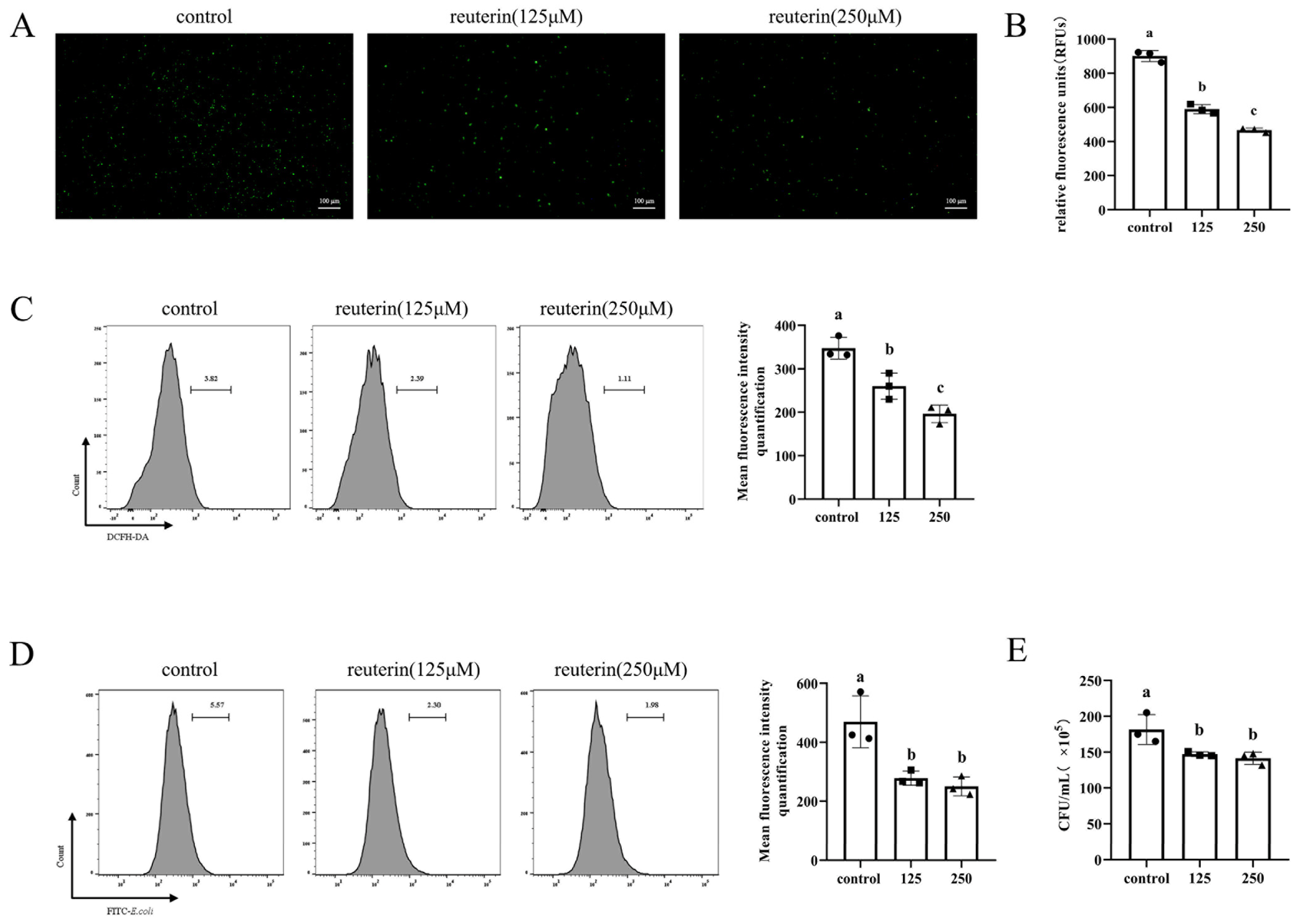
3.3. Reuterin Suppressed ROS Production and Phagocytosis Activity of HD11 Cells
3.4. Pretreatment and Cotreatment of HD11 Macrophages with Reuterin Inhibited Nitric Oxide Production When Exposed to LPS Stimulation
3.5. Reuterin Inhibited the Oxidative Stress in LPS-Stimulated HD11 Cells by Nrf2/HO-1 Signaling Pathway
3.6. Reuterin Inhibited the Inflammatory Response in LPS-Stimulated HD11 Cells
3.7. Reuterin Inhibited the LPS-Stimulated NF-κB p65 Nuclear Translocation in HD11 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramond, E.; Jamet, A.; Coureuil, M.; Charbit, A. Pivotal Role of Mitochondria in Macrophage Response to Bacterial Pathogens. Front. Immunol 2019, 10, 2461. [Google Scholar] [CrossRef]
- Twigg, H.L. Macrophages in Innate and Acquired Immunity. Semin. Respir. Crit. Care Med. 2004, 25, 21–31. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef]
- Fernando, M.R.; Reyes, J.L.; Iannuzzi, J.; Leung, G.; McKay, D.M. The Pro-Inflammatory Cytokine, Interleukin-6, Enhances the Polarization of Alternatively Activated Macrophages. PLoS ONE 2014, 9, e94188. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Rath, M.; Muller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Mosser, D.M. The many faces of macrophage activation. J. Leukoc. Biol. 2003, 73, 209–212. [Google Scholar] [CrossRef]
- Jaffar, N.; Okinaga, T.; Nishihara, T.; Maeda, T. Enhanced phagocytosis of Aggregatibacter actinomy-cetemcomitans cells by macrophages activated by a probiotic Lactobacillus strain. J. Dairy Sci. 2018, 101, 5789–5798. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.K.; Wu, Y.P.; Liu, R.R.; Xu, H.; Mei, X.Q.; Shang, Q.Q.; Liu, S.J.; Yu, D.Y.; Li, W.F. Lactobacillus rhamnosus GG promotes M1 polarization in murine bone marrow-derived macrophages by activating TLR2/MyD88/MAPK signaling pathway. Anim. Sci. J. 2020, 91, e13439. [Google Scholar] [CrossRef]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsua, W.H.; Pan, T.M. Immunomodulatory and antioxidant po-tential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid.-Based Complementary Altern. Med. 2018, 2018, 1756308. [Google Scholar] [CrossRef]
- Ji, J.; Shu, D.M.; Zheng, M.Z.; Wang, J.; Luo, C.L.; Wang, Y.; Guo, F.Y.; Zou, X.; Lv, X.H.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Jang, S.E.; Han, M.J.; Kim, S.Y.; Kim, D.H. Lactobacillus plantarum CLP-0611 ameliorates colitis in mice by polarizing M1 to M2-like macrophages. Int. Immunopharmacol. 2014, 21, 186–192. [Google Scholar] [CrossRef]
- Vollenweider, S.; Lacroix, C. 3-Hydroxypropionaldehyde: Applications and perspectives of biotechno-logical production. Appl. Microbiol. Biotechnol. 2004, 64, 16–27. [Google Scholar] [CrossRef]
- Talarico, T.L.; Dobrogosz, W.J. Purification and Characterization of Glycerol Dehydratase from Lacto-bacillus-Reuteri. Appl. Environ. Microbiol. 1990, 56, 1195–1197. [Google Scholar] [CrossRef]
- Cleusix, V.; Lacroix, C.; Vollenweider, S.; Duboux, M.; Le Blay, G. Inhibitory activity spectrum of reuterin produced by Lactobacillus reuteri against intestinal bacteria. BMC Microbiol. 2007, 7, 1–9. [Google Scholar] [CrossRef]
- Avila, M.; Gomez-Torres, N.; Hernandez, M.; Garde, S. Inhibitory activity of reuterin, nisin, lysozyme and nitrite against vegetative cells and spores of dairy-related Clostridium species. Int. J. Food Microbiol. 2014, 172, 70–75. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Bell, H.N.; Rebernick, R.J.; Goyert, J.; Singhal, R.; Kuljanin, M.; Kerk, S.A.; Huang, W.; Das, N.K.; Andren, A.; Solanki, S. Reuterin in the healthy gut microbiome suppresses colorectal cancer growth through altering redox balance. Cancer Cell 2022, 40, 185–200. [Google Scholar] [CrossRef]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Roth, N.; Kasbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Ben, Y.J.; Fu, C.X.; Hu, M.; Liu, L.; Wong, M.H.; Zheng, C.M. Human health risk assessment of antibiotic re-sistance associated with antibiotic residues in the environment: A review. Environ. Res. 2019, 169, 483–493. [Google Scholar] [CrossRef]
- Lee, S.C.; Wang, S.Y.; Li, C.C.; Liu, C.T. Anti-inflammatory effect of cinnamaldehyde and linalool from the leaf essential oil of Cinnamomum osmophloeum Kanehira in endotoxin-induced mice. J. Food Drug Anal. 2018, 26, 211–220. [Google Scholar] [CrossRef]
- Xun, W.J.; Shi, L.G.; Zhou, H.L.; Hou, G.Y.; Cao, T.; Zhao, C.P. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef]
- Wang, K.L.; Zhang, H.R.; Han, Q.J.; Lan, J.H.; Chen, G.Y.; Cao, G.T.; Yang, C.M. Effects of astragalus and ginseng polysaccharides on growth performance, immune function and intestinal barrier in weaned piglets chal-lenged with lipopolysaccharide. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1096–1105. [Google Scholar] [CrossRef]
- Beug, H.; Vonkirchbach, A.; Doderlein, G.; Conscience, J.F.; Graf, T. Chicken Hematopoietic-Cells Trans-formed by 7 Strains of Defective Avian Leukemia Viruses Display 3 Distinct Phenotypes of Differentiation. Cell 1979, 18, 375–390. [Google Scholar] [CrossRef]
- Slawinska, A.; Hsieh, J.C.; Schmidt, C.J.; Lamont, S.J. Heat Stress and Lipopolysaccharide Stimulation of Chicken Macrophage-Like Cell Line Activates Expression of Distinct Sets of Genes. PLoS ONE 2016, 11, e0164575. [Google Scholar] [CrossRef]
- Selvaraj, R.K.; Klasing, K.C. Lutein and eicosapentaenoic acid interact to modify iNOS mRNA levels through the PPAR gamma/RXR pathway in chickens and HD11 cell lines. J. Nutr. 2006, 136, 1610–1616. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.Y.; Li, C.; Qin, Y.Q.; Yin, R.L.; Du, S.W.; Liu, H.F.; Zhang, Y.F.; Wang, C.Y.; Rong, F.J.; Jin, N.Y. Evaluation of immunomodulatory activity of two potential probiotic Lactobacillus strains by in vivo tests. Anaerobe 2015, 35, 22–27. [Google Scholar] [CrossRef]
- de LeBlanc, A.D.; Chaves, S.; Carmuega, E.; Weill, R.; Antoine, J.; Perdigon, G. Effect of long-term continu-ous consumption of fermented milk containing probiotic bacteria on mucosal immunity and the activity of peritoneal macrophages. Immunobiology 2008, 213, 97–108. [Google Scholar] [CrossRef]
- Cao, X.F.; Tang, L.; Zeng, Z.H.; Wang, B.K.; Zhou, Y.H.; Wang, Q.; Zou, P.; Li, W.F. Effects of Probiotics BaSC06 on Intes-tinal Digestion and Absorption, Antioxidant Capacity, Microbiota Composition, and Macrophage Polariza-tion in Pigs for Fattening. Front. Vet. Sci. 2020, 7, 570593. [Google Scholar] [CrossRef]
- Lim, S.M.; Jeong, J.J.; Jang, S.E.; Han, M.J.; Kim, D.H. A mixture of the probiotic strains Bifidobacterium longum CH57 and Lactobacillus brevis CH23 ameliorates colitis in mice by inhibiting macrophage activa-tion and restoring the Th17/Treg balance. J. Funct. Foods 2016, 27, 295–309. [Google Scholar] [CrossRef]
- Ohue-Kitano, R.; Yasuoka, Y.; Goto, T.; Kitamura, N.; Park, S.B.; Kishino, S.; Kimura, I.; Kasubuchi, M.; Takahashi, H.; Li, Y.; et al. α-Linolenic acid-derived metabolites from gut lactic acid bacteria induce differentia-tion of anti-inflammatory M2 macrophages through G protein-coupled receptor 40. Fed. Am. Soc. Exp. Biol. J. 2018, 32, 304–318. [Google Scholar] [CrossRef]
- Liu, T.F.; Li, J.; Liu, Y.X.; Xiao, N.; Suo, H.T.; Xie, K.; Yang, C.L.; Wu, C. Short-Chain Fatty Acids Suppress Lipopolysac-charide-Induced Production of Nitric Oxide and Proinflammatory Cytokines Through Inhibition of NF-kappa B Pathway in RAW264.7 Cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.M.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intes-tinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, E.J.; Lee, J.C.; Kim, W.K.; Kim, H.S. Anti-inflammatory effects of short chain fatty acids in IFN-gamma-stimulated RAW 264.7 murine macrophage cells: Involvement of NF-kappa B and ERK sig-naling pathways. Int. Immunopharmacol. 2007, 7, 70–77. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.W.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Pro-duced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37. [Google Scholar] [CrossRef]
- Krishnan, S.; Ding, Y.F.; Saeidi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K.B. Gut Microbiota-Derived Tryptophan Metabolites Modulate Inflammatory Response in Hepatocytes and Macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef]
- Talarico, T.L.; Casas, I.A.; Chung, T.C.; Dobrogosz, W.J. Production and Isolation of Reuterin, a Growth In-hibitor Produced by Lactobacillus-Reuteri. Antimicrob. Agents Chemother. 1988, 32, 1854–1858. [Google Scholar] [CrossRef] [PubMed]
- Engevik, M.A.; Danhof, H.A.; Shrestha, R.; Chang-Graham, A.L.; Hyser, J.M.; Haag, A.M.; Mohammad, M.A.; Britton, R.A.; Versalovic, J.; Sorg, J.A.; et al. Reuterin dis-rupts Clostridioides difficile metabolism and pathogenicity through reactive oxygen species generation. Gut Microbes 2020, 12, 1795388. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Park, S.H.; Kang, M.K.; Kim, Y.H.; Lee, E.J.; Kim, D.Y.; Kim, S.I.; Oh, S.Y.; Lim, S.S.; Kang, Y.H. Asaronic Acid Attenuates Macrophage Activation toward M1 Phenotype through Inhibition of NF-kappa B Pathway and JAK-STAT Signaling in Glucose-Loaded Murine Macrophages. J. Agric. Food Chem. 2019, 67, 10069–10078. [Google Scholar] [CrossRef]
- Pengal, R.A.; Ganesan, L.P.; Wei, G.; Fang, H.Q.; Ostrowski, M.C.; Tridandapani, S. Lipopolysaccha-ride-induced production of interleukin-10 is promoted by the serine/threonine kinase Akt. Mol. Immunol. 2006, 43, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Chanteux, H.; Guisset, A.C.; Pilette, C.; Sibille, Y. LPS induces IL-10 production by human alveolar mac-rophages via MAPKinases- and Sp1-dependent mechanisms. Respir. Res. 2007, 8, 1–10. [Google Scholar] [CrossRef]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Luo, J.F.; Shen, X.Y.; Lio, C.K.; Dai, Y.; Cheng, C.S.; Liu, J.X.; Yao, Y.D.; Yu, Y.; Xie, Y.; Luo, P.; et al. Activation of Nrf2/HO-1 Pathway by Nar-dochinoid C Inhibits Inflammation and Oxidative Stress in Lipopolysaccharide-Stimulated Macrophages. Front. Pharmacol. 2018, 9, 911. [Google Scholar] [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjornsson, B. The Role of Mitogen-Activated Protein Kinase-Activated Protein Kinases (MAPKAPKs) in Inflammation. Genes 2013, 4, 101–133. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Forward Sequence (5′-3′) | Reward Sequence (5′-3′) |
|---|---|---|---|
| TLR4 | NM_001030693 | ATCTTTCAAGGTGCCACATC | GGATATGCTTGTTTCCACCA |
| MyD88 | NM_001030962 | CTGGCATCTTCTGAGTAGT | TTCCTTATAGTTCTGGCTTCT |
| TRAF6 | XM_040673311 | TCTGTTTGTCCACACGATGC | TATCTCTGGCTTGGCTTCCA |
| iNOS | NM_204961 | AAAGATGATGCCAAATTACACA | TCCGACAATTGATAACCTCC |
| CD206 | NM_001397660 | GAGGACTGCGTTGTTATGA | TCTTCTGTCGGTGCTTCT |
| CD86 | NM_001037839 | GGATGTCTTACAGGATGCT | CTGCTCTCCAAGGTGAAG |
| IL-1β | NM_204524 | ACTGGGCATCAAGGGCTA | GGTAGAAGATGAAGCGGGTC |
| IL-6 | NM_204628 | ATCCTCTGTTACCAATCTGC | ACATTTTCTTTGGCGTTGAC |
| IL-10 | NM_001004414 | TGCTGCGCTTCTACACAGAT | TGGCTTTGCTCCTCTTCTCG |
| TGF-β1 | NM_001318456 | GATGGACCCGATGAGTATTG | CGTTGAACACGAAGAAGATG |
| β-actin | NM_205518 | GAGAAATTGTGCGTGACATCA | CCTGAACCTCTCATTGCCA |
| Antibody Name | Dilution Ratio | Manufacturers |
|---|---|---|
| iNOS | 1:1000 | HuaAn Biotechnology Co., Ltd., Hangzhou, China |
| IL-1β | 1:500 | Cloud-clone crop, Co., Ltd., Wuhan, China |
| IL-10 | 1:500 | Cloud-clone crop, Co., Ltd., Wuhan, China |
| p-Nrf2 | 1:1000 | HuaAn Biotechnology Co., Ltd., Hangzhou, China |
| Nrf2 | 1:1000 | HuaAn Biotechnology Co., Ltd., Hangzhou, China |
| HO-1 | 1:500 | Boster Biological Technology, Wuhan, China |
| TRAF6 | 1:1000 | HuaAn Biotechnology Co., Ltd., Hangzhou, China |
| NF-κB p65 | 1:500 | Abcam, Cambridge, UK |
| p-ERK | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| ERK | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| p-JNK | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| JNK | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| p-p38 | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| p38 | 1:1000 | Cell signaling technology Inc, Beverly, MA, USA |
| β-actin | 1:10,000 | Abclonal, Biotechnology Co., Ltd., Wuhan, China |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Ding, X.; Wang, Y.; Li, D.; Xie, L.; Liang, S.; Zhang, Y.; Li, W.; Fu, A.; Zhan, X. Bacterial Metabolite Reuterin Attenuated LPS-Induced Oxidative Stress and Inflammation Response in HD11 Macrophages. Antioxidants 2022, 11, 1662. https://doi.org/10.3390/antiox11091662
Xu Y, Ding X, Wang Y, Li D, Xie L, Liang S, Zhang Y, Li W, Fu A, Zhan X. Bacterial Metabolite Reuterin Attenuated LPS-Induced Oxidative Stress and Inflammation Response in HD11 Macrophages. Antioxidants. 2022; 11(9):1662. https://doi.org/10.3390/antiox11091662
Chicago/Turabian StyleXu, Yibin, Xiaoqing Ding, Yuanyuan Wang, Danlei Li, Lingyu Xie, Shuang Liang, Yunfeng Zhang, Weifen Li, Aikun Fu, and Xiuan Zhan. 2022. "Bacterial Metabolite Reuterin Attenuated LPS-Induced Oxidative Stress and Inflammation Response in HD11 Macrophages" Antioxidants 11, no. 9: 1662. https://doi.org/10.3390/antiox11091662