
Selenoprotein: Potential Player in Redox Regulation in Chlamydomonas reinhardtii



Abstract
:1. Introduction
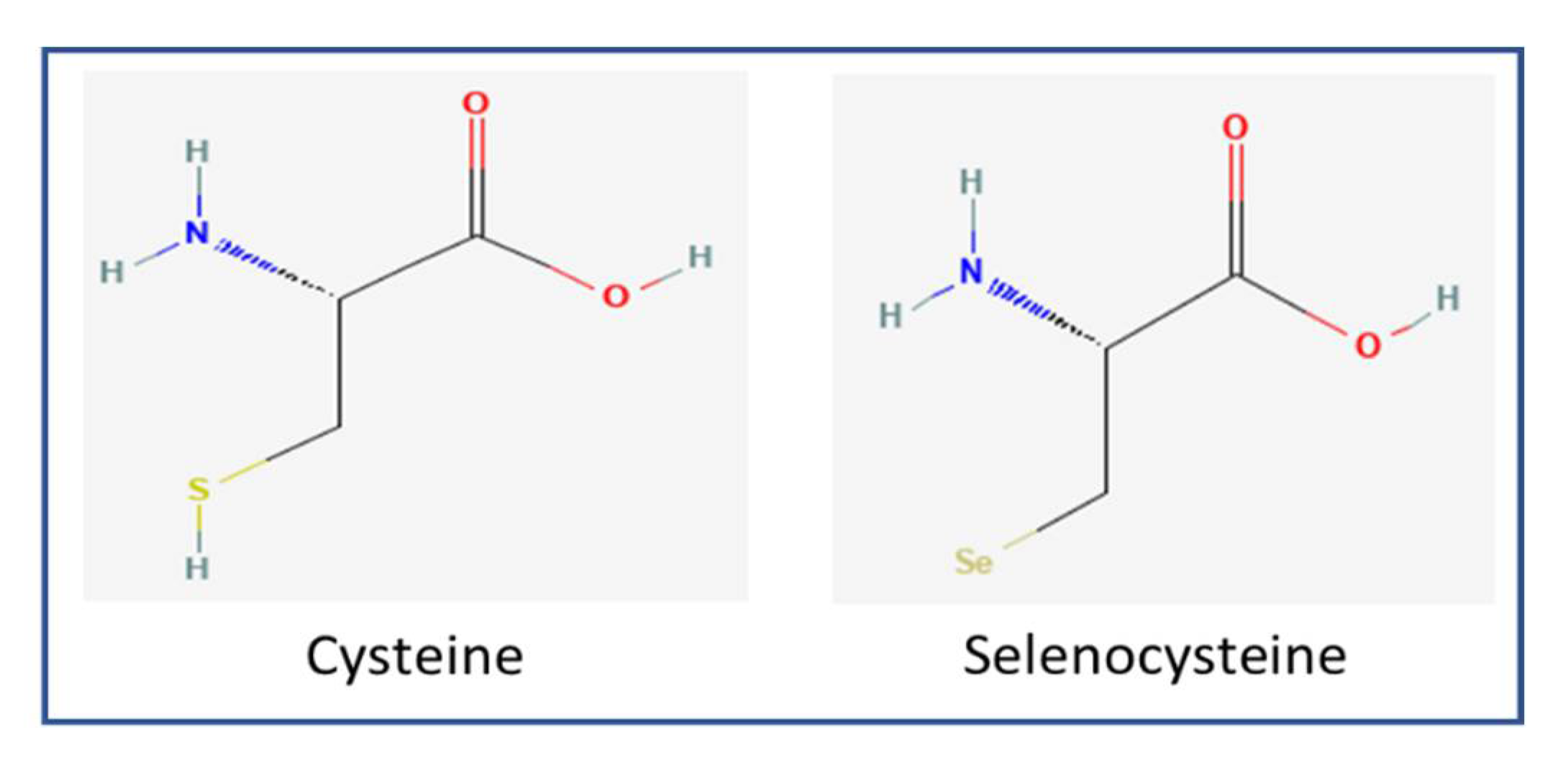
2. Selenoproteins and Cysteine-Containing Homologs
3. Selenoproteins from Algae
3.1. Glutathione Peroxidase (GPX)
3.2. Thioredoxin Reductase (TrxR)
3.3. Other Selenoproteins
4. Selenoproteins from Chlamydomonas Can Be Potential Antioxidants
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-Dependent Antioxidant Enzymes: Actions and Properties of Selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [Green Version]
- Seale, L.A. Selenocysteine β-Lyase: Biochemistry, Regulation and Physiological Role of the Selenocysteine Decomposition Enzyme. Antioxidants 2019, 8, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qazi, I.H.; Angel, C.; Yang, H.; Zoidis, E.; Pan, B.; Wu, Z.; Ming, Z.; Zeng, C.-J.; Meng, Q.; Han, H.; et al. Role of Selenium and Selenoproteins in Male Reproductive Function: A Review of Past and Present Evidences. Antioxidants 2019, 8, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, L.; Gafvelin, G.; Arnér, E.S.J. Selenocysteine in proteins—Properties and biotechnological use. Biochim. Biophys. Acta-Gen. Subj. 2005, 1726, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bulteau, A.L.; Chavatte, L. Update on Selenoprotein Biosynthesis. Antioxid. Redox Signal. 2015, 23, 775–794. [Google Scholar] [CrossRef]
- Turanov, A.A.; Xu, X.-M.; Carlson, B.A.; Yoo, M.-H.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of Selenocysteine, the 21st Amino Acid in the Genetic Code, and a Novel Pathway for Cysteine Biosynthesis. Adv. Nutr. Int. Rev. J. 2011, 2, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Shetty, S.; Copeland, P.R. Molecular Mechanism of Selenoprotein P Synthesis. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 2506–2510. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, R.; Rozovsky, S. Synthesis and semisynthesis of selenopeptides and selenoproteins. Curr. Opin. Chem. Biol. 2018, 46, 41–47. [Google Scholar] [CrossRef]
- Serrão, V.H.B.; Silva, I.R.; da Silva, M.T.A.; Scortecci, J.F.; de Freitas Fernandes, A.; Thiemann, O.H. The unique tRNASec and its role in selenocysteine biosynthesis. Amino Acids 2018, 50, 1145–1167. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-M.; Carlson, B.A.; Mix, H.; Zhang, Y.; Saira, K.; Glass, R.S.; Berry, M.J.; Gladyshev, V.N.; Hatfield, D.L. Biosynthesis of Selenocysteine on Its tRNA in Eukaryotes. PLoS Biol. 2006, 5, e4. [Google Scholar] [CrossRef]
- Sherrer, R.L.; O’Donoghue, P.; Söll, D. Characterization and evolutionary history of an archaeal kinase involved in selenocysteinyl-tRNA formation. Nucleic Acids Res. 2008, 36, 1247–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.-M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Palioura, S.; Salazar, J.C.; Su, D.; O’Donoghue, P.; Hohn, M.J.; Cardoso, A.M.; Whitman, W.B.; Söll, D. RNA-dependent conversion of phosphoserine forms selenocysteine in eukaryotes and archaea. Proc. Natl. Acad. Sci. USA 2006, 103, 18923–18927. [Google Scholar] [CrossRef] [Green Version]
- Dobosz-Bartoszek, M.; Simonović, M. Structure and Mechanism of Selenocysteine Synthases. In Selenium; Springer International Publishing: Cham, Switzerland, 2016; pp. 101–112. [Google Scholar]
- Veres, Z.; Kim, I.; Scholz, T.; Stadtman, T. Selenophosphate synthetase. Enzyme properties and catalytic reaction. J. Biol. Chem. 1994, 269, 10597–10603. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular Pathways and Physiological Roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [Green Version]
- Ambrogelly, A.; Palioura, S.; Söll, D. Natural expansion of the genetic code. Nat. Chem. Biol. 2006, 3, 29–35. [Google Scholar] [CrossRef]
- Fu, X.; Söll, D.; Sevostyanova, A. Challenges of site-specific selenocysteine incorporation into proteins by Escherichia coli. RNA Biol. 2018, 15, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Berry, M.; Banu, L.; Harney, J.; Larsen, P. Functional characterization of the eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons. EMBO J. 1993, 12, 3315–3322. [Google Scholar] [CrossRef]
- Ringquist, S.; Schneider, D.; Gibson, T.; Baron, C.; Böck, A.; Gold, L. Recognition of the mRNA selenocysteine insertion sequence by the specialized translational elongation factor SELB. Genes Dev. 1994, 8, 376–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Reches, M.; Groisman, I.; Engelberg-Kulka, H. The nature of the minimal ‘selenocysteine insertion sequence’ (SECIS) in Escherichia coli. Nucleic Acids Res. 1998, 26, 896–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krol, A. Evolutionarily different RNA motifs and RNA–protein complexes to achieve selenoprotein synthesis. Biochimie 2002, 84, 765–774. [Google Scholar] [CrossRef]
- Chavatte, L.; Brown, B.A.; Driscoll, D.M. Ribosomal protein L30 is a component of the UGA-selenocysteine recoding machinery in eukaryotes. Nat. Struct. Mol. Biol. 2005, 12, 408–416. [Google Scholar] [CrossRef]
- Wessjohann, L.A.; Schneider, A.; Abbas, M.; Brandt, W. Selenium in chemistry and biochemistry in comparison to sulfur. Biol. Chem. 2007, 388, 997–1006. [Google Scholar] [CrossRef]
- Arnér, E.S.J. Selenoproteins—What unique properties can arise with selenocysteine in place of cysteine? Exp. Cell Res. 2010, 316, 1296–1303. [Google Scholar] [CrossRef]
- Lobanov, A.V.; Hatfield, D.L.; Gladyshev, V.N. Eukaryotic Selenoproteins and Selenoproteomes. Biochim. Biophys. Acta-Gen. Subj. 2009, 1790, 1424–1428. [Google Scholar] [CrossRef] [Green Version]
- Mousa, R.; Dardashti, R.N.; Metanis, N. Selenium and Selenocysteine in Protein Chemistry. Angew. Chem. Int. Ed. 2017, 56, 15818–15827. [Google Scholar] [CrossRef]
- Gromer, S.; Wissing, J.; Behne, D.; Ashman, K.; Schirmer, R.H.; Floh, L.; Becker, K. A hypothesis on the catalytic mechanism of the selenoenzyme thioredoxin reductase. Biochem. J. 1998, 332, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Gorlatov, S.N.; Stadtman, T.C. Human Selenium-Dependent Thioredoxin Reductase from HeLa Cells: Properties of Forms with Differing Heparin Affinities. Arch. Biochem. Biophys. 1999, 369, 133–142. [Google Scholar] [CrossRef]
- Chung, S.S.; Kim, M.; Youn, B.-S.; Lee, N.S.; Park, J.W.; Lee, I.K.; Lee, Y.S.; Kim, J.B.; Cho, Y.M.; Lee, H.K.; et al. Glutathione Peroxidase 3 Mediates the Antioxidant Effect of Peroxisome Proliferator-Activated Receptor γ in Human Skeletal Muscle Cells. Mol. Cell. Biol. 2009, 29, 20–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-R.; Bar-Noy, S.; Kwon, J.; Levine, R.L.; Stadtman, T.C.; Rhee, S.G. Mammalian thioredoxin reductase: Oxidation of the C-terminal cysteine/selenocysteine active site forms a thioselenide, and replacement of selenium with sulfur markedly reduces catalytic activity. Proc. Natl. Acad. Sci. USA 2000, 97, 2521–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, S.; Schneider, I.; Pan, J.; Von Hacht, A.; Bardwell, J.C.A. The CXXC Motif Is More than a Redox Rheostat. J. Biol. Chem. 2007, 282, 28823–28833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From Selenium to Selenoproteins: Synthesis, Identity, and Their Role in Human Health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Peterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs Regulate Actin Assembly and Macrophage Function via Reversible Stereoselective Methionine Oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, W.C.; Alkan, Z. Regulation of Redox Signaling by Selenoproteins. Biol. Trace Elem. Res. 2010, 134, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, J.; Ronga, L.; Szpunar, J.; Lobinski, R. Characterization and Quantification of Selenoprotein P: Challenges to Mass Spectrometry. Int. J. Mol. Sci. 2021, 22, 6283. [Google Scholar] [CrossRef]
- Arnér, E.S.J.; Sarioglu, H.; Lottspeich, F.; Holmgren, A.; Böck, A. High-level expression in Escherichia coli of selenocysteine-containing rat thioredoxin reductase utilizing gene fusions with engineered bacterial-type SECIS elements and co-expression with the selA, selB and selC genes. J. Mol. Biol. 1999, 292, 1003–1016. [Google Scholar] [CrossRef]
- Arnér, E.S.J. Recombinant Expression of Mammalian Selenocysteine-Containing Thioredoxin Reductase and Other Selenoproteins in Escherichia coli. Methods Enzymol. 2002, 347, 226–235. [Google Scholar] [PubMed]
- Johansson, L.; Chen, C.; Thorell, J.-O.; Fredriksson, A.; Stone-Elander, S.; Gafvelin, G.; Arnér, E.S.J. Exploiting the 21st amino acid—purifying and labeling proteins by selenolate targeting. Nat. Chem. Biol. 2004, 1, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Stone-Elander, S.; Arnér, E.S.J. Tagging recombinant proteins with a Sel-tag for purification, labeling with electrophilic compounds or radiolabeling with 11C. Nat. Protoc. 2006, 1, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Novoselov, S.; Rao, M.; Onoshko, N.V.; Zhi, H.; Kryukov, G.; Xiang, Y.; Weeks, D.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins and selenocysteine insertion system in the model plant cell system, Chlamydomonas reinhardtii. EMBO J. 2002, 21, 3681–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.-H.; Wang, X.-F.; Eyal, Y.; She, Y.-M.; Donald, L.J.; Standing, K.G.; Ben-Hayyim, G. A Selenoprotein in the Plant Kingdom. J. Biol. Chem. 2002, 277, 25983–25991. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Lu, Y.; Zheng, L.; Li, G.; Chen, L.; Zhang, M.; Ni, J.; Liu, Q.; Zhang, Y. The algal selenoproteomes. BMC Genom. 2020, 21, 699. [Google Scholar] [CrossRef]
- Jiang, L.; Ni, J.; Liu, Q. Evolution of selenoproteins in the metazoan. BMC Genom. 2012, 13, 446. [Google Scholar] [CrossRef] [Green Version]
- Flohé, L. Glutathione Peroxidase Brought into Focus. In Free Radicals in Biology; Pryor, W.A., Ed.; Academic Press: New York, NY, USA, 2012; Volume V, pp. 223–277. [Google Scholar]
- Flohé, L.; Günzler, W.A. [12] Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–120. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigoó, R.; Gladyshev, V.N. Characterization of Mammalian Selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- El Haddad, M.; Jean, E.; Turki, A.; Hugon, G.; Vernus, B.; Bonnieu, A.; Passerieux, E.; Hamade, A.; Mercier, J.; Laoudj-Chenivesse, D.; et al. Glutathione peroxidase 3, a new retinoid target gene, is crucial for human skeletal muscle precursor cell survival. J. Cell Sci. 2012, 125, 6147–6156. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.J.; Diamond, A.M. Role of glutathione peroxidase 1 in breast cancer: Loss of heterozygosity and allelic differences in the response to selenium. Cancer Res. 2003, 63, 3347–3351. [Google Scholar] [PubMed]
- Hu, Y.; Benya, R.V.; Carroll, R.E.; Diamond, A.M. Allelic Loss of the Gene for the GPX1 Selenium-Containing Protein Is a Common Event in Cancer. J. Nutr. 2005, 135, 3021S–3024S. [Google Scholar] [CrossRef] [PubMed]
- Carlson, B.A.; Tobe, R.; Yefremova, E.; Tsuji, P.A.; Hoffmann, V.J.; Schweizer, U.; Gladyshev, V.N.; Hatfield, D.L.; Conrad, M. Glutathione peroxidase 4 and vitamin E cooperatively prevent hepatocellular degeneration. Redox Biol. 2016, 9, 22–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachara, B.A.; Gromadzinska, J.; Zbrog, Z.; Swiech, R.; Wasowicz, W.; Twardowska, E.; Jablonska, E.; Sobala, W. Selenium supplementation to chronic kidney disease patients on hemodialysis does not induce the synthesis of plasma glutathione peroxidase. Acta Biochim. Pol. 2009, 56, 183–187. [Google Scholar] [CrossRef]
- Taccone-Gallucci, M.; Noce, A.; Bertucci, P.; Fabbri, C.; Manca-Di-Villahermosa, S.; Della-Rovere, F.R.; De Francesco, M.; Lonzi, M.; Federici, G.; Scaccia, F.; et al. Chronic treatment with statins increases the availability of selenium in the antioxidant defence systems of hemodialysis patients. J. Trace Elem. Med. Biol. 2010, 24, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, B.R.; Hare, D.J.; Bush, A.I.; Roberts, B.R. Glutathione Peroxidase 4: A New Player in Neurodegeneration? Mol. Psychiatry 2017, 22, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Hambright, W.S.; Fonseca, R.S.; Chen, L.; Na, R.; Ran, Q. Ablation of ferroptosis regulator glutathione peroxidase 4 in forebrain neurons promotes cognitive impairment and neurodegeneration. Redox Biol. 2017, 12, 8–17. [Google Scholar] [CrossRef]
- Chen, L.; Hambright, W.S.; Na, R.; Ran, Q. Ablation of the Ferroptosis Inhibitor Glutathione Peroxidase 4 in Neurons Results in Rapid Motor Neuron Degeneration and Paralysis. J. Biol. Chem. 2015, 290, 28097–28106. [Google Scholar] [CrossRef] [Green Version]
- Beeor-Tzahar, T.; Ben-Hayyim, G.; Holland, D.; Faltin, Z.; Eshdat, Y. A stress-associated citrus protein is a distinct plant phospholipid hydroperoxide glutathione peroxidase. FEBS Lett. 1995, 366, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Stadtman, T.C. Selenocysteine. Annu. Rev. Biochem. 1996, 65, 83–100. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical Role as a Component of Glutathione Peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F. Molecular biology of selenium with implications for its metabolism 1. FASEB J. 1991, 5, 2274–2279. [Google Scholar] [CrossRef]
- Florian, S.; Krehl, S.; Loewinger, M.; Kipp, A.; Banning, A.; Esworthy, S.; Chu, F.-F.; Brigelius-Flohé, R. Loss of GPx2 increases apoptosis, mitosis, and GPx1 expression in the intestine of mice. Free Radic. Biol. Med. 2010, 49, 1694–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brigelius-Flohé, R.; Banning, A.; Schnurr, K. Selenium-Dependent Enzymes in Endothelial Cell Function. Antioxid. Redox Signal. 2003, 5, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Nishinaka, T.; Matsuno, K.; Yabe-Nishimura, C. Increased Gene Expression of Glutathione Peroxidase-3 in Diabetic Mouse Heart. Biol. Pharm. Bull. 2006, 29, 1042–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of Adipose Glutathione Peroxidase 3 in Obesity Contributes to Local and Systemic Oxidative Stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.P.; Maiorino, M.; Ursini, F.; Girotti, A.W. Protective action of phospholipid hydroperoxide glutathione peroxidase against membrane-damaging lipid peroxidation. In situ reduction of phospholipid and cholesterol hydroperoxides. J. Biol. Chem. 1990, 265, 454–461. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M.; Roveri, A. Phospholipid Hydroperoxide Glutathione Peroxidase (PHGPx): More Than an Antioxidant Enzyme? Biomed. Environ. Sci. 1997, 10, 327–332. [Google Scholar]
- Seiler, A.; Schneider, M.; Förster, H.; Roth, S.; Wirth, E.K.; Culmsee, C.; Plesnila, N.; Kremmer, E.; Rådmark, O.; Wurst, W.; et al. Glutathione Peroxidase 4 Senses and Translates Oxidative Stress into 12/15-Lipoxygenase Dependent- and AIF-Mediated Cell Death. Cell Metab. 2008, 8, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Yoo, M.-H.; Gu, X.; Xu, X.-M.; Kim, J.-Y.; Carlson, B.A.; Patterson, A.D.; Cai, H.; Gladyshev, V.N.; Hatfield, D.L. Delineating the Role of Glutathione Peroxidase 4 in Protecting Cells Against Lipid Hydroperoxide Damage and in Alzheimer’s Disease. Antioxid. Redox Signal. 2010, 12, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Dammeyer, P.; Damdimopoulos, A.E.; Nordman, T.; Jiménez, A.; Miranda-Vizuete, A.; Arnér, E.S.J. Induction of Cell Membrane Protrusions by the N-terminal Glutaredoxin Domain of a Rare Splice Variant of Human Thioredoxin Reductase 1. J. Biol. Chem. 2008, 283, 2814–2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.A.; Ludueña, R.F. Possible regulation of the in vitro assembly of bovine brain tubulin by the bovine thioredoxin system. Biochim. Biophys. Acta (BBA)/Protein Struct. Mol. 1991, 1076, 289–297. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Koizumi, S.; Ohno, Y.; Nagao, T.; Inoue, K. Extracellular ATP counteracts the ERK1/2-mediated death-promoting signaling cascades in astrocytes. Glia 2006, 54, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Hellfritsch, J.; Kirsch, J.; Schneider, M.; Fluege, T.; Wortmann, M.; Frijhoff, J.; Dagnell, M.; Fey, T.; Esposito, I.; Koelle, P.; et al. Knockout of Mitochondrial Thioredoxin Reductase Stabilizes Prolyl Hydroxylase 2 and Inhibits Tumor Growth and Tumor-Derived Angiogenesis. Antioxid. Redox Signal. 2015, 22, 938–950. [Google Scholar] [CrossRef]
- Kondo, N.; Nakamura, H.; Masutani, H.; Yodoi, J. Redox Regulation of Human Thioredoxin Network. Antioxid. Redox Signal. 2006, 8, 1881–1890. [Google Scholar] [CrossRef]
- Arnér, E.S.J. Focus on mammalian thioredoxin reductases—Important selenoproteins with versatile functions. Biochim. Biophys. Acta-Gen. Subj. 2009, 1790, 495–526. [Google Scholar] [CrossRef]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Gladyshev, V.N.; Stadtman, T.C.; Hatfield, D.L.; Jeang, K.-T. Levels of major selenoproteins in T cells decrease during HIV infection and low molecular mass selenium compounds increase. Proc. Natl. Acad. Sci. USA 1999, 96, 835–839. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Arnér, E.S.J.; Ljung, J.; Åslund, F.; Holmgren, A. Rat and Calf Thioredoxin Reductase Are Homologous to Glutathione Reductase with a Carboxyl-terminal Elongation Containing a Conserved Catalytically Active Penultimate Selenocysteine Residue. J. Biol. Chem. 1998, 273, 8581–8591. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.H.; David Arscott, L.; Müller, S.; Lennon, B.W.; Ludwig, M.L.; Wang, P.F.; Veine, D.M.; Becker, K.; Heiner Schirmer, R. Thioredoxin Reductase: Two Modes of Catalysis Have Evolved. Eur. J. Biochem. 2000, 267, 6110–6117. [Google Scholar] [CrossRef] [Green Version]
- Snider, G.W.; Dustin, C.M.; Ruggles, E.L.; Hondal, R.J. A Mechanistic Investigation of the C-Terminal Redox Motif of Thioredoxin Reductase from Plasmodium falciparum. Biochemistry 2014, 53, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Carlson, B.A.; Labunskyy, V.M.; Gladyshev, V.N.; Hatfield, D.L. Selenoprotein T deficiency alters cell adhesion and elevates selenoprotein W expression in murine fibroblast cells. Biochem. Cell Biol. 2009, 87, 953–961. [Google Scholar] [CrossRef]
- Boukhzar, L.; Hamieh, A.; Cartier, D.; Tanguy, Y.; Alsharif, I.; Castex, M.; Arabo, A.; El Hajji, S.; Bonnet, J.-J.; Errami, M.; et al. Selenoprotein T Exerts an Essential Oxidoreductase Activity That Protects Dopaminergic Neurons in Mouse Models of Parkinson’s Disease. Antioxid. Redox Signal. 2016, 24, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Chivers, P.T.; Laboissière, M.C.; Raines, R.T. The CXXC motif: Imperatives for the formation of native disulfide bonds in the cell. EMBO J. 1996, 15, 2659–2667. [Google Scholar] [CrossRef]
- Gobler, C.J.; Berry, D.L.; Dyhrman, S.T.; Wilhelm, S.W.; Salamov, A.; Lobanov, A.V.; Zhang, Y.; Collier, J.L.; Wurch, L.L.; Kustka, A.B.; et al. Niche of harmful alga Aureococcus anophagefferens revealed through ecogenomics. Proc. Natl. Acad. Sci. USA 2011, 108, 4352–4357. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Selenoprotein (Containing Sec Residue) | Functions | References |
|---|---|---|
| Glutathione peroxidase 1 (GPX 1) | As an antioxidant, also functions as Se storage house | [63,64] |
| GPX 2 | As an antioxidant, anti-apoptotic function in the colon regulates mucosal homeostasis | [65] |
| GPX 3 | As an antioxidant, preventing plasma LDL oxidation, functions in the reduction of H2O2 | [66,67,68] |
| GPX 4 | As an antioxidant protects brain membranes from peroxidative degradation, catalyzes the reduction of hydroperoxides, inhibits lipid peroxidation | [69,70,71,72] |
| GPX 6 | Unknown | - |
| Thioredoxin reductases 1 (TrxR1) | As an antioxidant, reduction of thioredoxin, actin polymerization for cell membrane restructuring | [37,73] |
| TrxR2 | Regulation of mitochondrial redox homeostasis, Maintains thioredoxin in a reduced state | [37] |
| TrxR3 | Unknown | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghuge, S.A.; Kadam, U.S.; Hong, J.C. Selenoprotein: Potential Player in Redox Regulation in Chlamydomonas reinhardtii. Antioxidants 2022, 11, 1630. https://doi.org/10.3390/antiox11081630
Ghuge SA, Kadam US, Hong JC. Selenoprotein: Potential Player in Redox Regulation in Chlamydomonas reinhardtii. Antioxidants. 2022; 11(8):1630. https://doi.org/10.3390/antiox11081630
Chicago/Turabian StyleGhuge, Sandip A., Ulhas Sopanrao Kadam, and Jong Chan Hong. 2022. "Selenoprotein: Potential Player in Redox Regulation in Chlamydomonas reinhardtii" Antioxidants 11, no. 8: 1630. https://doi.org/10.3390/antiox11081630