
Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response

 , , , ,
, , , ,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Database Searches, Sequence Retrieval and Analysis of Physical Properties
2.2. Classification of APX and CAT Genes of Soybeans and Paralogous Gene Pair Prediction
2.3. Intron–Exon and Motif Analysis of APX and CAT Genes
2.4. Promoter Analysis for Cis-Acting Elements
2.5. Chromosomal Localization and Gene Duplication Analysis
2.6. Determining Gene Duplication Event of Paralogous CAT and APX Genes
2.7. Comparative Phylogenetic Analysis
2.8. Comparative Analysis of Ortholog Gene Pairs Identified in Soybeans
2.9. Expression Pattern Analysis from Fourteen Soybean Tissues
2.10. Plant Materials, Stress Treatments and Tissue Sampling
2.11. RNA Extraction and Quantitative Real-Time PCR (RT-qPCR)
3. Results
3.1. Identification of APX and CAT Subfamily Members in Soybeans
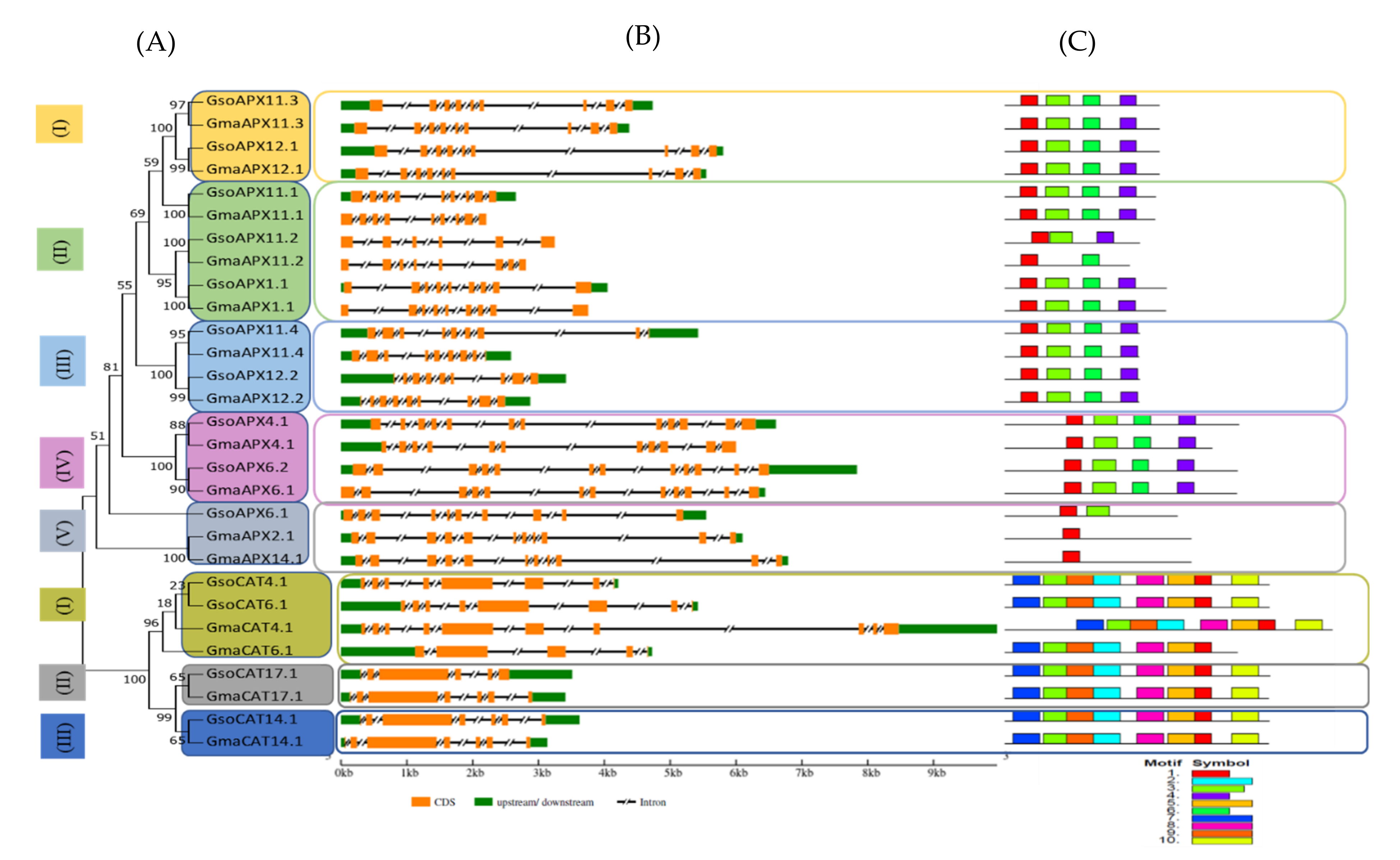
3.2. Phylogenetic Tree and Gene Structure Analysis of APX and CAT Proteins
3.3. Conserved Domain Detection in APX and CAT Proteins
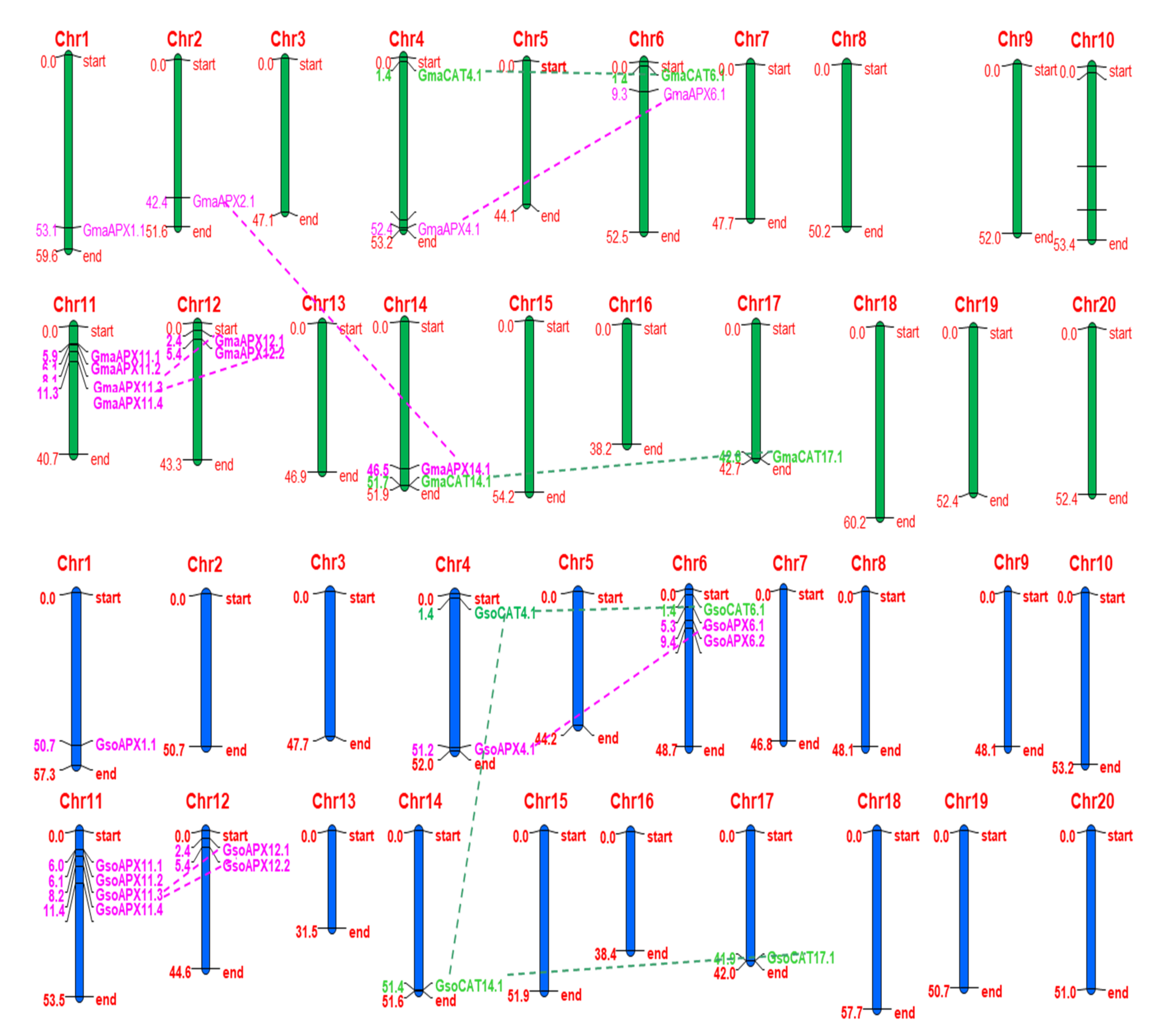
3.4. Chromosome Localization and Gene Duplication of APX and CAT Genes
3.5. Estimation of Divergence Time of G. max and G. soja APX and CAT PGPs
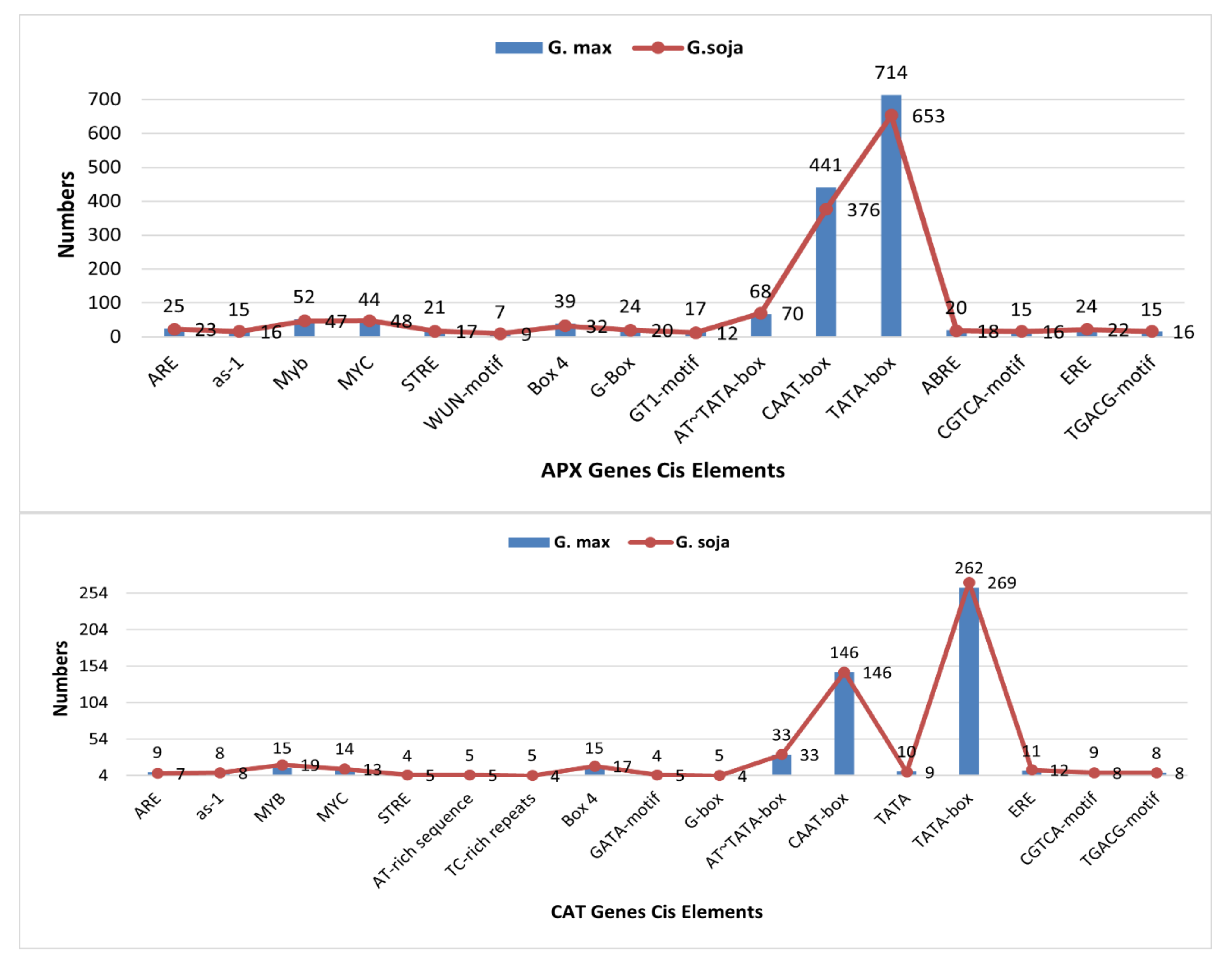
3.6. Cis Regulatory Elements
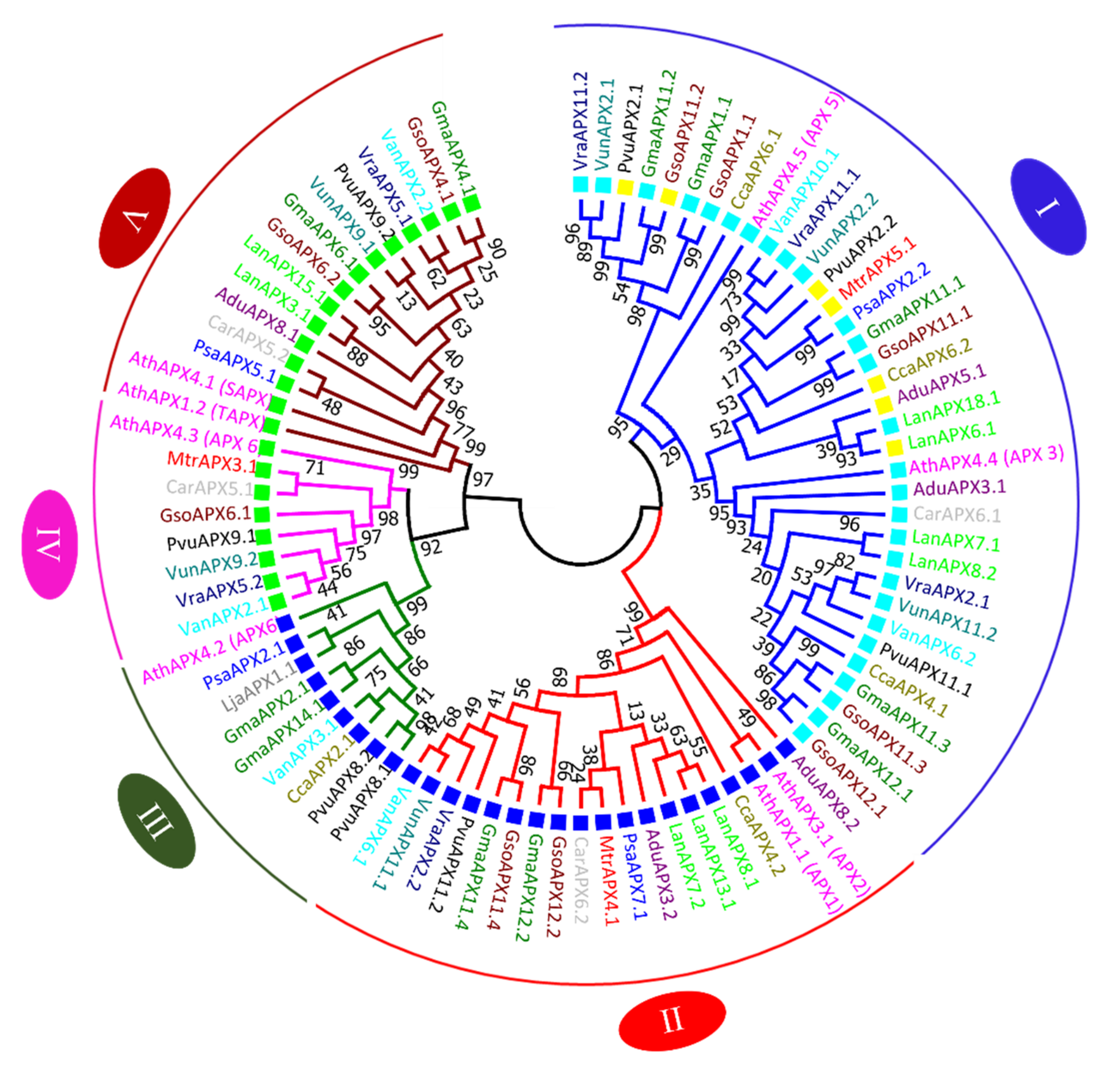
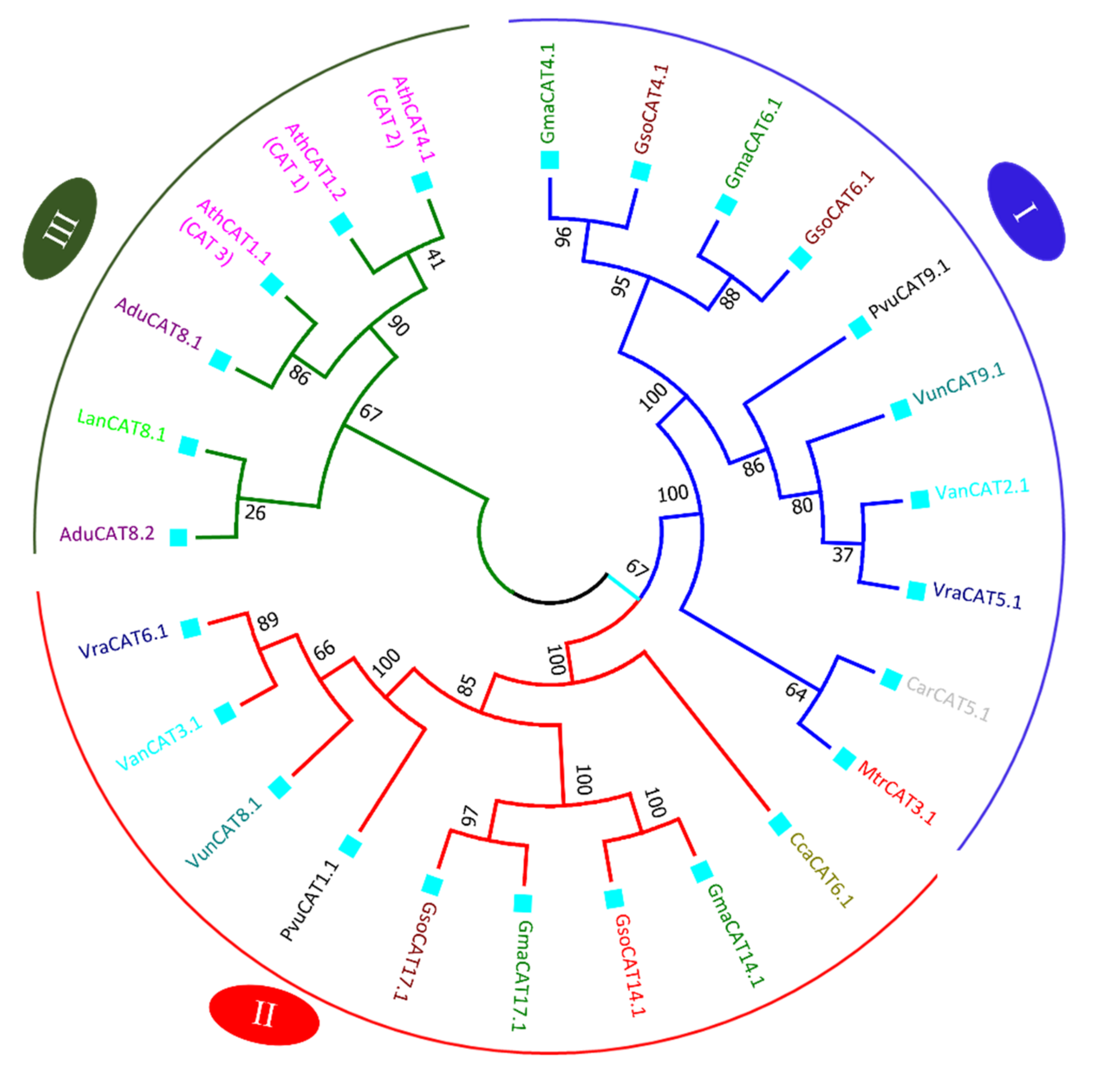
3.7. APX and CAT Genes Comparative Relationship from G. max and G. soja with Other Legumes and Arabidopsis thaliana
3.8. Comparison of Copy Number of G. max APX and CAT Genes with G. soja
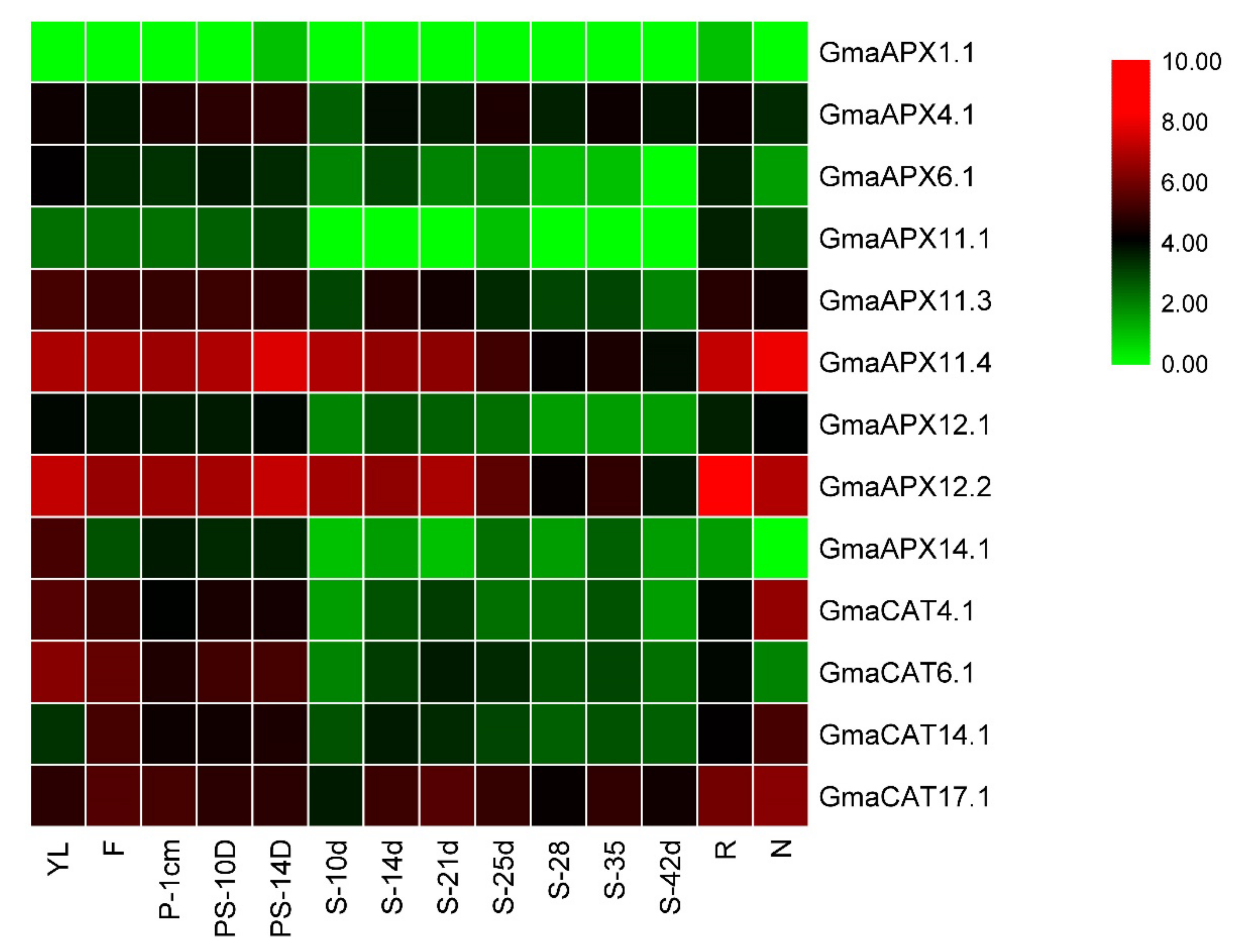
3.9. Expression Pattern of APXs and CATs in different Underground and Aerial Tissues of G. max Science Identifiers
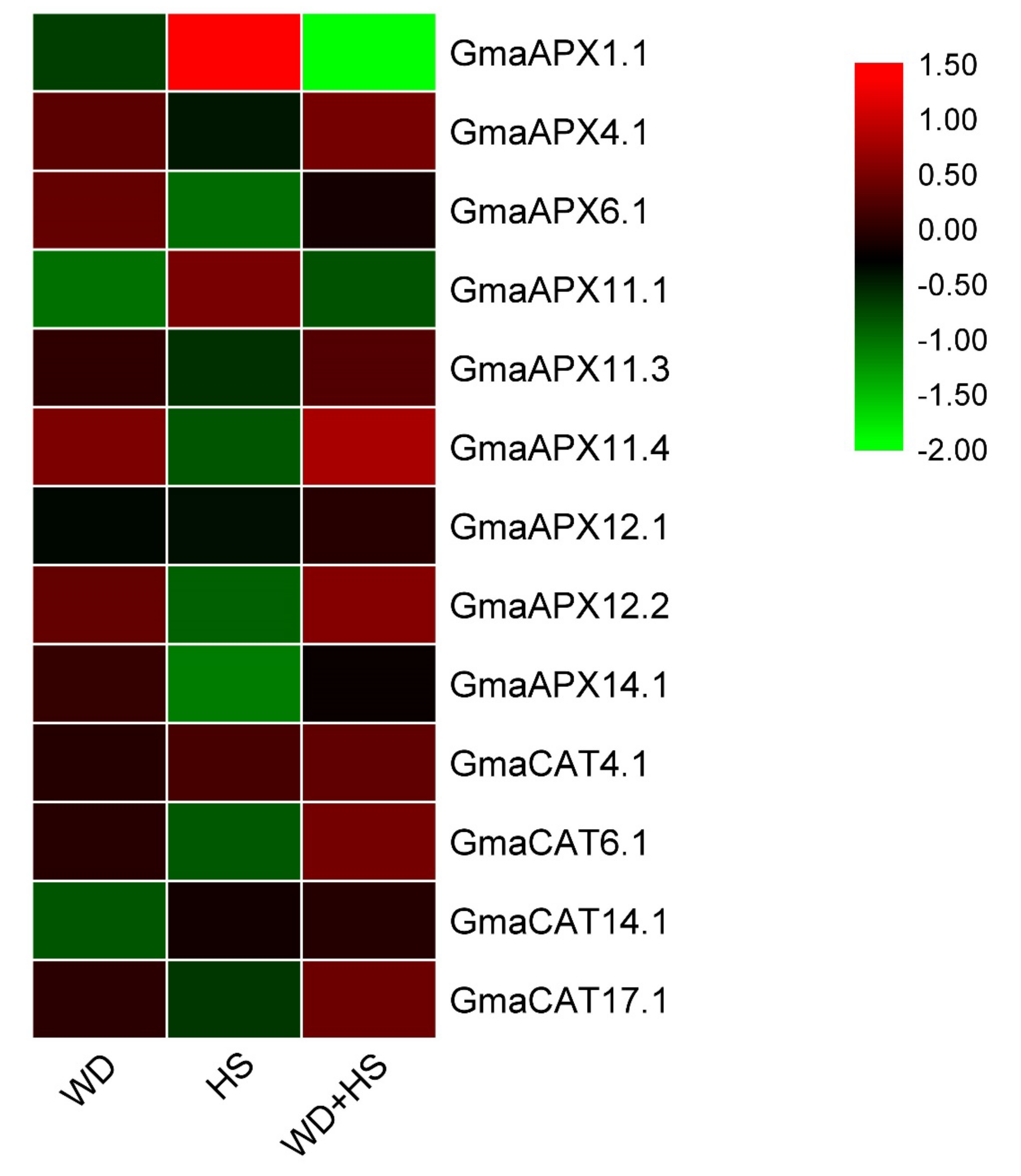
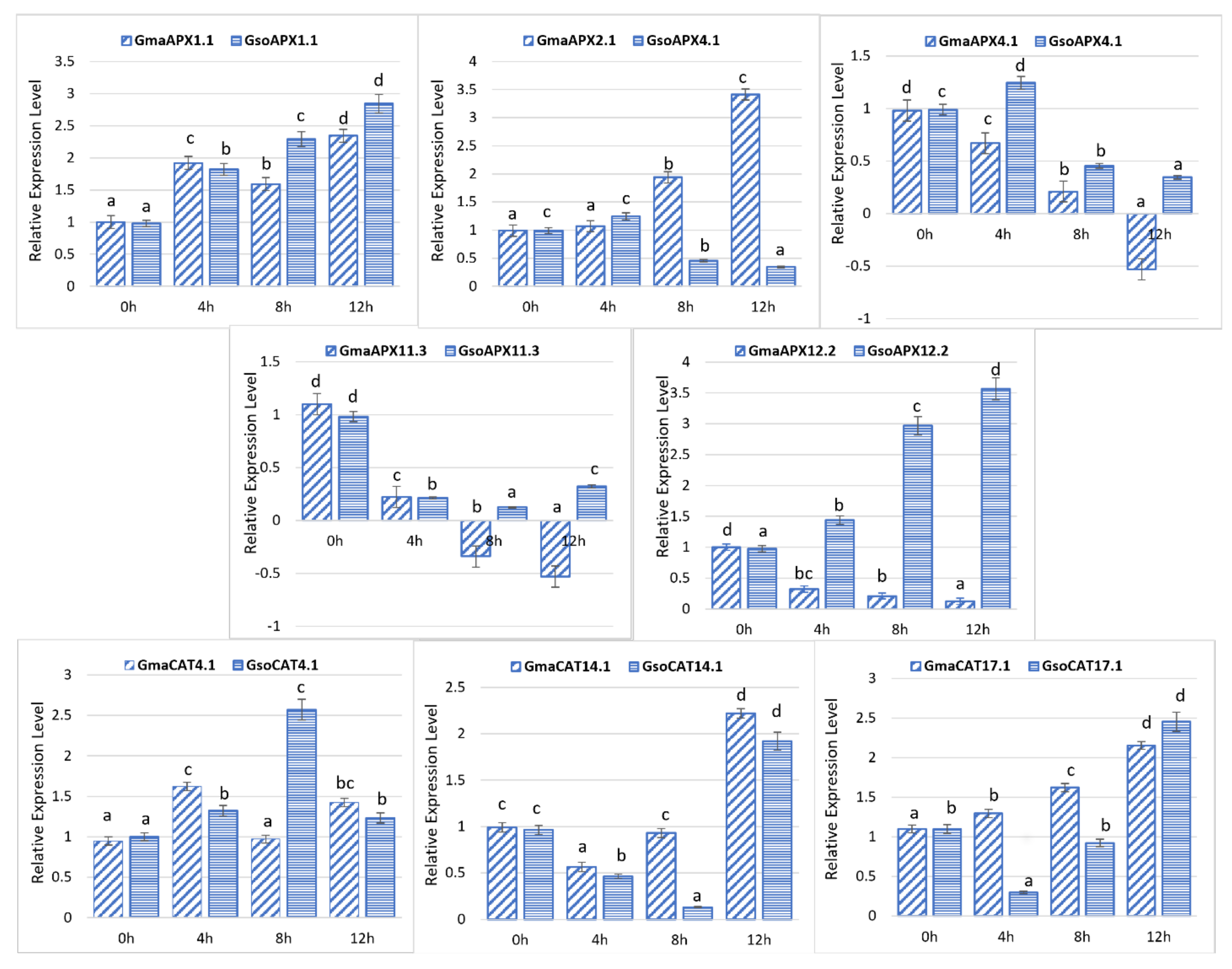
3.10. Response of G. soja and G. max APX and CAT Genes to Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jamil, S.; Shahzad, R.; Ahmad, S.; Fatima, R.; Zahid, R.; Anwar, M.; Iqbal, M.Z.; Wang, X. Role of Genetics, Genomics and Breeding approaches to combat stripe rust of wheat. Front. Nutr. 2020, 7, 173. [Google Scholar]
- Ahmad, S.; Tang, L.; Shahzad, R.; Mawia, A.M.; Rao, G.S.; Jamil, S.; Wei, C.; Sheng, Z.; Shao, G.; Wei, X. CRISPR-based crop improvements: A way forward to achieve zero hunger. J. Agric. Food Chem. 2021, 69, 8307–8323. [Google Scholar]
- Shahzad, R.; Shakra Jamil, S.A.; Nisar, A.; Amina, Z.; Saleem, S.; Iqbal, M.Z.; Atif, R.M.; Wang, X. Harnessing the potential of plant transcription factors in developing climate resilient crops to improve global food security: Current and future perspectives. Saudi J. Biol. Sci. 2021, 28, 2323. [Google Scholar] [PubMed]
- Jamil, S.; Shahzad, R.; Ahmad, S.; Ali, Z.; Shaheen, S.; Shahzadee, H.; Fatima, N.; Sharif, I.; Aleem, M.; Sultana, R. Climate Change and Role of Genetics and Genomics in Climate-Resilient Sorghum. In Developing Climate-Resilient Crops; Fahad, S., Sonmez, O., Saud, S., Wang, D., Wu, C., Adnan, M., Turan, V., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 111–138. [Google Scholar]
- Gupta, S.; Dong, Y.; Dijkwel, P.P.; Mueller-Roeber, B.; Gechev, T.S. Genome-wide analysis of ROS antioxidant genes in resurrection species suggest an involvement of distinct ROS detoxification systems during desiccation. Int. J. Mol. Sci. 2019, 20, 3101. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar]
- Kuluev, B.; Mikhaylova, E.; Berezhneva, Z.; Nikonorov, Y.; Postrigan, B.; Kudoyarova, G.; Chemeris, A. Expression profiles and hormonal regulation of tobacco NtEXGT gene and its involvement in abiotic stress response. Plant Physiol. Biochem. 2017, 111, 203–215. [Google Scholar]
- Gill, S.S.; Khan, N.A.; Anjum, N.A.; Tuteja, N. Amelioration of cadmium stress in crop plants by nutrients management: Morphological, physiological and biochemical aspects. Plant Stress 2011, 5, 1–23. [Google Scholar]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar]
- Mittler, R.; Zilinskas, B.A. Purification and characterization of pea cytosolic ascorbate peroxidase. Plant Physiol. 1991, 97, 962–968. [Google Scholar]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [PubMed] [Green Version]
- Teixeira, F.K.; Menezes-Benavente, L.; Margis, R.; Margis-Pinheiro, M. Analysis of the molecular evolutionary history of the ascorbate peroxidase gene family: Inferences from the rice genome. J. Mol. Evol. 2004, 59, 761–770. [Google Scholar] [PubMed]
- Najami, N.; Janda, T.; Barriah, W.; Kayam, G.; Tal, M.; Guy, M.; Volokita, M. Ascorbate peroxidase gene family in tomato: Its identification and characterization. Mol. Genet. Genom. 2008, 279, 171–182. [Google Scholar]
- Tao, C.; Jin, X.; Zhu, L.; Xie, Q.; Wang, X.; Li, H. Genome-wide investigation and expression profiling of APX gene family in Gossypium hirsutum provide new insights in redox homeostasis maintenance during different fiber development stages. Mol. Genet. Genom. 2018, 293, 685–697. [Google Scholar]
- Akbudak, M.A.; Filiz, E.; Vatansever, R.; Kontbay, K. Genome-Wide Identification and Expression Profiling of Ascorbate Peroxidase (APX) and Glutathione Peroxidase (GPX) Genes Under Drought Stress in Sorghum (Sorghum bicolor L.). J. Plant Growth Regul. 2018, 37, 925–936. [Google Scholar] [CrossRef]
- Panchuk, I.I.; Zentgraf, U.; Volkov, R.A. Expression of the Apx gene family during leaf senescence of Arabidopsis thaliana. Planta 2005, 222, 926–932. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.-J.; Jana, C.K.; Das, N. Role of catalase in oxidative stress-and age-associated degenerative diseases. Oxidative Med. Cell. Longev. 2019, 2019, 9613090. [Google Scholar]
- Sharma, I.; Ahmad, P. Catalase: A versatile antioxidant in plants. In Oxidative Damage to Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 131–148. [Google Scholar]
- Loewen, P.C.; Klotz, M.G.; Hassett, D.J. Catalase—An “old” enzyme that continues to surprise us. ASM News 2000, 66, 76–82. [Google Scholar]
- Chen, H.-J.; Wu, S.-D.; Huang, G.-J.; Shen, C.-Y.; Afiyanti, M.; Li, W.-J.; Lin, Y.-H. Expression of a cloned sweet potato catalase SPCAT1 alleviates ethephon-mediated leaf senescence and H2O2 elevation. J. Plant Physiol. 2012, 169, 86–97. [Google Scholar]
- GONZÁLEZ, E. The C-terminal domain of plant catalases Implications for a glyoxysomal targeting sequence. Eur. J. Biochem. 1991, 199, 211–215. [Google Scholar]
- Skadsen, R.W.; Schulze-Lefert, P.; Herbst, J.M. Molecular cloning, characterization and expression analysis of two catalase isozyme genes in barley. Plant Mol. Biol. 1995, 29, 1005–1014. [Google Scholar] [PubMed]
- Niewiadomska, E.; Polzien, L.; Desel, C.; Rozpadek, P.; Miszalski, Z.; Krupinska, K. Spatial patterns of senescence and development-dependent distribution of reactive oxygen species in tobacco (Nicotiana tabacum) leaves. J. Plant Physiol. 2009, 166, 1057–1068. [Google Scholar] [PubMed]
- Du, Y.Y.; Wang, P.C.; Chen, J.; Song, C.P. Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [PubMed]
- Guan, L.; Scandalios, J.G. Developmentally related responses of maize catalase genes to salicylic acid. Proc. Natl. Acad. Sci. USA 1995, 92, 5930–5934. [Google Scholar]
- Iwamoto, M.; Higo, H.; Higo, K. Differential diurnal expression of rice catalase genes: The 5′-flanking region of CatA is not sufficient for circadian control. Plant Sci. 2000, 151, 39–46. [Google Scholar]
- Hu, L.; Yang, Y.; Jiang, L.; Liu, S. The catalase gene family in cucumber: Genome-wide identification and organization. Genet. Mol. Biol. 2016, 39, 408–415. [Google Scholar]
- Wang, W.; Cheng, Y.; Chen, D.; Liu, D.; Hu, M.; Dong, J.; Zhang, X.; Song, L.; Shen, F. The catalase gene family in cotton: Genome-wide characterization and bioinformatics analysis. Cells 2019, 8, 86. [Google Scholar]
- Queval, G.; Issakidis-Bourguet, E.; Hoeberichts, F.A.; Vandorpe, M.; Gakière, B.; Vanacker, H.; Miginiac-Maslow, M.; Van Breusegem, F.; Noctor, G. Conditional oxidative stress responses in the Arabidopsis photorespiratory mutant cat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression, and define photoperiod as a crucial factor in the regulation of H2O2-induced cell death. Plant J. 2007, 52, 640–657. [Google Scholar]
- Joshi, T.; Valliyodan, B.; Wu, J.-H.; Lee, S.-H.; Xu, D.; Nguyen, H.T. Genomic differences between cultivated soybean, G. max and its wild relative G. soja. BMC Genom. 2013, 14, 1–11. [Google Scholar]
- Manavalan, L.P.; Guttikonda, S.K.; Phan Tran, L.-S.; Nguyen, H.T. Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol. 2009, 50, 1260–1276. [Google Scholar]
- Wang, L.; Liu, L.; Ma, Y.; Li, S.; Dong, S.; Zu, W. Transcriptome profilling analysis characterized the gene expression patterns responded to combined drought and heat stresses in soybean. Comput. Biol. Chem. 2018, 77, 413–429. [Google Scholar] [PubMed]
- Zhang, G.; Chen, M.; Chen, X.; Xu, Z.; Guan, S.; Li, L.-C.; Li, A.; Guo, J.; Mao, L.; Ma, Y. Phylogeny, gene structures, and expression patterns of the ERF gene family in soybean (Glycine max L.). J. Exp. Bot. 2008, 59, 4095–4107. [Google Scholar] [PubMed] [Green Version]
- Yin, G.; Xu, H.; Xiao, S.; Qin, Y.; Li, Y.; Yan, Y.; Hu, Y. The large soybean (Glycine max) WRKY TF family expanded by segmental duplication events and subsequent divergent selection among subgroups. BMC Plant Biol. 2013, 13, 148. [Google Scholar]
- Xu, H.; Li, Y.; Yan, Y.; Wang, K.; Gao, Y.; Hu, Y. Genome-scale identification of soybean BURP domain-containing genes and their expression under stress treatments. BMC Plant Biol. 2010, 10, 197. [Google Scholar]
- Fan, C.-M.; Wang, X.; Wang, Y.-W.; Hu, R.-B.; Zhang, X.-M.; Chen, J.-X.; Fu, Y.-F. Genome-wide expression analysis of soybean MADS genes showing potential function in the seed development. PLoS ONE 2013, 8, e62288. [Google Scholar]
- Du, H.; Yang, S.-S.; Liang, Z.; Feng, B.-R.; Liu, L.; Huang, Y.-B.; Tang, Y.-X. Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC Plant Biol. 2012, 12, 106. [Google Scholar]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar]
- Ma, T.; Wang, J.; Zhou, G.; Yue, Z.; Hu, Q.; Chen, Y.; Liu, B.; Qiu, Q.; Wang, Z.; Zhang, J. Genomic insights into salt adaptation in a desert poplar. Nat. Commun. 2013, 4, 2797. [Google Scholar]
- Ozyigit, I.I.; Filiz, E.; Vatansever, R.; Kurtoglu, K.Y.; Koc, I.; Ozturk, M.X.; Anjum, N.A. Identification and Comparative Analysis of H2O2-Scavenging Enzymes (Ascorbate peroxidase and Glutathione peroxidase) in Selected Plants Employing Bioinformatics Approaches. Front. Plant Sci. 2016, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.; Copley, R.R.; Doerks, T.; Ponting, C.P.; Bork, P. SMART: A web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 571–607. [Google Scholar]
- Chi, S.-M.; Nam, D. WegoLoc: Accurate prediction of protein subcellular localization using weighted Gene Ontology terms. Bioinformatics 2012, 28, 1028–1030. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Zhang, J. Evolutionary Distance: Estimation. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd: Chichester, UK, 2005. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Mifsud, W.; Bateman, A. Membrane-bound progesterone receptors contain a cytochrome b 5-like ligand-binding domain. Genome Biol. 2002, 3, 1–5. [Google Scholar] [CrossRef]
- Korber, B. HIV signature and sequence variation analysis. In Computational Analysis of HIV Molecular Sequences; Rodrigo, A.G., Jr., Learn, G.H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 55–74. [Google Scholar]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Du, H.; Liu, Y.; Ni, L.; Wang, Z.; Liang, C.; Tian, Z. Update soybean Zhonghuang 13 genome to a golden reference. Sci. China Life Sci. 2019, 62, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Zandalinas, S.I.; Fritschi, F.B.; Sengupta, S.; Fichman, Y.; Azad, R.K.; Mittler, R. The impact of water deficit and heat stress combination on the molecular response, physiology, and seed production of soybean. Physiol. Plant. 2021, 172, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.; Riaz, A.; Raza, Q.; Aleem, M.; Aslam, M.; Kong, K.; Atif, R.M.; Kashif, M.; Bhat, J.A.; Zhao, T. Genome-wide characterization and functional analysis of class III peroxidase gene family in soybean reveal regulatory roles of GsPOD40 in drought tolerance. Genomics 2022, 114, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ahmed, S.; Rashid, M.A.R.; Zafar, S.A.; Azhar, M.T.; Waqas, M.; Uzair, M.; Rana, I.A.; Azeem, F.; Chung, G.; Ali, Z. Genome-wide investigation and expression analysis of APETALA-2 transcription factor subfamily reveals its evolution, expansion and regulatory role in abiotic stress responses in Indica Rice (Oryza sativa L. ssp. indica). Genomics 2021, 113, 1029–1043. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, S.; Van, K.; Kim, T.-H.; Jeong, S.-C.; Choi, I.-Y.; Kim, D.-S.; Lee, Y.-S.; Park, D.; Ma, J. Whole-genome sequencing and intensive analysis of the undomesticated soybean (Glycine soja Sieb. and Zucc.) genome. Proc. Natl. Acad. Sci. USA 2010, 107, 22032–22037. [Google Scholar] [CrossRef] [Green Version]
- Konishi, S.; Izawa, T.; Lin, S.Y.; Ebana, K.; Fukuta, Y.; Sasaki, T.; Yano, M. An SNP caused loss of seed shattering during rice domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Pfeiffer, T.W.; Cornelius, P.L. Soybean QTL for yield and yield components associated with Glycine soja alleles. Crop Sci. 2008, 48, 571–581. [Google Scholar]
- Qin, Y.-M.; Hu, C.-Y.; Zhu, Y.-X. The ascorbate peroxidase regulated by H2O2 and ethylene is involved in cotton fiber cell elongation by modulating ROS homeostasis. Plant Signal. Behav. 2008, 3, 194–196. [Google Scholar] [CrossRef] [Green Version]
- Nan, H.; Li, W.; Lin, Y.-l.; Gao, L.-z. Genome-wide analysis of WRKY genes and their response to salt stress in the wild progenitor of Asian cultivated rice, Oryza rufipogon. Front. Genet. 2020, 11, 359. [Google Scholar] [CrossRef]
- Jia, Q.; Xiao, Z.-X.; Wong, F.-L.; Sun, S.; Liang, K.-J.; Lam, H.-M. Genome-wide analyses of the soybean F-box gene family in response to salt stress. Int. J. Mol. Sci. 2017, 18, 818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, S.U.; Qanmber, G.; Tahir, M.H.N.; Irshad, A.; Fiaz, S.; Ahmad, F.; Ali, Z.; Sajjad, M.; Shees, M.; Usman, M. Characterization of Vascular plant One-Zinc finger (VOZ) in soybean (Glycine max and Glycine soja) and their expression analyses under drought condition. PLoS ONE 2021, 16, e0253836. [Google Scholar] [CrossRef]
- Mir, Z.A.; Ali, S.; Shivaraj, S.; Bhat, J.A.; Singh, A.; Yadav, P.; Rawat, S.; Paplao, P.K.; Grover, A. Genome-wide identification and characterization of Chitinase gene family in Brassica juncea and Camelina sativa in response to Alternaria brassicae. Genomics 2020, 112, 749–763. [Google Scholar] [CrossRef]
- Qu, C.; Wang, L.; Zhao, Y.; Liu, C. Molecular Evolution of Maize Ascorbate Peroxidase Genes and Their Functional Divergence. Genes 2020, 11, 1204. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Yu, H.; Cao, P.B.; Fawal, N.; Mathé, C.; Azar, S.; Cassan-Wang, H.; Myburg, A.A.; Grima-Pettenati, J.; Marque, C. Explosive tandem and segmental duplications of multigenic families in Eucalyptus grandis. Genome Biol. Evol. 2015, 7, 1068–1081. [Google Scholar] [CrossRef] [Green Version]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Starr, T.K.; Jameson, S.C.; Hogquist, K.A. Positive and negative selection of T cells. Annu. Rev. Immunol. 2003, 21, 139–176. [Google Scholar] [CrossRef]
- Zhang, J.; Li, B.; Yang, Y.; Hu, W.; Chen, F.; Xie, L.; Fan, L. Genome-wide characterization and expression profiles of the superoxide dismutase gene family in Gossypium. Int. J. Genom. 2016, 1–11. [Google Scholar]
- Zhang, J.; Rosenberg, H.F.; Nei, M. Positive Darwinian selection after gene duplication in primate ribonuclease genes. Proc. Natl. Acad. Sci. USA 1998, 95, 3708–3713. [Google Scholar] [CrossRef] [Green Version]
- Leng, X.; Wang, H.; Zhang, S.; Qu, C.; Yang, C.; Xu, Z.; Liu, G. Identification and Characterization of the APX Gene Family and Its Expression Pattern under Phytohormone Treatment and Abiotic Stress in Populus trichocarpa. Genes 2021, 12, 334. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleem, M.; Raza, M.M.; Haider, M.S.; Atif, R.M.; Ali, Z.; Bhat, J.A.; Zhao, T. Comprehensive RNA-seq analysis revealed molecular pathways and genes associated with drought tolerance in wild soybean (Glycine soja Sieb. and Zucc.). Physiol. Plant. 2021, 172, 707–732. [Google Scholar] [CrossRef]
- Harb, A.; Awad, D.; Samarah, N. Gene expression and activity of antioxidant enzymes in barley (Hordeum vulgare L.) under controlled severe drought. J. Plant Interact. 2015, 10, 109–116. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soybase ID | Rename ID | Protein Domain Family (PFAM) | Chrom. No. | Gene Starting Position | Gene End Position | Orientation | Protein Length | MW (Kda) | Theor. Pi | Localization Cello | Localization Wegoloc |
|---|---|---|---|---|---|---|---|---|---|---|---|
| G. max | |||||||||||
| glyma.Zh13.gnm1.ann1.SoyZH13_01G148100 | GmaAPX1.1 | PF00041 | 1 | 53076021 | 53079776 | + | 300 | 33.81 | 9.05 | Nuclear | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_02G190600 | GmaAPX2.1 | PF00041 | 2 | 42405706 | 42411803 | - | 347 | 37.75 | 6.24 | Extracellular | Chloroplast |
| glyma.Zh13.gnm1.ann1.SoyZH13_04G223500 | GmaAPX4.1 | PF00041 | 4 | 52362952 | 52368952 | - | 386 | 41.95 | 6.73 | Chloroplast | Chloroplast |
| glyma.Zh13.gnm1.ann1.SoyZH13_06G109600 | GmaAPX6.1 | PF00041 | 6 | 9291175 | 9297616 | + | 432 | 46.98 | 7.13 | Chloroplast | Chloroplast |
| glyma.Zh13.gnm1.ann1.SoyZH13_11G076100 | GmaAPX11.1 | PF00041 | 11 | 5894217 | 5896422 | - | 280 | 31.18 | 9.08 | Cytoplasmic | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_11G078900 | GmaAPX11.2 | PF00041 | 11 | 6066134 | 6068941 | + | 232 | 26.64 | 8.74 | Cytoplasmic | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_11G104300 | GmaAPX11.3 | PF00041 | 11 | 8138680 | 8143056 | - | 287 | 31.72 | 6.62 | Cytoplasmic | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_11G142000 | GmaAPX11.4 | PF00041 | 11 | 11329025 | 11331607 | + | 250 | 27.05 | 5.5 | Cytoplasmic | Cytoplasm |
| glyma.Zh13.gnm1.ann1.SoyZH13_12G031100 | GmaAPX12.1 | PF00041 | 12 | 2425213 | 2430761 | - | 287 | 31.76 | 6.27 | Cytoplasmic | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_12G067000 | GmaAPX12.2 | PF00041 | 12 | 5412074 | 5414947 | - | 250 | 27.13 | 5.65 | Cytoplasmic | Cytoplasm |
| glyma.Zh13.gnm1.ann1.SoyZH13_14G162900 | GmaAPX14.1 | PF00041 | 14 | 46547958 | 46554747 | - | 347 | 37.92 | 6.76 | Extracellular | Chloroplast |
| glyma.Zh13.gnm1.ann1.SoyZH13_04G016700 | GmaCAT4.1 | PF00199 | 4 | 1374434 | 1384397 | - | 611 | 69.88 | 6.61 | Peroxisomal | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_06G016600 | GmaCAT6.1 | PF00199 | 6 | 1350172 | 1354895 | - | 434 | 49.79 | 6.26 | Peroxisomal | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_14G205900 | GmaCAT14.1 | PF00199 | 14 | 51701327 | 51704461 | + | 492 | 56.91 | 6.77 | Peroxisomal | Peroxisome |
| glyma.Zh13.gnm1.ann1.SoyZH13_17G248900 | GmaCAT17.1 | PF00199 | 17 | 42607546 | 42610955 | + | 492 | 56.95 | 6.77 | Peroxisomal | Peroxisome |
| G. soja | |||||||||||
| glyso.W05.gnm1.ann1.Glysoja.01G001741 | GsoAPX1.1 | PF00041 | 1 | 50740356 | 50744403 | + | 300 | 33.8 | 9.05 | Nuclear | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.04G010747 | GsoAPX4.1 | PF00041 | 4 | 51176378 | 51182983 | - | 435 | 47.3 | 7.1 | Chloroplast | Chloroplast |
| glyso.W05.gnm1.ann1.Glysoja.06G013959 | GsoAPX6.1 | PF00041 | 6 | 5275075 | 5280621 | + | 321 | 34.4 | 7.56 | Chloroplast | Cytoplasm |
| glyso.W05.gnm1.ann1.Glysoja.06G014412 | GsoAPX6.2 | PF00041 | 6 | 9350953 | 9358789 | + | 432 | 47.1 | 7.13 | Chloroplast | Chloroplast |
| glyso.W05.gnm1.ann1.Glysoja.11G029301 | GsoAPX11.1 | PF00041 | 11 | 5950676 | 5953331 | - | 280 | 31.2 | 9.08 | Cytoplasmic | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.11G029327 | GsoAPX11.2 | PF00041 | 11 | 6122599 | 6125845 | + | 250 | 28.1 | 8.65 | Mitochondrial | Plasma Membrane |
| glyso.W05.gnm1.ann1.Glysoja.11G029583 | GsoAPX11.3 | PF00041 | 11 | 8214021 | 8218754 | - | 287 | 31.7 | 6.62 | Cytoplasmic | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.11G029970 | GsoAPX11.4 | PF00041 | 11 | 11420791 | 11423997 | + | 250 | 27.1 | 5.51 | Cytoplasmic | Cytoplasm |
| glyso.W05.gnm1.ann1.Glysoja.12G033246 | GsoAPX12.1 | PF00041 | 12 | 2440656 | 2446460 | - | 287 | 31.8 | 6.27 | Cytoplasmic | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.12G033638 | GsoAPX12.2 | PF00041 | 12 | 5431487 | 5434903 | - | 250 | 27.1 | 5.52 | Cytoplasmic | Cytoplasm |
| glyso.W05.gnm1.ann1.Glysoja.04G008496 | GsoCAT4.1 | PF00199 | 4 | 1367980 | 1372190 | - | 492 | 56.7 | 6.8 | Peroxisomal | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.06G013473 | GsoCAT6.1 | PF00199 | 6 | 1361430 | 1366850 | - | 492 | 56.7 | 6.77 | Peroxisomal | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.14G039529 | GsoCAT14.1 | PF00199 | 14 | 51391433 | 51395055 | + | 492 | 56.9 | 6.77 | Peroxisomal | Peroxisome |
| glyso.W05.gnm1.ann1.Glysoja.17G047079 | GsoCAT17.1 | PF00199 | 17 | 41904865 | 41908375 | + | 494 | 57.4 | 6.93 | Peroxisomal | Peroxisome |
| Duplicated Pair | Ka | Ks | Ka/Ks | Data (Mya) | Duplicate Type | Purifying Selection | Type |
|---|---|---|---|---|---|---|---|
| GmaAPX14.1/GmaAPX2.1 | 0.04 | 0.06 | 0.69 | 4.92 | Segmental | Yes | Negative or purifying selection |
| GmaAPX4.1/GmaAPX6.1 | 0.04 | 0.07 | 0.56 | 5.74 | Segmental | Yes | Negative or purifying selection |
| GmaAPX11.3/GmaAPX12.1 | 0.04 | 0.02 | 2.26 | 1.64 | Segmental | No | Positive selection |
| GmaAPX11.4/GmaAPX12.2 | 0.05 | 0.05 | 1.11 | 4.10 | Segmental | No | Positive selection |
| GsoAPX4.1/GsoAPX6.2 | 0.03 | 0.09 | 0.36 | 7.38 | Segmental | Yes | Negative or purifying selection |
| GsoAPX11.4/GsoAPX12.2 | 0.04 | 0.09 | 0.39 | 7.38 | Segmental | Yes | Negative or purifying selection |
| GsoAPX11.3/GsoAPX12.1 | 0.02 | 0.09 | 0.23 | 7.38 | Segmental | Yes | Negative or purifying selection |
| GmaCAT4.1/GmaCAT6.1 | 0.03 | 0.1 | 0.3 | 8.20 | Segmental | Yes | Negative or purifying selection |
| GmaCAT14.1/GmaCAT17.1 | 0.01 | 0.13 | 0.05 | 10.66 | Segmental | Yes | Negative or purifying selection |
| GsoCAT6.1/GsoCAT4.1 | 0.01 | 0.08 | 0.08 | 6.56 | Segmental | Yes | Negative or purifying selection |
| GsoCAT4.1/GsoCAT14.1 | 0.06 | 0.74 | 0.08 | 60.66 | Segmental | Yes | Negative or purifying selection |
| GsoCAT14.1/GsoCAT17.1 | 0.02 | 0.14 | 0.14 | 11.48 | Segmental | Yes | Negative or purifying selection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleem, M.; Aleem, S.; Sharif, I.; Aleem, M.; Shahzad, R.; Khan, M.I.; Batool, A.; Sarwar, G.; Farooq, J.; Iqbal, A.; et al. Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response. Antioxidants 2022, 11, 1626. https://doi.org/10.3390/antiox11081626
Aleem M, Aleem S, Sharif I, Aleem M, Shahzad R, Khan MI, Batool A, Sarwar G, Farooq J, Iqbal A, et al. Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response. Antioxidants. 2022; 11(8):1626. https://doi.org/10.3390/antiox11081626
Chicago/Turabian StyleAleem, Muqadas, Saba Aleem, Iram Sharif, Maida Aleem, Rahil Shahzad, Muhammad Imran Khan, Amina Batool, Gulam Sarwar, Jehanzeb Farooq, Azeem Iqbal, and et al. 2022. "Whole-Genome Identification of APX and CAT Gene Families in Cultivated and Wild Soybeans and Their Regulatory Function in Plant Development and Stress Response" Antioxidants 11, no. 8: 1626. https://doi.org/10.3390/antiox11081626