
Ameliorative Effects of Silicon against Salt Stress in Gossypium hirsutum L.


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Measurement Traits and Methods
2.2.1. Plant Growth and Biomass Accumulation
2.2.2. Measurement of Gas Exchange Parameters
2.2.3. Determination of ROS and Antioxidant Enzymes
2.2.4. Measurement of Photosynthetic Pigments
2.2.5. Assay of Chlorophyll Fluorescence Parameters
2.2.6. Activity of Ferredoxin-NADP Reductase (FNR), ATP Synthase and Ribulose-1, 5-Bisphosphate Carboxylase/Oxygenase (RubisCO)
2.2.7. Estimation of Stomatal State
2.2.8. Transmission Electron Microscope Assay for Chloroplast Ultrastructure
2.2.9. Quantitative Real-Time PCR Analysis
2.3. Statistical Analysis
3. Results
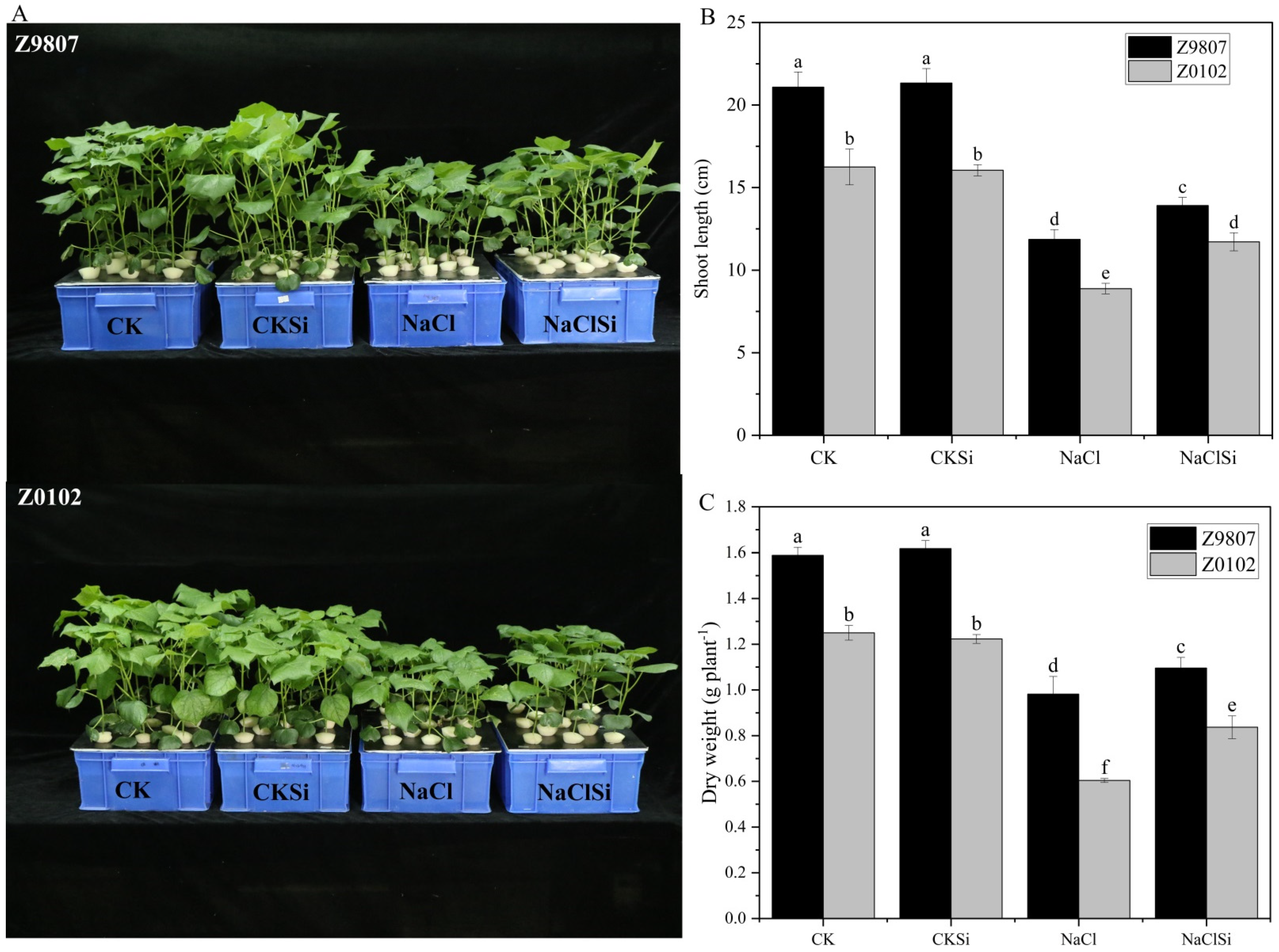
3.1. Effect of Exogenous Si on the Growth of Cotton Seedlings under Salt Stress Subsection
3.2. Regulation of Gas Exchange Parameters upon Exogenous Silicon
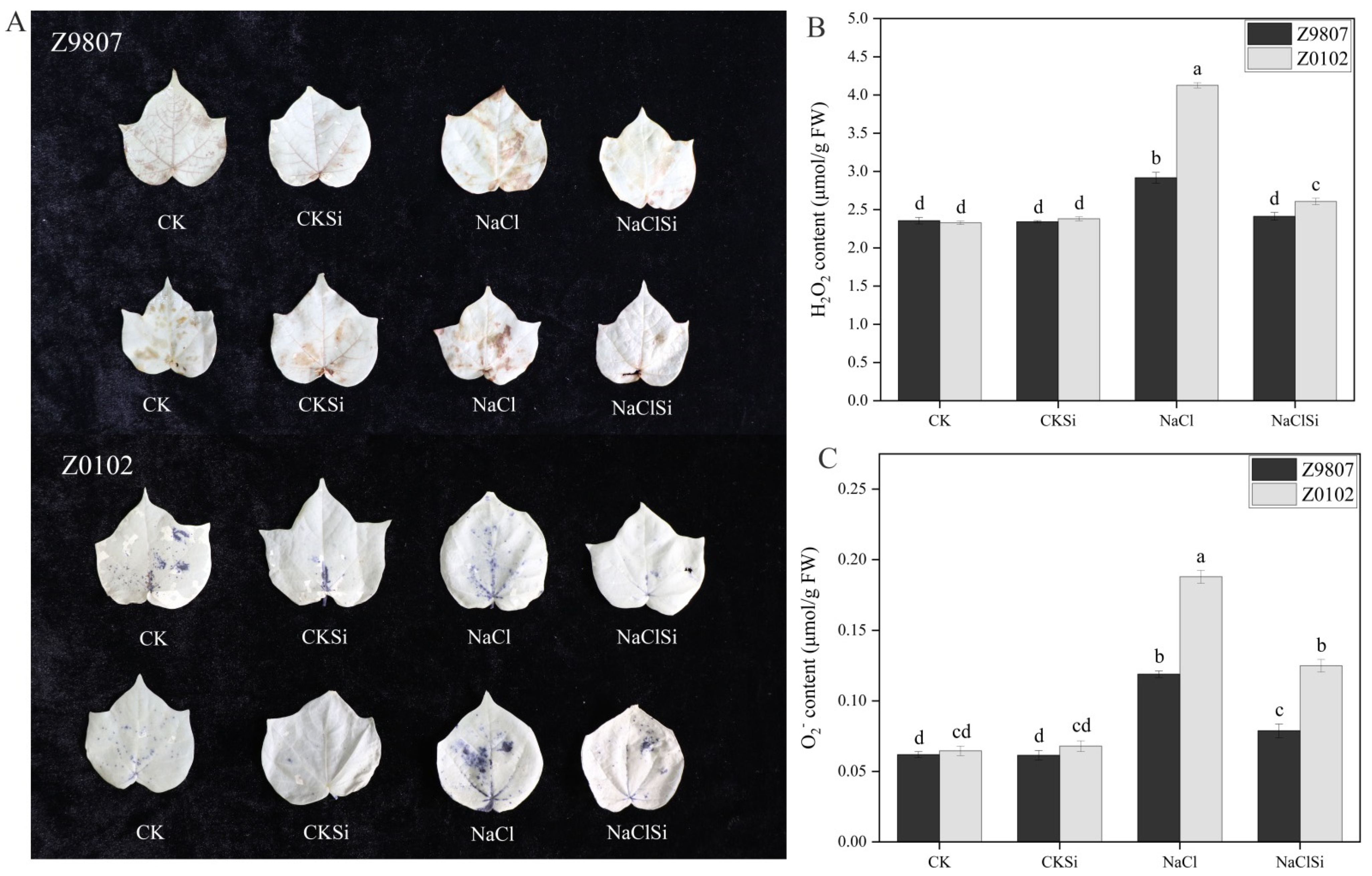
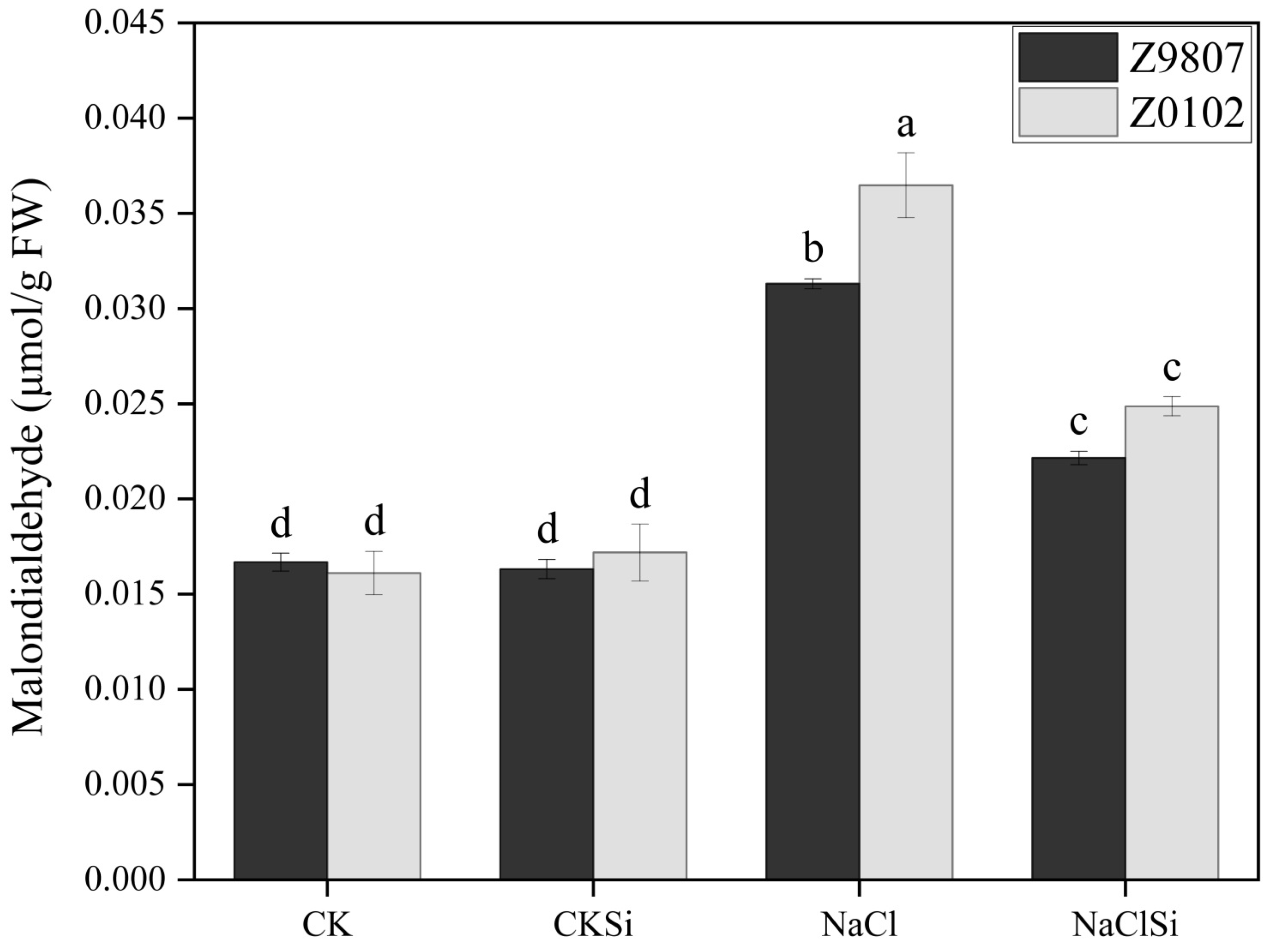
3.3. Effect of Si on the Accumulations of H2O2, O2− and MDA in Cotton Seedlings under Salt Stress
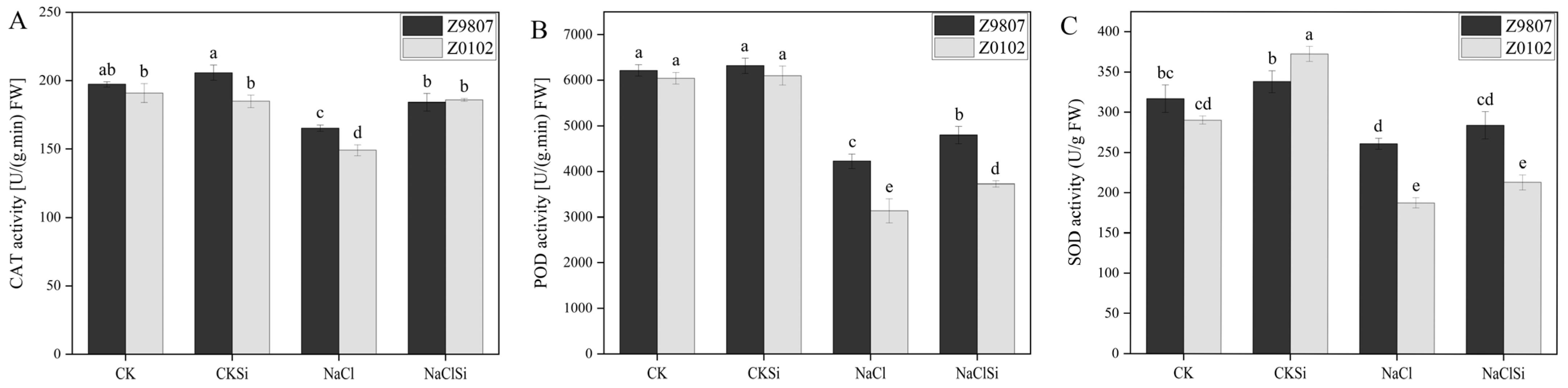
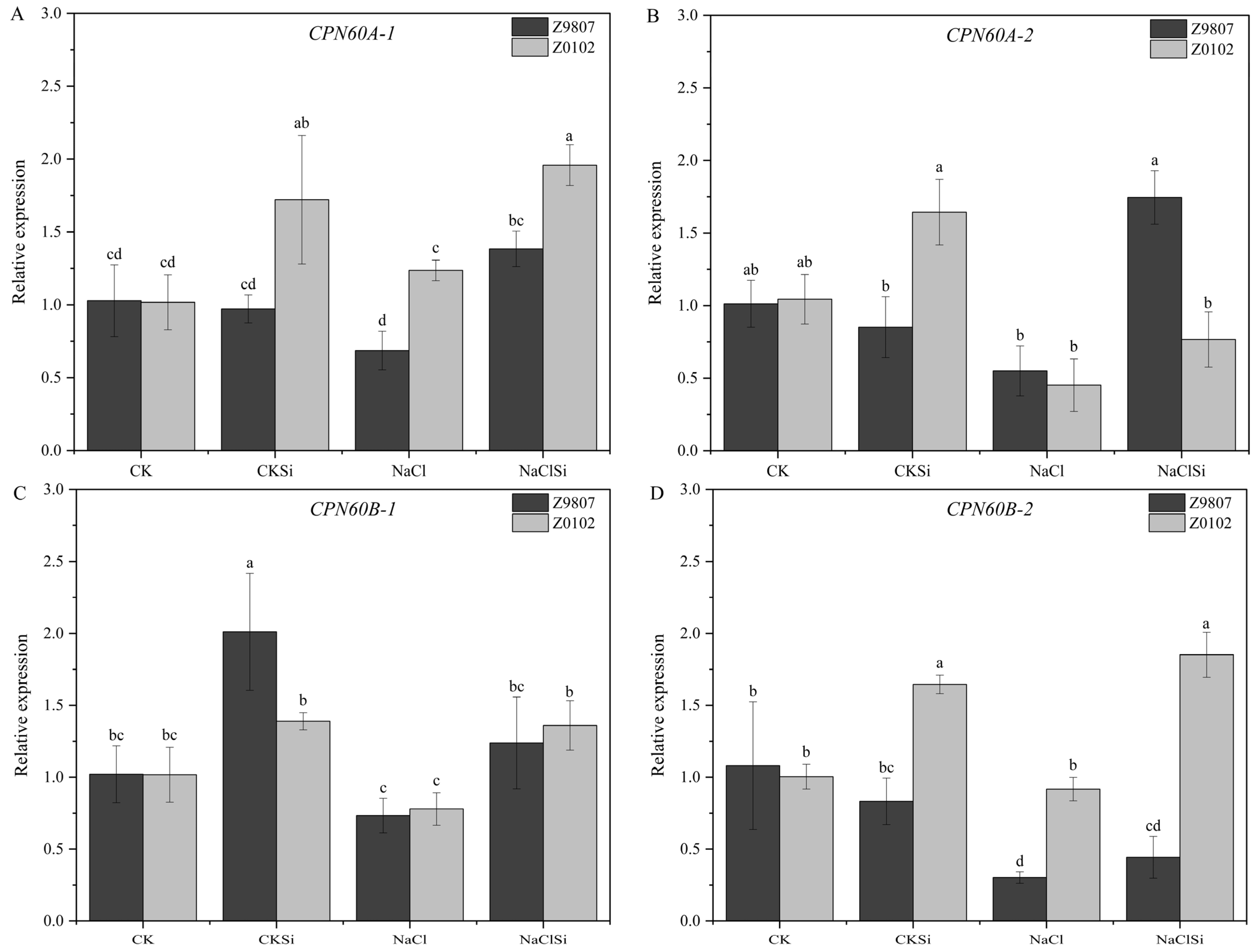
3.4. Effects of Exogenous Si on Antioxidase Activity and Antioxidase Gene Expression
3.5. Effects of Exogenous Silicon on Chlorophyll Content
3.6. Exogenous Silicon Facilitates Stomatal Opening
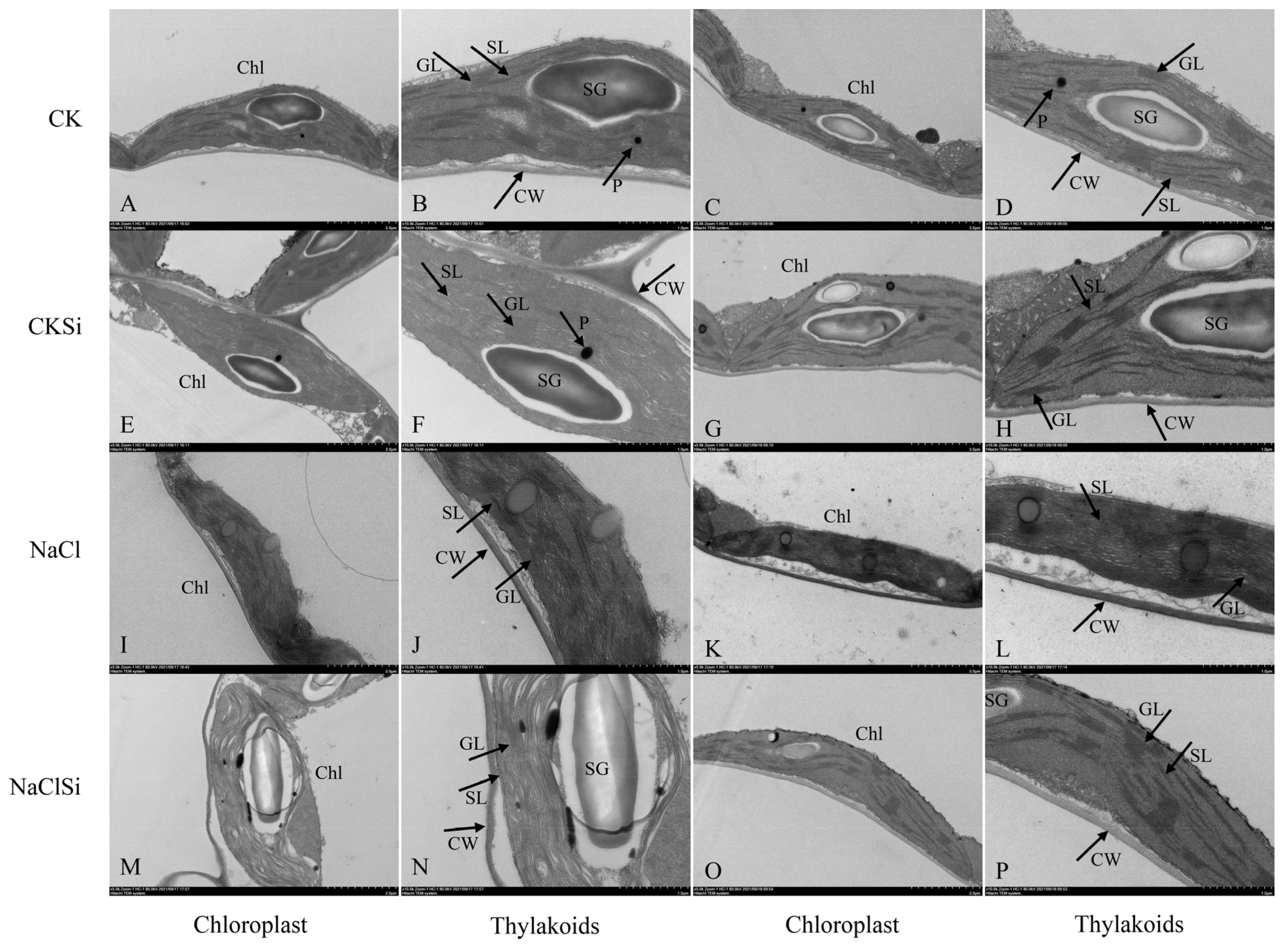
3.7. Ultrastructure of the Chloroplasts
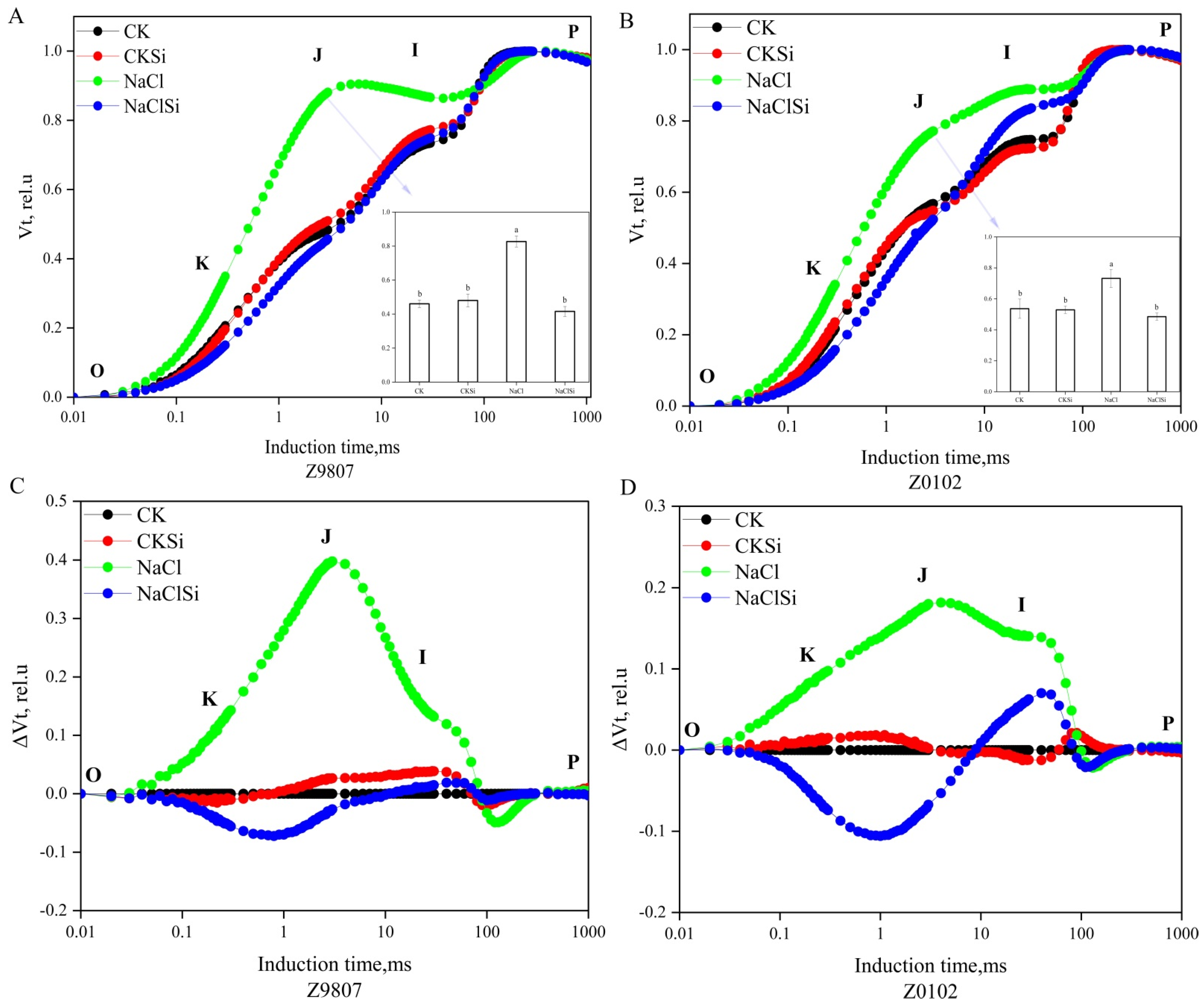
3.8. Impact of Exogenous Si on the Fluorescence Rise Kinetics O-J-I-P Curves in Cotton Leaves under Salt Stress
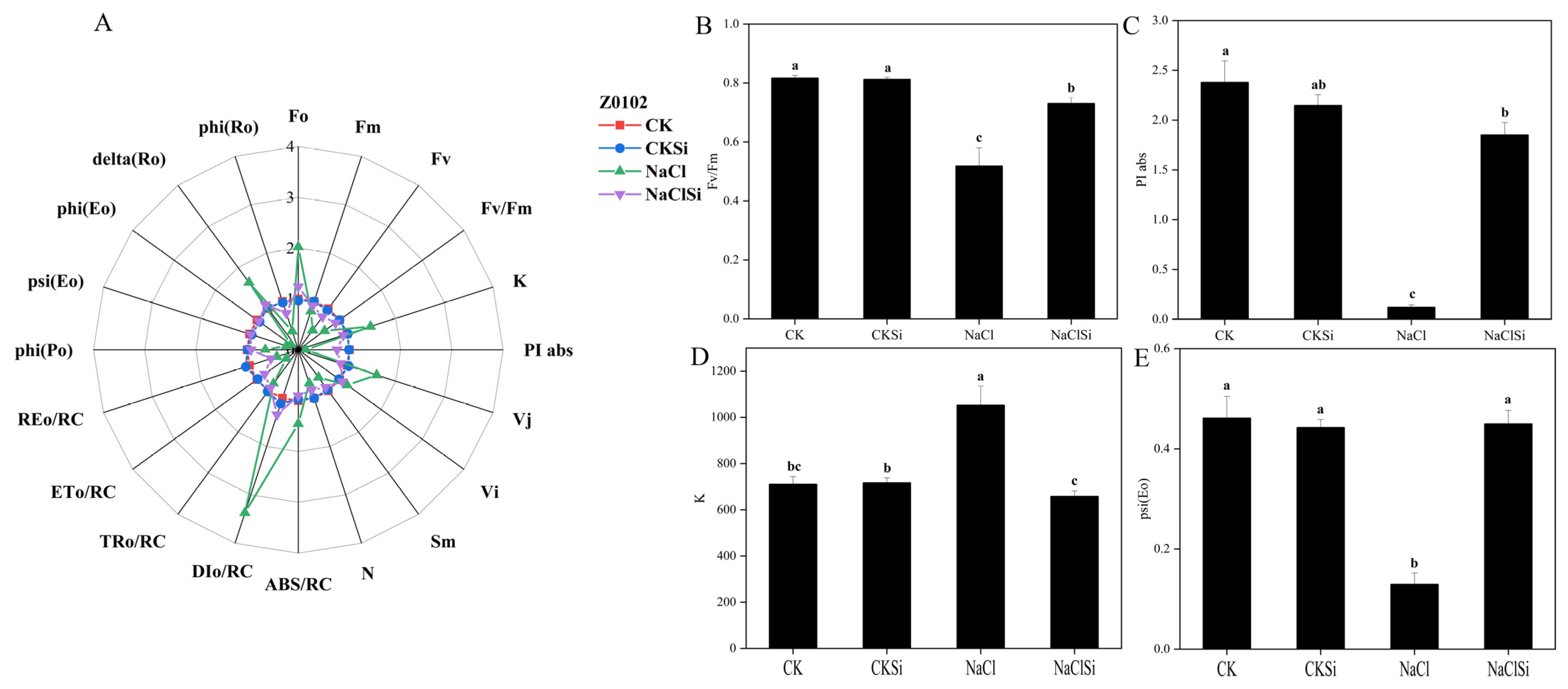
3.9. Fluorescence Transient OJIP-Detection Analysis
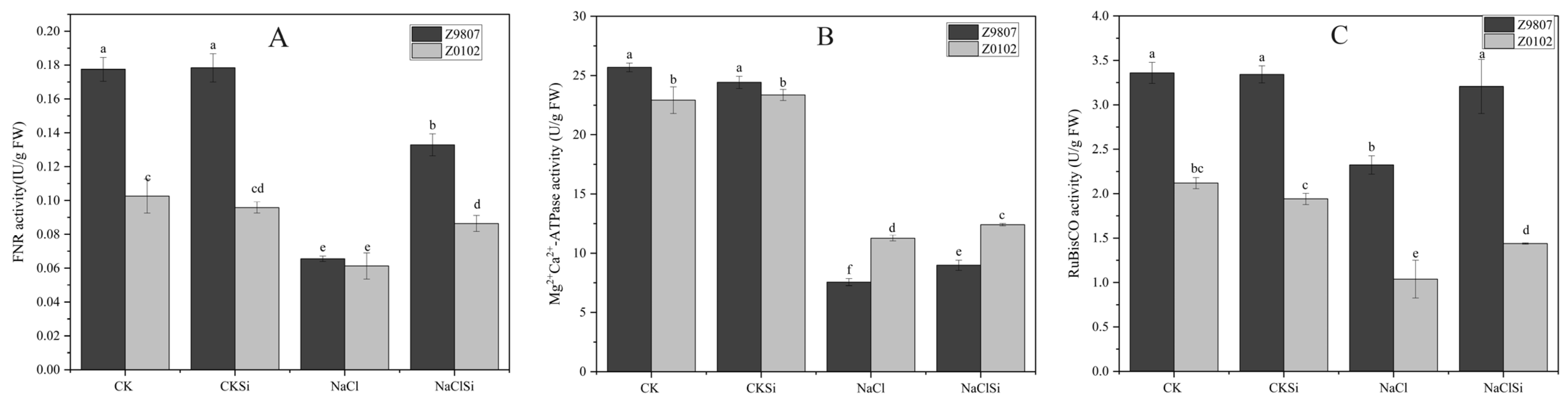
3.10. Si Enhanced the Activities of Key Enzymes in Photosynthesis and the Expression Levels of RubsiCO Genes in Cotton Seedlings under Salt Stress
4. Discussion
4.1. Si Alleviates Salt Damage by Modulating the Improvement of Photosynthesis
4.2. Si Alleviates Salt Induced Oxidative Stress by Promoting the Effective Transport of Photosynthetic Electrons and Regulating the Activities of Antioxidant Enzymes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wang, X.; Zhang, H.; Liu, X.; Shi, J.; Dong, Q.; Xu, Q.; Gui, H.; Song, M.; Yan, G. Early ABA-stimulated maintenance of Cl− homeostasis by mepiquat chloride priming confers salt tolerance in cotton seeds. Crop. J. 2021, 9, 387–399. [Google Scholar] [CrossRef]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef]
- He, Y.; Yu, C.; Zhou, L.; Chen, Y.; Liu, A.; Jin, J.; Hong, J.; Qi, Y.; Jiang, D. Rubisco decrease is involved in chloroplast protrusion and Rubisco-containing body formation in soybean (Glycine max.) under salt stress. Plant Physiol. Bioch. 2014, 74, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.H.; Wang, G.Z.; Si, W.T.; Zhou, Y.; Liu, Z.; Jia, J. Effects of salt stress on photosynthetic pigments and activity of ribulose-1,5-bisphosphate Carboxylase/Oxygenase in Kalidium foliatum1. Russ. J. Plant Phys. 2018, 65, 98–103. [Google Scholar] [CrossRef]
- Yang, X.; Lu, C. Photosynthesis is improved by exogenous glycinebetaine in salt-stressed maize plants. Physiol. Plant. 2005, 124, 343–352. [Google Scholar] [CrossRef]
- Sikder, R.K.; Wang, X.; Jin, D.; Zhang, H.; Gui, H.; Dong, Q.; Pang, N.; Zhang, X.; Song, M. Screening and evaluation of reliable traits of upland cotton (Gossypium hirsutum L.) genotypes for salt tolerance at the seedling growth stage. J. Cotton Res. 2020, 3, 11. [Google Scholar] [CrossRef]
- Satir, O.; Berberoglu, S. Crop yield prediction under soil salinity using satellite derived vegetation indices. Field Crop. Res. 2016, 192, 134–143. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Liu, B.; Soundararajan, P.; Manivannan, A. Mechanisms of silicon-mediated amelioration of salt stress in plants. Plants 2019, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E. Review The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Hodoson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot Lond. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [Green Version]
- Guntzer, F.; Keller, C.; Meunier, J. Benefits of plant silicon for crops: A review. Agron Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Wang, H.; Hu, Y.; Mao, L.; Sun, L.; Dong, T.; Nan, W.; Bi, Y. Silicon does not mitigate cell death in cultured tobacco BY-2 cells subjected to salinity without ethylene emission. Plant Cell Rep. 2015, 34, 331–343. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotox. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef]
- Alam, P.; Balawi, T.H.; Altalayan, F.H.; Hatamleh, A.A.; Ashraf, M.; Ahmad, P. Silicon attenuates the negative effects of chromium stress in tomato plants by modifying antioxidant enzyme activities, ascorbate—Glutathione cycle and glyoxalase system. Acta Physiol. Plant 2021, 43, 110. [Google Scholar] [CrossRef]
- Misuthova, A.; Slovakova, L.; Kollarova, K.; Vaculik, M. Effect of silicon on root growth, ionomics and antioxidant performance of maize roots exposed to as toxicity. Plant Physiol. Biochem. 2021, 168, 155–166. [Google Scholar] [CrossRef]
- Jain, S.; Rai, P.; Singh, J.; Singh, V.P.; Prasad, R.; Rana, S.; Deshmukh, R.; Tripathi, D.K.; Sharma, S. Exogenous addition of silicon alleviates metsulfuron methyl induced stress in wheat seedlings. Plant Physiol. Biochem. 2021, 167, 705–712. [Google Scholar] [CrossRef]
- Zhang, W.; Xie, Z.; Wang, L.; Li, M.; Lang, D.; Zhang, X. Silicon alleviates salt and drought stress of Glycyrrhiza uralensis seedling by altering antioxidant metabolism and osmotic adjustment. J. Plant Res. 2017, 130, 611–624. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant 2015, 37, 71. [Google Scholar] [CrossRef]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Thorne, S.J.; Hartley, S.E.; Maathuis, F. Is silicon a panacea for alleviating drought and salt stress in crops? Front. Plant Sci. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, G. Exploring the significant contribution of silicon in regulation of cellular redox homeostasis for conferring stress tolerance in plants. Plant Physiol. Bioch. 2021, 166, 393–404. [Google Scholar] [CrossRef]
- Liang, Y.C.N.A. Effects of silicon on enzyme activity and sodium, potassium and calcium concentration in barley under salt stress. Plant Soil 1999, 209, 217–224. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. Amst. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Zhao, W.; Zhu, X. Silicon improves photosynthesis and strengthens enzyme activities in the C3 succulent xerophyte Zygophyllum xanthoxylum under drought stress. J. Plant Physiol. 2016, 199, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Cheema, M.A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Nawaz, A.; Abbas, T.; Ali, Q. Physiological and biochemical attributes of bread wheat (Triticum aestivum L.) seedlings are influenced by foliar application of silicon and selenium under water deficit. Acta Physiol. Plant 2019, 41, 146. [Google Scholar] [CrossRef]
- Muneer, S.; Park, Y.; Manivannan, A.; Soundararajan, P.; Jeong, B. Physiological and proteomic analysis in chloroplasts of Solanum lycopersicum L. under silicon efficiency and salinity stress. Int. J. Mol. Sci. 2014, 15, 21803–21824. [Google Scholar] [CrossRef]
- Kong, X.; Luo, Z.; Zhang, Y.; Li, W.; Dong, H. Soaking in H2O2 regulates ABA biosynthesis and GA catabolism in germinating cotton seeds under salt stress. Acta Physiol. Plant 2017, 39, 2. [Google Scholar] [CrossRef]
- Jin, D.; Wang, X.; Xu, Y.; Gui, H.; Zhang, H.; Dong, Q.; Sikder, R.K.; Yang, G.; Song, M. Chemical defoliant promotes leaf abscission by altering ROS metabolism and photosynthetic efficiency in Gossypium hirsutum. Int. J. Mol. Sci. 2020, 21, 2738. [Google Scholar] [CrossRef] [Green Version]
- Shu, S.; Yuan, Y.; Chen, J.; Sun, J.; Zhang, W.; Tang, Y.; Zhong, M.; Guo, S. The role of putrescine in the regulation of proteins and fatty acids of thylakoid membranes under salt stress. Sci. Rep. 2015, 5, 14390. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Chen, K. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Meng, Y.; Yin, Q.; Yan, Z.; Wang, Y.; Niu, J.; Zhang, J.; Fan, K. Exogenous silicon enhanced salt resistance by maintaining K+/Na+ homeostasis and antioxidant performance in Alfalfa leaves. Front. Plant Sci. 2020, 11, 1183. [Google Scholar] [CrossRef]
- Abbas, T.; Balal, R.M.; Shahid, M.A.; Pervez, M.A.; Ayyub, C.M.; Aqueel, M.A.; Javaid, M.M. Silicon-induced alleviation of NaCl toxicity in okra (Abelmoschus esculentus) is associated with enhanced photosynthesis, osmoprotectants and antioxidant metabolism. Acta Physiol. Plant 2015, 37, 6. [Google Scholar] [CrossRef]
- Huang, X.; Chao, D.; Gao, J.; Zhu, M.; Shi, M.; Lin, H. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Gene Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum “Bugwang”. Biomed. Res. Int. 2016, 2016, 3076357. [Google Scholar] [CrossRef] [Green Version]
- Cao, B.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hortic. Amst. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Ali, S.; Farooq, M.A.; Yasmeen, T.; Hussain, S.; Arif, M.S.; Abbas, F.; Bharwana, S.A.; Zhang, G. The influence of silicon on barley growth, photosynthesis and ultra-structure under chromium stress. Ecotox. Environ. Saf. 2013, 89, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.C.; Loik, M.E. Water relations and photosynthesis along an elevation gradient for Artemisia tridentata during an historic drought. Oecologia 2016, 181, 65–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Ghosh, I.; Khan, S.; Banerjee, G.; Dziarski, A.; Vinyard, D.J.; Debus, R.J.; Brudvig, G.W. Insights into proton-transfer pathways during water oxidation in photosystem II. J. Phys. Chem. B 2019, 123, 8195–8202. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhao, N.; Qin, D.; Liu, S.; Jiang, S.; Xu, L.; Sun, Z.; Yan, D.; Hu, A. The synergistic effects of silicon and selenium on enhancing salt tolerance of maize plants. Environ. Exp. Bot. 2021, 187, 104482. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis Press: London, UK, 2000; pp. 445–483. [Google Scholar]
- Chen, H.X.; Gao, H.Y.; An, S.Z.; Li, W.J. Dissipation of excess energy in mehler-peroxidase reaction in Rumex leaves during salt shock. Photosynthetica 2004, 42, 117–122. [Google Scholar] [CrossRef]
- Peng, C.; Su, X.; Pan, X.; Liu, Z.; Mei, L. Structure, assembly and energy transfer of plant photosystem II supercomplex. Biochim. Biophys. Acta 2018, 1859, 633–644. [Google Scholar]
- Chen, X.; Zhou, Y.; Cong, Y.; Zhu, P.; Xing, J.; Cui, J.; Xu, W.; Shi, Q.; Diao, M.; Liu, H.Y. Ascorbic acid-induced photosynthetic adaptability of processing tomatoes to salt stress probed by fast OJIP fluorescence rise. Front. Plant Sci. 2021, 12, 594400. [Google Scholar] [CrossRef]
- Wang, X.; Ding, X.; Qi, Y.; Shi, M. Calcium amendment for improved germination, plant growth, and leaf photosynthetic electron transport in oat (Avena sativa) under NaCl stress. PLoS ONE 2021, 16, e256529. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, Y.; Cheng, Y.; Zou, T.; Duan, X.; Gong, M.; Zou, Z. Generation of reactive oxygen species and their functions and deleterious effects in plants. Acta Bot. Boreal. Occident. Sin. 2014, 34, 1916–1926, (In Chinese with English Abstract). [Google Scholar]
- Boris, I.; Sergey, K. Participation of photosynthetic electron transport in production and scavenging of reactive oxygen species. Antioxid. Redox Sign. 2003, 5, 43–53. [Google Scholar]
- Saleh, J.; Najafi, N.; Oustan, S. Effects of silicon application on wheat growth and some physiological characteristics under different levels and sources of salinity. Commun. Soil Sci. Plan 2017, 48, 1114–1122. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Qin, H.; Ding, H.; Li, Y.; Guo, T. Silicon application alleviates drought stress in wheat through transcriptional regulation of multiple antioxidant defense pathways. J. Plant Growth Regul. 2016, 35, 1–10. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Pn (µmol m−2 s−1) | Ci (µmol mol−1) | Gs (mol m−2 s−1) | Tr (mmol m−2 s−1) |
|---|---|---|---|---|---|
| Z9807 | CK | 8.2 a | 322.7 b | 0.20 b | 3.35 a |
| CKSi | 8.1 a | 332.9 b | 0.23 b | 3.80 a | |
| NaCl | 5.3 c | 223.3 d | 0.05 e | 1.20 d | |
| NaClSi | 7.3 b | 266.8 c | 0.09 c | 1.60 c | |
| Z0102 | CK | 7.4 b | 329.1 b | 0.20 b | 2.70 b |
| CKSi | 7.2 b | 347.3 a | 0.28 a | 3.61 a | |
| NaCl | 4.7 d | 220.3 d | 0.06 e | 1.12 d | |
| NaClSi | 7.0 b | 278.3 c | 0.11 c | 1.96 c |
| Genotype | Treatment | Chlorophyll Content (mg g−1 FW) | Carotenoids (mg g−1 FW) | ||
|---|---|---|---|---|---|
| Chlorophyll a | Chlorophyll b | Chl (a + b) | |||
| Z9807 | CK | 1.54 a | 0.84 ab | 2.37 a | 0.083 b |
| CKSi | 1.53 a | 0.88 a | 2.41 a | 0.102 a | |
| NaCl | 0.92 d | 0.52 de | 1.44 d | 0.063 cd | |
| NaClSi | 1.29 b | 0.66 c | 1.95 b | 0.066 cd | |
| Z0102 | CK | 1.27 b | 0.78 b | 2.05 b | 0.075 bc |
| CKSi | 1.27 b | 0.78 b | 2.04 b | 0.087 b | |
| NaCl | 0.75 e | 0.49 e | 1.23 e | 0.054 d | |
| NaClSi | 1.05 c | 0.56 d | 1.61 c | 0.065 cd | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Qi, Q.; Zhang, H.; Dong, Q.; Iqbal, A.; Gui, H.; Kayoumu, M.; Song, M.; Zhang, X.; Wang, X. Ameliorative Effects of Silicon against Salt Stress in Gossypium hirsutum L. Antioxidants 2022, 11, 1520. https://doi.org/10.3390/antiox11081520
Li L, Qi Q, Zhang H, Dong Q, Iqbal A, Gui H, Kayoumu M, Song M, Zhang X, Wang X. Ameliorative Effects of Silicon against Salt Stress in Gossypium hirsutum L. Antioxidants. 2022; 11(8):1520. https://doi.org/10.3390/antiox11081520
Chicago/Turabian StyleLi, Leilei, Qian Qi, Hengheng Zhang, Qiang Dong, Asif Iqbal, Huiping Gui, Mirezhatijiang Kayoumu, Meizhen Song, Xiling Zhang, and Xiangru Wang. 2022. "Ameliorative Effects of Silicon against Salt Stress in Gossypium hirsutum L." Antioxidants 11, no. 8: 1520. https://doi.org/10.3390/antiox11081520