
Zinc Oxide Nanoparticles (ZnO-NPs) Suppress Fertility by Activating Autophagy, Apoptosis, and Oxidative Stress in the Developing Oocytes of Female Zebrafish


 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
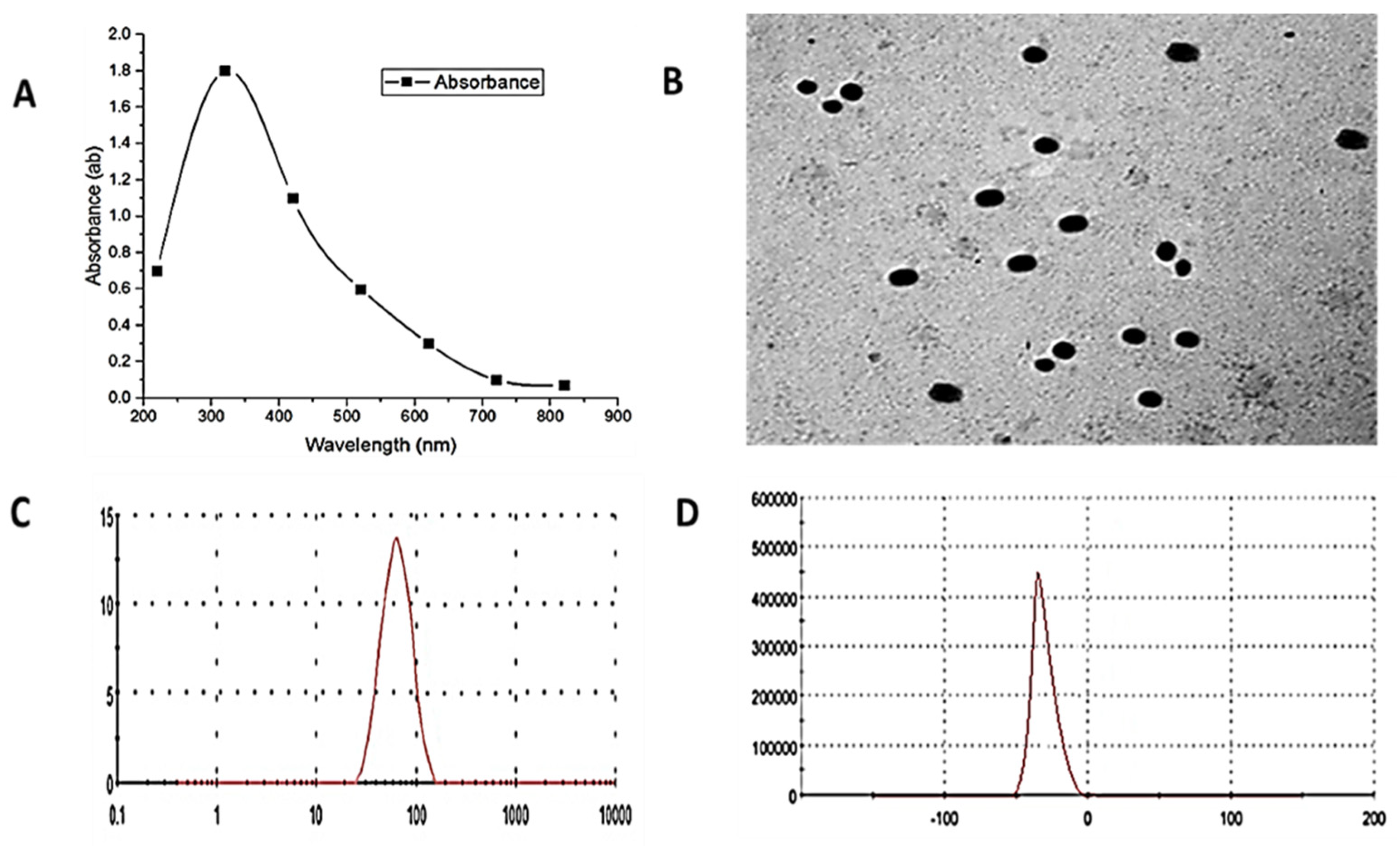
2.1. ZnO-NPs Preparation and Characterization
2.2. Animal Husbandry and Experiment Design
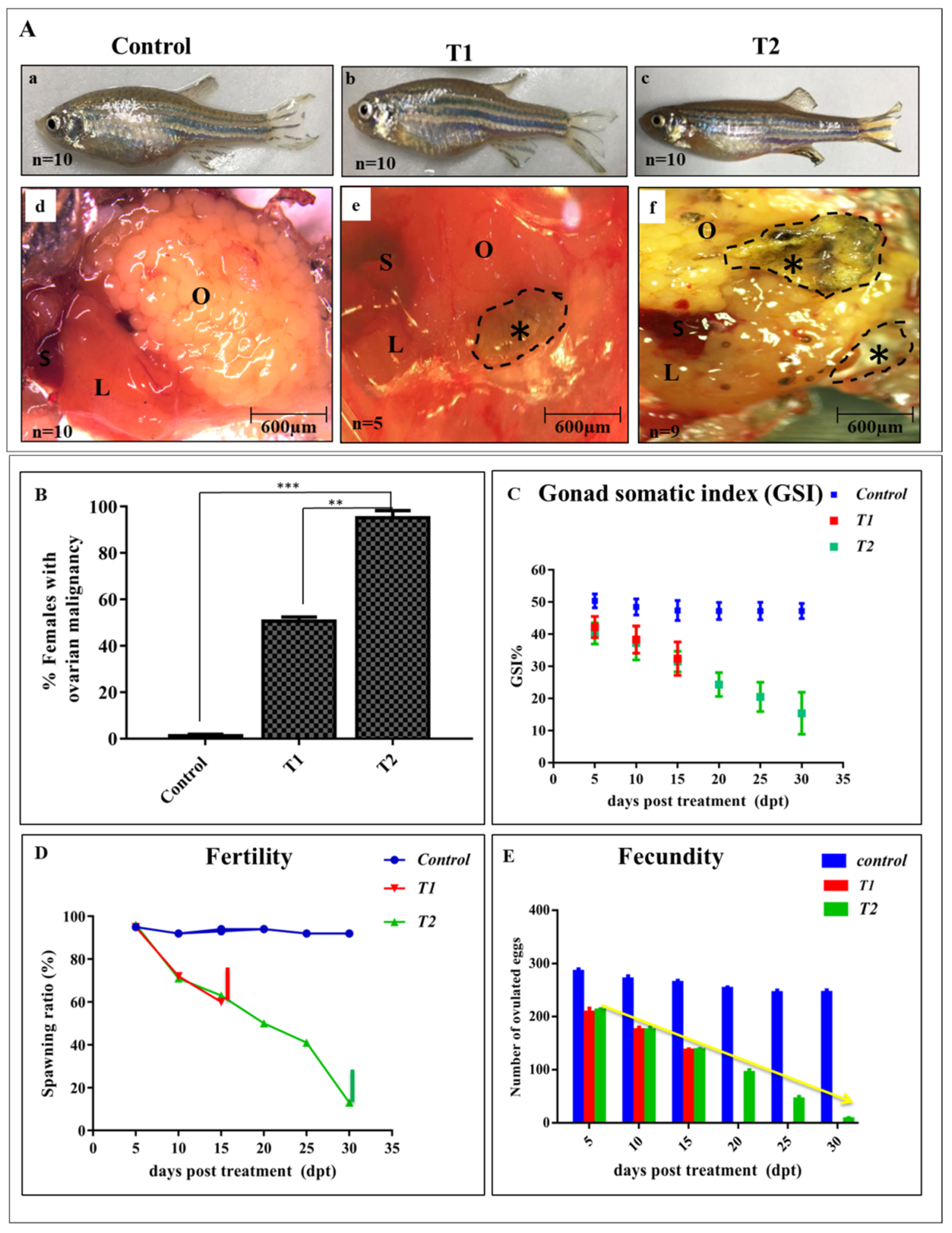
2.3. Fertility, Fecundity, and Gonad Somatic Index Calculations
2.4. Determination of Zinc Residues in the Whole Fish Body
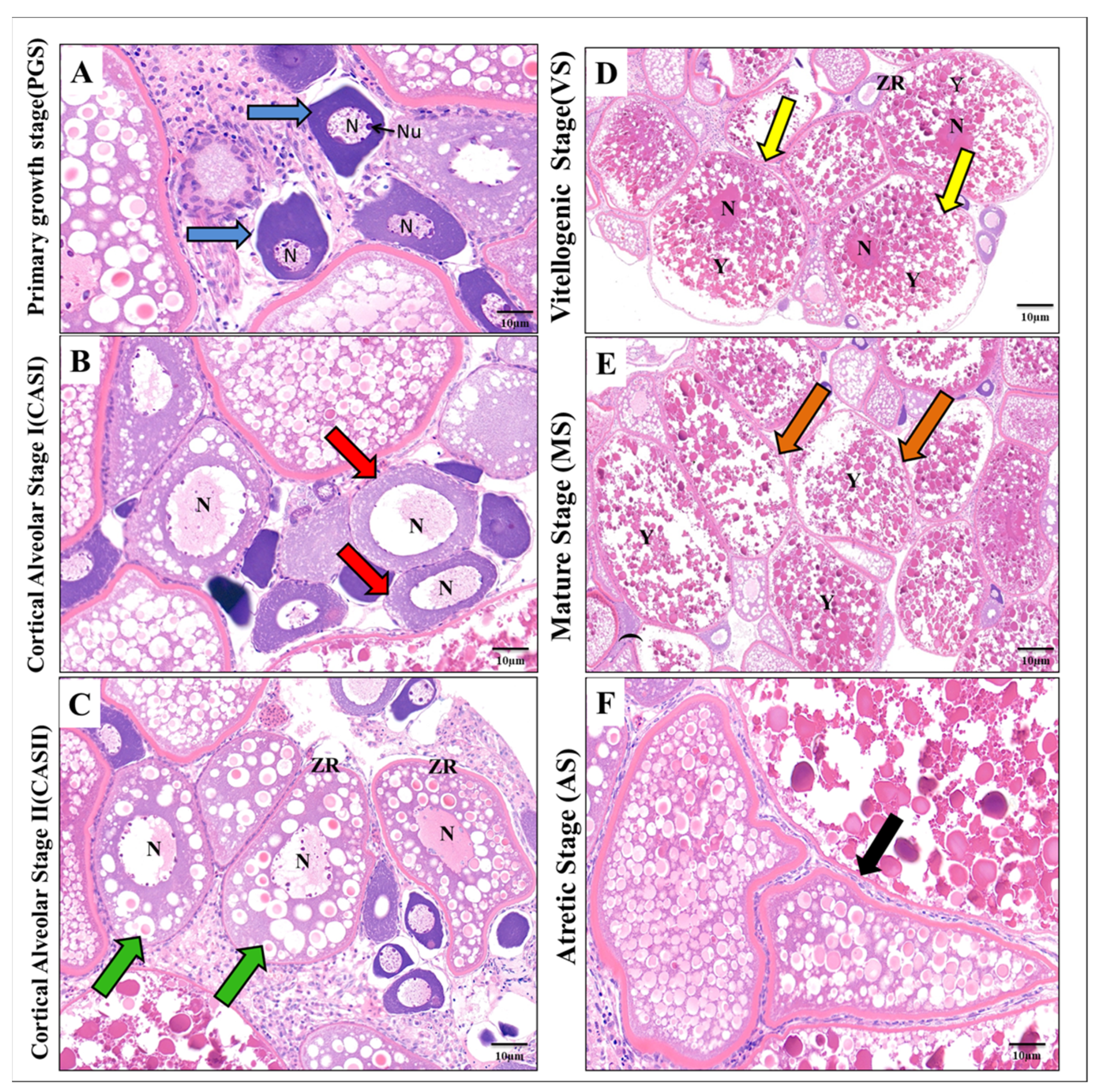
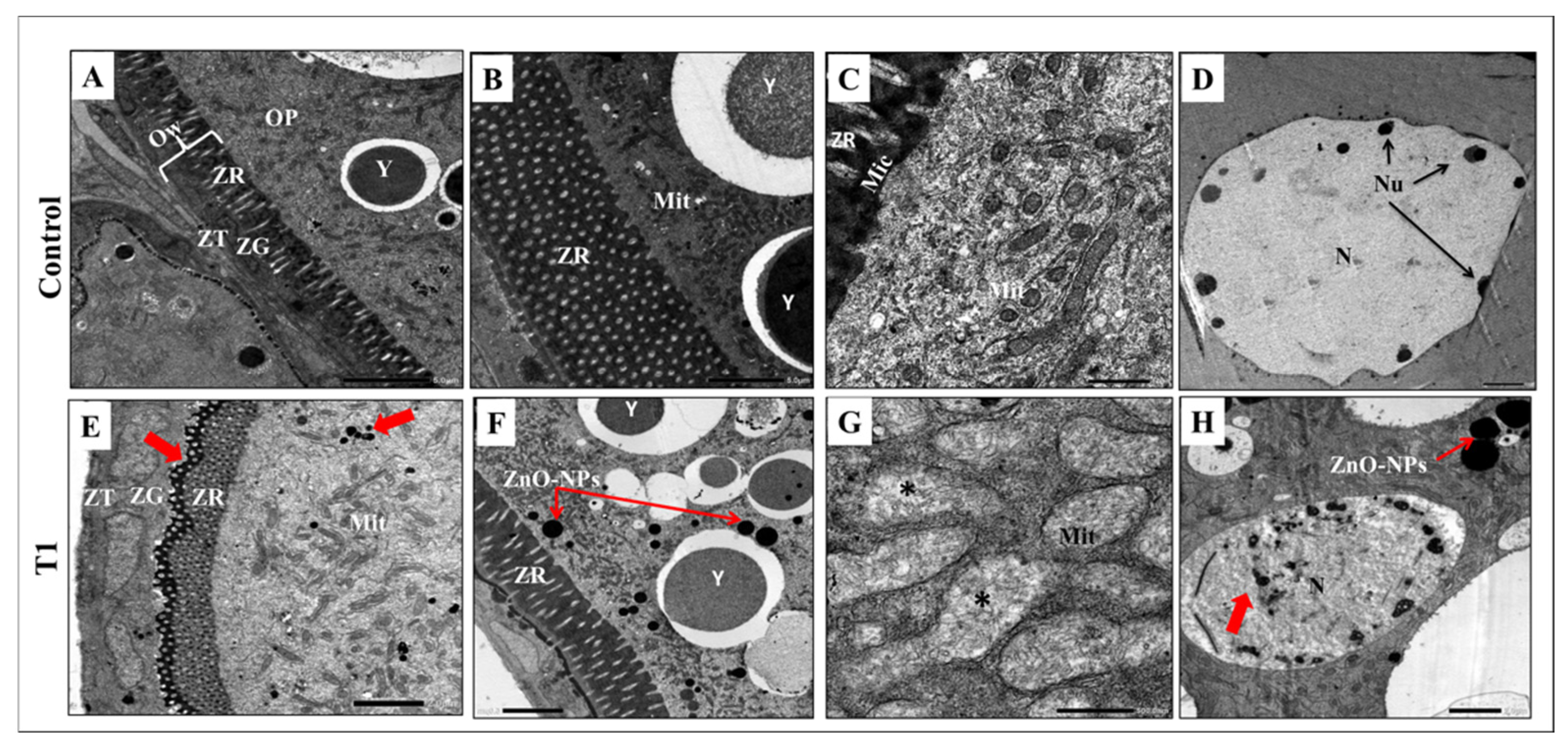
2.5. Histological Assessments and Transmission Electron Microscope (TEM) Preparations
2.6. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
2.7. Protein Extraction and Western Blot Analysis
2.8. Statistical Analysis and Graph Preparation
3. Results
3.1. Zinc Oxide Nanoparticles (ZnO-NPs) Characterization
3.2. ZnO-NPs Affect Ovarian Morphology and Alter Female Fertility and Fecundity
3.3. Zn Residues in the Female’s Body
3.4. Effect of ZnO-NPs on Histological Findings of Oocyte Maturation Levels
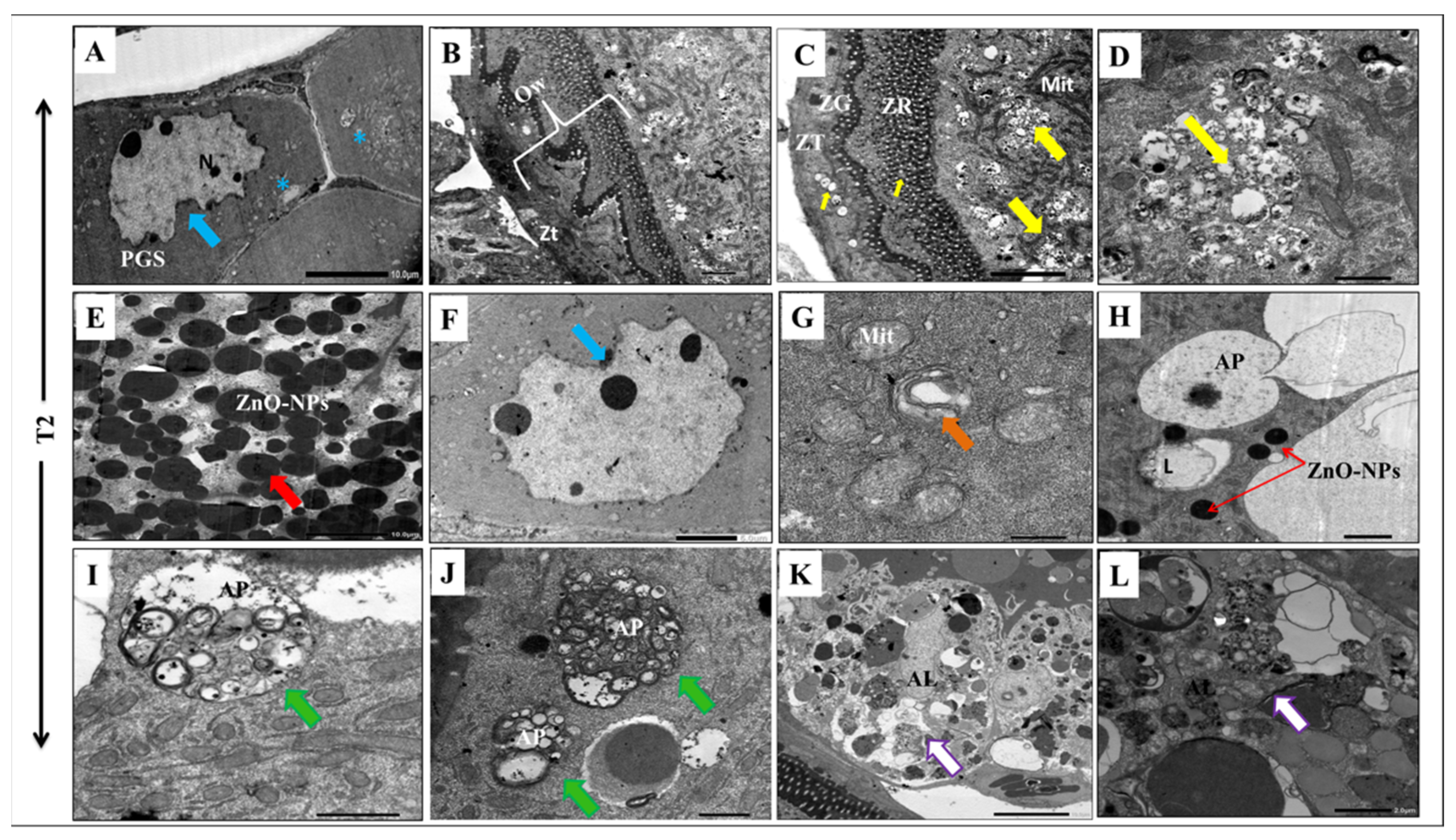
3.5. ZnO-NPs Induce Cytotoxicity and Autophagosome Formation at the Ultrastructural Level
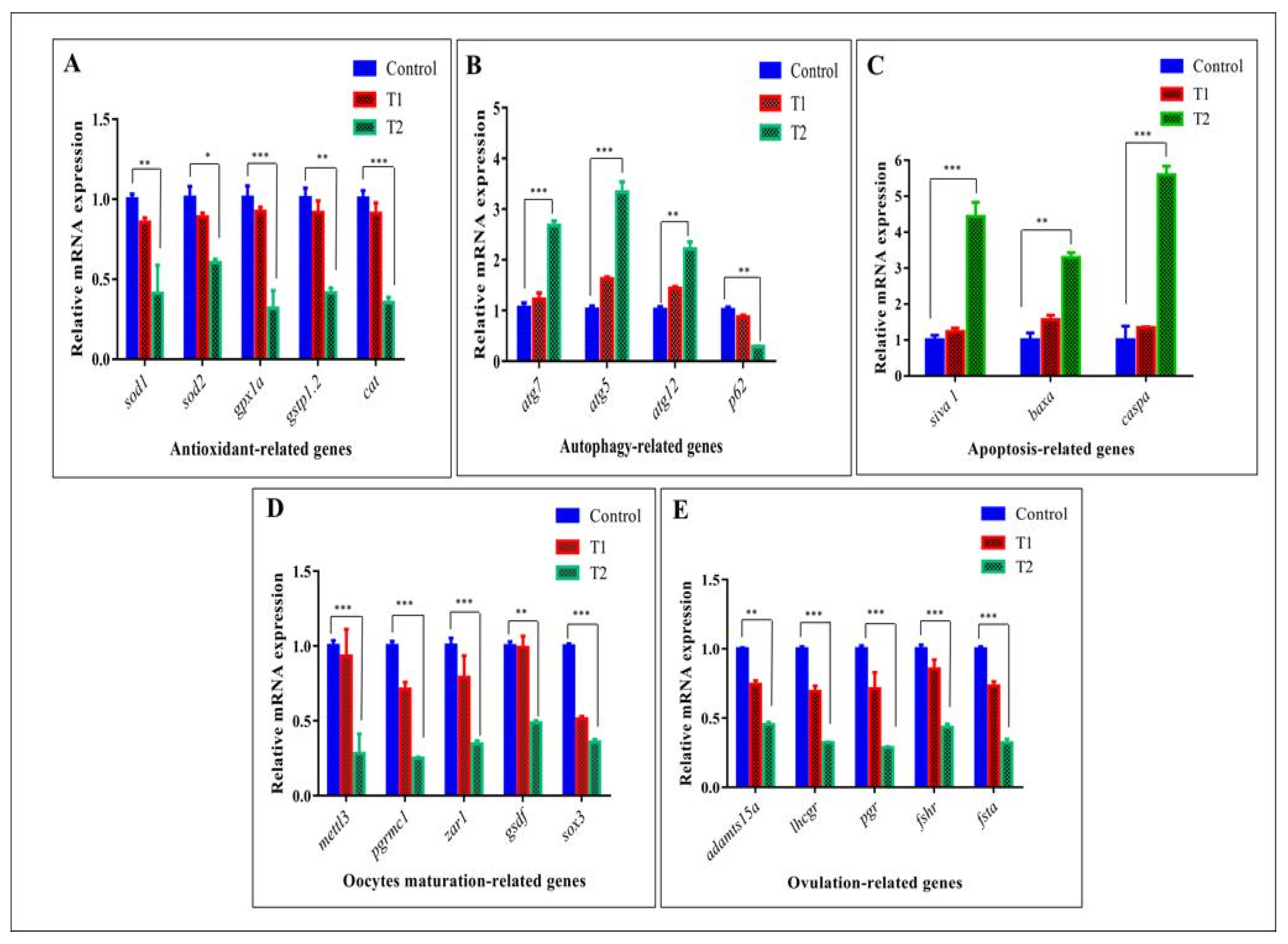
3.6. Effect of ZnO-NPs on the Expression of Specific Genes (Antioxidant, Autophagy, Apoptosis, Oocyte Maturation, and Ovulation-Related Genes) in Zebrafish Ovarian Cells
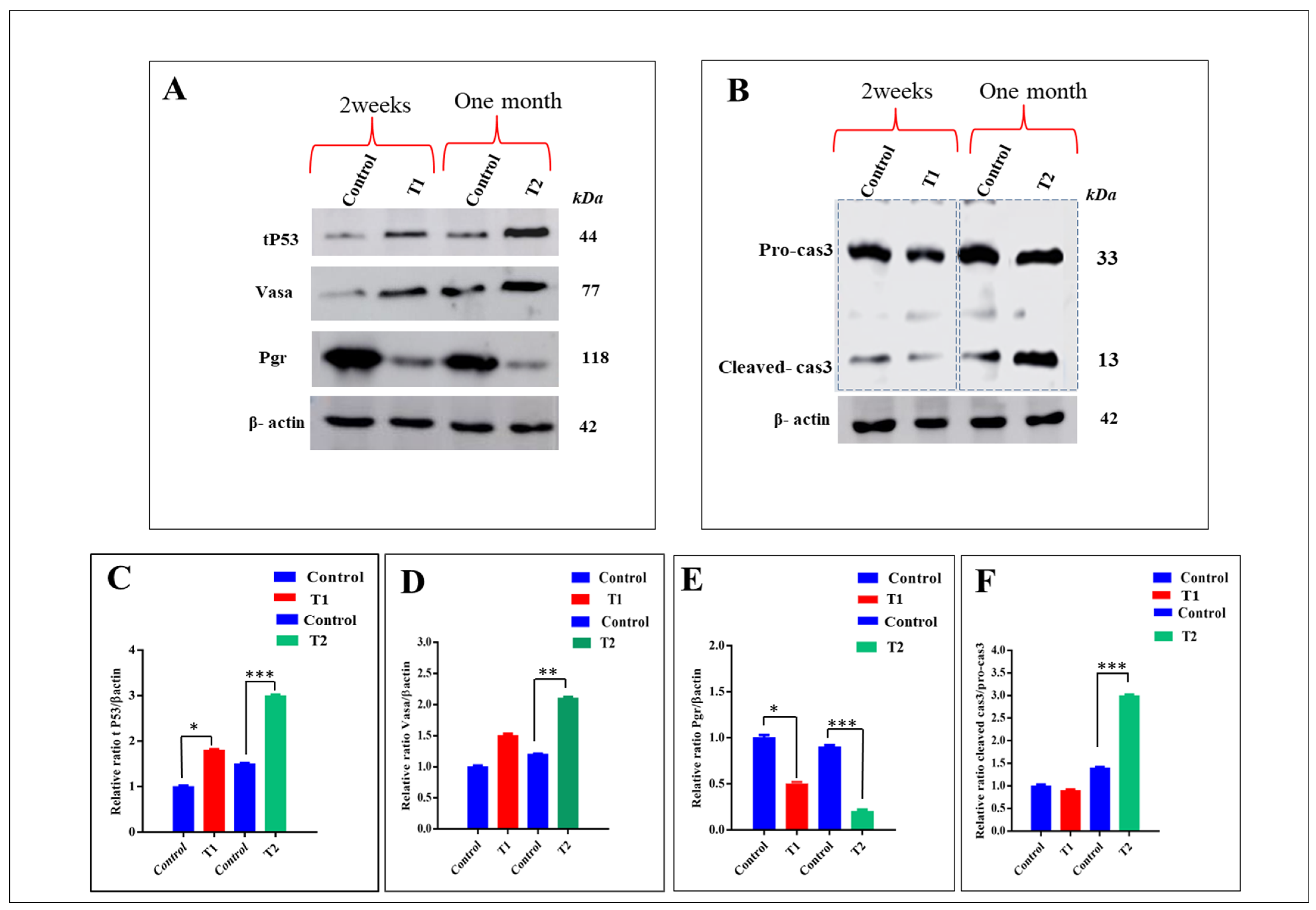
3.7. Effect of ZnO-NPs on the Expression of Proteins Related to Apoptosis Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heiligtag, F.J.; Niederberger, M. The fascinating world of nanoparticle research. Mater. Today 2013, 16, 262–271. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Pezzuto, F.; Scarano, A. Cytotoxic and Bacteriostatic Activity of Nanostructured TiO2 Coatings. Pol. J. Microbiol. 2016, 65, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Guildford, A.L.; Poletti, T.; Osbourne, L.H.; Di Cerbo, A.; Gatti, A.M.; Santin, M. Nanoparticles of a different source induce different patterns of activation in key biochemical and cellular components of the host response. J. R. Soc. Interf. 2009, 6, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Darroudi, M. Zinc oxide nanoparticles: Biological synthesis and biomedical applications. Ceram. Int. 2017, 43, 907–914. [Google Scholar] [CrossRef]
- Islam, F.; Shohag, S.; Uddin, M.J.; Islam, M.R.; Nafady, M.H.; Akter, A.; Mitra, S.; Roy, A.; Emran, T.B.; Cavalu, S. Exploring the Journey of Zinc Oxide Nanoparticles (ZnO-NPs) toward Biomedical Applications. Materials 2022, 15, 2160. [Google Scholar] [CrossRef]
- Jiang, J.; Pi, J.; Cai, J. The Advancing of Zinc Oxide Nanoparticles for Biomedical Applications. Bioinorg. Chem. Appl. 2018, 2018, 1062562. [Google Scholar] [CrossRef]
- Scarano, A.; Murmura, G.; Vantaggiato, G.; Lauritano, D.; Silvestre-Rangil, J.; Di Cerbo, A.; Lorusso, F. Delayed expansion of atrophic mandible (deam): A case report. Oral Implantol. (Rome) 2017, 10, 190–196. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, S.; Wu, Y.; You, H.; Lv, L. Acute ZnO nanoparticles exposure induces developmental toxicity, oxidative stress and DNA damage in embryo-larval zebrafish. Aquat. Toxicol. 2013, 136, 49–59. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, J.; Zhang, X.; Chang, Y.; Chen, Y. The impact of ZnO nanoparticle aggregates on the embryonic development of zebrafish (Danio rerio). Nanotechnology 2009, 20, 195103. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Ge, W.; Zhang, P.; Liu, X.; Zhang, W.; Hao, Y.; Yu, S.; Li, L.; Chu, M.; et al. Oocyte exposure to ZnO nanoparticles inhibits early embryonic development through the gamma-H2AX and NF-kappaB signaling pathways. Oncotarget 2017, 8, 42673–42692. [Google Scholar] [CrossRef]
- Saber, M.; Hayaei-Tehrani, R.S.; Mokhtari, S.; Hoorzad, P.; Esfandiari, F. In vitro cytotoxicity of zinc oxide nanoparticles in mouse ovarian germ cells. Toxicol. Vitr. 2021, 70, 105032. [Google Scholar] [CrossRef] [PubMed]
- Selman, K.; Wallace, R.A.; Sarka, A.; Qi, X. Stages of oocyte development in the zebrafish, Brachydanio rerio. J. Morphol. 1993, 218, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Cakici, O.; İsisag Ucuncu, S. Oocyte Development In Zebrafish, Danio rerio (Teleostei: Cyprinidae). J. Fish.Aqua. Sci. 2007, 24, 137–141. [Google Scholar]
- Lessman, C.A. Oocyte maturation: Converting the zebrafish oocyte to the fertilizable egg. Gen. Comp. Endocrinol. 2009, 161, 53–57. [Google Scholar] [CrossRef]
- Eppig, J.J. Oocyte control of ovarian follicular development and function in mammals. Reproduction 2001, 122, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.Q.; Sugiura, K.; Eppig, J.J. Mouse oocyte control of granulosa cell development and function: Paracrine regulation of cumulus cell metabolism. Semin. Reprod. Med. 2009, 27, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Gawriluk, T.R.; Hale, A.N.; Flaws, J.A.; Dillon, C.P.; Green, D.R.; Rucker, E.B., 3rd. Autophagy is a cell survival program for female germ cells in the murine ovary. Reproduction 2011, 141, 759–765. [Google Scholar] [CrossRef]
- Agnello, M.; Chiarelli, R.; Martino, C.; Bosco, L.; Roccheri, M.C. Autophagy is required for sea urchin oogenesis and early development. Zygote 2016, 24, 918–926. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Emr, S.D. Autophagy as a regulated pathway of cellular degradation. Science 2000, 290, 1717–1721. [Google Scholar] [CrossRef]
- Yoshimori, T. Autophagy: Paying Charon’s toll. Cell 2007, 128, 833–836. [Google Scholar] [CrossRef]
- D’Herde, K.; De Prest, B.; Roels, F. Subtypes of active cell death in the granulosa of ovarian atretic follicles in the quail (Coturnix coturnix japonica). Reprod. Nutr. Dev. 1996, 36, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Jo, M.W.; Lee, E.Y.; Yoon, B.K.; Choi, D.S. The role of autophagy in follicular development and atresia in rat granulosa cells. Fertil. Steril. 2010, 93, 2532–2537. [Google Scholar] [CrossRef]
- Gioacchini, G.; Dalla Valle, L.; Benato, F.; Fimia, G.M.; Nardacci, R.; Ciccosanti, F.; Piacentini, M.; Borini, A.; Carnevali, O. Interplay between autophagy and apoptosis in the development of Danio rerio follicles and the effects of a probiotic. Reprod. Fertil. Dev. 2013, 25, 1115–1125. [Google Scholar] [CrossRef] [PubMed]
- Thome, R.G.; Santos, H.B.; Arantes, F.P.; Domingos, F.F.; Bazzoli, N.; Rizzo, E. Dual roles for autophagy during follicular atresia in fish ovary. Autophagy 2009, 5, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Kuma, A.; Mizushima, N. The role of autophagy during the oocyte-to-embryo transition. Autophagy 2008, 4, 1076–1078. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Aoki, A.; Kusabiraki, T.; Shima, T.; Yoshino, O.; Cheng, S.B.; Sharma, S.; Saito, S. Role of autophagy in oocytogenesis, embryogenesis, implantation, and pathophysiology of pre-eclampsia. J Obstet Gynaecol. Res. 2017, 43, 633–643. [Google Scholar] [CrossRef]
- Ge, W. Intrafollicular paracrine communication in the zebrafish ovary: The state of the art of an emerging model for the study of vertebrate folliculogenesis. Mol. Cell. Endocrinol. 2005, 237, 1–10. [Google Scholar] [CrossRef]
- Clelland, E.; Peng, C. Endocrine/paracrine control of zebrafish ovarian development. Mol. Cell Endocrinol. 2009, 312, 42–52. [Google Scholar] [CrossRef]
- Elkouby, Y.M. All in one - integrating cell polarity, meiosis, mitosis and mechanical forces in early oocyte differentiation in vertebrates. Int. J. Dev. Biol. 2017, 61, 179–193. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alkhatib, F.M.; Alzahrani, S.O.; Shafi, M.E.; El Abdel-Hamid, S.; Taha, T.F.; Aboelenin, S.M.; Soliman, M.M.; Ahmed, N.H. Impact of mycogenic zinc nanoparticles on performance, behavior, immune response, and microbial load in Oreochromis niloticus. Saudi J. Biol. Sci. 2021, 28, 4592–4604. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio); University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Tang, H.; Liu, Y.; Luo, D.; Ogawa, S.; Yin, Y.; Li, S.; Zhang, Y.; Hu, W.; Parhar, I.S.; Lin, H.; et al. The kiss/kissr systems are dispensable for zebrafish reproduction: Evidence from gene knockout studies. Endocrinology 2015, 156, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, H.; Wang, L.; He, J.; Guo, Y.; Liu, Y.; Liu, X.; Lin, H. Fertility Enhancement but Premature Ovarian Failure in esr1-Deficient Female Zebrafish. Front. Endocrinol. (Lausanne) 2018, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Mahaffey, K.R.; Capar, S.G.; Gladen, B.C.; Fowler, B.A. Concurrent exposure to lead, cadmium, and arsenic. Effects on toxicity and tissue metal concentrations in the rat. J. Lab. Clin. Med. 1981, 98, 463–481. [Google Scholar] [PubMed]
- Julshman, K. Analysis of Major and Minor Elements in Mollusks from Norway. Ph.D. Thesis, Institute of Nutrition Direction of Bergen Nygardsangen, Bergen University, Bergen, Norway, 1983. [Google Scholar]
- Suvarna, K.; Layton, C. The Hematoxylin and eosin, Connective and mesenchymal tissues with their stains, Immunohistochemical techniques and Transmission electron microscopy. In Bancroft’s Theory and Practice of Histological Technique; Livingstone, C., Ed.; Elsevier: Oxford, UK, 2013. [Google Scholar]
- Mawed, S.A.; He, Y.; Zhang, J.; Mei, J. Strategy of Hepatic Metabolic Defects Induced by beclin1 Heterozygosity in Adult Zebrafish. Int. J. Mol. Sci. 2020, 21, 1533. [Google Scholar] [CrossRef]
- Dulta, K.; Koşarsoy Ağçeli, G.; Chauhan, P.; Jasrotia, R.; Chauhan, P.K. A Novel Approach of Synthesis Zinc Oxide Nanoparticles by Bergenia ciliata Rhizome Extract: Antibacterial and Anticancer Potential. J. Inorg. Organomet. Polym. Mater. 2021, 31, 180–190. [Google Scholar] [CrossRef]
- Iravani, S.; Korbekandi, H.; Mirmohammadi, S.V.; Zolfaghari, B. Synthesis of silver nanoparticles: Chemical, physical and biological methods. Res. Pharm. Sci. 2014, 9, 385–406. [Google Scholar]
- Ezealisiji, K.M.; Siwe-Noundou, X.; Maduelosi, B.; Nwachukwu, N.; Krause, R.W.M. Green synthesis of zinc oxide nanoparticles using Solanum torvum (L) leaf extract and evaluation of the toxicological profile of the ZnO nanoparticles–hydrogel composite in Wistar albino rats. Intern. Nano Lett. 2019, 9, 99–107. [Google Scholar] [CrossRef]
- Talam, S.; Karumuri, S.R.; Gunnam, N. Synthesis, Characterization, and Spectroscopic Properties of ZnO Nanoparticles. ISRN Nanotec. 2012, 2012, 372505. [Google Scholar] [CrossRef]
- Agarwal, H.; Nakara, A.; Menon, S.; Shanmugam, V. Eco-friendly synthesis of zinc oxide nanoparticles using Cinnamomum Tamala leaf extract and its promising effect towards the antibacterial activity. J. Drug Deliv. Sci. Technol. 2019, 53, 101212. [Google Scholar] [CrossRef]
- Kime, D.E. The effects of pollution on reproduction in fish. Rev. Fish Biol. Fish. 1995, 5, 52–95. [Google Scholar] [CrossRef]
- Reading, B.; Sullivan, C. The Reproductive Organs and Processes: Vitellogenesis in Fishes. Encyclopedia of Fish Physiology 2011, 1, 635–646. [Google Scholar] [CrossRef]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria mediated apoptosis in human liver cells (HepG2). Apoptosis 2012, 17, 852–870. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K.; Khan, M.N.; Jabeen, F.; Kosour, N.; Chaudhry, A.S.; Sohail, M.; Ahmad, N. Toxicity of zinc oxide nanoparticles (ZnO-NPs) in tilapia (Oreochromis mossambicus): Tissue accumulation, oxidative stress, histopathology and genotoxicity. Int. J. Environ. Sci. Technol. 2019, 16, 1973–1984. [Google Scholar] [CrossRef]
- Yao, Y.; Zang, Y.; Qu, J.; Tang, M.; Zhang, T. The Toxicity Of Metallic Nanoparticles On Liver: The Subcellular Damages, Mechanisms, And Outcomes. Int. J. Nanomed. 2019, 14, 8787–8804. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhu, L.; Duan, Z.; Qi, R.; Li, Y.; Lang, Y. Comparative toxicity of several metal oxide nanoparticle aqueous suspensions to Zebrafish (Danio rerio) early developmental stage. J. Environ. Sci. Health Part A Toxic/Hazard. Subst. Environ. Eng. 2008, 43, 278–284. [Google Scholar] [CrossRef]
- Tavabe, K.R.; Kuchaksaraei, B.S.; Javanmardi, S. Effects of ZnO nanoparticles on the Giant freshwater prawn (Macrobrachium rosenbergii, de Man, 1879): Reproductive performance, larvae development, CHH concentrations and anti-oxidative enzymes activity. Anim. Reprod. Sci. 2020, 221, 106603. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, X.; Zhang, X.; Zhao, Z.; Liu, H.; George, R.; Wilson-Rawls, J.; Chang, Y.; Chen, Y. Disruption of zebrafish (Danio rerio) reproduction upon chronic exposure to TiO2 nanoparticles. Chemosphere 2011, 83, 461–467. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, B.; Hong, W.; Chen, L.; Yao, L.; Zhao, Y.; Aguilar, Z.P.; Xu, H. ZnO Nanoparticles Induced Male Reproductive Toxicity Based on the Effects on the Endoplasmic Reticulum Stress Signaling Pathway. Int. J. Nanomed. 2019, 14, 9563–9576. [Google Scholar] [CrossRef]
- Bacchetta, R.; Santo, N.; Marelli, M.; Nosengo, G.; Tremolada, P. Chronic toxicity effects of ZnSO4 and ZnO nanoparticles in Daphnia magna. Environ. Res. 2017, 152, 128–140. [Google Scholar] [CrossRef]
- Sayadi, M.H.; Pavlaki, M.D.; Martins, R.; Mansouri, B.; Tyler, C.R.; Kharkan, J.; Shekari, H. Bioaccumulation and toxicokinetics of zinc oxide nanoparticles (ZnO NPs) co-exposed with graphene nanosheets (GNs) in the blackfish (Capoeta fusca). Chemosphere 2021, 269, 128689. [Google Scholar] [CrossRef]
- Kaya, H.; Aydın, F.; Gürkan, M.; Yılmaz, S.; Ates, M.; Demir, V.; Arslan, Z. Effects of zinc oxide nanoparticles on bioaccumulation and oxidative stress in different organs of tilapia (Oreochromis niloticus). Environ. Toxicol. Pharmacol. 2015, 40, 936–947. [Google Scholar] [CrossRef] [PubMed]
- Johnston, B.D.; Scown, T.M.; Moger, J.; Cumberland, S.A.; Baalousha, M.; Linge, K.; van Aerle, R.; Jarvis, K.; Lead, J.R.; Tyler, C.R. Bioavailability of Nanoscale Metal Oxides TiO2, CeO2, and ZnO to Fish. Environ. Sci. Technol. 2010, 44, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Choy, J.H. Biokinetics of zinc oxide nanoparticles: Toxicokinetics, biological fates, and protein interaction. Int. J. Nanomed. 2014, 9 Suppl 2, 261–269. [Google Scholar] [CrossRef]
- Suganthi, P.; Murali, M.; Athif, P.; Sadiq Bukhari, A.; Syed Mohamed, H.E.; Basu, H.; Singhal, R.K. Haemato-immunological studies in ZnO and TiO2 nanoparticles exposed euryhaline fish, Oreochromis mossambicus. Environ. Toxicol. Pharmacol. 2019, 66, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, S.; You, H.; Jiang, R.; Zhuang, C.; Zhang, X. Developmental toxicity and DNA damage to zebrafish induced by perfluorooctane sulfonate in the presence of ZnO nanoparticles. Environ. Toxicol. 2016, 31, 360–371. [Google Scholar] [CrossRef]
- Liu, X.Q.; Zhang, H.F.; Zhang, W.D.; Zhang, P.F.; Hao, Y.N.; Song, R.; Li, L.; Feng, Y.N.; Hao, Z.H.; Shen, W.; et al. Regulation of neuroendocrine cells and neuron factors in the ovary by zinc oxide nanoparticles. Toxicol. Lett. 2016, 256, 19–32. [Google Scholar] [CrossRef]
- Akbulut, C.; Kotil, T.; Yön, N. Ultrastructural evaluation of oocyte envelopes of zebrafish (Danio Rerio) (Hamilton, 1822) after TiO2 nanoparticle exposure. Arch. Biol. Sci. 2017, 70, 35. [Google Scholar] [CrossRef]
- Ramsden, C.S.; Henry, T.B.; Handy, R.D. Sub-lethal effects of titanium dioxide nanoparticles on the physiology and reproduction of zebrafish. Aquat. Toxicol. 2013, 126, 404–413. [Google Scholar] [CrossRef]
- Han, Z.; Yan, Q.; Ge, W.; Liu, Z.G.; Gurunathan, S.; De Felici, M.; Shen, W.; Zhang, X.F. Cytotoxic effects of ZnO nanoparticles on mouse testicular cells. Int. J. Nanomed. 2016, 11, 5187–5203. [Google Scholar] [CrossRef]
- Chen, G.-H.; Song, C.-C.; Zhao, T.; Hogstrand, C.; Wei, X.-L.; Lv, W.-H.; Song, Y.-F.; Luo, Z. Mitochondria-Dependent Oxidative Stress Mediates ZnO Nanoparticle (ZnO NP)-Induced Mitophagy and Lipotoxicity in Freshwater Teleost Fish. Environ. Sci. Technol. 2022, 56, 2407–2420. [Google Scholar] [CrossRef]
- Rajkumar, K.S.; Sivagaami, P.; Ramkumar, A.; Murugadas, A.; Srinivasan, V.; Arun, S.; Senthil Kumar, P.; Thirumurugan, R. Bio-functionalized zinc oxide nanoparticles: Potential toxicity impact on freshwater fish Cyprinus carpio. Chemosphere 2022, 290, 133220. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Jiang, H.; Xu, G.; Li, C.; Li, D.; Tang, S. Lethality of Zinc Oxide Nanoparticles Surpasses Conventional Zinc Oxide via Oxidative Stress, Mitochondrial Damage and Calcium Overload: A Comparative Hepatotoxicity Study. Int. J. Mol. Sci. 2022, 23, 6724. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Hong, W.; Tang, Y.; Zhao, Y.; Aguilar, Z.P.; Xu, H. Protective effect of the NAC and Sal on zinc oxide nanoparticles-induced reproductive and development toxicity in pregnant mice. Food Chem. Toxicol. 2020, 143, 111552. [Google Scholar] [CrossRef]
- Yu, K.N.; Yoon, T.J.; Minai-Tehrani, A.; Kim, J.E.; Park, S.J.; Jeong, M.S.; Ha, S.W.; Lee, J.K.; Kim, J.S.; Cho, M.H. Zinc oxide nanoparticle induced autophagic cell death and mitochondrial damage via reactive oxygen species generation. Toxicol. Vitr. 2013, 27, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Singh, S.K.; Chauhan, L.K.; Das, M.; Tripathi, A.; Dwivedi, P.D. Zinc oxide nanoparticles induce apoptosis by enhancement of autophagy via PI3K/Akt/mTOR inhibition. Toxicol. Lett. 2014, 227, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Klionsky, D.J. Protein turnover via autophagy: Implications for metabolism. Annu. Rev. Nutr. 2007, 27, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, F.; Li, Y.; Shan, X.; Yin, L.; Hao, X.; Zhong, Y. More serious autophagy can be induced by ZnO nanoparticles than single-walled carbon nanotubes in rat tracheal epithelial cells. Environ. Toxicol. 2021, 36, 238–248. [Google Scholar] [CrossRef]
- Shen, J.; Yang, D.; Zhou, X.; Wang, Y.; Tang, S.; Yin, H.; Wang, J.; Chen, R.; Chen, J. Role of Autophagy in Zinc Oxide Nanoparticles-Induced Apoptosis of Mouse Leydig Cells. Int. J. Mol. Sci. 2019, 20, 4042. [Google Scholar] [CrossRef]
- Wang, J.; Deng, X.; Zhang, F.; Chen, D.; Ding, W. ZnO nanoparticle-induced oxidative stress triggers apoptosis by activating JNK signaling pathway in cultured primary astrocytes. Nanoscale Res. Lett. 2014, 9, 117. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Raja, M.; Ahmad, I.; Siddiqui, M.K.; AlSalhi, M.S.; Alrokayan, S.A. ZnO nanorod-induced apoptosis in human alveolar adenocarcinoma cells via p53, survivin and bax/bcl-2 pathways: Role of oxidative stress. Nanomed. 2011, 7, 904–913. [Google Scholar] [CrossRef]
- Chen, S.X.; Yang, X.Z.; Deng, Y.; Huang, J.; Li, Y.; Sun, Q.; Yu, C.-P.; Zhu, Y.; Hong, W.S. Silver nanoparticles induce oocyte maturation in zebrafish (Danio rerio). Chemosphere 2017, 170, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Applerot, G.; Lipovsky, A.; Dror, R.; Perkas, N.; Nitzan, Y.; Lubart, R.; Gedanken, A. Enhanced Antibacterial Activity of Nanocrystalline ZnO Due to Increased ROS-Mediated Cell Injury. Adv. Funct. Mater. 2009, 19, 842–852. [Google Scholar] [CrossRef]
- Mishra, P.K.; Mishra, H.; Ekielski, A.; Talegaonkar, S.; Vaidya, B. Zinc oxide nanoparticles: A promising nanomaterial for biomedical applications. Drug Discov. Today 2017, 22, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Peng, X.; Mei, S. Autophagy in Ovarian Follicular Development and Atresia. Int. J. Biol. Sci. 2019, 15, 726–737. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.K.; Gupta, A.; Sharma, A.; Yadav, A.K.; Tiwari, M.; Pandey, A.N.; Prasad, S.; Shrivastav, T.G.; Chaube, S.K. Fate of the germ cells in mammalian ovary: A review. J. Reprod. Health. Med. 2020, 1. [Google Scholar] [CrossRef]
- Di Cerbo, A.; Palmieri, B. Review: The market of probiotics. Pak. J. Pharm. Sci. 2015, 28, 2199–2206. [Google Scholar]
- Peluso, J.J. Progesterone signaling mediated through progesterone receptor membrane component-1 in ovarian cells with special emphasis on ovarian cancer. Steroids 2011, 76, 903–909. [Google Scholar] [CrossRef]
- Peluso, J.J.; Gawkowska, A.; Liu, X.; Shioda, T.; Pru, J.K. Progesterone receptor membrane component-1 regulates the development and Cisplatin sensitivity of human ovarian tumors in athymic nude mice. Endocrinology 2009, 150, 4846–4854. [Google Scholar] [CrossRef]
- Ahmed, I.S.; Rohe, H.J.; Twist, K.E.; Mattingly, M.N.; Craven, R.J. Progesterone receptor membrane component 1 (Pgrmc1): A heme-1 domain protein that promotes tumorigenesis and is inhibited by a small molecule. J. Pharmacol. Exp. Ther. 2010, 333, 564–573. [Google Scholar] [CrossRef]
- Lin, S.-T.; Chou, H.-C.; Chang, S.-J.; Chen, Y.-W.; Lyu, P.-C.; Wang, W.-C.; Chang, M.D.-T.; Chan, H.-L. Proteomic analysis of proteins responsible for the development of doxorubicin resistance in human uterine cancer cells. J. Proteom. 2012, 75, 5822–5847. [Google Scholar] [CrossRef]
- Patiño, R.; Thomas, P.; Yoshizaki, G. Ovarian follicle maturation and ovulation: An integrated perspective. Fish Physiol. Biochem. 2003, 28, 305–308. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, D.; Shaner, Z.C.; Chen, S.; Hong, W.; Stellwag, E.J. Nuclear progestin receptor (pgr) knockouts in zebrafish demonstrate role for pgr in ovulation but not in rapid non-genomic steroid mediated meiosis resumption. Front. Endocrinol. (Lausanne) 2015, 6, 37. [Google Scholar] [CrossRef]
- Morselli, E.; Tasdemir, E.; Maiuri, M.C.; Galluzzi, L.; Kepp, O.; Criollo, A.; Vicencio, J.M.; Soussi, T.; Kroemer, G. Mutant p53 protein localized in the cytoplasm inhibits autophagy. Cell Cycle 2008, 7, 3056–3061. [Google Scholar] [CrossRef] [PubMed]
- Petitjean, A.; Achatz, M.I.; Borresen-Dale, A.L.; Hainaut, P.; Olivier, M. TP53 mutations in human cancers: Functional selection and impact on cancer prognosis and outcomes. Oncogene 2007, 26, 2157–2165. [Google Scholar] [CrossRef] [PubMed]
- Raz, E. The function and regulation of vasa-like genes in germ-cell development. Genome Biol. 2000, 1, reviews1017. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, E.A.; Wessel, G.M. Vasa genes: Emerging roles in the germ line and in multipotent cells. Bioessays 2010, 32, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Sudo, T.; Mikami, Y.; Otani, M.; Takano, M.; Tsuda, H.; Itamochi, H.; Katabuchi, H.; Ito, M.; Nishimura, R. Germ cell specific protein VASA is over-expressed in epithelial ovarian cancer and disrupts DNA damage-induced G2 checkpoint. Gynecol. Oncol. 2008, 111, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Poon, J.; Wessel, G.M.; Yajima, M. An unregulated regulator: Vasa expression in the development of somatic cells and in tumorigenesis. Dev. Biol. 2016, 415, 24–32. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Accession Number in NCBI | Primer Sequences |
|---|---|---|
| Reference Gene | ||
| βactin | NM_131031 | F: 5′ ATGGATGAGGAAATCGCTGC 3′ |
| R:5′CTTTCTGTCCCATGCCAACC 3′ | ||
| Antioxidant enzymes | ||
| Superoxide dismutase1 (sod1) | NM_131294 | F: 5′ CGCACTTCAACCCTCATGAC 3′ |
| R: 5′ TGAATCACCATGGTCCTCCC 3′ | ||
| Superoxide dismutase2 (sod2) | NM_199976 | F:5′CCTCCAGACAGAAGCA 3′ |
| R:5′CTGAAATGAGCCAAAGT 3′ | ||
| Glutathione peroxidase 1a (gpx1a) | NM_001007281 | F:5′GCACAACAGTCAGGGAT 3′ |
| R:5′TCAGGAACGCAAACAG 3′ | ||
| Glutathione S-transferase pi 1.2 (gstp1.2) | NM_131734 | F:5′CCAACCACCTCAAATGCT 3′ |
| R:5′ACGGGAAAGAGTCCAGACAG 3′ | ||
| Catalase (cat) | NM_130912 | F:5′TGTGGAAGGAGGGTCG 3′ |
| R:5′CTTTGGCTTTGGAGTAG 3′ | ||
| Autophagy-related genes | ||
| Autophagy-related gene-7 homolog (atg7) | XM 021479676 | F:5′ACGGTGATGCTGTTGGTCTG 3′ |
| R: 5′ TTTGTCGGTGGATTTGAAGG 3′ | ||
| Autophagy-related gene-5 homolog (atg5) | NM_205618 | F: 5′ TGGAGTATCCCACCGAAGA3′ |
| R:5′ CACTGGTCGGAAGAGC3′ | ||
| Autophagy-related gene-12 homolog (atg12) | NM_001246200 | F: 5′ TCATCTCACGCTTCCTCAA 3′ |
| R: 5′ TCACTTCCGAAACACTCAAA 3′ | ||
| Sequestosome 1 (sqstm1) (p62) | NM_001312913 | F: 5′ TGGTGCTACTGCCTCTTCTCA 3′ |
| R: 5′ GGGTTACTTTGGTCCGCTTT 3′ | ||
| Apoptosis-related genes | ||
| Apoptosis-inducing factor (siva1) | NM_001327928 | F: 5′ CCGCTACCGACAGGAGATCTACGA 3′ |
| R: 5′ GGTGTGGAGCGCGCTCTGTGCAGT 3′ | ||
| BCL2 associated X, apoptosis regulator (baxa) | NM_131562 | F: 5′ GACAGGGATGCTGAAGTGA 3′ |
| R: 5′ TGAGTCGGCTGAAGATTAGA 3′ | ||
| Caspase a (caspa) | NM_131505 | F: 5′ GACGGTGAGCCTGATGAGCCAA 3′ |
| R: 5′ CCTGAACAGTTCCTCGATGTGA 3′ | ||
| Oocyte maturation genes | ||
| Methyltransferase like 3 (mettl3) | NM_212780 | F: 5′ CCTAGAGCTGCTGAATACCAGT 3′ |
| R: 5′ GATGATTCGCCTGAAGTGC 3′ | ||
| Progesterone receptor membrane component-1 (pgrmc1) | NM_001007392 | F: 5′ CAGACTATGGCCCGGTTGAGGAG 3′ |
| R: 5′ CTGCATGGCATTGAGATCGG 3′ | ||
| Zygote arrest 1 (zar1) | NM_194381 | F: 5′ CAACCCGAAGACCGAC 3′ |
| R: 5′ CACCACCGCTGCTGAC 3′ | ||
| Gonadal soma-derived factor variant 2 (gsdf) | NM_001114668 | F: 5′ GCTCCATCCGTCACCT 3′ |
| R: 5′ TCACCGTAGACAGAACCAG 3′ | ||
| SRY-box transcription factor 3 (sox3) | NM_001001811 | F: 5′ ATTCCGCAGTCCAACA 3′ |
| R: 5′ TTCTCCTGAGCCATCTTC 3′ | ||
| Ovulation-related genes | ||
| Metallopeptidase with thrombospondin type 1 motif, 15a (adamts15a) | NM_001126429 | F:5′GAGAGCAAAGATAACAAGGCACAAA3′ |
| R: 5′TTTTCCACCTTTATTGACTCCACCT3′ | ||
| Luteinizing hormone/choriogonadotropin receptor (lhcgr) | NM_205625 | F: 5′ CGCTCTGATCAACTGGGACA 3′ |
| R: 5′ GGCGCTGTTGGCATAAATCC 3′ | ||
| Progesterone receptor (pgr) | NM_001166335 | F: 5′ ACAGACAGCATACACCGC3′ |
| R: 5′TCCACAGGTCAGAACTCC3′ | ||
| Follicle-stimulating hormone receptor (fshr) | NM_001001812 | F: 5′ CAAATGCGTCTACGCCATGC3′ |
| R: 5′AAAGCGGGATTACGGACGGT3′ | ||
| Follistatin a (fsta) | NM_131037 | F: 5′ CATCAAGGCCAAGTCATGCG3′ |
| R: 5′GCCTGCTTCATGGCACACTC3′ | ||
| Groups | Zn Residues (μg/g Wet Weight) |
|---|---|
| control | 15.83 ± 1.57 c |
| T1 | 32.10 ± 4.01 b |
| T2 | 57.85 ± 1.78 a |
| SEM | 6.26 |
| p-value | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mawed, S.A.; Marini, C.; Alagawany, M.; Farag, M.R.; Reda, R.M.; El-Saadony, M.T.; Elhady, W.M.; Magi, G.E.; Di Cerbo, A.; El-Nagar, W.G. Zinc Oxide Nanoparticles (ZnO-NPs) Suppress Fertility by Activating Autophagy, Apoptosis, and Oxidative Stress in the Developing Oocytes of Female Zebrafish. Antioxidants 2022, 11, 1567. https://doi.org/10.3390/antiox11081567
Mawed SA, Marini C, Alagawany M, Farag MR, Reda RM, El-Saadony MT, Elhady WM, Magi GE, Di Cerbo A, El-Nagar WG. Zinc Oxide Nanoparticles (ZnO-NPs) Suppress Fertility by Activating Autophagy, Apoptosis, and Oxidative Stress in the Developing Oocytes of Female Zebrafish. Antioxidants. 2022; 11(8):1567. https://doi.org/10.3390/antiox11081567
Chicago/Turabian StyleMawed, Suzan Attia, Carlotta Marini, Mahmoud Alagawany, Mayada R. Farag, Rasha M. Reda, Mohamed T. El-Saadony, Walaa M. Elhady, Gian E. Magi, Alessandro Di Cerbo, and Wafaa G. El-Nagar. 2022. "Zinc Oxide Nanoparticles (ZnO-NPs) Suppress Fertility by Activating Autophagy, Apoptosis, and Oxidative Stress in the Developing Oocytes of Female Zebrafish" Antioxidants 11, no. 8: 1567. https://doi.org/10.3390/antiox11081567