
Beneficial Effects of Standardized Extracts from Wastes of Red Oranges and Olive Leaves

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Industrial Extraction
2.2. HPLC–PDA Analyses of Anthocyanins, Hydroxycinnamic Acids and Flavanones of ROE
2.3. Determination of Total Polyphenols Content of OLE
2.4. HPLC–PDA Analysis of Oleuropein of OLE
2.5. Inhibition of DPPH
2.6. Cell Culture and Cell Viability Assay
2.7. Cellular Model of Hepatic Steatosis
2.8. Determination of Heme Oxygenase-1 (HO-1) Levels (ELISA)
2.9. Oil Red O’Staining
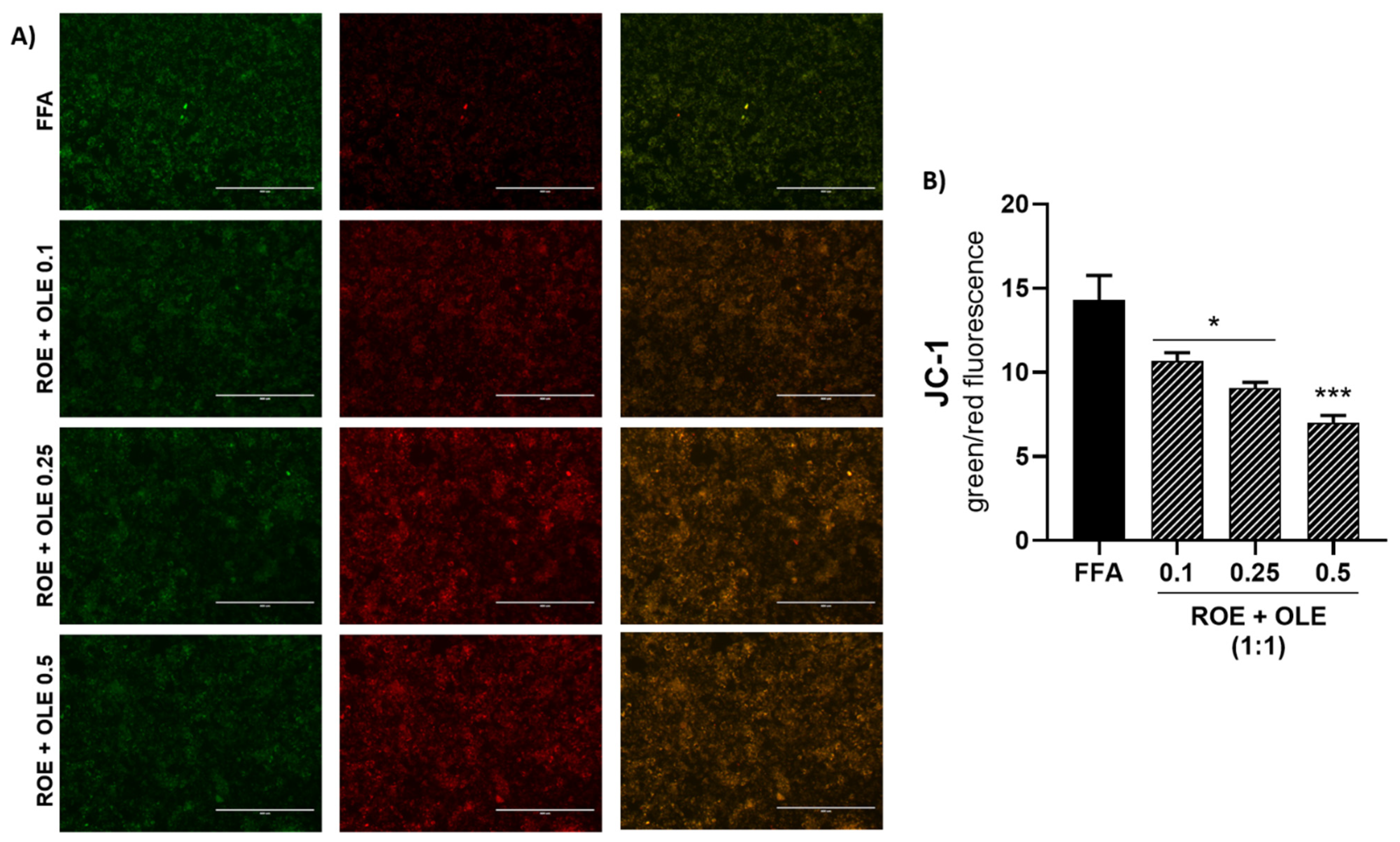
2.10. Measurement of Mitochondrial Membrane Potential
2.11. Thiol (RSH) Group Determination
2.12. Measurement of Lipid Peroxidation
2.13. Measurement of HMG-CoA Reductase Activity
2.14. Statistical Analysis
3. Results
3.1. Chemical Comosition of ROE and OLE
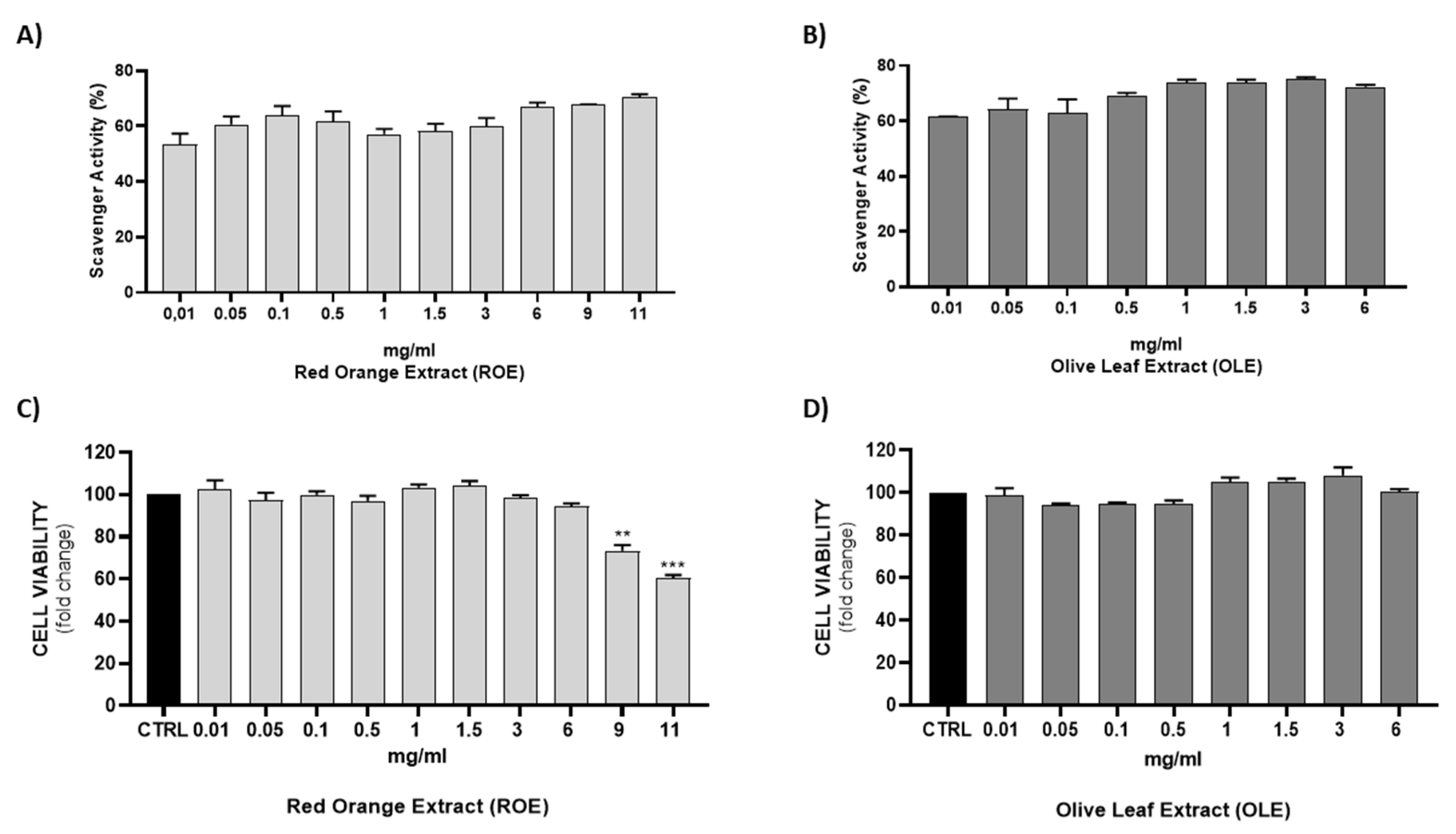
3.2. Free Radical Scavenging Activity of ROE and OLE
3.3. Effect of ROE and OLE on Cell Viability
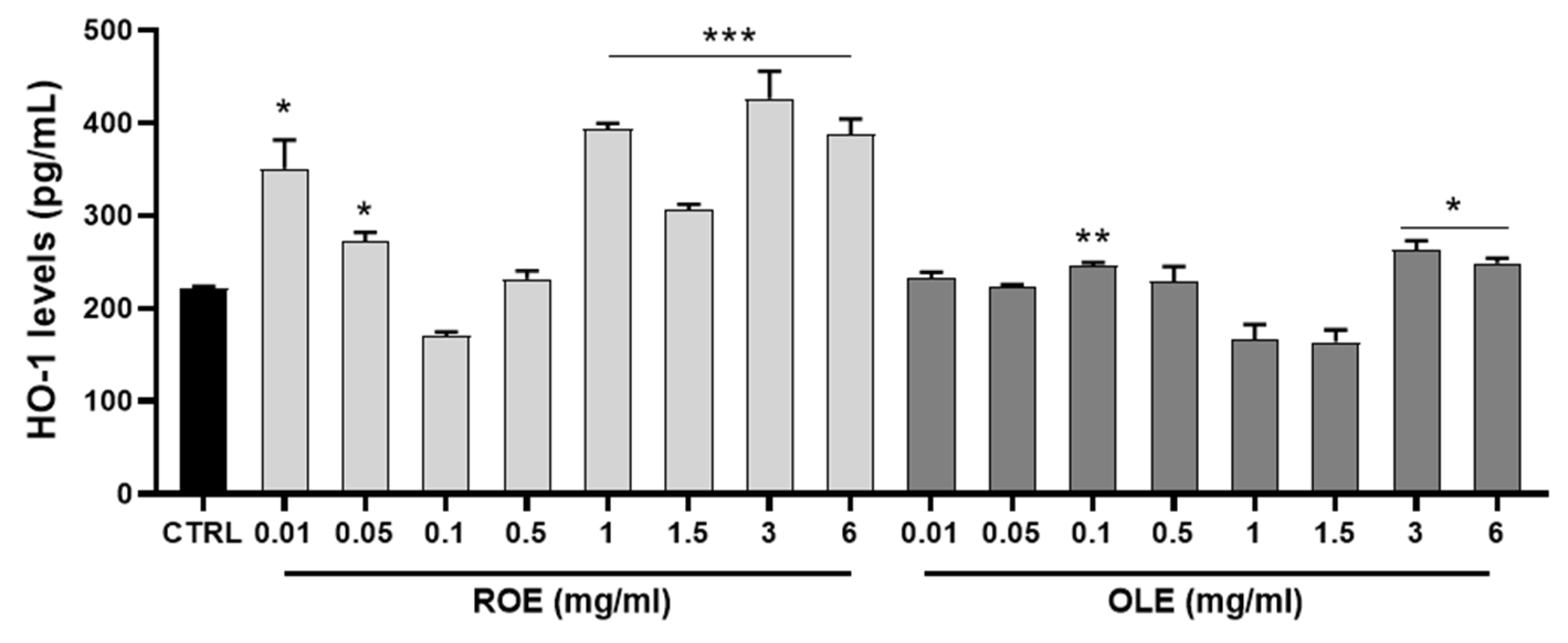
3.4. Antioxidant Activity of ROE and OLE Mediated by HO-1 Induction
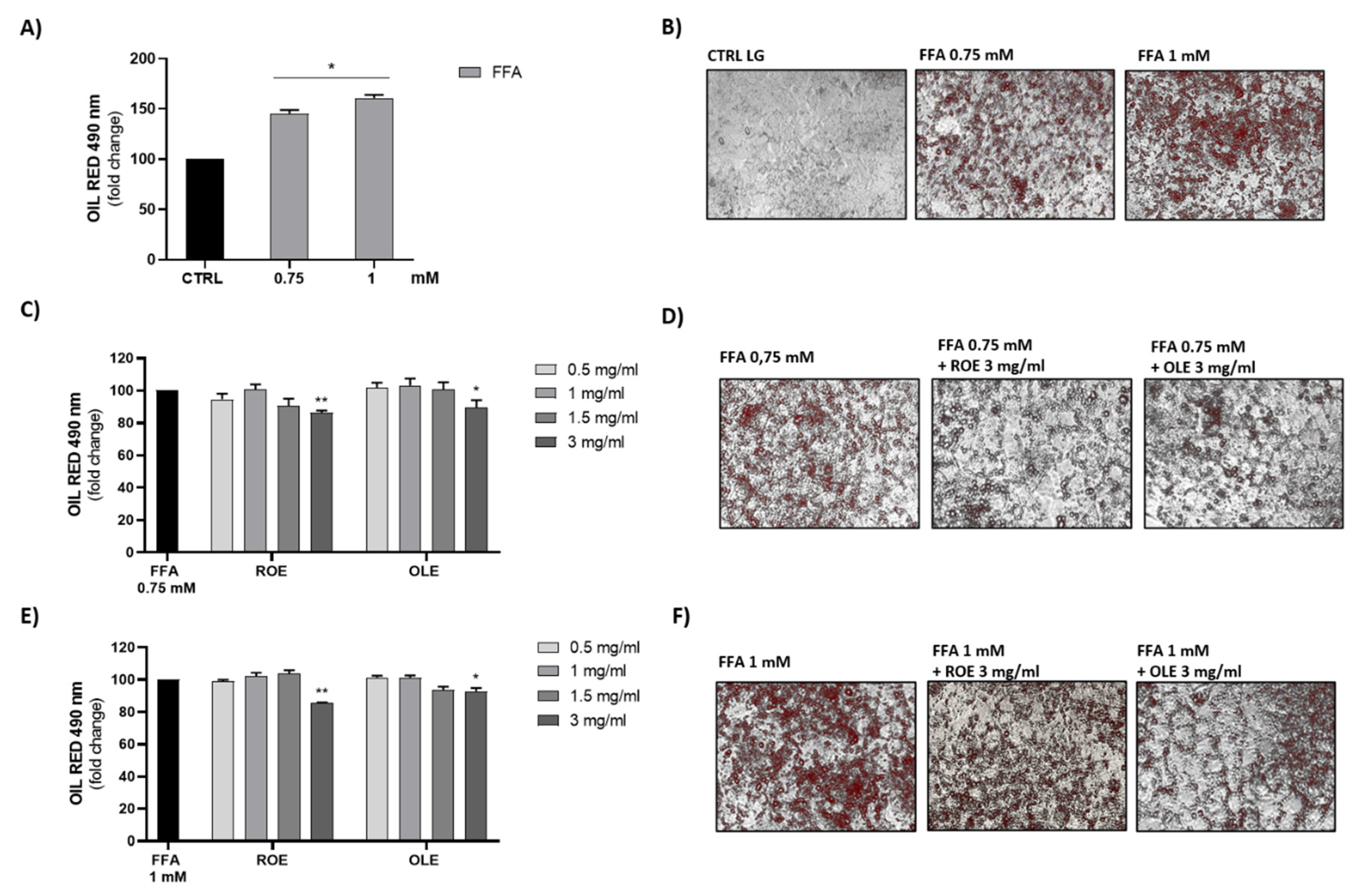
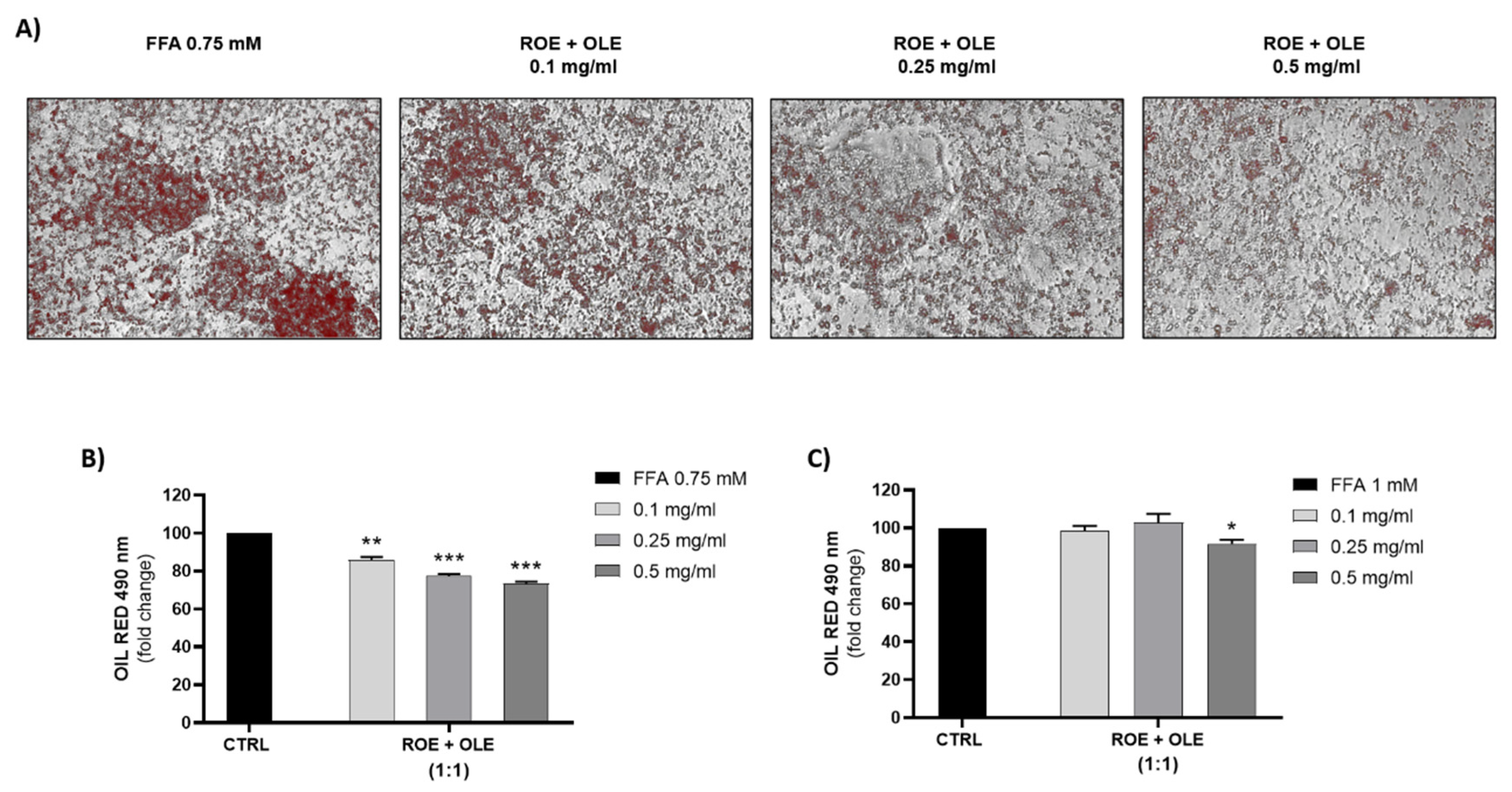
3.5. Effect of ROE and OLE on HepG2 Fatty Storage
3.6. Effect of ROE and OLE on Oxidative Damage and Mitochondrial Dysfunction
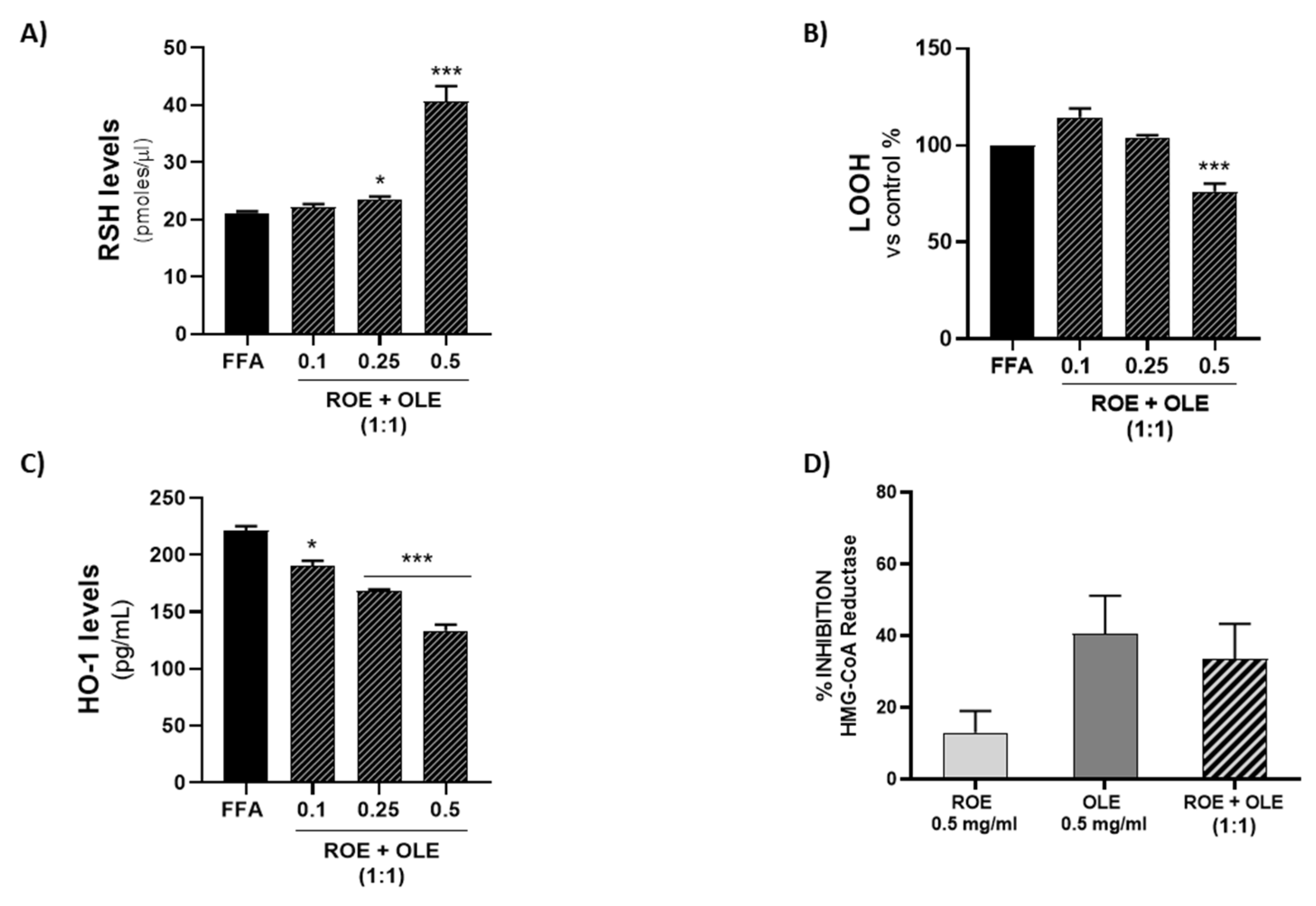
3.7. Effect of ROE and OLE on Thiol (RSH) Group Levels
3.8. Effect of ROE and OLE on LOOH Levels
3.9. Effect of ROE and OLE on HO-1 Expression in Steatosis Model
3.10. Effect of ROE and OLE on HMG-CoA Reductase Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maina, S.; Kachrimanidou, V.; Apostolis, K. A roadmap towards a circular and sustainable bioeconomy through waste valorization. Curr. Opin. Green Sustain. Chem. 2017, 8, 18–23. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, M.; Adhikari, B. Handbook of Food Powders: Processes and Properties; Woodhead Publishing: Cambridge, UK, 2013. [Google Scholar]
- Karam, M.C.; Petit, J.; Zimmer, D.; Elie; Joël, S. Effects of drying and grinding in production of fruit and vegetable powders: A review. J. Food Eng. 2016, 188, 32–49. [Google Scholar] [CrossRef]
- Neacsu, M.; Vaughan, N.; Raikos, V.; Multari, S.; Duncan, G.J.; Duthie, G.G.; Russell, W.R. Phytochemical profile of commercially available food plant powders: Their potential role in healthier food reformulations. Food Chem. 2015, 179, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Bas-Bellver, C.; Barrera, C.; Betoret, N.; Seguí, L. Turning Agri-Food Cooperative Vegetable Residues into Functional Powdered Ingredients for the Food Industry. Sustainability 2020, 12, 1284. [Google Scholar] [CrossRef] [Green Version]
- Sorrenti, V.; Di Giacomo, C.; Russo, A.; Acquaviva, R.; Barcellona, M.L.; Vanella, A. Inhibition of LDL Oxidation by Red Orange (Citrus sinensis) Extract and its Active Components. J. Food Sci. 2006, 69, C480–C484. [Google Scholar] [CrossRef]
- Sorrenti, V.; Di Giacomo, C.; Acquaviva, R.; Cosenza, J.; Carota, G.; Galvano, F. Blond and blood juice supplementation in high fat diet fed mice: Effect on antioxidant status and DDAH/ADMA pathway. RSC Adv. 2019, 9, 11406–11412. [Google Scholar] [CrossRef] [Green Version]
- Russo, M.; Bonaccorsi, I.; Inferrera, V.; Dugo, P.; Luigi, M. Underestimated sources of flavonoids, limonoids and dietary fiber: Availability in orange’s by-products. J. Funct. Foods 2015, 12, 150–157. [Google Scholar] [CrossRef]
- Vergara-Barberán, M.; Navarro-Huerta, J.A.; Torres-Lapasió, J.R.; Simó-Alfonso, E.F.; García-Alvarez-Coque, M.C. Classification of olive leaves and pulp extracts by comprehensive two-dimensional liquid chromatography of polyphenolic fingerprints. Food Chem. 2020, 320, 126630. [Google Scholar] [CrossRef]
- Suri, S.; Singh, A.; Prabhat, K.N. Current applications of citrus fruit processing waste: A scientific outlook. Appl. Food Res. 2022, 2, 100050. [Google Scholar] [CrossRef]
- Espeso, J.; Isaza, A.; Lee, J.Y.; Sörensen, P.M.; Jurado, P.; Avena-Bustillos, R.d.J.; Olaizola, M.; Arboleya, J.C. Olive Leaf Waste Management. Front. Sustain. Food Syst. 2021, 5, 660582. [Google Scholar] [CrossRef]
- Acar-Tek, N.; Ağagündüz, D. Olive Leaf (Olea europaea L. folium): Potential Effects on Glycemia and Lipidemia. Ann. Nutr. Metab. 2020, 76, 10–15. [Google Scholar] [CrossRef]
- Fabroni, S.; Ballistreri, G.; Amenta, M.; Rapisarda, P. Anthocyanins in different Citrus species: An UHPLC-PDA-ESI/MS. J. Sci. Food Agric. 2016, 96, 4797–4808. [Google Scholar] [CrossRef] [PubMed]
- Rapisarda, P.; Bianco, M.L.; Pannuzzo, P.; Nicolina, T. Effect of cold storage on vitamin C, phenolics and antioxidant activity of five orange genotypes Citrus sinensis (L.) Osbeck. Postharvest Biol. Technol. 2008, 49, 348–354. [Google Scholar] [CrossRef]
- Rouseff, L.R. Quantitative Survey of Narirutin, Naringin, Hesperidin, and Neohesperidin in Citrus. J. Agric. Food Chem. 1987, 35, 1027–1030. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M.; Lester, P. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999. [Google Scholar] [CrossRef]
- D’Antuono, I.; Kontogianni, V.G.; Kotsiou, K.; Linsalata, V.; Logrieco, A.F.; Tasioula-Margari, M.; Cardinali, A. Polyphenolic characterization of olive mill wastewaters, coming from Italian and Greek olive cultivars, after membrane technology. Food Res. Int. 2014, 65, 301–310. [Google Scholar] [CrossRef]
- Swapna Sasi, U.S.; Sindhu, G.; Raghu, K.G. Fructose-palmitate based high calorie induce steatosis in HepG2 cells via mitochondrial dysfunction: An in vitro approach. Toxicol. Vitr. 2020, 68, 104952. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Bendary, E.; Francis, R.R.; Ali, H.M.G.; Sarwat, M.I.; El Hady, S. Antioxidant and structure–activity relationships (SARs) of some phenolic and anilines compounds. Ann. Agric. Sci. 2013, 58, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Hahn, D.; Shin, S.H.; Bae, J.-S. Natural Antioxidant and Anti-Inflammatory Compounds in Foodstuff or Medicinal Herbs Inducing Heme Oxygenase-1 Expression. Antioxidants 2020, 9, 1191. [Google Scholar] [CrossRef] [PubMed]
- Chiang, S.K.; Chen, S.E.; Chang, L.C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef] [Green Version]
- Drummond, G.S.; Baum, J.; Greenberg, M.; Lewis, D.; Abraham, N.G. HO-1 overexpression and underexpression: Clinical implications. Arch. Biochem. Biophys. 2019, 673, 108073. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Diaconeasa, Z.; Cătoi, A.-F.; Vodnar, D.C. Screening of Ten Tomato Varieties Processing Waste for Bioactive Components and Their Related Antioxidant and Antimicrobial Activities. Antioxidants 2019, 8, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrenti, V.; Randazzo, C.L.; Caggia, C.; Ballistreri, G.; Romeo, F.V.; Fabroni, S.; Timpanaro, N.; Raffaele, M.; Vanella, L. Beneficial Effects of Pomegranate Peel Extract and Probiotics on Pre-adipocyte Differentiation. Front. Microbiol. 2019, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, D.S.; An, T.H.; Park, H.-J.; Kim, W.K.; Bae, K.-H.; Oh, K.-J. Metabolic Spectrum of Liver Failure in Type 2 Diabetes and Obesity: From NAFLD to NASH to HCC. Int. J. Mol. Sci. 2021, 22, 4495. [Google Scholar] [CrossRef] [PubMed]
- Moravcová, A.; Červinková, Z.; Kučera, O.; Mezera, V.; Rychtrmoc, D.; Lotková, H. The effect of oleic and palmitic acid on induction of steatosis and cytotoxicity on rat hepatocytes in primary culture. Physiol. Res. 2015, 64, S627–S636. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.C.; Amankwa-Sakyi, M.; Flynn, T.J. Cellular glutathione in fatty liver in vitro models. Toxicol. Vitr. 2011, 25, 1501–1506. [Google Scholar] [CrossRef]
- Raffaele, M.; Carota, G.; Sferrazzo, G.; Licari, M.; Barbagallo, I.; Sorrenti, V.; Signorelli, S.S.; Vanella, L. Inhibition of Heme Oxygenase Antioxidant Activity Exacerbates Hepatic Steatosis and Fibrosis In Vitro. Antioxidants 2019, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, A.L.; Traverso, N.; Domenicotti, C.; Piras, S.; Moretta, L.; Marinari, U.M.; Pronzato, M.A.; Nitti, M. The Nrf2/HO-1 Axis in Cancer Cell Growth and Chemoresistance. Oxid. Med. Cell. Longev. 2016, 2016, 1958174. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Gambino, R.; Maurizio, C. Cholesterol metabolism and the pathogenesis of non-alcoholic steatohepatitis. Prog. Lipid Res. 2013, 52, 175–191. [Google Scholar] [CrossRef] [Green Version]
- Arguello, G.; Balboa, E.; Arrese, M.; Silvana, Z. Recent insights on the role of cholesterol in non-alcoholic fatty liver disease. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2015, 1852, 1765–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrikx, T.; Walenbergh, S.M.A.; Hofker, M.H.; Shiri-Sverdlov, R. Lysosomal cholesterol accumulation: Driver on the road to inflammation during atherosclerosis and non-alcoholic steatohepatitis. Obes. Rev. 2014, 15, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N. The Role of Cholesterol in the Pathogenesis of NASH. Trends Endocrinol. Metab. 2016, 27, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated Fatty Acids and Risk of Coronary Heart Disease: Modulation by Replacement Nutrients. Curr. Atheroscler. Rep. 2010, 12, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Dongiovanni, P.; Petta, S.; Mannisto, V.; Mancina, R.M.; Pipitone, R.; Karja, V.; Maggioni, M.; Kakela, P.; Wiklund, O.; Mozzi, E.; et al. Statin use and non-alcoholic steatohepatitis in at risk individuals. J. Hepatol. 2015, 63, 705–712. [Google Scholar] [CrossRef]
- Mancini, G.B.J.; Baker, S.; Bergeron, J.; Fitchett, D.; Frohlich, J.; Genest, J.; Gupta, M.; Hegele, R.A.; Ng, D.; Pope, J. Diagnosis, Prevention, and Management of Statin Adverse Effects and Intolerance: Proceedings of a Canadian Working Group Consensus Conference. Can. J. Cardiol. 2011, 27, 635–662. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | ROE | OLE |
|---|---|---|
| Total Polyphenols | not determined | 3.82 g/100 g a |
| Hydroxytyrosol | absent | 0.10 g/100 g |
| Tyrosol | absent | 0.05 g/100 g |
| Oleuropein | absent | 0.40 g/100 g |
| Total Phenols | absent | 0.55 g/100 g b |
| Cyanidin 3,5-diglucoside | 0.06 g/100 g | absent |
| Delphinidin 3-glucoside | 0.02 g/100 g | absent |
| Cyanidin 3-glucoside | 0.31 g/100 g | absent |
| Cyanidin 3-rutinoside | 0.02 g/100 g | absent |
| Petunidin 3-glucoside | 0.02 g/100 g | absent |
| Delphinidin 3-(6″-malonyl)glucoside | 0.02 g/100 g | absent |
| Peonidin 3-glucoside | 0.02 g/100 g | absent |
| Petunidin 3-(6″-malonyl)glucoside | 0.03 g/100 g | absent |
| Cyanidin derivative | 0.03 g/100 g | absent |
| Cyanidin 3-(6″-malonyl)glucoside | 0.27 g/100 g | absent |
| Cyanidin 3-(6″-dioxalyl)glucoside | 0.06 g/100 g | absent |
| Pelargonidin derivative | 0.02 g/100 g | absent |
| Peonidin 3-(6″-malonyl)glucoside | 0.02 g/100 g | absent |
| Cyanidin derivative | 0.03 g/100 g | absent |
| Peonidin derivative | 0.01 g/100 g | absent |
| Cyanidin derivative | 0.03 g/100 g | absent |
| Cyanidin derivative | 0.02 g/100 g | absent |
| Cyanidin derivative | 0.03 g/100 g | absent |
| Total Anthocyanins | 1.02 g/100 g c | absent |
| p-Coumaric acid | 0.17 g/100 g | absent |
| Ferulic acid | 0.62 g/100 g | absent |
| Sinapic acid | 0.24 g/100 g | absent |
| Total Hydroxycinnamic Acids | 1.03 g/100 g d | absent |
| Narirutin | 3.17 g/100 g | absent |
| Hesperidin | 10.1 g/100 g | absent |
| Didymin | 1.13 g/100 g | absent |
| Total Flavanones | 14.4 g/100 g e | absent |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burò, I.; Consoli, V.; Castellano, A.; Vanella, L.; Sorrenti, V. Beneficial Effects of Standardized Extracts from Wastes of Red Oranges and Olive Leaves. Antioxidants 2022, 11, 1496. https://doi.org/10.3390/antiox11081496
Burò I, Consoli V, Castellano A, Vanella L, Sorrenti V. Beneficial Effects of Standardized Extracts from Wastes of Red Oranges and Olive Leaves. Antioxidants. 2022; 11(8):1496. https://doi.org/10.3390/antiox11081496
Chicago/Turabian StyleBurò, Ilaria, Valeria Consoli, Angela Castellano, Luca Vanella, and Valeria Sorrenti. 2022. "Beneficial Effects of Standardized Extracts from Wastes of Red Oranges and Olive Leaves" Antioxidants 11, no. 8: 1496. https://doi.org/10.3390/antiox11081496