
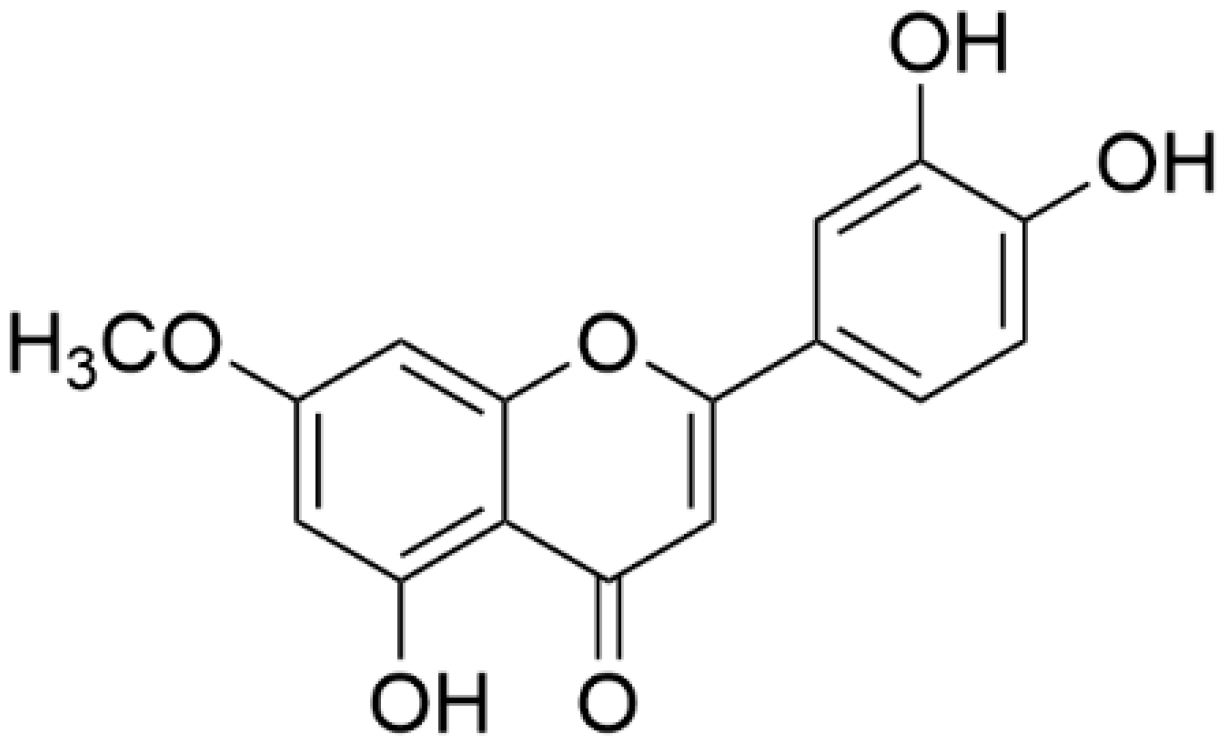
7-O-Methylluteolin Suppresses the 2,4-Dinitrochlorobenzene-Induced Nrf2/HO-1 Pathway and Atopic Dermatitis-like Lesions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction and Isolation of 7-ML from W. ganpi
2.3. Cell Culture
2.4. Dual-Luciferase Assay
2.5. Animals
2.6. DNCB-Induced Animal Experiments
2.7. Histological Examination
2.8. Measurement of Total Serum IgE and IL-4 Levels
2.9. Western Blotting
2.10. Measurement of TEWL and Skin Hydration
2.11. Statistical Analysis
3. Results
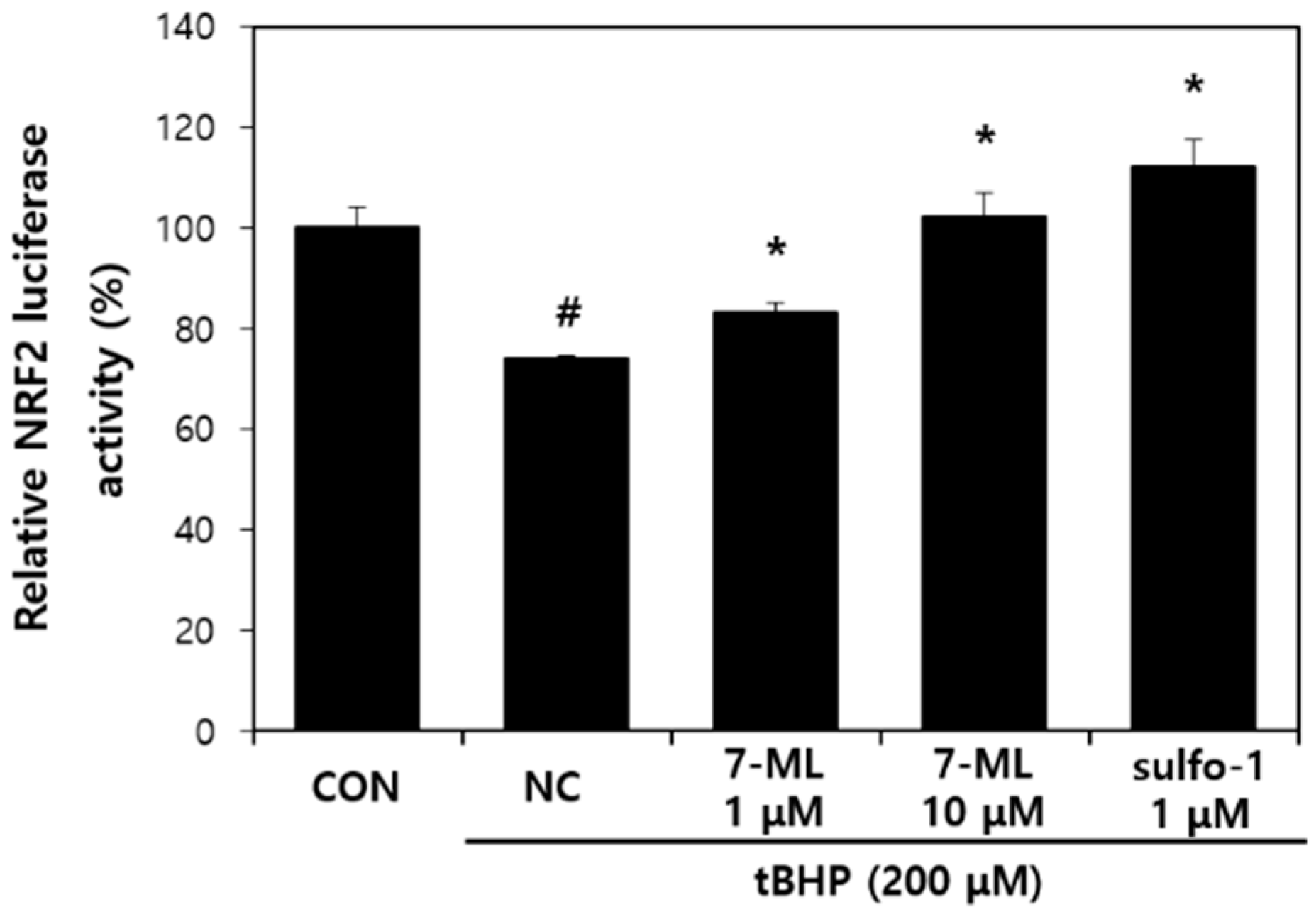
3.1. Effect of 7-ML on tBHP-Induced Nrf2 Transactivation Activity
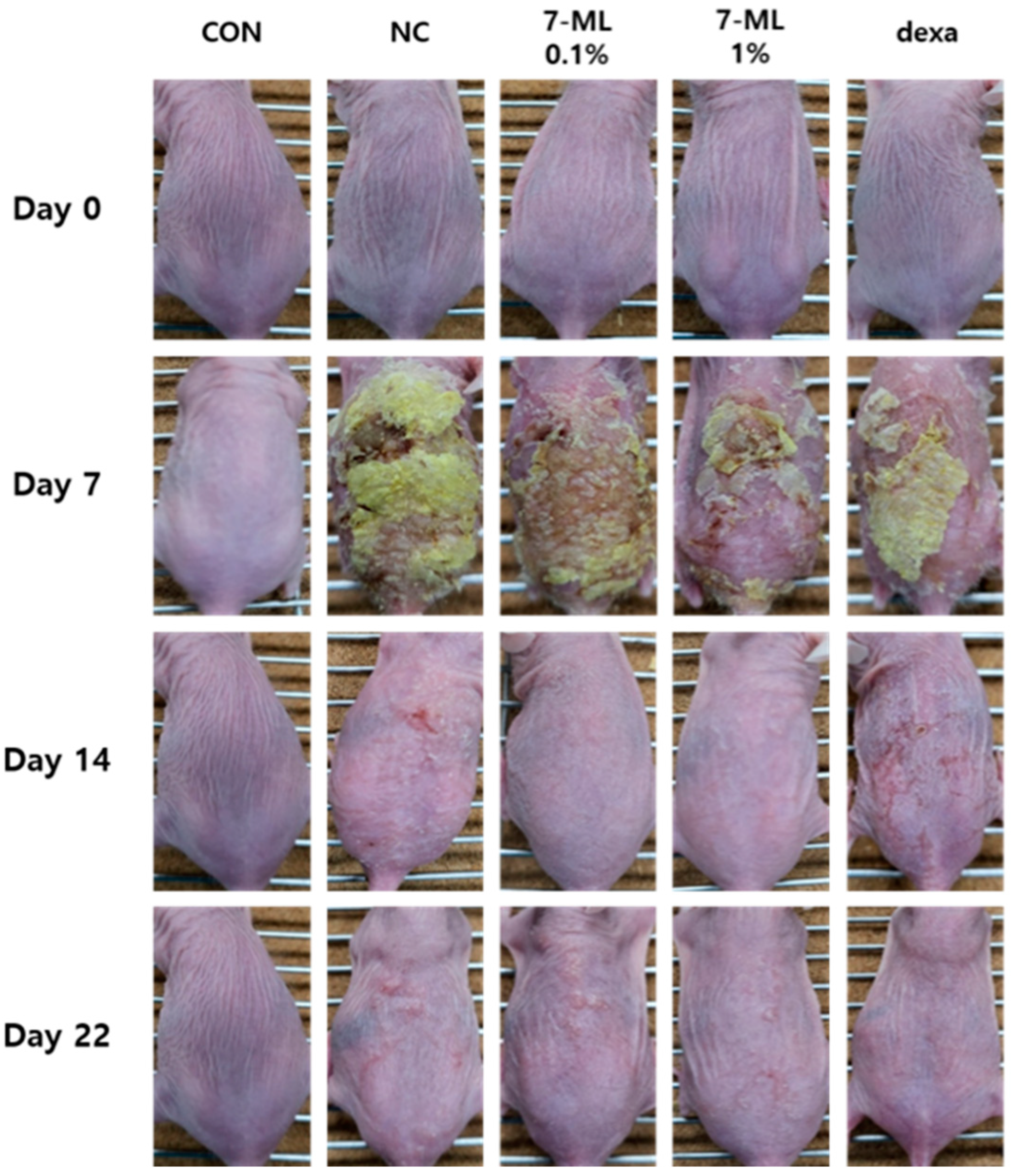
3.2. Histological Evaluation of 7-ML on DNCB-Induced AD-like Skin Lesions
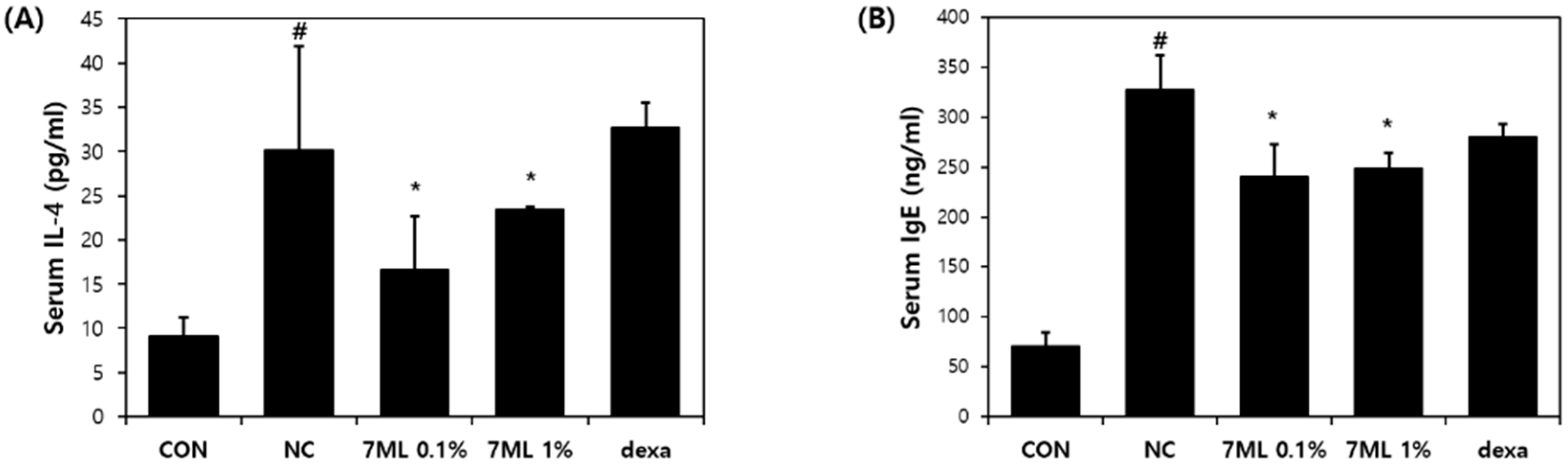
3.3. Effects of 7-ML on DNCB-Induced Inflammatory Cytokine Expression
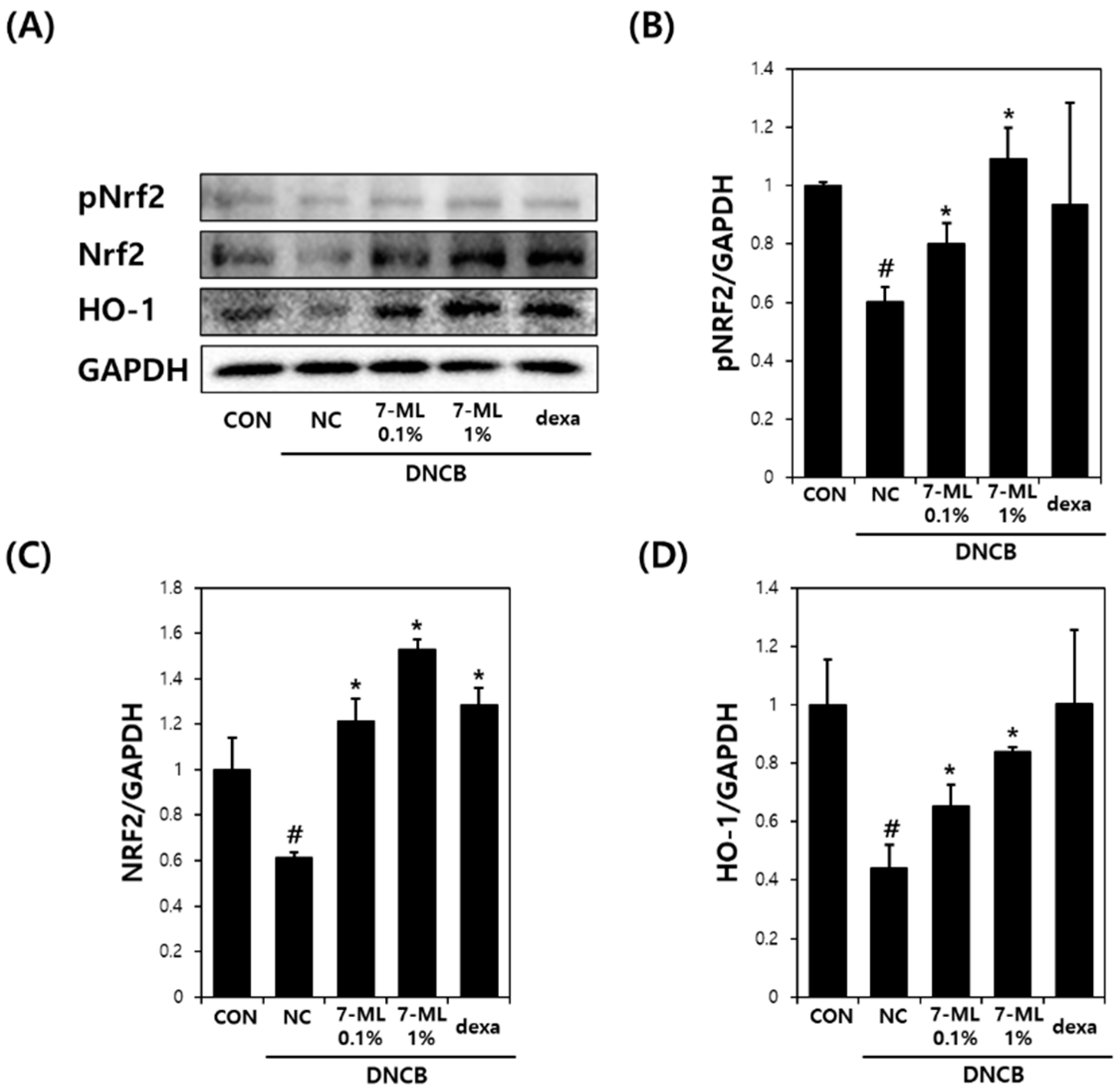
3.4. Effects of 7-ML on DNCB-Induced Antioxidant Enzyme Expression
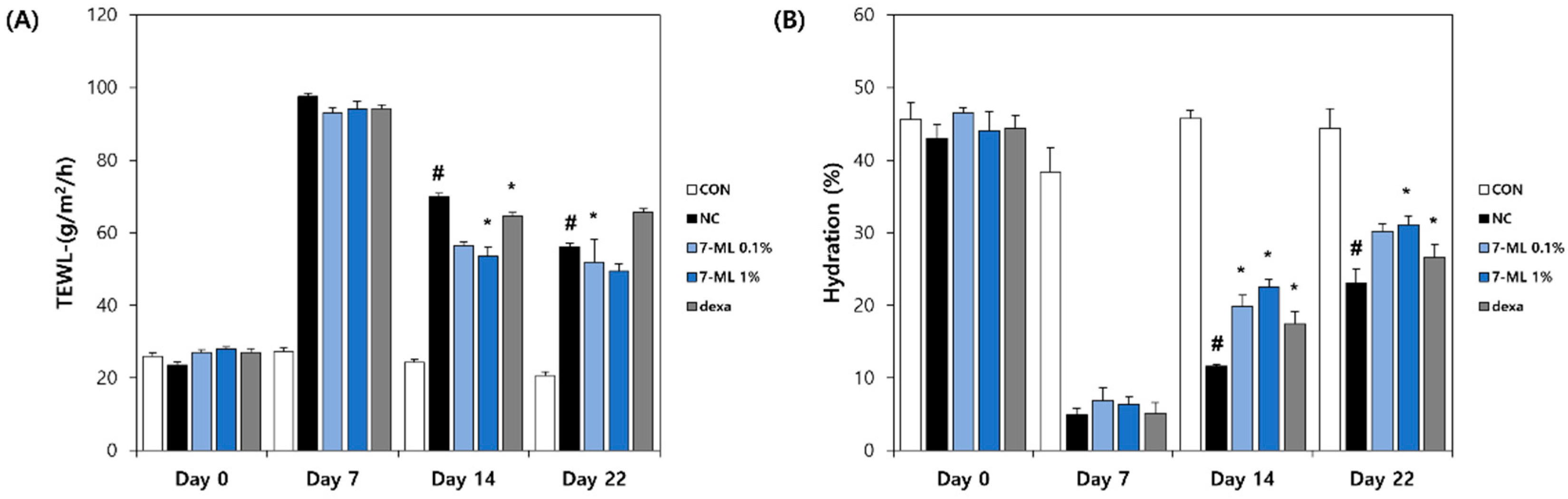
3.5. Effects of 7-ML on Skin Barrier Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simpson, E.L. Atopic dermatitis: A review of topical treatment options. Curr. Med. Res. Opin. 2010, 26, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Correale, C.E.; Walker, C.; Murphy, L.; Craig, T.J. Atopic dermatitis: A review of diagnosis and treatment. Am. Fam. Physician 1999, 60, 1191–1198. [Google Scholar] [PubMed]
- Furue, M. Regulation of Skin Barrier Function via Competition between AHR Axis versus IL-13/IL-4‒JAK‒STAT6/STAT3 Axis: Pathogenic and Therapeutic Implications in Atopic Dermatitis. J. Clin. Med. 2020, 9, 3741. [Google Scholar] [CrossRef]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Won, S.-Y.; Kim, S.R.; Maeng, S.; Jin, B.K. Interleukin-13/Interleukin-4-induced oxidative stress contributes to death of prothrombinkringle-2 (pKr-2)-activated microglia. J. Neuroimmunol. 2013, 265, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Xian, D.; Guo, M.; Xu, J.; Yang, Y.; Zhao, Y.; Zhong, J. Current evidence to support the therapeutic potential of flavonoids in oxidative stress-related dermatoses. Redox Rep. 2021, 26, 134–146. [Google Scholar] [CrossRef]
- Ji, H.; Li, X.K. Oxidative Stress in Atopic Dermatitis. Oxid. Med. Cell. Longev. 2016, 2016, 2721469. [Google Scholar] [CrossRef]
- Bertino, L.; Guarneri, F.; Cannavò, S.P.; Casciaro, M.; Pioggia, G.; Gangemi, S. Oxidative Stress and Atopic Dermatitis. Antioxidants 2020, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Kader, H.A.; Azeem, M.; Jwayed, S.A.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.A.; Hetta, H.F.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef]
- Gendrisch, F.; Esser, P.R.; Schempp, C.M.; Wölfle, U. Luteolin as a modulator of skin aging and inflammation. BioFactors 2021, 47, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Kim, M.Y.; Cho, J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Song, Y.S. Luteolin and luteolin-7-O-glucoside inhibit lipopolysaccharide-induced inflammatory responses through modulation of NF-κB/AP-1/PI3K-Akt signaling cascades in RAW 264.7 cells. Nutr. Res. Pract. 2013, 7, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.S.; Park, C.M. Luteolin and luteolin-7-O-glucoside strengthen antioxidative potential through the modulation of Nrf2/MAPK mediated HO-1 signaling cascade in RAW 264.7 cells. Food Chem. Toxicol. 2014, 65, 70–75. [Google Scholar] [CrossRef]
- Lemańska, K.; van der Woude, H.; Szymusiak, H.; Boersma, M.G.; Gliszczyńska-Swigło, A.; Rietjens, I.M.; Tyrakowska, B. The effect of catechol O-methylation on radical scavenging characteristics of quercetin and luteolin—A mechanistic insight. Free Radical Res. 2004, 38, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Patel, A.B.; Panagiotidou, S.; Theoharides, T.C. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J. Allergy Clin. Immunol. 2015, 135, 1044–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, H.; Morikawa, T.; Ando, S.; Toguchida, I.; Yoshikawa, M. Structural requirements of flavonoids for nitric oxide production inhibitory activity and mechanism of action. Bioorg. Med. Chem. 2003, 11, 1995–2000. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Palma, E.; Cordaro, M.; D’Amico, R.; Peritore, A.F.; Licata, P.; Crupi, R. Canine atopic dermatitis: Role of luteolin as new natural treatment. Vet. Med. Sci. 2020, 6, 926–932. [Google Scholar] [CrossRef]
- Hirano, T.; Higa, S.; Arimitsu, J.; Naka, T.; Shima, Y.; Ohshima, S.; Fujimoto, M.; Yamadori, T.; Kawase, I.; Tanaka, T. Flavonoids such as luteolin, fisetin and apigenin are inhibitors of interleukin-4 and interleukin-13 production by activated human basophils. Int. Arch. Allergy Immunol. 2004, 134, 135–140. [Google Scholar] [CrossRef]
- Lee, D.-H.; Park, J.-K.; Choi, J.; Jang, H.; Seol, J.-W. Anti-inflammatory effects of natural flavonoid diosmetin in IL-4 and LPS-induced macrophage activation and atopic dermatitis model. Int. Immunopharmacol. 2020, 89, 107046. [Google Scholar] [CrossRef]
- Yoon, J.-W.; Yi, M.-H.; Kim, Y.-S. Growth Environment and Vegetation Structure of Native Habitats of Wikstroemia ganpi (Sieb. et Zucc). Korean J. Environ. Ecol. 2014, 28, 331–341. [Google Scholar] [CrossRef]
- Qian, S.-J.; Zhang, Y.-H.; Li, G.-D. The complete chloroplast genome of a medicinal plant, Wikstroemia chamaedaphne (Thymelaeaceae). Mitochondrial DNA B Resour. 2020, 5, 648–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, J.-P.; Yuan, J.-J.; Pi, S.-H.; Wang, R.; Tan, R.; Ma, C.-Y.; Zhang, T.; Jiang, H.-Z. Flavones and Lignans from the Stems of Wikstroemia scytophylla Diels. Pharmacogn. Mag. 2017, 13, 488–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-Y.; Park, N.-J.; Jegal, J.; Choi, S.-h.; Lee, S.W.; Hang, J.; Kim, S.-N.; Yang, M.H. Chamaejasmine Isolated from Wikstroemia dolichantha Diels Suppresses 2,4-Dinitrofluoro-benzene-Induced Atopic Dermatitis in SKH-1 Hairless Mice. Biomolecules 2019, 9, 697. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-M.; Zhu, L.; Jiang, J.-G.; Yang, L.; Wang, D.-Y. Bioactive components and pharmacological action of Wikstroemia indica (L.) CA Mey and its clinical application. Curr. Pharm. Biotechnol. 2009, 10, 743–752. [Google Scholar] [CrossRef]
- Jegal, J.; Park, N.-J.; Jo, B.-G.; Kim, T.-Y.; Bong, S.-K.; Choi, S.-h.; Paik, J.-H.; Kim, J.-W.; Kim, S.-N.; Yang, M.H. Wikstroemia ganpi Extract Improved Atopic Dermatitis-Like Skin Lesions via Suppression of Interleukin-4 in 2,4-Dinitrochlorobenzene-Induced SKH-1 Hairless Mice. Molecules 2021, 26, 2016. [Google Scholar] [CrossRef]
- Jegal, J.; Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Jo, G.-A.; Choi, H.-S.; Kim, S.-N.; Yang, M.H. Inhibitory Effects of Luteolin 7-Methyl Ether Isolated from Wikstroemia ganpi on TNF-α/IFN-γ Mixture-Induced Inflammation in Human Keratinocyte. Nutrients 2021, 13, 4387. [Google Scholar] [CrossRef]
- Venditti, A.; Serrilli, A.M.; Cecco, M.D.; Ciaschetti, G.; Bianco, A. Coumarins and other components of Daphne oleoides Schreb. subsp. oleoides from Majella National Park. Biochem. Syst. Ecol. 2019, 83, 39–46. [Google Scholar] [CrossRef]
- Afzal, M.; Mehdi, F.S.; Abbasi, F.; Ahmad, H. Efficacy of Avicennia marina (Forsk.) Vierh. leaves extracts against some atmospheric fungi. Afr. J. Biotechnol. 2011, 10, 10790–11094. [Google Scholar] [CrossRef] [Green Version]
- Meselhy, M.R.; El-Mekkawy, S.; Ma, C.; Nakamura, N.; Tewtrakul, S.; Hattori, M. Developing of anti-HIV agents from natural resources. J. Trad. Med. 2005, 22, 116–128. [Google Scholar]
- Miura, K.; Kikuzaki, H.; Nakatani, N. Antioxidant Activity of Chemical Components from Sage (Salvia officinalis L.) and Thyme (Thymus vulgaris L.) Measured by the Oil Stability Index Method. J. Agric. Food Chem. 2002, 50, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- Saewan, N.; Koysomboon, S.; Chantrapromma, K. Anti-tyrosinase and anti-cancer activities of flavonoids from Blumea balsamifera DC. J. Med. Plant Res. 2011, 5, 1018–1025. [Google Scholar] [CrossRef]
- Lin, P.; Ren, Y.; Yan, X.; Luo, Y.; Zhang, H.; Kesarwani, M.; Bu, J.; Zhan, D.; Zhou, Y.; Tang, Y.; et al. The high NRF2 expression confers chemotherapy resistance partly through up-regulated DUSP1 in myelodysplastic syndromes. Haematologica 2019, 104, 485–496. [Google Scholar] [CrossRef]
- Shi, M.; Wang, J.; Bi, F.; Bai, Z. Diosmetin alleviates cerebral ischemia-reperfusion injury through Keap1-mediated Nrf2/ARE signaling pathway activation and NLRP3 inflammasome inhibition. Environ. Toxicol. 2022, 37, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Botchway, B.O.A.; Liu, X. Curcumin Can Activate the Nrf2/HO-1 Signaling Pathway and Scavenge Free Radicals in Spinal Cord Injury Treatment. Neurorehabil. Neural Repair 2021, 35, 576–584. [Google Scholar] [CrossRef]
- Helou, D.G.; Martin, S.F.; Pallardy, M.; Chollet-Martin, S.; Kerdine-Römer, S. Nrf2 Involvement in Chemical-Induced Skin Innate Immunity. Front. Immunol. 2019, 10, 1004. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.-Y.; Yang, M.-Q.; Wang, C.-F.; Ding, J.; Li, J.Y. Inhibiting mast cell degranulation by HO-1 affects dendritic cell maturation in vitro. Inflamm. Res. 2014, 63, 527–537. [Google Scholar] [CrossRef]
- Hwang, E.; Lin, P.; Ngo, H.T.T.; Yi, T.-H. Clove attenuates UVB-induced photodamage and repairs skin barrier function in hairless mice. Food Funct. 2018, 9, 4936–4947. [Google Scholar] [CrossRef]
- Chiricozzi, A.; Maurelli, M.; Peris, K.; Girolomoni, G. Targeting IL-4 for the Treatment of Atopic Dermatitis. Immunotargets Ther. 2020, 9, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Woodfolk, J.A. Skin Barrier Defects in Atopic Dermatitis. Curr. Allergy Asthma Rep. 2014, 14, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, B.C.; Lovato, P.; Bäumer, W.; Olivry, T. Translational Animal Models of Atopic Dermatitis for Preclinical Studies. Yale J. Biol. Med. 2017, 90, 389–402. [Google Scholar] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Paik, J.-H.; Choi, S.; Kim, S.-N.; Yang, M.H. 7-O-Methylluteolin Suppresses the 2,4-Dinitrochlorobenzene-Induced Nrf2/HO-1 Pathway and Atopic Dermatitis-like Lesions. Antioxidants 2022, 11, 1344. https://doi.org/10.3390/antiox11071344
Kim T-Y, Park N-J, Jo B-G, Paik J-H, Choi S, Kim S-N, Yang MH. 7-O-Methylluteolin Suppresses the 2,4-Dinitrochlorobenzene-Induced Nrf2/HO-1 Pathway and Atopic Dermatitis-like Lesions. Antioxidants. 2022; 11(7):1344. https://doi.org/10.3390/antiox11071344
Chicago/Turabian StyleKim, Tae-Young, No-June Park, Beom-Geun Jo, Jin-Hyub Paik, Sangho Choi, Su-Nam Kim, and Min Hye Yang. 2022. "7-O-Methylluteolin Suppresses the 2,4-Dinitrochlorobenzene-Induced Nrf2/HO-1 Pathway and Atopic Dermatitis-like Lesions" Antioxidants 11, no. 7: 1344. https://doi.org/10.3390/antiox11071344