
The Impact of Quercetin and Its Methylated Derivatives 3-o-Methylquercetin and Rhamnazin in Lipopolysaccharide-Induced Inflammation in Porcine Intestinal Cells


 , , and
, , and 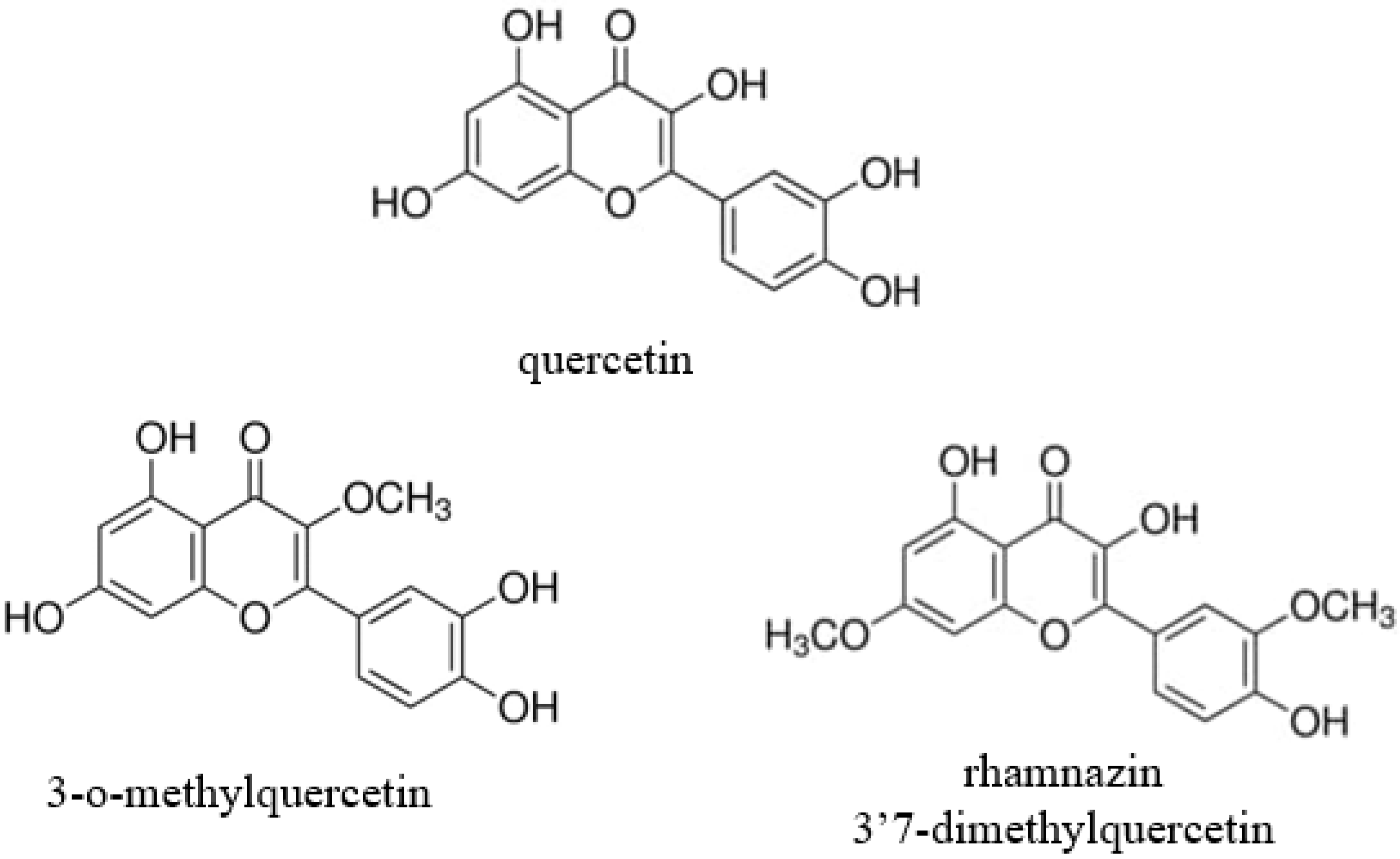{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Line and Culture Conditions
2.3. Cell Viability Measurement by the Neutral Red Uptake Assay
2.4. Treatment of Enterocytes with Q, QM, R, and LPS
2.5. Determination of the Amount of Intracellular Reactive Oxygen Species (ROS) and Extracellular H2O2 Levels in IPEC-J2 Cells
2.6. Measurement of IL-6 Levels
2.7. Paracellular Permeability Measurement
2.8. Statistics
3. Results
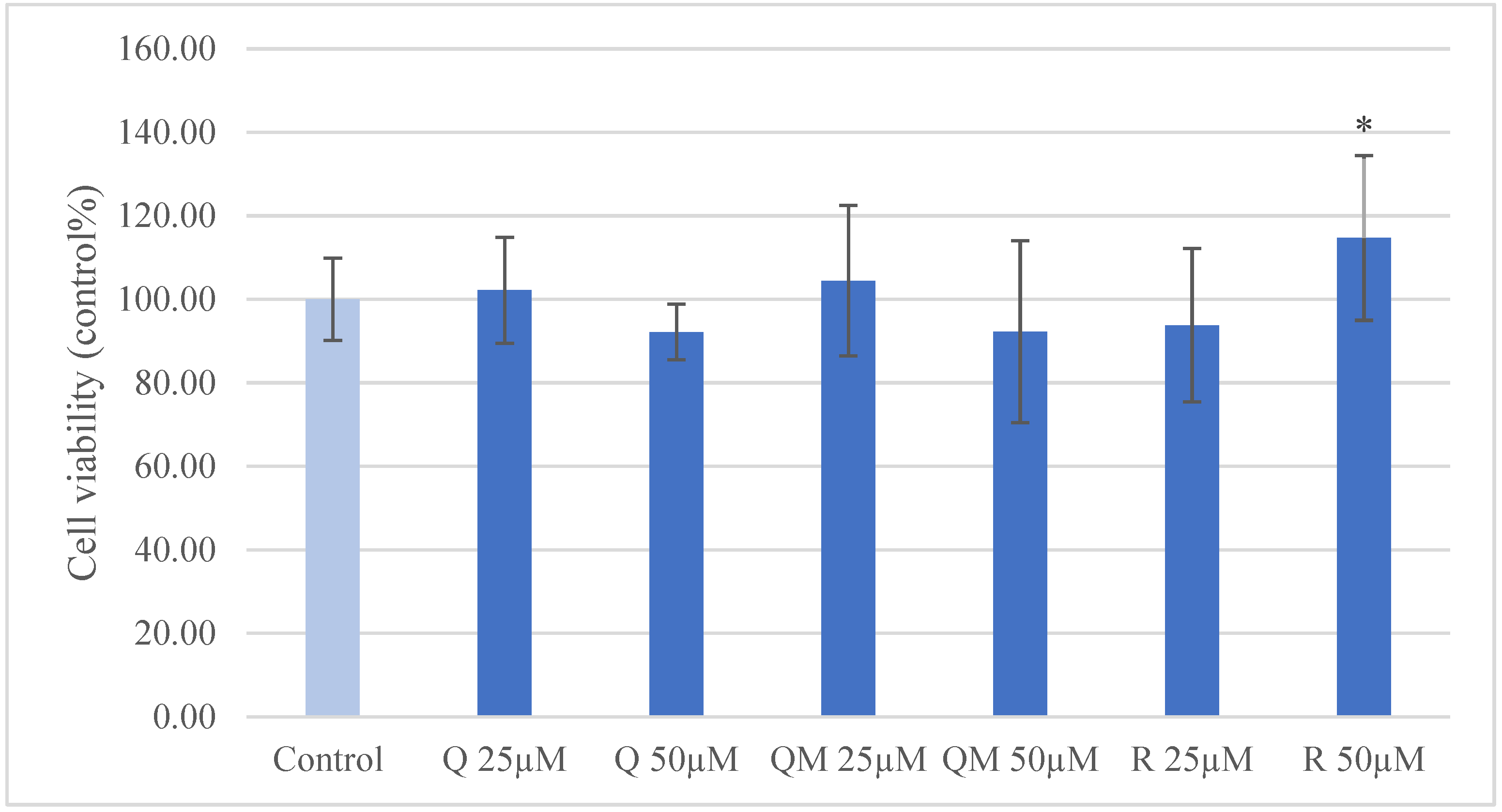
3.1. IPEC-J2 Cell Viability
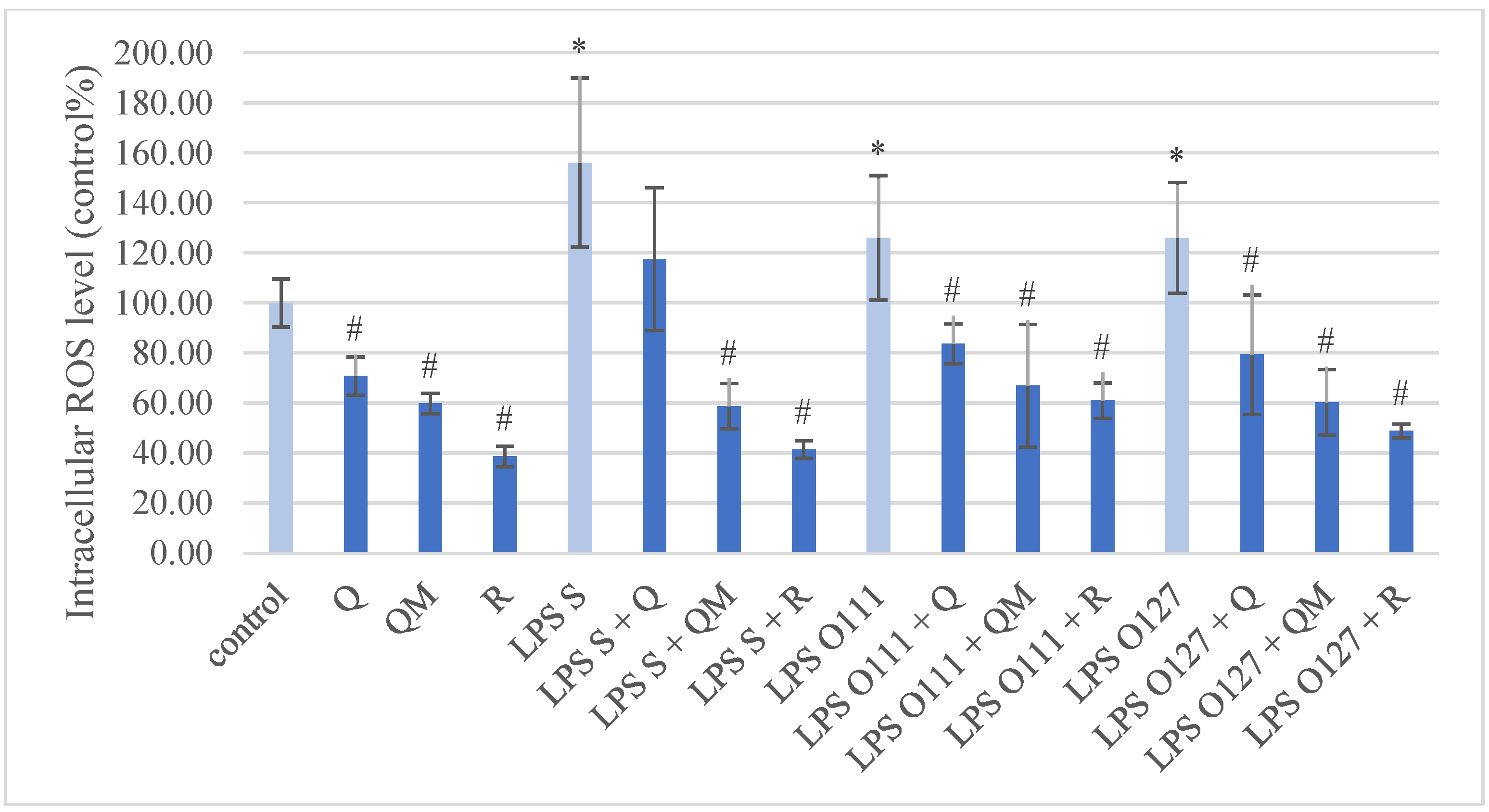
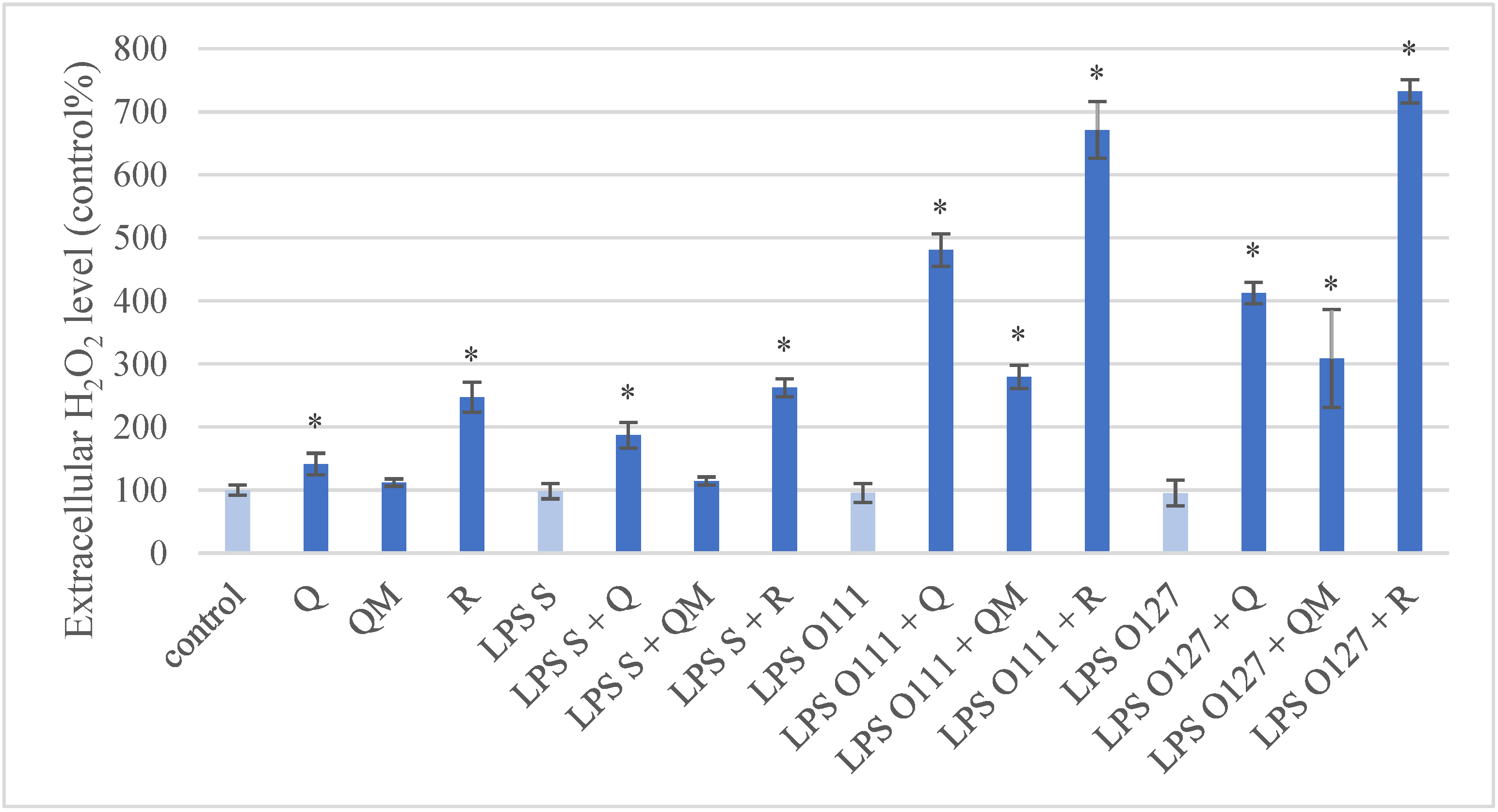
3.2. Production of ROS in IPEC-J2 Cells after Treatment of Q, QM, and R
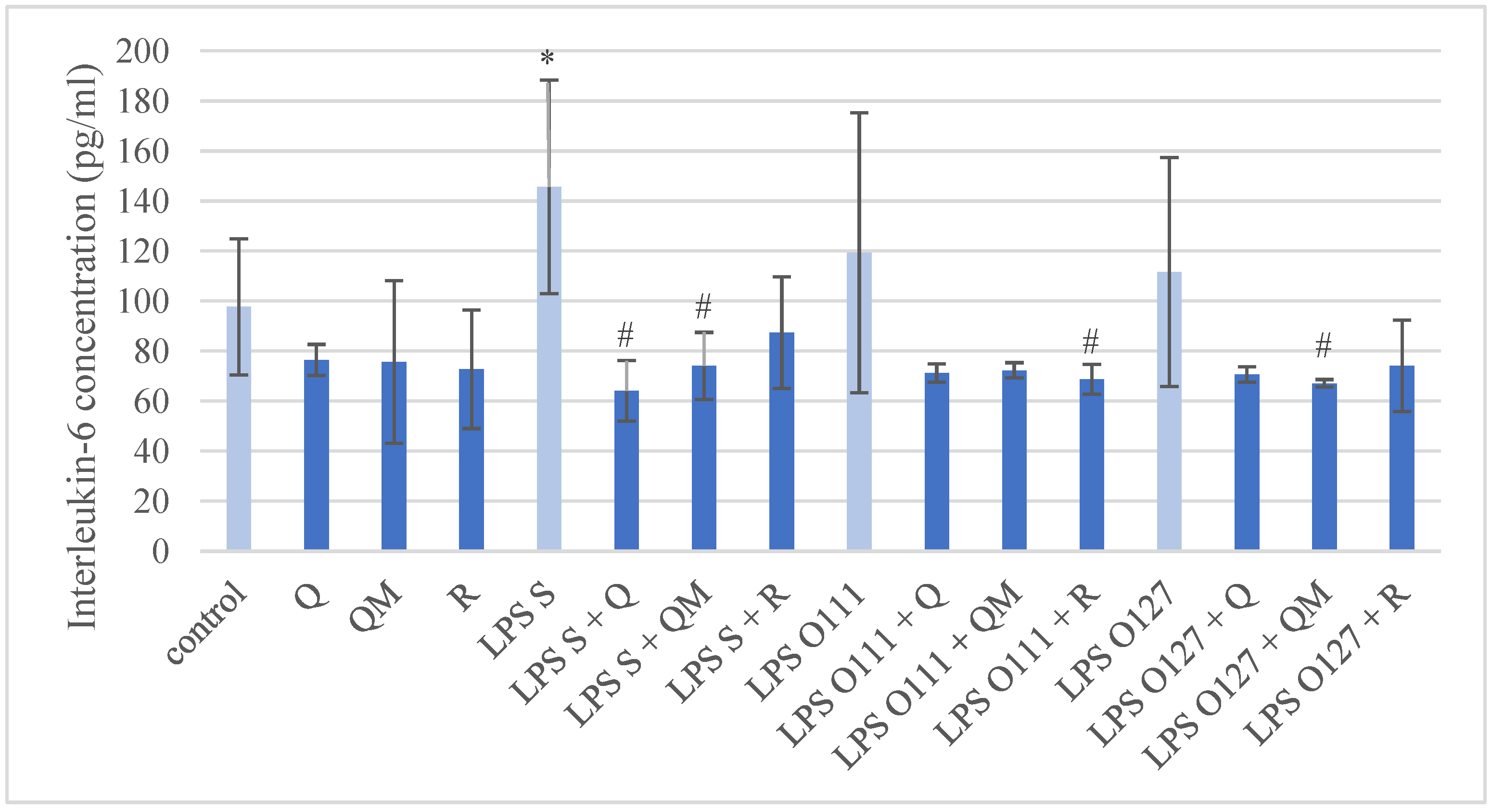
3.3. Production of IL-6 in IPEC-J2 Cells after Treatment of Q, QM and R
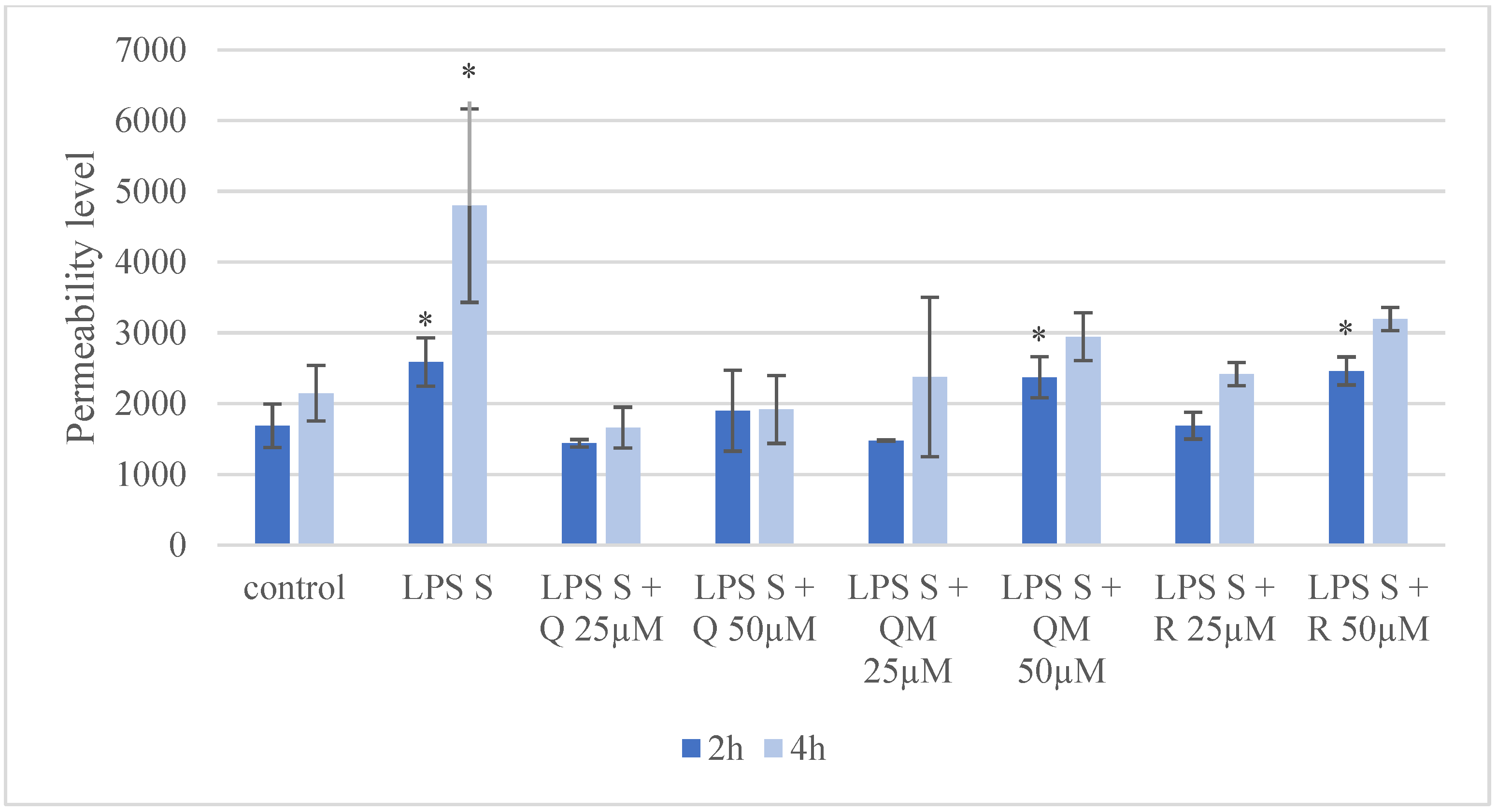
3.4. Paracellular Permeability of IPEC-J2 Cells after Treatment of Q, QM, and R
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Z.Y.; Sun, L.H.; Lin, Y.C.; Ma, X.Y.; Zheng, C.T.; Zhou, G.L.; Chen, F.; Zou, S.T. Effects of dietary glycyl-glutamine on growth performance, small intestinal integrity, and immune responses of weaning piglets challenged with lipopolysaccharide1. J. Anim. Sci. 2009, 87, 4050–4056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Chen, L.; Zheng, W.; Shi, M.; Liu, L.; Xie, C.; Wang, X.; Niu, Y.; Hou, Q.; Xu, X.; et al. Lactobacillus frumenti Facilitates Intestinal Epithelial Barrier Function Maintenance in Early-Weaned Piglets. Front. Microbiol. 2018, 9, 897. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuan, Q.; Xu, G.; Chen, H.; Lei, H.; Su, J. Effects of Quercetin on Proliferation and H2O2-Induced Apoptosis of Intestinal Porcine Enterocyte Cells. Molecules 2018, 23, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Beshbishy, A.M.; Ikram, M.; Mulla, Z.S.; El-Hack, M.E.A.; Taha, A.E.; Algammal, A.M.; Elewa, Y.H.A. The Pharmacological Activity, Biochemical Properties, and Pharmacokinetics of the Major Natural Polyphenolic Flavonoid: Quercetin. Foods 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Yu, Z.; Fan, R.; Gao, X.; Yu, M.; Li, H.; Wei, H.; Bi, K. Simultaneous Determination of Ten Flavonoids from Viscum coloratum Grown on Different Host Species and Different Sources by LC-MS. Chem. Pharm. Bull. 2011, 59, 1322–1328. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; El-Kassem, L.T.A.; Shaher, F.; Al-Farawati, R. In vitro inhibition of Hepatitis C virus protease and antioxidant by flavonoid glycosides from the Saudi costal plant Sarcocornia fruticosa. Nat. Prod. Res. 2018, 33, 3364–3371. [Google Scholar] [CrossRef]
- Goda, Y.; Hoshino, K.; Akiyama, H.; Ishikawa, T.; Abe, Y.; Nakamura, T.; Otsuka, H.; Takeda, Y.; Tanimura, A.; Toyoda, M. Constituents in Watercress: Inhibitors of Histamine Release from RBL-2H3 Cells Induced by Antigen Stimulation. Biol. Pharm. Bull. 1999, 22, 1319–1326. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, S.E.; Kaiser, S.; Pittol, V.; Doneda, E.; De Souza, K.C.B.; Bassani, V.L. Semi-preparative isolation and purification of phenolic compounds from Achyrocline satureioides (Lam) D.C. by high-performance counter-current chromatography. Phytochem. Anal. 2018, 30, 182–192. [Google Scholar] [CrossRef]
- Kumar, A.D.N.; Bevara, G.B.; Kaja, L.K.; Badana, A.K.; Malla, R.R. Protective effect of 3-O-methyl quercetin and kaempferol from Semecarpus anacardium against H2O2 induced cytotoxicity in lung and liver cells. BMC Complement. Altern. Med. 2016, 16, 376. [Google Scholar] [CrossRef] [Green Version]
- Boers, F.; Deng, B.-L.; Lemière, G.; Lepoivre, J.; De Groot, A.; Dommisse, R.; Vlietinck, A.J. An Improved Synthesis of the Anti-Picornavirus Flavone 3-O-Methylquercetin. Arch. der Pharm. 1997, 330, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.V.; Seshadri, T.R. Synthesis of rhamnazin. J. Chem. Soc. 1946, 771–772. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Yun, B.-S.; Lee, I.-K.; Kim, J.-P.; Chung, S.-H.; Shim, G.-S.; Yoo, I.-D. Lipid peroxidation inhibitory activity of some constituents isolated from the stem bark ofEucalyptus globulus. Arch. Pharmacal Res. 2000, 23, 147–150. [Google Scholar] [CrossRef]
- Martini, N.; Katerere, D.; Eloff, J. Biological activity of five antibacterial flavonoids from Combretum erythrophyllum (Combretaceae). J. Ethnopharmacol. 2004, 93, 207–212. [Google Scholar] [CrossRef]
- Wu, G.; Dai, X.; Li, X.; Jiang, H. Antioxidant and Anti-Inflammatory Effects of Rhamnazin on Lipopolysaccharide-Induced Acute Lung Injury and Inflammation in Rats. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lim, H.J.; Ryu, J.-H. In vitro anti-inflammatory activity of 3-O-methyl-flavones isolated from Siegesbeckia glabrescens. Bioorg. Med. Chem. Lett. 2008, 18, 1511–1514. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Identification of Six Flavonoids as Novel Cellular Antioxidants and Their Structure-Activity Relationship. Oxidative Med. Cell. Longev. 2020, 2020, 4150897. [Google Scholar] [CrossRef]
- Wang, J.; Lou, J.; Luo, C.; Zhou, L.; Wang, M.; Wang, L. Phenolic Compounds from Halimodendron halodendron (Pall.) Voss and Their Antimicrobial and Antioxidant Activities. Int. J. Mol. Sci. 2012, 13, 11349–11364. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.-C.; Shih, C.-M.; Chen, M.-C.; Lai, Y.-H.; Chen, J.-H.; Chen, C.-M.; Lin, C.-N. Suppressive Effects of 3-O-Methylquercetin on Ovalbumin-Induced Airway Hyperresponsiveness. Planta Med. 2004, 70, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-S.; Shih, C.-M.; Wang, S.-H.; Chen, T.-T.; Lin, C.-N.; Ko, W.-C. Mechanisms of suppression of nitric oxide production by 3-O-methylquercetin in RAW 264.7 cells. J. Ethnopharmacol. 2006, 103, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Brosnahan, A.J.; Brown, D.R. Porcine IPEC-J2 intestinal epithelial cells in microbiological investigations. Vet. Microbiol. 2012, 156, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, O.; Mátis, G.; Pászti-Gere, E.; Palócz, O.; Kulcsár, A.; Petrilla, J.; Csikó, G.; Neogrády, Z.; Gálfi, P. Effects of Lactobacillus plantarum 2142 and sodium n-butyrate in lipopolysaccharide-triggered inflammation: Comparison of a porcine intestinal epithelial cell line and primary hepatocyte monocultures with a porcine enterohepatic co-culture system12. J. Anim. Sci. 2014, 92, 3835–3845. [Google Scholar] [CrossRef] [Green Version]
- Farkas, O.; Palócz, O.; Pászti-Gere, E.; Gálfi, P. Polymethoxyflavone Apigenin-Trimethylether Suppresses LPS-Induced Inflammatory Response in Nontransformed Porcine Intestinal Cell Line IPEC-J2. Oxidative Med. Cell. Longev. 2015, 2015, 673847. [Google Scholar] [CrossRef] [Green Version]
- Palócz, O.; Pászti-Gere, E.; Gálfi, P.; Farkas, O. Chlorogenic Acid Combined with Lactobacillus plantarum 2142 Reduced LPS-Induced Intestinal Inflammation and Oxidative Stress in IPEC-J2 Cells. PLoS ONE 2016, 11, e0166642. [Google Scholar] [CrossRef]
- Karancsi, Z.; Móritz, A.V.; Lewin, N.; Veres, A.M.; Farkas, O. Beneficial Effect of a Fermented Wheat Germ Extract in Intestinal Epithelial Cells in case of Lipopolysaccharide-Evoked Inflammation. Oxidative Med. Cell. Longev. 2020, 2020, 1482482. [Google Scholar] [CrossRef]
- Schierack, P.; Nordhoff, M.; Pollmann, M.; Weyrauch, K.D.; Amasheh, S.; Lodemann, U.; Jores, J.; Tachu, B.; Kleta, S.; Blikslager, A.; et al. Characterization of a porcine intestinal epithelial cell line for in vitro studies of microbial pathogenesis in swine. Histochemistry 2005, 125, 293–305. [Google Scholar] [CrossRef]
- Vergauwen, H. The IPEC-J2 Cell Line. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; Chapter 12. [Google Scholar]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Mohanty, J.; Jaffe, J.S.; Schulman, E.S.; Raible, D.G. A highly sensitive fluorescent micro-assay of H2O2 release from activated human leukocytes using a dihydroxyphenoxazine derivative. J. Immunol. Methods 1997, 202, 133–141. [Google Scholar] [CrossRef]
- Wang, H.; Joseph, J.A. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic. Biol. Med. 1999, 27, 612–616. [Google Scholar] [CrossRef]
- Wen, X.; Walle, T. Methylated Flavonoids Have Greatly Improved Intestinal Absorption and Metabolic Stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walgren, A.R.; Walle, U.; Walle, T. Transport of Quercetin and Its Glucosides across Human Intestinal Epithelial Caco-2 Cells. Biochem. Pharmacol. 1998, 55, 1721–1727. [Google Scholar] [CrossRef]
- Walle, T. Absorption and metabolism of flavonoids. Free Radic. Biol. Med. 2004, 36, 829–837. [Google Scholar] [CrossRef]
- Maciej, J.; Schäff, C.; Kanitz, E.; Tuchscherer, A.; Bruckmaier, R.; Wolffram, S.; Hammon, H. Bioavailability of the flavonol quercetin in neonatal calves after oral administration of quercetin aglycone or rutin. J. Dairy Sci. 2015, 98, 3906–3917. [Google Scholar] [CrossRef]
- Ader, P. Bioavailability and metabolism of the flavonol quercetin in the pig. Free Radic. Biol. Med. 2000, 28, 1056–1067. [Google Scholar] [CrossRef]
- Skjolaas, K.; Burkey, T.; Dritz, S.; Minton, J. Effects of Salmonella enterica serovars Typhimurium (ST) and Choleraesuis (SC) on chemokine and cytokine expression in swine ileum and jejunal epithelial cells. Veter-Immunol. Immunopathol. 2006, 111, 199–209. [Google Scholar] [CrossRef]
- Vergauwen, H.; Prims, S.; DeGroote, J.; Wang, W.; Casteleyn, C.; Van Cruchten, S.; De Smet, S.; Michiels, J.; Van Ginneken, C. In Vitro Investigation of Six Antioxidants for Pig Diets. Antioxidants 2016, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Burkey, T.; Skjolaas, K.; Dritz, S.; Minton, J. Expression of Toll-like receptors, interleukin 8, macrophage migration inhibitory factor, and osteopontin in tissues from pigs challenged with Salmonella enterica serovar Typhimurium or serovar Choleraesuis. Veter-Immunol. Immunopathol. 2007, 115, 309–319. [Google Scholar] [CrossRef]
- Devriendt, B.; Stuyven, E.; Verdonck, F.; Goddeeris, B.; Cox, E. Enterotoxigenic Escherichia coli (K88) induce proinflammatory responses in porcine intestinal epithelial cells. Dev. Comp. Immunol. 2010, 34, 1175–1182. [Google Scholar] [CrossRef]
- Kovács, D.; Karancsi, Z.; Farkas, O.; Jerzsele, Á. Antioxidant Activity of Flavonoids in LPS-Treated IPEC-J2 Porcine Intestinal Epithelial Cells and Their Antibacterial Effect against Bacteria of Swine Origin. Antioxidants 2020, 9, 1267. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Liang, M.; Sun, X.; Mohyuddin, S.G.; Chen, S.; Wen, J.; Yong, Y.; Ma, X.; Yu, Z.; Ju, X.; et al. Baicalin Alleviates LPS-Induced Oxidative Stress via NF-κB and Nrf2–HO1 Signaling Pathways in IPEC-J2 Cells. Front. Vet. Sci. 2022, 8, 808233. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Zhang, Y.; Si, X.; Jin, Y.; Jiang, D.; Dai, Z.; Wu, Z. Quercetin Alleviates Oxidative Damage by Activating Nuclear Factor Erythroid 2-Related Factor 2 Signaling in Porcine Enterocytes. Nutrients 2021, 13, 375. [Google Scholar] [CrossRef] [PubMed]
- Kessler, M.; Ubeaud, G.; Jung, L. Anti- and pro-oxidant activity of rutin and quercetin derivatives. J. Pharm. Pharmacol. 2003, 55, 131–142. [Google Scholar] [CrossRef]
- Dueñas, M.; González-Manzano, S.; González-Paramás, A.; Santos-Buelga, C. Antioxidant evaluation of O-methylated metabolites of catechin, epicatechin and quercetin. J. Pharm. Biomed. Anal. 2010, 51, 443–449. [Google Scholar] [CrossRef]
- Cai, S.-Q.; Zhang, Q.; Zhao, X.-H.; Shi, J. The In Vitro Anti-Inflammatory Activities of Galangin and Quercetin towards the LPS-Injured Rat Intestinal Epithelial (IEC-6) Cells as Affected by Heat Treatment. Molecules 2021, 26, 7495. [Google Scholar] [CrossRef]
- Song, Y.; Han, M.; Zhang, X. Quercetin suppresses the migration and invasion in human colon cancer Caco-2 cells through regulating toll-like receptor 4/Nuclear Factor-kappa B pathway. Pharmacogn. Mag. 2016, 12, S237–S244. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Diao, P.; Shu, X.; Li, L.; Xiong, L. Quercetin and Quercitrin Attenuates the Inflammatory Response and Oxidative Stress in LPS-Induced RAW264.7 Cells: In Vitro Assessment and a Theoretical Model. BioMed Res. Int. 2019, 2019, 7039802. [Google Scholar] [CrossRef] [Green Version]
- Sul, O.-J.; Ra, S.W. Quercetin Prevents LPS-Induced Oxidative Stress and Inflammation by Modulating NOX2/ROS/NF-kB in Lung Epithelial Cells. Molecules 2021, 26, 6949. [Google Scholar] [CrossRef]
- Ren, X.; Han, L.; Li, Y.; Zhao, H.; Zhang, Z.; Zhuang, Y.; Zhong, M.; Wang, Q.; Ma, W.; Wang, Y. Isorhamnetin attenuates TNF -α-induced inflammation, proliferation, and migration in human bronchial epithelial cells via MAPK and NF-κB pathways. Anat. Rec. 2020, 304, 901–913. [Google Scholar] [CrossRef]
- Belchor, M.N.; Gaeta, H.H.; Rodrigues, C.F.B.; Costa, C.R.D.C.; Toyama, D.D.O.; Passero, L.F.D.; Laurenti, M.D.; Toyama, M.H. Evaluation of Rhamnetin as an Inhibitor of the Pharmacological Effect of Secretory Phospholipase A2. Molecules 2017, 22, 1441. [Google Scholar] [CrossRef] [Green Version]
- During, A.; Larondelle, Y. The O-methylation of chrysin markedly improves its intestinal anti-inflammatory properties: Structure–activity relationships of flavones. Biochem. Pharmacol. 2013, 86, 1739–1746. [Google Scholar] [CrossRef]
- Zhao, X.; Dong, B.; Friesen, M.; Liu, S.; Zhu, C.; Yang, C. Capsaicin Attenuates Lipopolysaccharide-Induced Inflammation and Barrier Dysfunction in Intestinal Porcine Epithelial Cell Line-J2. Front. Physiol. 2021, 12, 715469. [Google Scholar] [CrossRef]
- Jiang, Q.; Yin, J.; Chen, J.; Ma, X.; Wu, M.; Li, X.; Yao, K.; Tan, B.; Yin, Y. 4-Phenylbutyric acid accelerates rehabilitation of barrier function in IPEC-J2 cell monolayer model. Anim. Nutr. 2021, 7, 1061–1069. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karancsi, Z.; Kovács, D.; Palkovicsné Pézsa, N.; Gálfi, P.; Jerzsele, Á.; Farkas, O. The Impact of Quercetin and Its Methylated Derivatives 3-o-Methylquercetin and Rhamnazin in Lipopolysaccharide-Induced Inflammation in Porcine Intestinal Cells. Antioxidants 2022, 11, 1265. https://doi.org/10.3390/antiox11071265
Karancsi Z, Kovács D, Palkovicsné Pézsa N, Gálfi P, Jerzsele Á, Farkas O. The Impact of Quercetin and Its Methylated Derivatives 3-o-Methylquercetin and Rhamnazin in Lipopolysaccharide-Induced Inflammation in Porcine Intestinal Cells. Antioxidants. 2022; 11(7):1265. https://doi.org/10.3390/antiox11071265
Chicago/Turabian StyleKarancsi, Zita, Dóra Kovács, Nikolett Palkovicsné Pézsa, Péter Gálfi, Ákos Jerzsele, and Orsolya Farkas. 2022. "The Impact of Quercetin and Its Methylated Derivatives 3-o-Methylquercetin and Rhamnazin in Lipopolysaccharide-Induced Inflammation in Porcine Intestinal Cells" Antioxidants 11, no. 7: 1265. https://doi.org/10.3390/antiox11071265