
Mangifera indica L. Leaves as a Potential Food Source of Phenolic Compounds with Biological Activity

 , , , , ,
, , , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of the Extract
2.3. HPLC/DAD and HPLC/ESI-MS Analyses
2.4. Antioxidant Activity Measurement
2.5. In Vitro Evaluation of Antibacterial Activity of Mango Leaf Extract
2.6. Measurement of Cell Viability: MTT Assay
2.7. Alpha-Glucosidase Inhibition Assay
2.8. Anti-Inflammatory Activity In Vitro
2.9. RNA Extraction and qRT-PCR
2.10. Statistical Analysis
3. Results
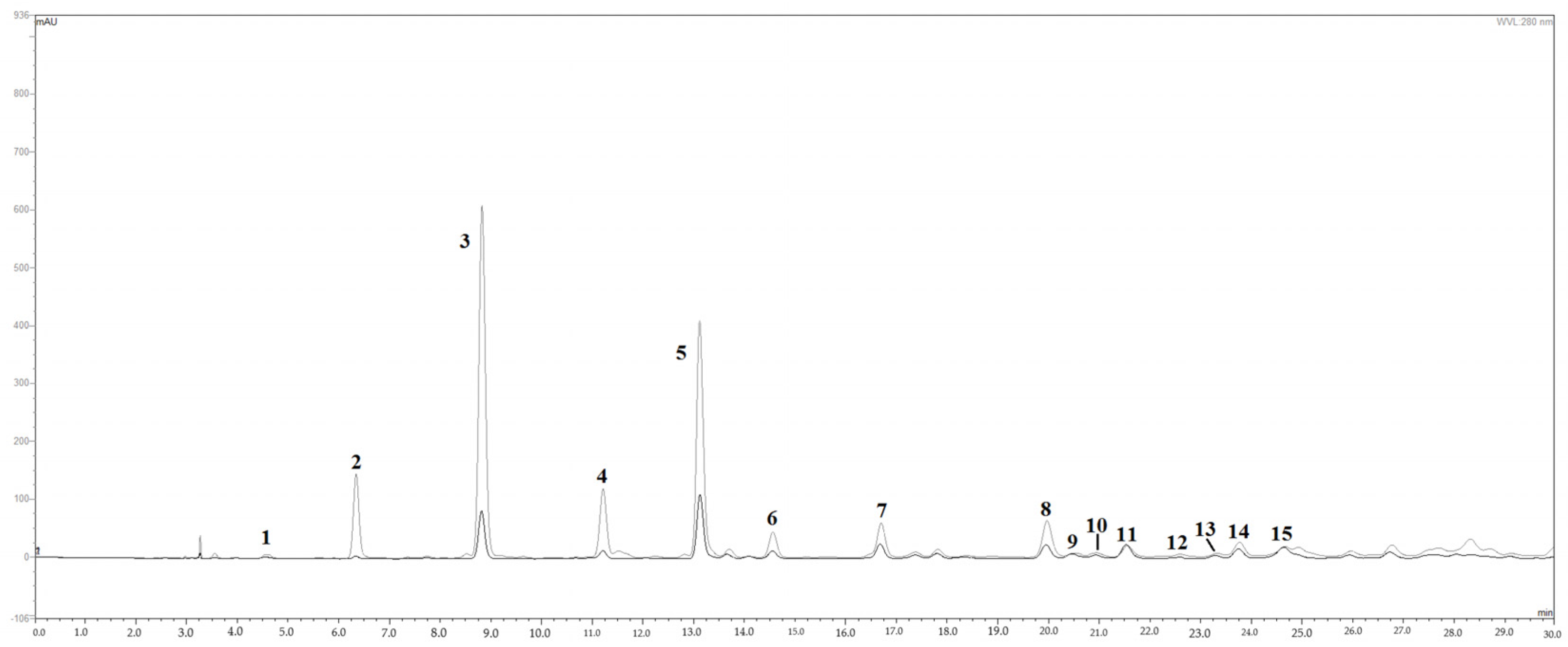
3.1. Polyphenol Profile and Content of MLE
3.2. Antioxidant Activity of Mango Leaf Extracts
3.3. Antibacterial Activity of Mango Leaf Extract
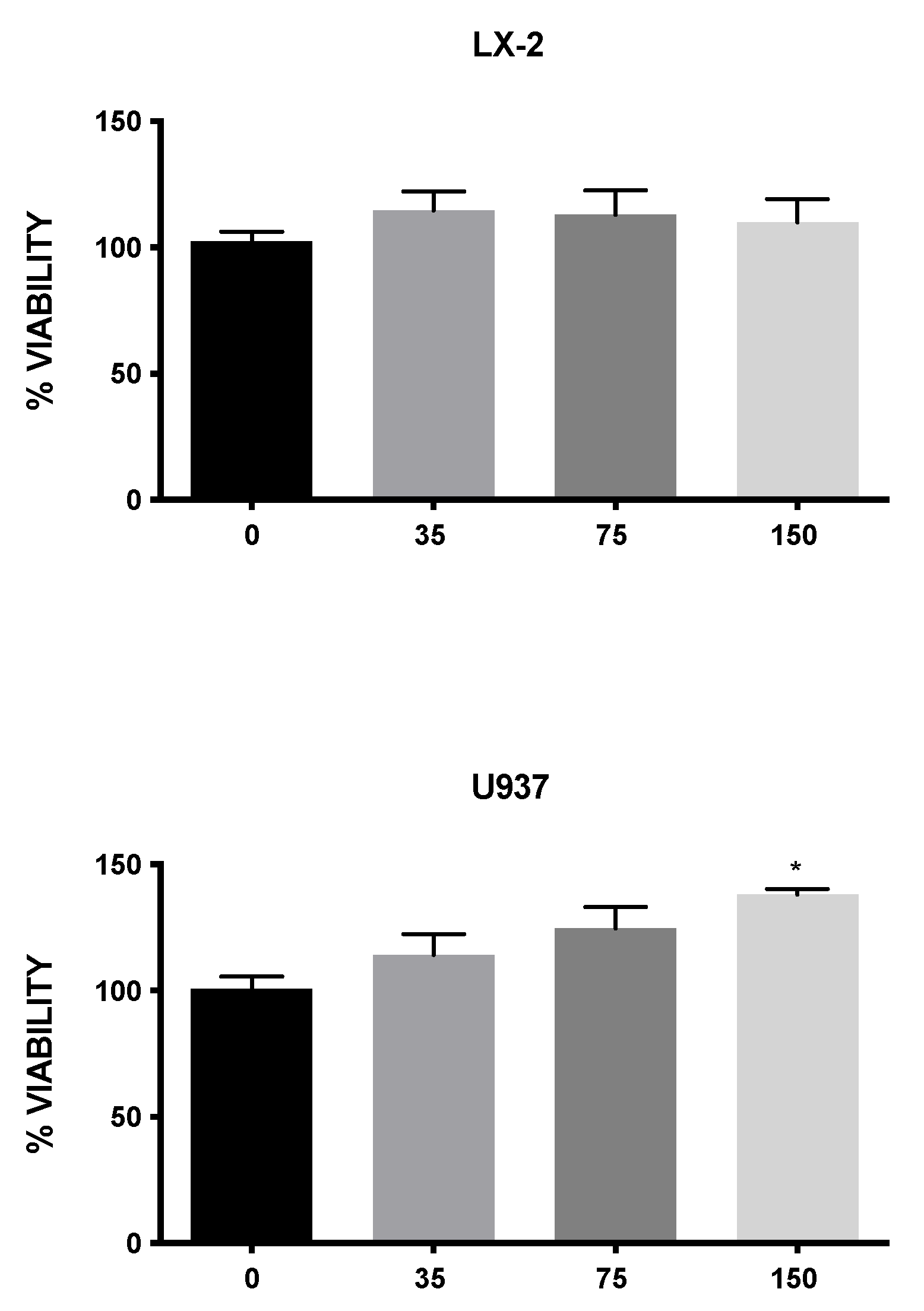
3.4. Effect of MLE on Cell Viability
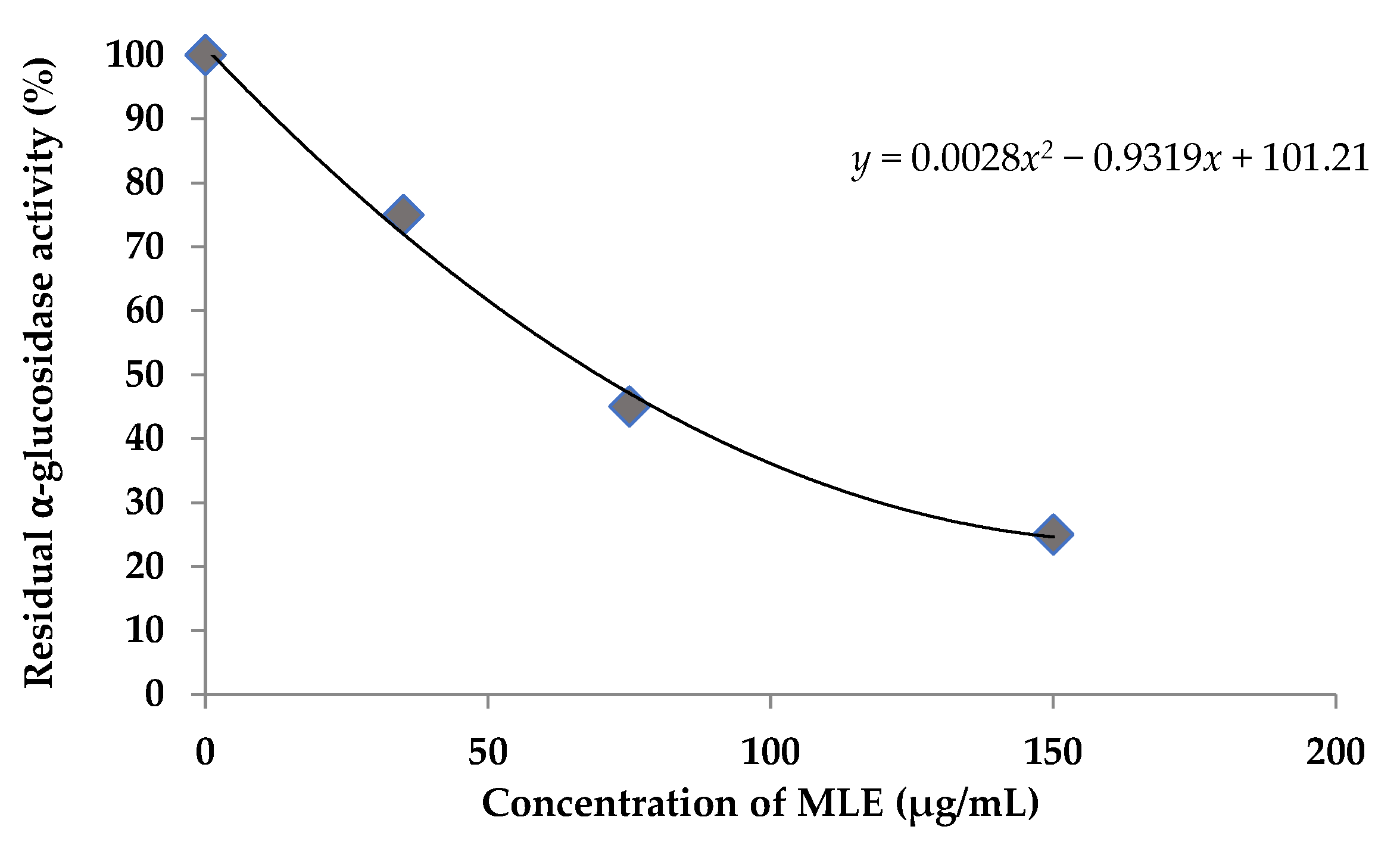
3.5. Alpha-Glucosidase Inhibition
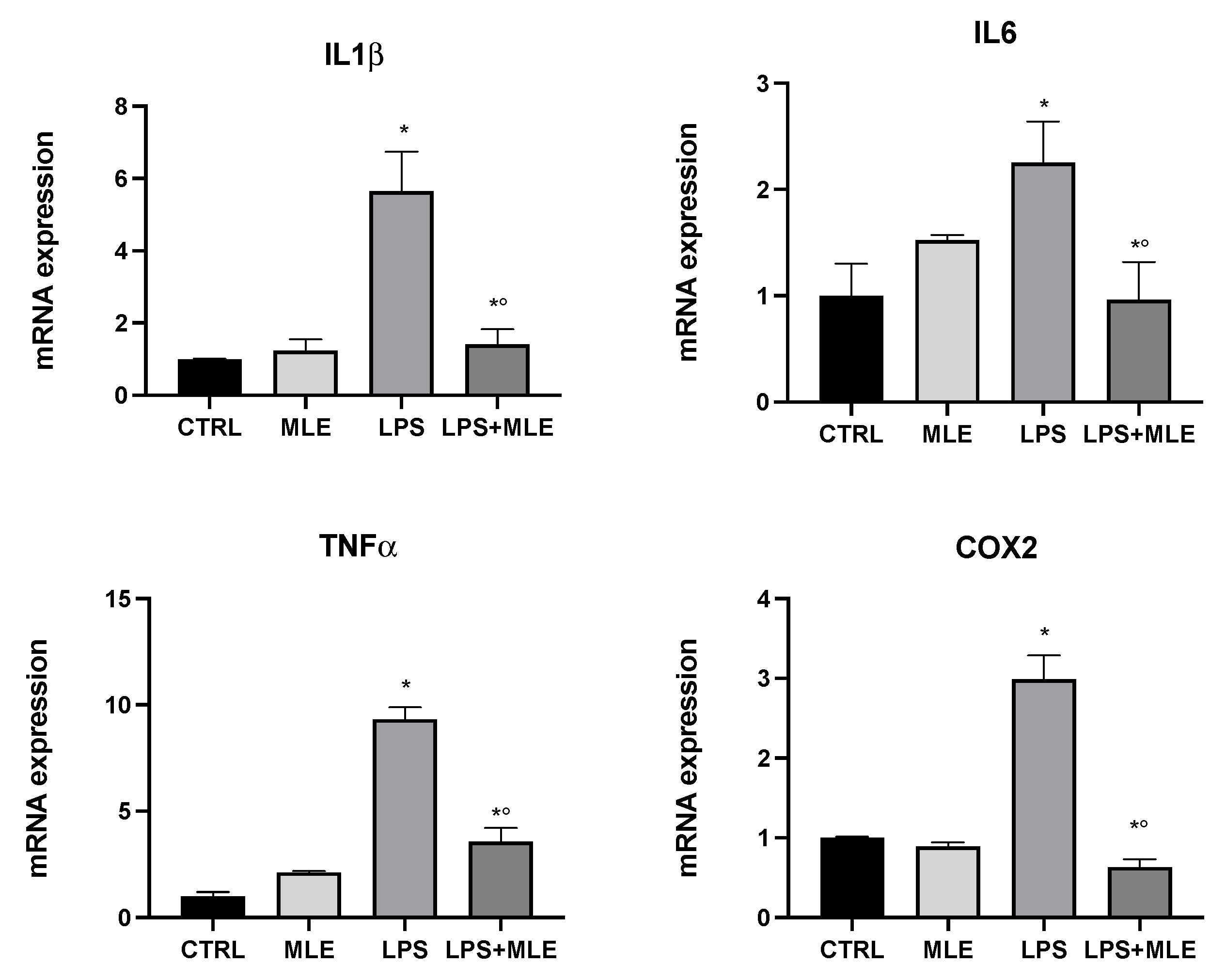
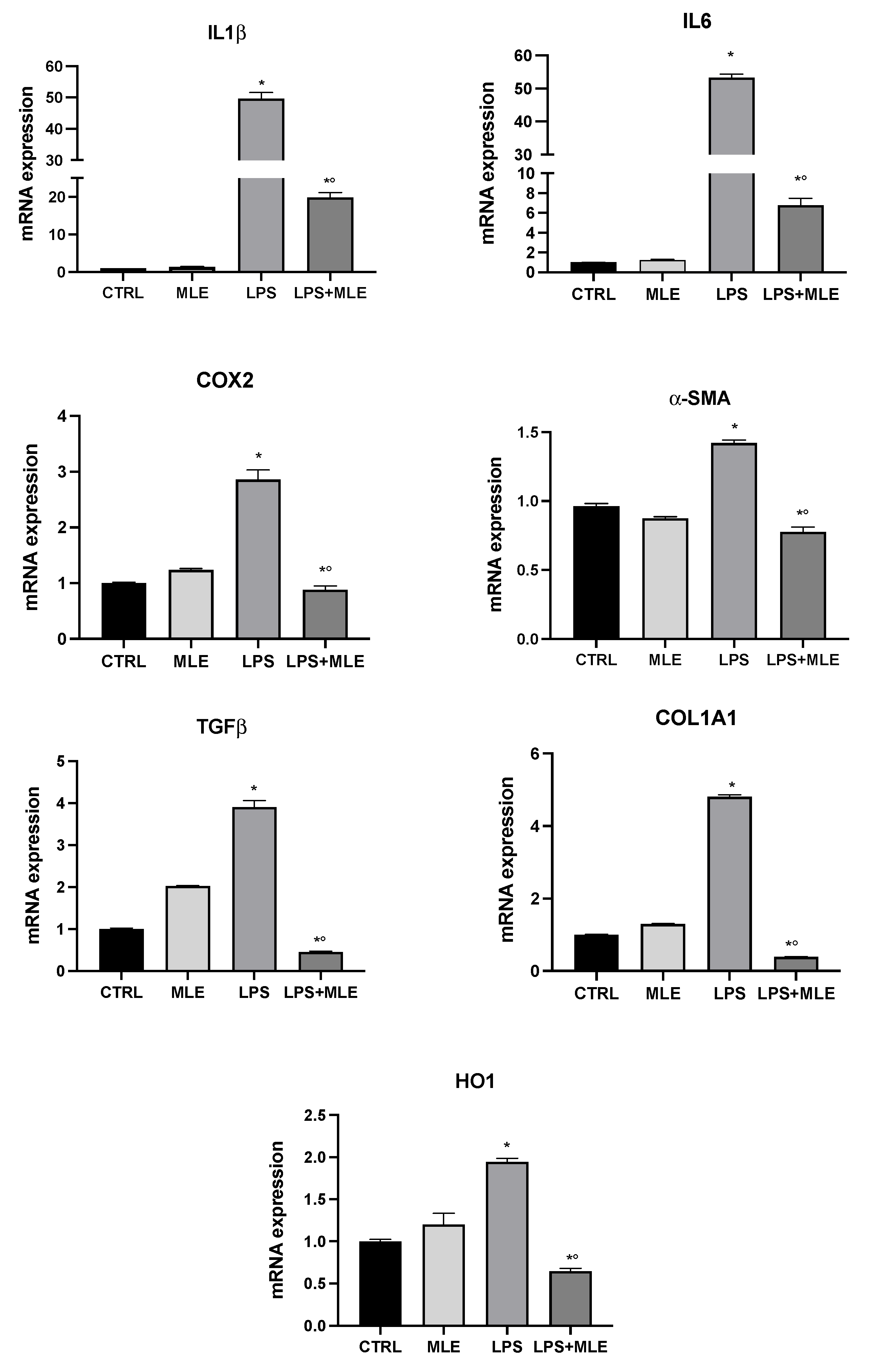
3.6. MLE Reduces In Vitro Inflammation and Hepatic Fibrosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aversa, R.P.; Rowen, V.V.; Apicella, A.; Petrescu, F.I.T. One Can Slow Down the Aging through Antioxidants. Am. J. Eng. Appl. Sci. 2016, 9, 2991. [Google Scholar] [CrossRef]
- Kumar, D.; Ladaniya, M.S.; Gurjar, M.; Kumar, S. Impact of drying methods on natural antioxidants, phenols and flavanones of immature dropped Citrus sinensis L. Osbeck fruits. Sci. Rep. 2022, 12, 6684. [Google Scholar] [CrossRef]
- Zhu, L.; Hu, W.; Murtaza, A.; Iqbal, A.; Li, J.; Zhang, J.; Li, J.; Kong, M.; Xu, X.; Pan, S. Eugenol treatment delays the flesh browning of fresh-cut water chestnut (Eleocharis tuberosa) through regulating the metabolisms of phenolics and reactive oxygen species. Food Chem. X 2022, 14, 100307. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S. Antioxidative properties of 34 green leafy vegetables. J. Funct. Foods 2016, 26, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Schieber, A.U.W.; Carle, R. Characterization of polyphenols in mango puree concentrate by HPLC with diode array and mass spectrometric detection. Innov. Food Sci. Emerg. Technol. 2000, 1, 161–166. [Google Scholar] [CrossRef]
- Noratto, G.D.; Bertoldi, M.C.; Krenek, K.; Talcott, S.T.; Stringheta, P.C.; Mertens-Talcott, S.U. Anticarcinogenic effects of polyphenolics from mango (Mangifera indica) varieties. J. Agric. Food Chem. 2010, 58, 4104–4112. [Google Scholar] [CrossRef]
- Garrido, G.; Gonzalez, D.; Lemus, Y.; Garcia, D.; Lodeiro, L.; Quintero, G.; Delporte, C.; Nunez-Selles, A.J.; Delgado, R. In vivo and in vitro anti-inflammatory activity of Mangifera indica L. extract (VIMANG). Pharmacol. Res. 2004, 50, 143–149. [Google Scholar] [CrossRef]
- Pardo-Andreu, G.L.; Paim, B.A.; Castilho, R.F.; Velho, J.A.; Delgado, R.; Vercesi, A.E.; Oliveira, H.C. Mangifera indica L. extract (Vimang) and its main polyphenol mangiferin prevent mitochondrial oxidative stress in atherosclerosis-prone hypercholesterolemic mouse. Pharmacol. Res. 2008, 57, 332–338. [Google Scholar] [CrossRef]
- Sferrazzo, G.; Palmeri, R.; Vanella, L.; Parafati, L.; Ronsisvalle, S.; Biondi, A.; Basile, F.; Li Volti, G.; Barbagallo, I. Mangifera indica L. Leaf Extract Induces Adiponectin and Regulates Adipogenesis. Int. J. Mol. Sci. 2019, 20, 3211. [Google Scholar] [CrossRef] [Green Version]
- Restuccia, C.; Lombardo, M.; Scavo, A.; Mauromicale, G.; Cirvilleri, G. Combined application of antagonistic Wickerhamomyces anomalus BS91 strain and Cynara cardunculus L. leaf extracts for the control of postharvest decay of citrus fruit. Food Microbiol. 2020, 92, 103583. [Google Scholar] [CrossRef]
- Kharchoufi, S.; Parafati, L.; Licciardello, F.; Muratore, G.; Hamdi, M.; Cirvilleri, G.; Restuccia, C. Edible coatings incorporating pomegranate peel extract and biocontrol yeast to reduce Penicillium digitatum postharvest decay of oranges. Food Microbiol. 2018, 74, 107–112. [Google Scholar] [CrossRef]
- Barbagallo, I.; Vanella, L.; Distefano, A.; Nicolosi, D.; Maravigna, A.; Lazzarino, G.; Di Rosa, M.; Tibullo, D.; Acquaviva, R.; Li Volti, G. Moringa oleifera Lam. improves lipid metabolism during adipogenic differentiation of human stem cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5223–5232. [Google Scholar]
- Odyek, G.S.; Bincher, A.L.N.; Mingeoun, D.B.K.; Poljy, W.J.; Owen, O.O. The activity of Mangifera indica L. leaf extracts against the tetanus causing bacterium, Clostridium tetani. Afr. J. Ecol. 2007, 45, 54–58. [Google Scholar]
- Alka Sahrawat, S.P.; Alka Anter, S.K.S. Antibacterial activity of Mangifera indica (mango) leaves against drug resistant bacterial strains. Int. J. Adv. Res. 2013, 6, 82. [Google Scholar]
- Olasehinde, G.I.; Smith, K.J.; Openibo, J.O.; Taiwo, O.S.; Bello, O.A.; Ajayi, J.B.; Ayepola, O.O.; Ajayi, A.A. Phytochemical and Antimicrobial Properties of Mangifera indica Leaf Extracts. Covenant J. Phys. Life Sci. 2018, 6, 1. [Google Scholar]
- Kanwal, Q.; Hussain, I.; Latif Siddiqui, H.; Javaid, A. Antifungal activity of flavonoids isolated from mango (Mangifera indica L.) leaves. Nat. Prod. Res. 2010, 24, 1907–1914. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Han, L.; Gao, X.; Liu, E.; Wang, T. Regulation of lipid and glucose homeostasis by mango tree leaf extract is mediated by AMPK and PI3K/AKT signaling pathways. Food Chem. 2013, 141, 2896–2905. [Google Scholar] [CrossRef]
- Panel, W.; Brand-Williams, M.M.E.; Cuvelier, C.; Berset, A.L.O.O. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 1, 25–30. [Google Scholar]
- Severi, J.A.; Lima, Z.P.; Kushima, H.; Brito, A.R.; Santos, L.C.; Vilegas, W.; Hiruma-Lima, C.A. Polyphenols with antiulcerogenic action from aqueous decoction of mango leaves (Mangifera indica L.). Molecules 2009, 14, 1098–1110. [Google Scholar] [CrossRef] [Green Version]
- Brenes-Arguedas, T.; Horton, M.W.; Coley, P.D.; Lokvam, J.; Waddell, R.A.; Meizoso-O’Meara, B.E.; Kursar, T.A. Contrasting mechanisms of secondary metabolite accumulation during leaf development in two tropical tree species with different leaf expansion strategies. Oecologia 2006, 149, 91–100. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Busquets-Cortes, C.; Capo, X.; Tejada, S.; Tur, J.A.; Pons, A.; Sureda, A. Cyclooxygenase-2 Inhibitors as a Therapeutic Target in Inflammatory Diseases. Curr. Med. Chem. 2019, 26, 3225–3241. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, I.; Marrazzo, G.; Frigiola, A.; Zappala, A.; Li Volti, G. Role of carbon monoxide in vascular diseases. Curr. Pharm. Biotechnol. 2012, 13, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, I.; Nicolosi, A.; Calabrese, G.; David, S.; Cimino, S.; Madonia, M.; Cappello, F. The role of the heme oxygenase system in the metabolic syndrome. Curr. Pharm. Des. 2014, 20, 4970–4974. [Google Scholar] [CrossRef]
- Riyaphan, J.; Pham, D.C.; Leong, M.K.; Weng, C.F. In Silico Approaches to Identify Polyphenol Compounds as alpha-Glucosidase and alpha-Amylase Inhibitors against Type-II Diabetes. Biomolecules 2021, 11, 1877. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Afrose, A.; Rahman, M.R.; Chowdhury, R.; Nirzhor, S.S.R.; Khan, R.I.; Kabir, M.T. A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes. Int. J. Mol. Sci. 2021, 22, 4180. [Google Scholar] [CrossRef]
- Stephan, F.; Fajac, A.; Grenet, D.; Honderlick, P.; Ricci, S.; Frachon, I.; Friard, S.; Caubarrere, I.; Bernaudin, J.F.; Stern, M. Predictive value of cytomegalovirus DNA detection by polymerase chain reaction in blood and bronchoalveolar lavage in lung transplant patients. Transplantation 1997, 63, 1430–1435. [Google Scholar] [CrossRef]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Maynard, S.; Schurman, S.H.; Harboe, C.; de Souza-Pinto, N.C.; Bohr, V.A. Base excision repair of oxidative DNA damage and association with cancer and aging. Carcinogenesis 2009, 30, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Barja, G. The mitochondrial free radical theory of aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar] [CrossRef]
- Imran, M.; Arshad, M.S.; Butt, M.S.; Kwon, J.H.; Arshad, M.U.; Sultan, M.T. Mangiferin: A natural miracle bioactive compound against lifestyle related disorders. Lipids Health Dis. 2017, 16, 84. [Google Scholar] [CrossRef]
- Selles, A.J.; Villa, D.G.; Rastrelli, L. Mango polyphenols and its protective effects on diseases associated to oxidative stress. Curr. Pharm. Biotechnol. 2015, 16, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Fomenko, E.V.; Chi, Y. Mangiferin modulation of metabolism and metabolic syndrome. Biofactors 2016, 42, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, G.F.; Carota, G.; Castruccio Castracani, C.; Spampinato, M.; Manti, S.; Papale, M.; Di Rosa, M.; Barbagallo, I.; Leonardi, S. Nutraceuticals in the Prevention of Viral Infections, including COVID-19, among the Pediatric Population: A Review of the Literature. Int. J. Mol. Sci. 2021, 22, 2465. [Google Scholar] [CrossRef] [PubMed]
- Marunaka, Y.; Marunaka, R.; Sun, H.; Yamamoto, T.; Kanamura, N.; Inui, T.; Taruno, A. Actions of Quercetin, a Polyphenol, on Blood Pressure. Molecules 2017, 22, 209. [Google Scholar] [CrossRef]
- Eid, H.M.; Haddad, P.S. The Antidiabetic Potential of Quercetin: Underlying Mechanisms. Curr. Med. Chem. 2017, 24, 355–364. [Google Scholar] [CrossRef]
- Singh, S.K.; Kumar, Y.; Kumar, S.S.; Sharma, V.K.; Dua, K.; Samad, A. Antimicrobial evaluation of mangiferin analogues. Indian J. Pharm. Sci. 2009, 71, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Scavo, A.P.G.; Restuccia, C.; Parafati, L.; Cirvilleri, G.; Mauromicale, G. Antimicrobial activity of cultivated cardoon (Cynara cardunculus L. var. altilis DC.) leaf extracts against bacterial species of agricultural and food interest. Ind. Crops Prod. 2019, 129, 206–211. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, G.; Liao, Y.; Gong, D. Inhibitory kinetics and mechanism of kaempferol on alpha-glucosidase. Food Chem. 2016, 190, 207–215. [Google Scholar] [CrossRef]
- Lawal, T.O.C.J.; Okonkwoa, E.K.; Adedoyin, H.J.; Shettima, M.S.; Muhammad, I.U.; Alhassanc, A.J. In Vitro and In Vivo Hypoglycaemic Effect of Camellia Sinensis on Alpha Glucosidase Activity and Glycaemic Index of White Bread. Appl. Food Res. 2022, 2, 37. [Google Scholar] [CrossRef]
- Kulkarni, V.R.; Vincent, K. Exploring the potential of Mangifera indica leaves extract versus mangiferin for therapeutic application. Agric. Nat. Resour. 2018, 52, 155–161. [Google Scholar] [CrossRef]
- Schmidt, A.M. Highlighting Diabetes Mellitus: The Epidemic Continues. Arterioscler. Thromb. Vasc. Biol. 2018, 38, e1–e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Liu, J.; Yang, W.; Ling, W. Lipopolysaccharide mediates hepatic stellate cell activation by regulating autophagy and retinoic acid signaling. Autophagy 2017, 13, 1813–1827. [Google Scholar] [CrossRef] [Green Version]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. LPS-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol. 2011, 31, 379–446. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, M.; Giallongo, C.; Romano, A.; Longhitano, L.; La Spina, E.; Avola, R.; Scandura, G.; Dulcamare, I.; Bramanti, V.; Di Rosa, M.; et al. Focus on Osteosclerotic Progression in Primary Myelofibrosis. Biomolecules 2021, 11, 122. [Google Scholar] [CrossRef]
- Raffaele, M.; Carota, G.; Sferrazzo, G.; Licari, M.; Barbagallo, I.; Sorrenti, V.; Signorelli, S.S.; Vanella, L. Inhibition of Heme Oxygenase Antioxidant Activity Exacerbates Hepatic Steatosis and Fibrosis In Vitro. Antioxidants 2019, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Tsukada, S.; Parsons, C.J.; Rippe, R.A. Mechanisms of liver fibrosis. Clin. Chim. Acta 2006, 364, 33–60. [Google Scholar] [CrossRef]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Blissett, D.; Blissett, R.; Henry, L.; Stepanova, M.; Younossi, Y.; Racila, A.; Hunt, S.; Beckerman, R. The economic and clinical burden of nonalcoholic fatty liver disease in the United States and Europe. Hepatology 2016, 64, 1577–1586. [Google Scholar] [CrossRef]
- Estes, C.; Razavi, H.; Loomba, R.; Younossi, Z.; Sanyal, A.J. Modeling the epidemic of nonalcoholic fatty liver disease demonstrates an exponential increase in burden of disease. Hepatology 2018, 67, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Zhu, X.; Xia, M.; Yan, H.; Chang, X.; Hu, X.; Pan, B.; Guo, W.; Li, X.; Gao, X. Impact of Type 2 Diabetes on Nonalcoholic Steatohepatitis and Advanced Fibrosis in Patients with Nonalcoholic Fatty Liver Disease. Endocr. Pract. 2020, 26, 444–453. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|
| α -SMA | NM_001141945.2 | TCGCATCAAGGCCCAAGAAA | GGATTCCCGTCTTAGTCCCG |
| COX-2 | NM_000963.4 | CTGGCGCTCAGCCATACAG | CGCACTTATACTGGTCAAATCCC |
| COL1A1 | NM_000088.4 | CTGGCCTCCCTGGAATGAAG | GGCAGCACCAGTAGCACC |
| GAPDH | NM_002046.7 | TTCTTTTGCGTCGCCAGCC | CTTCCCGTTCTCAGCCTTGAC |
| HO-1 | NM_002133.3 | GTTGGGGTGGTTTTTGAGCC | TTAGACCAAGGCCACAGTGC |
| IL-1β | NM_000576.3 | ATGATGGCTTATTACAGTGGCAA | GTCGGAGATTCGTAGCTGGA |
| IL-6 | NM_000600.5 | CCACCGGGAACGAAAGAGAA | GAGAAGGCAACTGGACCGAA |
| TGF- β | NM_000660.7 | GGAAATTGAGGGCTTTCGCC | CCGGTAGTGAACCCGTTGAT |
| TNF-α | NM_000594.4 | GCAACAAGACCACCACTTCG | GATCAAAGCTGTAGGCCCCA |
| Peak | Rt, min a | Compound Identification | Biochemical Class | mg/mL |
|---|---|---|---|---|
| 1 | 4.62 | Gallic acid b | Phenolic acid | 0.028 |
| 2 | 6.47 | Maclurin 3-C-glucoside | Benzophenones/xanthones | 0.608 |
| 3 | 8.92 | Iriflophenone 3-C-glucoside | Benzophenones/xanthones | 4.218 |
| 4 | 11.31 | Iriflophenone 3-C-(2-O-galloyl) glucoside | Benzophenones/xanthones | 1.089 |
| 5 | 13.15 | Mangiferin b | Benzophenones/xanthones | 2.023 |
| 6 | 14.66 | Iriflophenone derivative isomer 1 c | Benzophenones/xanthones | 0.368 |
| 7 | 16.77 | Iriflophenone 3-C-(2,6 di-O-galloyl) glucoside | Benzophenones/xanthones | 0.960 |
| 8 | 20.07 | Iriflophenone derivative isomer 2 c | Benzophenones/xanthones | 1.082 |
| 9 | 20.55 | Rutin b | Flavonols | 0.056 |
| 10 | 21.04 | Quercetin 3-O-glucoside | Flavonols | 0.030 |
| 11 | 21.62 | Galloyl hexose isomer 1 c | Phenolic acid | 0.212 |
| 12 | 22.59 | Quercetin pentoside isomer 1 c | Flavonols | 0.012 |
| 13 | 23.41 | Quercetin pentoside isomer 2 c | Flavonols | 0.019 |
| 14 | 23.89 | Galloyl hexose isomer 2 c | Phenolic acid | 0.269 |
| 15 | 24.82 | Galloyl hexose derivative c | Phenolic acid | 0.206 |
| Total gallic acid derivatives | 0.715 | |||
| Total benzophenones/xanthones | 10.350 | |||
| Total flavonols | 0.118 | |||
| Total polyphenols | 11.184 | |||
| Sample | RSA (%) |
|---|---|
| Leaf Extract | 90.19 ± 0.31 ns |
| Mangiferin (2.02 mg/mL) | 87.80 ± 1.86 ns |
| Antibacterial Activity | ||
|---|---|---|
| Inhibition Zone (cm) | ||
| Microorganism | MLE | Mangiferin (2.02 mg/mL) |
| B. cereus | 0.43 ± 0.06 c | 0.00 ± 0.00 ns |
| B. subtilis | 0.23 ± 0.06 d | 0.00 ± 0.00 ns |
| Ps. fluorescens | 0.70 ± 0.00 a | 0.00 ± 0.00 ns |
| St. aureus | 0.43 ± 0.06 c | 0.00 ± 0.00 ns |
| St. haemolyticus | 0.50 ± 0.00 b | 0.00 ± 0.00 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sferrazzo, G.; Palmeri, R.; Restuccia, C.; Parafati, L.; Siracusa, L.; Spampinato, M.; Carota, G.; Distefano, A.; Di Rosa, M.; Tomasello, B.; et al. Mangifera indica L. Leaves as a Potential Food Source of Phenolic Compounds with Biological Activity. Antioxidants 2022, 11, 1313. https://doi.org/10.3390/antiox11071313
Sferrazzo G, Palmeri R, Restuccia C, Parafati L, Siracusa L, Spampinato M, Carota G, Distefano A, Di Rosa M, Tomasello B, et al. Mangifera indica L. Leaves as a Potential Food Source of Phenolic Compounds with Biological Activity. Antioxidants. 2022; 11(7):1313. https://doi.org/10.3390/antiox11071313
Chicago/Turabian StyleSferrazzo, Giuseppe, Rosa Palmeri, Cristina Restuccia, Lucia Parafati, Laura Siracusa, Mariarita Spampinato, Giuseppe Carota, Alfio Distefano, Michelino Di Rosa, Barbara Tomasello, and et al. 2022. "Mangifera indica L. Leaves as a Potential Food Source of Phenolic Compounds with Biological Activity" Antioxidants 11, no. 7: 1313. https://doi.org/10.3390/antiox11071313