
Does Plant Breeding for Antioxidant-Rich Foods Have an Impact on Human Health?

 ,
,  , ,
, ,  , , ,
, , ,  , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
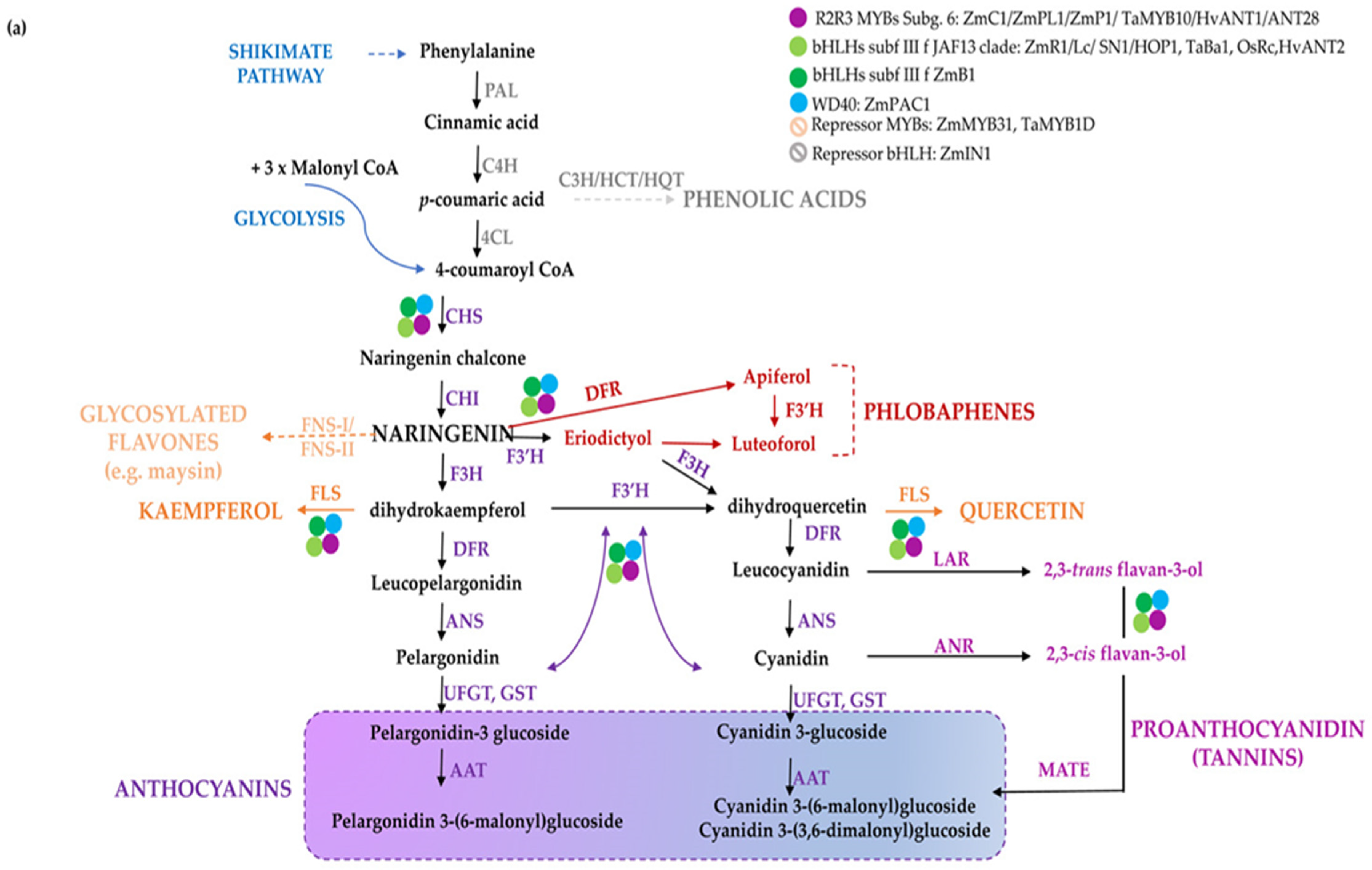
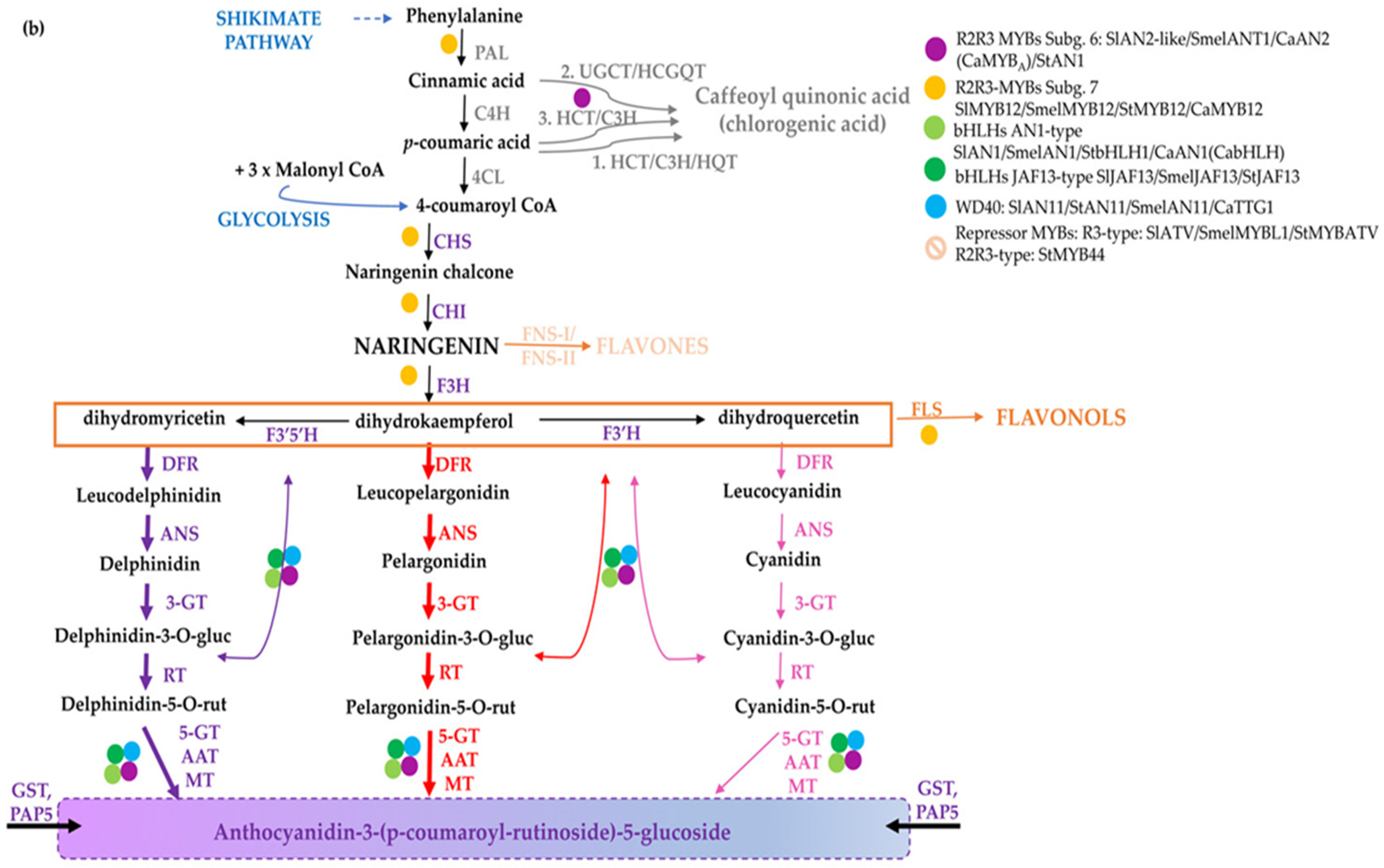
2. The Pigment Biosynthetic Pathways
2.1. The Conserved Flavonoid Pathway
2.2. Carotenoids Colouring Fruits and Grains
3. Antioxidants in Cereals
3.1. Wheat
3.2. Rice
3.3. Maize
3.4. Barley
3.5. Sorghum
4. Antioxidants in Solanaceae
4.1. Tomato

{kind=link}
{kind=link}
{kind=link}
| Typical Range of Concentration (mg/g) in the Edible Fruit of Cultivated Solanaceae Varieties | |||||||
|---|---|---|---|---|---|---|---|
| Antioxidant Class | Prevalent Compounds | Main Modifications | Distribution into the Fruit | Tomato (S. lycopersicum) | Potato (S. tuberosum) | * Eggplant (S. melongena) | Pepper (C. annum) |
| Carotenoids | lycopene and phytoene (tomato); antheraxanthin, violaxanthin, lutein; zeaxanthin (potato); luteolin (pepper) | n.d. | peel and flesh | Cultivated varieties (mg 100 g−1 FW) 7.8–18.1 (lycopene); 1.0–2.9 (phytoene) [133] | Cultivated varieties (mg/100 g DW): 1.1 (antheraxanthin); 0.8 (violaxanthin); 0.5 (lutein); 0.5 (zeaxanthin in S. tuberosum 4n); 2.2 (zeaxanthin in S. phureja 2n) [134], 0.3–3.6 (mg/100 g DW) in Andean landraces [135] | Local eggplant landraces 0.00146–0.00406 mg g−1 FW [136] | Red cultivars total carotenoids 13.51–43.32 mg/100 g DW; orange cultivars total carotenoids 109.69–190.43 mg/100 g DW; yellow cultivars total carotenoids 15.31–29.70 mg/100 g DW [137,138] |
| Phenolic acids | hydroxycinnamic acids (mainly CGA, caffeic acid) | mainly conjugated with organic acids | mainly in the flesh where they account for 70–90% of total phenolics in eggplant fruits; mainly in tuber skin (potato) | Cultivated varieties (mg 100 g−1 FW) 1.4–3.3 (CGA); 0.1–1.3 (caffeic acid) [133] | Cultivated varieties (tuber skin) 1020–2920 mg/100 g DW, CGA: 2.11 mg g−1 DW (cv. Bionica) [139,140]; total phenolics: 10.1–105.0 mg/100 g DW (Vitellotte), 6.7–108.0 mg/100 g DW (Blue Star) [141] | Cultivated eggplant CGA at stage A of fruit ripening (2319 mg/100 g DW) [142], 4 mg/100 g DW cv. Lunga Napoletana [143] | C. annuum cultivars 119.97 ± 3.44–2060.12 ± 20.56 mg GAE/100 g FW [127] |
| Flavonoids (considered as total content) | flavanones (naringenin chalcone), flavonols (mainly rutin, quercetin) | mainly conjugated (glycosides) | mainly in the fruit peel | Cultivated varieties (mg 100 g−1 FW) 0.9–18.2 (naringenin chalcone); 0.5–4.5 (rutin); 0.7–4.4 (quercetin) [133] | Cultivated varieties (tuber skin): total flavonoid content 510–960 mg/100 g DW [139], total flavonols 0–0,22 (Andean potatoes) and up to 3 (Phureja tubers) mg g−1 DW [144] | Cultivated varieties at commercial maturity (stage B) total flavonoid content 267.7 mg/100 g DW [145] | Cultivated varieties (mg 100 g−1 FW): 2.21 (quercetin); 4.71 (luteolin) (USDA) |
| Flavonoids (Anthocyanins) | mainly delphinidin, petunidin (potato), and pelargonidin-3-(p-coumaroyl-rutinoside9-5-glucoside | mainly conjugated (glycosides) | mainly in the fruit peel | n.d. | Anthocyanins: 16 (Andigenum group) and 41 (Phureja genotype) mg g−1 DW [144], 16.32 mg/100 g DW (Vitellotte), 18.12 mg/100 g DW (Magenta Love) [141] | cv. Lunga Napoletana D3R ∼1.2 mg/100 mg DW [143] | n.d. |
4.2. Potato
4.3. Eggplant
4.4. Pepper
5. Impact of Crop Management and Food Processing on Antioxidant Content
6. Impact of the Cereal and Solanaceae-Based Foods Enriched in Antioxidants on Human Health: Preclinical and Clinical Studies
6.1. Pre-Clinical Studies
6.2. Clinical Studies
| Antioxidant-Rich Food | Health Effects in Clinical Studies | Refs. | |
|---|---|---|---|
| Cereals | Antocyanin-rich rice | A reduced glycaemic response together with improvement of antioxidant status in healthy subjects | [273] |
| Purple/red rice | Significantly increases antioxidant activity and decreases plasma MDA and proinflammatory cytokines in healthy population | [274] | |
| Purple corn | Reduced plasma levels of inflammatory markers and improved the response to Infliximab, a chimeric monoclonal antibody against TNF-α, in Crohn’s Disease patients | [275] | |
| Pigmented wheat | Various medical advantages like obesity, type-2diabetes, cardiovascular disease, and cancer | [272] | |
| Black wheat | Decreased serum levels of glycated albumin and prevented the increase in TNF-α and IL-6 levels in patients with type 2 diabetes | [276] | |
| Purple wheat | Modest improvement in plasma markers of inflammation and oxidative stress in overweight and obese adults with evidence of chronic inflammation | [277] | |
| Solanaceae | Pigmented potatoes | Significant drop in blood pressure without weight gain in healthy adults | [279] |
| Reduced postprandial glycemia and insulinemia in healthy adults | [280,281] | ||
| Purple potatoes | Improved arterial stiffness in healthy adult | [282] |
7. New Perspectives to Promote Antioxidant Response in Human Cells: Plant miRNAs with Cross-Kingdom Activity
8. Does a Diet including Antioxidant-Rich Cereals and Solanaceae Have an Impact on Human Health?
Author Contributions
Funding
Conflicts of Interest
Gene and Enzyme Abbreviation List
| POD | peroxidase |
| APX | ascorbate peroxidase |
| MDA | malondialdehyde |
| OS | oxidative stress |
| ROS | reactive oxygen species |
| PAL | phenylalanine ammonia lyase |
| C4H | cinnamate 4-hydroxylase |
| C3H | p-coumarate 3′-hydroxlase |
| HQT | hydroxycinnamoyl CoA quinate hydroxycinnamoyl transferase |
| HCT | hydroxycinnamoyl CoA shikimate/quinate hydroxycinnamoyltransferase |
| UGCT | UDP glucose:cinnamate glucosyl transferase |
| HCGQT | hydroxycinnamoyl D-glucose:quinate hydroxycinnamoyl transferase |
| 4CL | 4-coumarate-CoA ligase |
| CHS | chalcone synthase |
| CHI | chalcone isomerase |
| F3H | flavanone 3-hydroxylase |
| F3′H | flavonoid 3′-hydroxylase |
| F3′5′H | flavonoid 3′,5′-hydroxylase |
| FNS | flavone synthase |
| FLS | flavonol synthase |
| DFR | dihydroflavonol 4-reductase |
| ANS | anthocyanidin synthase |
| LAR | leucoanthocyanidin reductase |
| ANR | anthocyanidin reductase |
| UFGT | UDP-glucose: flavonoid-3-O-glycosyltransferase |
| 3-GT | flavonoid 3-O-glucosyltransferase |
| RT | rhamnosyl transferase |
| 5-GT | flavonoid 5-O-glucosyl transferase |
| AAT | anthocyanin acyl transferase |
| MT | methyltransferase |
| GST | glutathione S-transferase |
| MATE | multi-antimicrobial extrusion protein |
| PAP5 | Putative Anthocyanin Permease 5 |
| DXS | 1-deoxy-D-xylulose 5-phosphate synthase |
| DXR | 1-deoxy-D-xylulose-5-phosphate reductor isomerase |
| MCT | 2-C-methyl-D-erythritol 4-phosphate cytidylyl transferase |
| CMK | 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase |
| MDS | 2-C-methyl-D-erythritol 2 4-cyclodiphosphate synthase |
| HDS | 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase |
| HDR | 4-hydroxy-3-methylbut-2-enyl diphosphate reductase |
| IPI | Isopentenyl-diphosphate delta-isomerase 1 |
| GGPP | Geranylgeranyl pyrophosphate synthase |
| PSY | Phytoene synthase |
| PDS | Phytoene desaturase |
| ZISO | 15-cis-zeta-carotene isomerase |
| ZDS | Zeta-carotene desaturase |
| CRTISO | Carotenoid isomerase |
| PTOX | Plastid terminal oxydase |
| LCYB1 | Lycopene beta-cyclase |
| CYCB | Chromoplast-specific lycopene beta-cyclase |
| LCYE | Lycopene epsilon cyclase |
| CYP97 | Cytochrome P450 |
| ZEP | Zeaxanthin epoxidase |
| VDE | Violaxanthin de-epoxidase |
| CHY | Beta-carotene hydroxylase |
| NXD1 | Neoxanthin-deficient 1 |
| ABA4 | Abscisic acid deficient 4 |
| CCD | Carotenoid cleavage dioxygenase |
| NCED | 9-cis-epoxycarotenoid dioxygenase |
| BCH2/CH2 | beta-carotene hydroxylase |
| CGA | Chlorogenic Acid |
| bHLH | basic helix loop helix |
| MYB | myeloblastosis |
| WD40 | tryptophan-aspartic acid repeat domains |
| SlAN2-like | S. lycopersicum Anthocyanin 2 like |
| SlAN1 | S. lycopersicum Anthocyanin 1 |
| SlAN11 | S. lycopersicum Anthocyanin 11 |
| SlMYB-ATV | S. lycopersicum MYB-atroviolacea |
| IN1 | Intensifier 1 |
| PAC1 | Pale Aleurone Color 1 |
| SGR1 | Stay Green 1 |
| TTG1 | Transparent Testa Glabra 1 |
| Aft | Anthocyanin fruit tomato |
| Atv | atroviolacea |
| Nrf2 | nuclear factor erythroid 2–related factor 2 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| iNOS | inducible nitric oxide synthase |
| COX-2 | cyclooxygenase-2 |
| MAPKs | mitogen-activated protein kinase |
| JNK | c-Jun N-terminal kinases |
| ERK | extracellular signal-regulated kinases |
| COX-1 | cyclooxygenase-1 |
| PGE2 | prostaglandins E2 |
| UCP3 | Uncoupling Protein 3 |
| apoE | apoliprotein E |
| SAPK/JNK | |
| IBD | inflammatory bowel disease |
| DSS | dextran sodium sulfate |
| T2DM | Type 2 diabetes |
| CVD | Cardiovascular disease |
| TNF-α | Tumor Necrosis Factor-α |
| IL-6 | Interleukin-6 |
References
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Martínez, V.; Mitjans, M.; Vinardell, M.P. Cytoprotective effects of polyphenols against oxidative damage. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Sherma, Z., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 275–288. [Google Scholar]
- Marone, D.; Mastrangelo, A.M.; Borrelli, G.M.; Mores, A.; Laidò, G.; Russo, M.A.; Ficco, D. Specialized metabolites: Physiological and biochemical role in stress resistance, strategies to improve their accumulation, and new applications in crop breeding and management. Plant Physiol. Biochem. 2022, 172, 48–55. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data (accessed on 3 November 2021).
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant. Phys. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schijlen, E.G.; Ric de Vos, C.H.; van Tunen, A.J.; Bovy, A.G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 2004, 65, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Markovic, J.M.D.; Milenkovic, D.; Amie, D.; Popovic-Bijelic, A.; Mojovic, M.; Pasti, I.A.; Markovic, Z.S. Energy requirements of the reactions of kaempferol and selected radical species in different media: Towards the prediction of the possible radical scavenging mechanisms. Struct. Chem. 2004, 25, 1795–1804. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.; Visser, R.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Albert, N.W.; Davies, K.M.; Schwinn, K.E. Gene regulation networks generate diverse pigmentation patterns in plants. Plant Signal Behav. 2014, 9, e29526. [Google Scholar] [CrossRef] [Green Version]
- Allan, A.C.; Espley, R.V. MYBs Drive Novel Consumer Traits in Fruits and Vegetables. Trends Plant Sci. 2018, 23, 693–705. [Google Scholar] [CrossRef]
- Petroni, K.; Pilu, R.; Tonelli, C. Anthocyanins in corn: A wealth of genes for human health. Planta 2014, 240, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Constabel, C.P. MYB repressors as regulators of phenylpropanoid metabolism in plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Silva, S.; Santos, A.; Chalfun-Júnior, A.; Zhao, J.; Peres, L.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants—Tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Colanero, S.; Perata, P.; Gonzali, S. What’s behind purple tomatoes? Insight into the mechanisms of anthocyanin synthesis in tomato Fruits. Plant Physiol. 2020, 182, 1841–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhang, Y. A transcriptional network makes normal tomato fruit not purple. Mol. Plant 2020, 13, 11–13. [Google Scholar] [CrossRef]
- Vershinin, A. Biological functions of carotenoids—Diversity and evolution. BioFactors 1999, 10, 99–104. [Google Scholar] [CrossRef]
- Giuliano, G.; Tavazza, R.; Diretto, G.; Beyer, P.; Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotechnol. 2008, 26, 139–145. [Google Scholar] [CrossRef]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid Biosynthesis during Tomato Fruit Development (Evidence for Tissue-Specific Gene Expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobson, G.E.; Davies, J.N. The tomato. In The Biochemistry of Fruits and Their Products; Hulme, A.C., Ed.; Academic Press: London, UK, 1971; pp. 437–482. [Google Scholar]
- Qin, X.; Zhang, W.; Dubcovsky, J.; Tian, L. Cloning and comparative analysis of carotenoid beta-hydroxylase genes provides new insights into carotenoid metabolism in tetraploid (Triticum turgidum ssp. durum) and hexaploid (Triticum aestivum) wheat grains. Plant Mol. Biol. 2012, 80, 631–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Concepción, M.; Boronat, A. Breaking new ground in the regulation of the early steps of plant isoprenoid biosynthesis. Curr. Opin. Plant Biol. 2015, 25, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Stanley, L.; Yuan, Y.W. Transcriptional regulation of carotenoid biosynthesis in plants: So many regulators, so little consensus. Front. Plant Sci. 2019, 10, 1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, R.; Bradbury, L.M.; Wurtzel, E.T. The carotenoid dioxygenase gene family in maize, sorghum, and rice. Arch. Biochem. Biophys. 2010, 504, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Barchi, L.; Pietrella, M.; Venturini, L.; Minio, A.; Toppino, L.; Acquadro, A.; Andolfo, G.; Aprea, G.; Avanzato, C.; Bassolino, L.; et al. A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci. Rep. 2019, 9, 11769. [Google Scholar] [CrossRef]
- El-Qudah, J.M. Identification and quantification of major carotenoids in some vegetables. Amer. J. Appl. Sci. 2009, 6, 492. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hort. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [Green Version]
- Martel, C.; Vrebalov, J.; Tafelmeyer, P.; Giovannoni, J.J. The tomato mads-box transcription factor ripening inhibitor interacts with promoters involved in numerous ripening processes in a colorless nonripening-dependent manner. Plant Physiol. 2011, 157, 1568–1579. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, M.; Nakano, T.; Shima, Y.; Ito, Y. A large-scale identification of direct targets of the tomato MADS box transcription factor ripening inhibitor reveals the regulation of fruit ripening. Plant Cell 2013, 25, 371–386. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A stay-green protein SlSGR1 regulates lycopene and β-carotene accumulation by interacting directly with SlPSY1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Awika, J.M. Major Cereal Grains Production and Use around the World. In Advances in Cereal Science: Implications to Food Processing and Health Promotion ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2011; Chapter 1; pp. 1–13. [Google Scholar]
- Asenstorfer, R.; Wang, Y.; Mares, D. Chemical structure of flavonoid compounds in wheat (Triticum aestivum L.) flour that contribute to the yellow colour of Asian Alkaline Noodles. J. Cereal Sci. 2006, 43, 108–119. [Google Scholar] [CrossRef]
- de Camargo, A.C.; de Souza Silva, A.P.; Soares, J.C.; de Alencar, S.M.; Handa, C.L.; Cordeiro, K.S.; Figueira, M.S.; Sampaio, G.R.; Torres, E.A.F.S.; Shahidi, F.; et al. Do flavonoids from durum wheat contribute to its bioactive properties? A prospective study. Molecules 2021, 26, 463. [Google Scholar] [CrossRef] [PubMed]
- Ficco, D.B.M.; De Simone, V.; Colecchia, S.A.; Pecorella, I.; Platani, C.; Nigro, F.; Finocchiaro, F.; Papa, R.; De Vita, P. Genetic variability in anthocyanin composition and nutritional properties of blue, purple, and red bread (Triticum aesti-vum L.) and durum (Triticum turgidum L. spp. turgidum var. durum) wheats. J. Agric. Food Chem. 2014, 62, 8686–8695. [Google Scholar] [CrossRef]
- Laddomada, B.; Caretto, S.; Mita, G. Wheat bran phenolic acids: Bioavailability and stability in whole wheat-based foods. Molecules 2015, 20, 15666–15685. [Google Scholar] [CrossRef]
- Borrelli, G.M.; De Leonardis, A.M.; Platani, C.; Troccoli, A. Distribution along durum wheat kernel of the components involved in semolina colour. J. Cereal Sci. 2008, 48, 494–502. [Google Scholar] [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic acids in wheat varieties in the healthgrain diversity Screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [Google Scholar] [CrossRef]
- Mateo Anson, N.M.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of ferulic acid is determined by its bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [Google Scholar] [CrossRef]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted Grains: A Comprehensive Review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ma, D.; Sun, D.; Wang, C.; Zhang, J.; Xie, Y.; Tiancai, G. Total phenolic, flavonoid content, and antioxidant activity of flour, noodles, and steamed bread made from different colored wheat grains by three milling methods. Crop J. 2015, 3, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Okarter, N.; Liu, C.S.; Sorrells, M.E.; Liu, R.H. Phytochemical content and antioxidant activity of six diverse varieties of whole wheat. Food Chem. 2010, 119, 249–257. [Google Scholar] [CrossRef]
- Rakszegi, M.; Boros, D.; Kuti, C.; Láng, L.; Bedo, Z.; Shewry, P.R. Composition and end-use quality of 150 wheat lines selected for the healthgrain diversity Screen. J. Agric. Food Chem. 2008, 56, 9750–9757. [Google Scholar] [CrossRef]
- Nigro, D.; Laddomada, B.; Mita, G.; Blanco, E.; Colasuonno, P.; Simeone, R.; Gadaleta, A.; Pasqualone, A.; Blanco, A. Genome wide association mapping of phenolic acids in tetraploid wheats. J. Cereal Sci. 2017, 75, 25–34. [Google Scholar] [CrossRef]
- Laddomada, B.; Durante, M.; Mangini, G.; D’Amico, L.; Lenucci, M.S.; Simeone, R.; Piarulli, L.; Mita, G.; Blanco, A. Genetic variation for phenolic acids concentration and composition in a tetraploid wheat (Triticum turgidum L.) collection. Genet. Resour. Crop Evol. 2017, 64, 587–597. [Google Scholar] [CrossRef]
- Digesù, A.M.; Platani, C.; Cattivelli, L.; Mangini, G.; Blanco, A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 2009, 50, 210–218. [Google Scholar] [CrossRef]
- Groth, S.; Wittmann, R.; Longin, C.F.; Böhm, V. Influence of variety and growing location on carotenoid and vitamin E contents of 184 different durum wheat varieties (Triticum turgidum ssp. durum) in Germany. Eur. Food Res. Technol. 2020, 246, 2079–2092. [Google Scholar] [CrossRef]
- Yilmaz, V.A.; Brandolini, A.; Hidalgo, A. Phenolic acids and antioxidant activity of wild, feral and domesticated diploid wheats. J. Cereal Sci. 2015, 64, 168–175. [Google Scholar] [CrossRef]
- Pasqualone, A.; Delvecchio, L.N.; Mangini, G.; Taranto, F.; Blanco, A. Variability of total soluble phenolic compounds and antioxidant activity in a collection of tetraploid wheat. Agric. Food Sci. 2014, 23, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Boukid, F.; Dall’Asta, M.; Bresciani, L.; Mena, P.; Del Rio, D.; Calani, L.; Sayar, R.; Seo, Y.W.; Yacoubi, I.; Mejri, M. Phenolic profile and antioxidant capacity of landraces, old and modern Tunisian durum wheat. Eur. Food Res. Technol. 2019, 245, 73–82. [Google Scholar] [CrossRef]
- Di Loreto, A.; Bosi, S.; Montero, L.; Bregola, V.; Marotti, I.; Sferrazza, R.E.; Dinelli, G.; Herrero, M.; Cifuentes, A. Determination of phenolic compounds in ancient and modern durum wheat genotypes. Electrophoresis 2018, 39, 2001–2010. [Google Scholar] [CrossRef]
- Zrckova, M.; Capouchova, I.; Eliášová, M.; Paznocht, L.; Pazderů, K.; Dvořák, P.; Konvalina, P.; Orsák, M.; Štěrba, Z. The effect of genotype, weather conditions and cropping system on antioxidant activity and content of selected antioxidant compounds in wheat with coloured grain. Plant Soil Environ. 2018, 64, 530–538. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Li, L.; Harflett, C.; Shewry, P.R.; Ward, J.L. Effects of environment and genotype on phenolic acids in wheat in the healthgrain diversity screen. J. Agric. Food Chem. 2010, 58, 9341–9352. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; De Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011, 54, 255–264. [Google Scholar] [CrossRef]
- Ziegler, J.U.; Wahl, S.; Würschum, T.; Longin, C.F.H.; Carle, R.; Schweiggert, R.M. Lutein and lutein esters in whole grain flours made from 75 genotypes of 5 triticum species grown at multiple sites. J. Agric. Food Chem. 2015, 63, 5061–5071. [Google Scholar] [CrossRef]
- Carrera, A.; Echenique, V.; Zhang, W.; Helguera, M.; Manthey, F.; Schrager, A.; Picca, A.; Cervigni, G.; Dubcovsky, J. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta colour in durum wheat, Triticum turgidum ssp. durum. J. Agric. Food Chem. 2007, 45, 67–77. [Google Scholar]
- Campos, K.M.; Royo, C.; Schulthess, A.; Villegas, D.; Matus, I.; Ammar, K.; Schwember, A.R. Association of Phytoene synthase Psy1-A1 and Psy1-B1 allelic variants with semolina yellowness in durum wheat (Triticum turgidum L. var. durum). Euphytica 2016, 207, 109–117. [Google Scholar] [CrossRef]
- Yu, S.; Li, M.; Dubcovsky, J.; Tian, L. Mutant combinations of lycopene ɛ-cyclase and β-carotene hydroxylase 2 homoeologs increased β-carotene accumulation in endosperm of tetraploid wheat (Triticum turgidum L.) grains. Plant Biotechnol. J. 2021, 20, 564. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Röder, M.S.; Börner, A. Mapping genes controlling anthocyanin pigmentation on the glume and pericarp in tetraploid wheat (Triticum durum L.). Euphytica 2010, 171, 65–69. [Google Scholar] [CrossRef]
- Top Crop Manager. Available online: https://www.topcropmanager.com/ (accessed on 15 October 2021).
- Burešová, V.; Kopecký, D.; Bartoš, J.; Martinek, P.; Watanabe, N.; Vyhnánek, T.; Doležel, J. Variation in genome composition of blue-aleurone wheat. Theor. Appl. Genet. 2015, 128, 273–282. [Google Scholar] [CrossRef]
- Jaafar, S.N.S.; Baron, J.; Siebenhandl-Ehn, S.; Rosenau, T.; Böhmdorfer, S.; Grausgruber, H. Increased anthocyanin content in purple pericarp × blue aleurone wheat crosses. Plant Breed. 2013, 132, 546–552. [Google Scholar] [CrossRef]
- Ndolo, V.U.; Beta, T. Distribution of carotenoids in endosperm, germ, and aleurone fractions of cereal grain kernels. Food Chem. 2013, 139, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Gayen, D.; Ghosh, S.; Paul, S.; Sarkar, S.N.; Datta, S.K.; Datta, K. Metabolic Regulation of Carotenoid-Enriched Golden Rice Line. Front. Plant Sci. 2016, 7, 1622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masisi, K.; Diehl-Jones, W.L.; Gordon, J.; Chapman, D.; Moghadasian, M.H.; Beta, T. Carotenoids of aleurone, germ, and endosperm fractions of barley, corn and wheat differentially inhibit oxidative stress. J. Agric. Food Chem. 2015, 63, 2715–2724. [Google Scholar] [CrossRef]
- Mendoza-Díaz, S.; Ortiz-Valerio, M.; Castaño-Tostado, E.; Figueroa-Cárdenas, J.; Reynoso-Camacho, R.; Ramos-Gómez, M.; Campos-Vega, R.; Loarca-Piña, G. Antioxidant capacity and antimutagenic activity of anthocyanin and carotenoid extracts from nixtamalized pigmented creole maize races (Zea Mays L.). Plant Foods Hum. Nutr. 2012, 67, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Pinheiro, S.S.; da Silva, L.L.; de Menezes, C.B.; de Carvalho, C.W.; Tardin, F.D.; Queiroz, V.A.; Martino, H.S.; Pinheiro-Sant’Ana, H.M. Tocochromanols and carotenoids in sorghum (Sorghum bicolor L.): Diversity and stability to the heat treatment. Food Chem. 2015, 172, 900–908. [Google Scholar] [CrossRef]
- Shammugasamy, B.; Ramakrishnan, Y.; Ghazali, H.M.; Muhammad, K. Tocopherol and tocotrienol contents of different varieties of rice in Malaysia. J. Sci. Food Agric. 2015, 95, 672–678. [Google Scholar] [CrossRef]
- Shao, Y.; Xu, F.; Sun, X.; Bao, J.; Beta, T. Phenolic acids, anthocyanins, and antioxidant capacity in rice (Oryza sativa L.) grains at four stages of development after flowering. Food Chem. 2014, 143, 90–96. [Google Scholar] [CrossRef]
- De La Parra, C.; Serna Saldivar, S.O.; Liu, R.H. Effect of processing on the phytochemical profiles and antioxidant activity of corn for production of masa, tortillas, and tortilla chips. J. Agric. Food Chem. 2007, 55, 4177–4183. [Google Scholar] [CrossRef]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Stoichiometric and kinetic studies of phenolic antioxidants from andean purple corn and red-fleshed sweetpotato. J. Agric. Food Chem. 2003, 51, 3313–3319. [Google Scholar] [CrossRef]
- Salazar-López, N.J.; González-Aguilar, G.A.; Rouzaud-Sández, O.; Robles-Sánchez, M. Bioaccessibility of hydroxycinnamic acids and antioxidant capacity from sorghum bran thermally processed during simulated in vitro gastrointestinal digestion. J. Food Sci. Technol. 2018, 55, 2021–2030. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Pilu, R.; Cassani, E.; Sirizzotti, A.; Petroni, K.; Tonelli, C. Effect of Flavonoid Pigments on the Accumulation of Fumonisin B1 in the Maize Kernel. J. Appl. Genet. 2011, 52, 145–152. [Google Scholar] [CrossRef]
- Habyarimana, E.; Lopez-Cruz, M. Genomic Selection for Antioxidant Production in a Panel of Sorghum bicolor and S. bicolor × S. halepense Lines. Genes 2019, 10, 841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, F.; Giusti, M.M. Quantification of purple corn (Zea mays L.) anthocyanins using spectrophotometric and hplc approaches: Method comparison and correlation. Food Anal. Methods 2016, 9, 1367–1380. [Google Scholar] [CrossRef]
- Tomay, F.; Marinelli, A.; Leoni, V.; Caccia, C.; Matros, A.; Mock, H.P.; Tonelli, C.; Petroni, K. Purple corn extract induces long-lasting reprogramming and M2 phenotypic switch of adipose tissue macrophages in obese mice. J. Transl. Med. 2019, 17, 237. [Google Scholar] [CrossRef] [Green Version]
- Finocchiaro, F.; Ferrari, B.; Gianinetti, A.; Dall’asta, C.; Galaverna, G.; Scazzina, F.; Pellegrini, N. Characterization of antioxidant compounds of red and white rice and changes in total antioxidant capacity during processing. Mol. Nut. Food Res. 2007, 51, 1006–1019. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; McClung, A.M.; Chen, M.H. Phytochemicals and antioxidant capacities in rice brans of different color. J. Food Sci. 2011, 76, C117–C126. [Google Scholar] [CrossRef]
- Sweeney, M.T.; Thomson, M.J.; Pfeil, B.E.; McCouch, S. Caught red-handed: Rc encodes a basic helix-loop-helix protein conditioning red pericarp in rice. Plant Cell 2006, 18, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Zhang, G.; Bao, J. Total phenolic content and antioxidant capacity of rice grains with extremely small size. Afr. J. Agric. Res. 2011, 6, 2289–2293. [Google Scholar]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529–532. [Google Scholar] [CrossRef]
- Oikawa, T.; Maeda, H.; Oguchi, T.; Yamaguchi, T.; Tanabe, N.; Ebana, K.; Yano, M.; Ebitani, T.; Izawa, T. The Birth of a Black Rice Gene and Its Local Spread by Introgression. Plant Cell 2015, 27, 2401–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [Green Version]
- Schaub, P.; Al-Babili, S.; Drake, R.; Beyer, P. Why is golden rice golden (yellow) instead of red? Plant Physiol. 2005, 138, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paine, J.A.; Shipton, C.A.; Chaggar, S.; Howells, R.M.; Kennedy, M.J.; Vernon, G.; Wright, S.Y.; Hinchliffe, E.; Adams, J.L.; Silverstone, A.L.; et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nat. Biotechnol. 2005, 23, 482–487. [Google Scholar] [CrossRef]
- Tang, G. Techniques for measuring vitamin A activity from β-carotene. Am. J. Clin. Nutr. 2012, 96, 1185S–1188S. [Google Scholar] [CrossRef] [Green Version]
- Colombo, F.; Di Lorenzo, C.; Petroni, K.; Silano, M.; Pilu, R.; Falletta, E.; Biella, S.; Restani, P. Pigmented corn varieties as functional ingredients for gluten-free products. Foods 2021, 10, 1770. [Google Scholar] [CrossRef] [PubMed]
- Suriano, S.; Balconi, C.; Valoti, P.; Redaelli, R. Comparison of total polyphenols, profile anthocyanins, color analysis, carotenoids and tocols in pigmented maize. LWT 2021, 144, 111257. [Google Scholar] [CrossRef]
- Kristina, K.; Grbeša, D. Carotenoid Content and Antioxidant Activity of Hexane Extracts from Selected Croatian Corn Hybrids. Food Chem. 2015, 167, 402–408. [Google Scholar]
- Hulshof, P.J.; Kosmeijer-Schuil, T.; West, C.E.; Hollman, P.C. Quick screening of maize kernels for provitamin a content. J. Food Compos. Anal. 2007, 20, 655–661. [Google Scholar] [CrossRef]
- Himi, E.; Yamashita, Y.; Haruyama, N.; Yanagisawa, T.; Maekawa, M.; Taketa, S. Ant28 gene for proanthocyanidin synthesis encoding the R2R3 MYB domain protein (Hvmyb10) highly affects grain dormancy in barley. Euphytica 2011, 188, 141–151. [Google Scholar] [CrossRef]
- Lago, C.; Cassani, E.; Zanzi, C.; Landoni, M.; Trovato, R.; Pilu, R. Development and study of a maize cultivar rich in anthocyanins: Coloured polenta, a new functional food. Plant Breed. 2014, 133, 210–217. [Google Scholar] [CrossRef]
- Lago, C.; Landoni, M.; Cassani, E.; Doria, E.; Nielsen, E.; Pilu, R. Study and characterization of a novel functional food: Purple popcorn. Mol. Breed. 2013, 31, 575–585. [Google Scholar] [CrossRef]
- Baik, B.-K.; Ullrich, S.-E. Barley for food: Characteristics, improvement, and renewed interest. J. Cereal Sci. 2008, 48, 233–242. [Google Scholar] [CrossRef]
- Idehen, E.; Tang, Y.; Sang, S. Bioactive phytochemicals in barley. J. Food Drug Anal. 2017, 25, 148–161. [Google Scholar] [CrossRef] [Green Version]
- Strygina, K.V.; Börner, A.; Khlestkina, E.K. Identification and characterization of regulatory network components for anthocyanin synthesis in barley aleurone. BMC Plant Biol. 2017, 17 (Suppl. 1), 184. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Jiang, Q.T.; Wei, Y.M.; Liu, C. Inheritance analysis and mapping of quantitative trait loci (QTL) controlling individual anthocyanin compounds in purple barley (Hordeum vulgare L.) grains. PLoS ONE 2017, 12, e0183704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Subirà, M.; Romero, M.-P.; Macià, A.; Puig, E.; Romagosa, I.; Moralejo, M. Bioactive compounds and antioxidant capacity in pearling fractions of hulled, partially hull-less and hull-less food barley genotypes. Foods 2021, 10, 565. [Google Scholar] [CrossRef]
- Martínez-Subirà, M.; Romero, M.P.; Macià, A.; Puig, E.; Macià, A.; Romagosa, I.; Moralejo, M. Purple, high β-glucan, hulless barley as valuable ingredient for functional food. Food Sci. Technol. 2020, 131, 109582. [Google Scholar] [CrossRef]
- Martínez-Subirà, M.; Moralejo, M.; Puig, E.; Romero, M.-P.; Savin, R.; Romagosa, I. Impact of rising temperature in the deposition patterns of bioactive compounds in field grown food barley grains. Plants 2021, 10, 598. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W. Phenolic compounds in cereal grains and their health benefits. Cereal Foods World 2007, 52, 105–111. [Google Scholar] [CrossRef]
- Alfieri, M.; Balconi, C.; Cabassi, G.; Habyarimana, E.; Redaelli, R. Antioxidant activity in a set of sorghum landraces and breeding lines. Maydica 2017, 62, 1–7. [Google Scholar]
- Dicko, M.H.; Gruppen, H.; Barro, C.; Traore, A.S.; van Berkel, W.J.; Voragen, A.G. Impact of phenolic compounds and related enzymes in sorghum varieties for resistance and susceptibility to biotic and abiotic stresses. J. Chem. Ecol. 2005, 31, 2671–2688. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, X.; Xiang, W.; Zhu, C.; Lin, Z.; Wu, Y.; Li, J.; Pandravada, S.; Ridder, D.D.; Bai, G.; et al. Presence of tannins in sorghum grains is conditioned by different natural alleles of Tannin1. Proc. Natl. Acad. Sci. USA 2012, 109, 10281–10286. [Google Scholar] [CrossRef] [Green Version]
- Habyarimana, E.; Dall’Agata, M.; De Franceschi, P.; Baloch, F.S. Genome-wide association mapping of total antioxidant capacity, phenols, tannins, and flavonoids in a panel of Sorghum bicolor and S. bicolor × S. halepense populations using multi-locus models. PLoS ONE 2019, 14, e0225979. [Google Scholar] [CrossRef] [Green Version]
- Habyarimana, E.; Lopez-Cruz, M.; Baloch, F.S. Genomic selection for optimum index with dry biomass yield, dry mass fraction of fresh material, and plant height in biomass sorghum. Genes 2020, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Mace, E.; Innes, D.; Hunt, C.; Wang, X.; Tao, Y.; Baxter, J.; Hassall, M.; Hathorn, A.; Jordan, D. The Sorghum QTL stlas: A powerful tool for trait dissection, comparative genomics and crop improvement. Theor. Appl. Genet. 2019, 132, 751–766. [Google Scholar] [CrossRef]
- Boddu, J.; Svabek, C.; Ibraheem, F.; Jones, A.D.; Chopra, S. Characterization of a deletion allele of a sorghum Myb gene yellow seed1 showing loss of 3-deoxyflavonoids. Plant Sci. 2005, 169, 542–552. [Google Scholar] [CrossRef]
- Rhodes, D.H.; Hoffmann, L., Jr.; Rooney, W.L.; Ramu, P.; Morris, G.P.; Kresovich, S. Genome-wide association study of grain polyphenol concentrations in global sorghum [Sorghum bicolor (L.) Moench] germplasm. J. Agric. Food Chem. 2014, 62, 10916–10927. [Google Scholar] [CrossRef]
- Rhodes, D.; Gadgil, P.; Perumal, R.; Tesso, T.; Herald, T.J. Natural Variation and Genome-Wide Association Study of Antioxidants in a Diverse Sorghum Collection. Cereal Chem. 2017, 94, 190–198. [Google Scholar] [CrossRef]
- Klee, H.J.; Giovannoni, J.J. Genetics and control of tomato fruit ripening and quality attributes. Annu. Rev. Genet. 2011, 45, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Gonzali, S.; Mazzucato, A.; Perata, P. Purple as a tomato: Towards high anthocyanin tomatoes. Trends Plant Sci. 2009, 14, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.M.; Mes, P.; Myers, J.R. Characterization and inheritance of the Anthocyanin fruit (Aft) tomato. J. Hered. 2003, 94, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paauw, M.; Koes, R.; Quattrocchio, F.M. Alteration of flavonoid pigmentation patterns during domestication of food crops. J. Exp. Bot. 2019, 70, 3719–3735. [Google Scholar] [CrossRef]
- Bedinger, P.A.; Chetelat, R.T.; McClure, B.; Moyle, L.C.; Rose, J.K.; Stack, S.M.; van der Knaap, E.; Baek, Y.S.; Lopez-Casado, G.; Covey, P.A.; et al. Interspecific reproductive barriers in the tomato clade: Opportunities to decipher mechanisms of reproductive isolation. Sex. Plant Reprod. 2011, 24, 171–187. [Google Scholar] [CrossRef]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.G.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Gonzali, S.; Perata, P. Anthocyanins from Purple Tomatoes as Novel Antioxidants to Promote Human Health. Antioxidants 2020, 9, 1017. [Google Scholar] [CrossRef]
- Sun, C.; Deng, L.; Du, M.; Zhao, J.; Chen, Q.; Huang, T.; Jiang, H.; Li, C.B.; Li, C. A transcriptional network promotes anthocyanin biosynthesis in tomato flesh. Mol. Plant 2020, 13, 42–58. [Google Scholar] [CrossRef]
- Cao, X.; Qiu, Z.; Wang, X.; Van Giang, T.; Liu, X.; Wang, J.; Wang, X.; Gao, J.; Guo, Y.; Du, Y.; et al. A putative R3 MYB repressor is the candidate gene underlying atroviolacium, a locus for anthocyanin pigmentation in tomato fruit. J. Exp. Bot. 2017, 68, 5745–5758. [Google Scholar] [CrossRef] [PubMed]
- Colanero, S.; Perata, P.; Gonzali, S. The atroviolacea gene encodes an R3- MYB protein repressing anthocyanin synthesis in tomato plants. Front. Plant Sci. 2018, 9, 830. [Google Scholar] [CrossRef] [Green Version]
- Povero, G.; Gonzali, S.; Bassolino, L.; Mazzucato, A.; Perata, P. Transcriptional analysis in high-anthocyanin tomatoes reveals synergistic effect of Aft and atv genes. J. Plant Physiol. 2011, 168, 270–279. [Google Scholar] [CrossRef]
- Blando, F.; Berland, H.; Maiorano, G.; Durante, M.; Mazzucato, A.; Picarella, M.E.; Nicoletti, I.; Gerardi, C.; Mita, G.; Andersen, Ø.M. Nutraceutical characterization of anthocyanin-rich fruits produced by “Sun Black” tomato line. Front. Nutr. 2019, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, Y.; Chen, S.; Tian, H.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Lycopene is enriched in tomato fruit by CRISPR/Cas9-mediated multiplex genome editing. Front. Plant Sci. 2018, 9, 559. [Google Scholar] [CrossRef]
- Galpaz, N.; Wang, Q.; Menda, N.; Zamir, D.; Hirschberg, J. Abscisic acid deficiency in the tomato mutant high-pigment 3 leading to increased plastid number and higher fruit lycopene content. Plant J. 2008, 53, 717–730. [Google Scholar] [CrossRef]
- Levin, I.; de Vos, C.H.; Tadmor, Y.; Bovy, A.G.; Lieberman, M.; Oren-Shamir, M.; Segev, O.; Kolotilin, I.; Keller, M.; Ovadia, R.; et al. High pigment tomato mutants—More than just lycopene (a review). Isr. J. Plant Sci. 2006, 54, 179–190. [Google Scholar] [CrossRef]
- Karniel, U.; Koch, A.; Zamir, D.; Hirschberg, J. Development of zeaxanthin-rich tomato fruit through genetic manipulations of carotenoid biosynthesis. Plant Biotechnol. J. 2020, 18, 2292–2303. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Bulling, K.; Hill, L.; Martin, C. Beyond Purple Tomatoes: Combined Strategies Targeting Anthocyanins to Generate Crimson, Magenta, and Indigo Fruit. Horticulturae 2021, 7, 327. [Google Scholar] [CrossRef]
- Scarano, A.; Butelli, E.; De Santis, S.; Cavalcanti, E.; Hill, L.; De Angelis, M.; Giovinazzo, G.; Chieppa, M.; Martin, C.; Santino, A. Combined Dietary Anthocyanins, Flavonols, and Stilbenoids Alleviate Inflammatory Bowel Disease Symptoms in Mice. Front. Nutr. 2018, 4, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a Source of Carotenoids and Polyphenols Targeted to Cancer Prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Sulli, M.; Mandolino, G.; Sturaro, M.; Onofri, C.; Diretto, G.; Parisi, B.; Giuliano, G. Molecular and biochemical characterization of a potato collection with contrasting tuber carotenoid content. PLoS ONE 2017, 12, e0184143. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; Herrera, M.; Hoffmann, L.; Hausman, J.F.; Larondelle, Y.; Evers, D. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant and mineral micronutrients. J. Agric. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef]
- Martínez-Ispizua, E.; Calatayud, Á.; Marsal, J.I.; Mateos-Fernández, R.; Díez, M.J.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.R. Phenotyping local eggplant varieties: Commitment to biodiversity and nutritional quality preservation. Front. Plant Sci. 2021, 12, 696272. [Google Scholar] [CrossRef]
- Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef] [Green Version]
- Sora, G.T.; Haminiuk, C.W.; da Silva, M.V.; Zielinski, A.A.; Gonçalves, G.A.; Bracht, A.; Peralta, R.M. A comparative study of the capsaicinoid and phenolic contents and in vitro antioxidant activities of the peppers of the genus Capsicum: An application of chemometrics. J. Food Sci. Technol. 2015, 52, 8086–8094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcio Gaudino, E.; Colletti, A.; Grillo, G.; Tabasso, S.; Cravotto, G. Emerging Processing Technologies for the Recovery of Valuable Bioactive Compounds from Potato Peels. Foods 2020, 9, 1598. [Google Scholar] [CrossRef]
- Pacifico, D.; Musmeci, S.; del Pulgar, J.S.; Onofri, C.; Parisi, B.; Sasso, R.; Mandolino, G.; Lombardi-Boccia, G. Caffeic Acid and α-Chaconine Influence the Resistance of Potato Tuber to Phthorimaea operculella (Lepidoptera: Gelechiidae). Am. J. Potato Res. 2019, 96, 403–413. [Google Scholar] [CrossRef]
- De Masi, L.; Bontempo, P.; Rigano, D.; Stiuso, P.; Carafa, V.; Nebbioso, A.; Piacente, S.; Montoro, P.; Aversano, R.; D’Amelia, V.; et al. Comparative phytochemical characterization, genetic profile, and antiproliferative activity of polyphenol-rich extracts from pigmented tubers of different solanum tuberosum varieties. Molecules 2020, 25, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; de Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef]
- Docimo, T.; Francese, G.; Ruggiero, A.; Batelli, G.; De Palma, M.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Mennella, G.; Tucci, M. Phenylpropanoids Accumulation in Eggplant Fruit: Characterization of Biosynthetic Genes and Regulation by a MYB Transcription Factor. Front. Plant Sci. 2016, 6, 1233. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, H.; Goyer, A.; Navarre, D.A. Antioxidants in potatoes: A functional view on one of the major food crops worldwide. Molecules 2021, 26, 2446. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Florio, F.E.; Fibiani, M.; Speranza, G.; Rabuffetti, M.; Gattolin, S.; Toppino, L.; Rotino, G.L. Scrapped but not neglected: Insights into the composition, molecular modulation and antioxidant capacity of phenols in peel of eggplant (Solanum melongena L.) fruits at different developmental stages. Plant Physiol. Biochem. 2021, 167, 678–690. [Google Scholar] [CrossRef]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engineering of potato carotenoid content through tuber-specific overexpression of a bacterial mini-pathway. PLoS ONE 2007, 2, e350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Römer, S.; Lübeck, J.; Kauder, F.; Steiger, S.; Adomat, C.; Sandmann, G. Genetic engineering of a zeaxanthin-rich potato by antisense inactivation and co-suppression of carotenoid epoxidation. Metab. Eng. 2002, 4, 263–272. [Google Scholar] [CrossRef] [PubMed]
- van Eck, J.; Conlin, B.; Garvin, D.F.; Mason, H.; Navarre, D.A.; Brown, C.R. Enhancing beta-carotene content in potato by RNAi-mediated silencing of the beta-carotene hydroxylase gene. Am. J. Potato Res. 2007, 84, 331–342. [Google Scholar] [CrossRef]
- Mishiba, K.I.; Nishida, K.; Inoue, N.; Fujiwara, T.; Teranishi, S.; Iwata, Y.; Takeda, S.; Koizumi, N. Genetic engineering of eggplant accumulating β-carotene in fruit. Plant Cell Rep. 2020, 39, 1029–1039. [Google Scholar] [CrossRef]
- Luo, J.; Butelli, E.; Hill, L.; Parr, A.; Niggeweg, R.; Bailey, P.; Weisshaar, B.; Martin, C. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: Expression in fruit results in very high levels of both types of polyphenol. Plant J. 2008, 56, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, W.; Chen, J.; Jia, R.; Ma, L.; Wang, S.; Wang, J.; Shen, X.; Chu, Z.; Zhu, C.; et al. Development of Marker-Free Transgenic Potato Tubers Enriched in Caffeoylquinic Acids and Flavonols. J. Agric. Food Chem. 2016, 64, 2932–2940. [Google Scholar] [CrossRef]
- Kostyn, K.; Szatkowski, M.; Kulma, A.; Kosieradzka, I.; Szopa, J. Transgenic potato plants with overexpression of dihydroflavonol reductase can serve as efficient nutrition sources. J. Agric. Food Chem. 2013, 61, 6743–6753. [Google Scholar] [CrossRef]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Martens, S.; Mock, H.P. Metabolite profiling of red and blue potatoes revealed cultivar and tissue specific patterns for anthocyanins and other polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef]
- Tatarowska, B.; Milczarek, D.; Wszelaczyńska, E.; Pobereżny, J.; Keutgen, N.; Keutgen, A.J.; Flis, B. Carotenoids variability of potato tubers in relation to genotype, growing location and year. Am. J. Potato Res. 2019, 96, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Thorup, T.A.; Tanyolac, B.; Livingstone, K.D.; Popovsky, S.; Paran, I.; Jahn, M. Candidate gene analysis of organ pigmentation loci in the Solanaceae. Proc. Natl. Acad. Sci. USA 2020, 97, 11192–11197. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.R.; Edwards, C.G.; Yang, C.P.; Dean, B.B. Segregation of Total carotenoid in high level potato germplasm and its relationship to beta-carotene hydroxylase polymorphism. Am. J. Potato Res. 2006, 83, 365–372. [Google Scholar] [CrossRef]
- Wolters, A.M.; Uitdewilligen, J.G.; Kloosterman, B.A.; Hutten, R.C.; Visser, R.G.; van Eck, H.J. Identification of alleles of carotenoid pathway genes important for zeaxanthin accumulation in potato tubers. Plant Mol. Biol. 2020, 73, 659–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellumori, M.; Chasquibol Silva, N.A.; Vilca, L.; Andrenelli, L.; Cecchi, L.; Innocenti, M.; Balli, D.; Mulinacci, N. A Study on the biodiversity of pigmented andean potatoes: Nutritional profile and phenolic composition. Molecules 2020, 25, 3169. [Google Scholar] [CrossRef] [PubMed]
- Florio, F.E.; Gattolin, S.; Toppino, L.; Bassolino, L.; Fibiani, M.; Lo Scalzo, R.; Rotino, G.L.A. SmelAAT acyltransferase variant causes a major difference in eggplant (solanum melongena L.) peel anthocyanin composition. Int. J. Mol. Sci. 2021, 22, 9174. [Google Scholar] [CrossRef]
- Plazas, M.; Andújar, I.; Vilanova, S.; Hurtado, M.; Gramazio, P.; Herraiz, F.J.; Prohens, J. Breeding for chlorogenic acid content in eggplant: Interest and prospects. Not. Bot. Horti. Agrobot. Cluj-Napoca 2013, 41, 26. [Google Scholar] [CrossRef] [Green Version]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 editing of three ppo genes reduces fruit flesh browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jang, S.J.; Jeong, H.B.; Lee, S.Y.; Venkatesh, J.; Lee, J.H.; Kwon, J.K.; Kang, B.C. A mutation in Zeaxanthin epoxidase contributes to orange coloration and alters carotenoid contents in pepper fruit (Capsicum annuum). Plant J. 2021, 106, 1692–1707. [Google Scholar] [CrossRef]
- Deepa, N.; Kaur, C.; Binoy, G.; Singh, B.; Kapoor, H.C. Antioxidant constituents in some sweet pepper (Capsicum annuum L.) genotypes during maturity. LWT-Food Sci. Technol. 2007, 40, 121–129. [Google Scholar] [CrossRef]
- Gómez-García, M.; Ochoa-Alejo, N. Biochemistry and molecular biology of carotenoid biosynthesis in chili peppers (Capsicum spp.). Int. J. Mol. Sci. 2013, 14, 19025–19053. [Google Scholar] [CrossRef] [Green Version]
- Matus, Z.; Deli, J.; Szabolcs, J. Carotenoid composition of yellow pepper during ripening: Isolation of β-cryptoxanthin 5,6-epoxide. J. Agric. Food Chem. 1991, 39, 1907–1914. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Barański, M.; Srednicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, C.; et al. Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analyses. Br. J. Nutr. 2014, 112, 794–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Génard, M. Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Michalskaa, A.; Wojdyłob, A.; Bogucka, B. The influence of nitrogen and potassium fertilisation on the content of polyphenolic compounds and antioxidant capacity of colored potato. J. Food Comp. Anal. 2016, 47, 69–75. [Google Scholar] [CrossRef]
- Mitchell, A.E.; Hong, Y.J.; Koh, E.; Barrett, D.M.; Bryant, D.E.; Denison, R.F.; Kaffka, S. Ten-year comparison of the influence of organic and conventional crop management practices on the content of flavonoids in tomatoes. J. Agric. Food Chem. 2007, 55, 6154–6159. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, D.; Casciani, R.; Ritota, M.; Mandolino, G.; Onofri, C.; Moschella, A.; Parisi, B.; Cafiero, C.; Valentini, M. NMR-Based metabolomics for organic farming traceability of early potatoes. J. Agric. Food Chem. 2013, 61, 11201–11211. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Średnicka-Tober, D.; Hallmann, E.; Kopczyńska, K.; Zarzyńska, K. The impact of organic vs. conventional agricultural practices on selected quality features of eight potato cultivars. Agronomy 2019, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, D.; Paris, R. Effect of organic potato farming on human and environmental health and benefits from new plant breeding techniques. is it only a matter of public acceptance? Sustainability 2016, 8, 1054. [Google Scholar] [CrossRef] [Green Version]
- Dimberg, L.H.; Gissén, C.; Nilsson, J. Phenolic compounds in oat grains (Avena sativa L.) grown in conventional and organic systems. Ambio 2005, 34, 331–337. [Google Scholar] [CrossRef]
- Kesarwani, A.; Chiang, P.Y.; Chen, S.S. Distribution of phenolic compounds and antioxidative activities of rice kernel and their relationships with agronomic practice. Science 2014, 2014, 620171. [Google Scholar] [CrossRef] [Green Version]
- Luthria, D.; Singh, A.P.; Wilson, T.; Vorsa, N.; Banuelos, G.S.; Vinyard, B.T. Influence of conventional and organic agricultural practices on the phenolic content in eggplant pulp: Plant-to-plant variation. Food Chem. 2010, 121, 406–411. [Google Scholar] [CrossRef]
- Szafirowska, A.; Elkner, K. The comparison of yielding and nutritive value of organic and conventional pepper fruits. Veg. Crops Res. Bull. 2019, 71, 111–121. [Google Scholar] [CrossRef]
- Chassy, A.W.; Bui, L.; Renaud, E.N.C.; Van Horn, M.; Mitchell, A.E. Three-year comparison of the content of antioxidant microconstituents and several quality characteristics in organic and conventionally managed tomatoes and bell peppers. J. Agric. Food Chem. 2006, 54, 8244–8252. [Google Scholar] [CrossRef] [PubMed]
- Marín, A.; Gil, M.I.; Flores, P.; Hellín, P.; Selma, M.V. Microbial quality and bioactive constituents of sweet peppers from sustainable production systems. J. Agric. Food Chem. 2008, 56, 11334–11341. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Suseela, V. Unraveling Arbuscular mycorrhiza-induced changes in plant primary and secondary metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Gundel, P.E.; Helander, M.; Saikkoonen, K. Endophytic mediation of reactive oxygen species and antioxidant activity in plants: A review. Fungal Divers. 2012, 54, 1–10. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the ideotype mycorrhizal symbionts for the production of healthy food. Front. Plant Sci. 2018, 9, 1089. [Google Scholar] [CrossRef]
- Skendi, A. Alternatives to increase the antioxidant capacity of bread with phenolics. In Trends in Wheat and Bread Making; Elsevier: Amsterdam, The Netherlands, 2021; Chapter 11. [Google Scholar]
- Giovannetti, M.; Avio, L.; Barale, R.; Ceccarelli, N.; Cristofani, R.; Iezzi, A.; Mignolli, F.; Picciarelli, P.; Pinto, B.; Reali, D.; et al. Nutraceutical value and safety of tomato fruits produced by mycorrhizal plants. Br. J. Nutr. 2012, 107, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Aal, E.S.; Young, J.C.; Rabalski, I. Anthocyanin Composition in Black, Blue, Pink, Purple, and Red Cereal Grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef]
- Ficco, D.B.M.; De Simone, V.; De Leonardis, A.M.; Giovanniello, V.; Del Nobile, M.A.; Padalino, L.; Lecce, L.; Borrelli, G.M.; De Vita, P. Use of purple durum wheat to produce naturally functional fresh and dry pasta. Food Chem. 2016, 205, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Kasote, D.; Tiozon, R.N.; Sartagoda, K.J.D.; Itagi, H.; Roy, P.; Kohli, A.; Regina, A.; Sreenivasulu, N. Food processing technologies to develop functional foods with enriched bioactive phenolic compounds in cereals. Front. Plant Sci. 2021, 12, 2660. [Google Scholar] [CrossRef] [PubMed]
- Mert, I.D. The applications of microfluidization in cereals and cereal-based products: An overview. Crit. Rev. Food Sci. Nutr. 2020, 60, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Ficco, D.B.M.; Borrelli, G.M.; Miedico, O.; Giovanniello, V.; Tarallo, M.; Pompa, C.; De Vita, P.; Chiaravalle, A.E. Effects of grain debranning on bioactive compounds, antioxidant capacity and essential and toxinc trace elements in purple durum wheats. LWT Food Sci. Technol. 2020, 118, 108734. [Google Scholar] [CrossRef]
- Ficco, D.B.M.; Borrelli, G.M.; Giovanniello, V.; Platani, C.; De Vita, P. Production of anthocyanin-enriched flours of durum and soft pigmented wheats by air-classification, as a potential ingredient for functional bread. J. Cereal Sci. 2018, 79, 118–126. [Google Scholar] [CrossRef]
- Cilla, A.; Bosch, L.; Barberá, R.; Alegría, A. Effect of processing on the bioaccessibility of bioactive compounds—A review focusing on carotenoids, minerals, ascorbic acid, tocopherols and polyphenols. J. Food Compos. Anal. 2018, 68, 3–15. [Google Scholar] [CrossRef]
- Paznocht, L.; Kotikova, Z.; Orsak, M.; Lachman, J.; Martinek, P. Carotenoid changes of colored-grain wheat flours during bun-making. Food Chem. 2017, 277, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Bartl, P.; Albreht, A.; Skrt, M.; Tremlova, B.; Ostadlova, M.; Smejkal, K.; Vovk, I.; Ulrih, N.P. Anthocyanins in purple and blue wheat grains and in resulting bread: Quantity, composition, and thermal stability. Int. J. Food Sci. Nutr. 2015, 66, 514–519. [Google Scholar] [CrossRef]
- Carcea, M.; Narducci, V.; Turfani, V.; Giannini, V. Polyphenols in Raw and Cooked Cereals/Pseudocereals/Legume Pasta and Couscous. Foods 2017, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Rytel, E.; Tajner-Czopek, A.; Kita, A.; Tkaczyńska, A.; Kucharska, A.Z.; Sokół-Łętowska, A. The Influence of the Production Process on the Anthocyanin Content and Composition in Dried Potato Cubes, Chips, and French Fries Made from Red-Fleshed Potatoes. Appl. Sci. 2021, 11, 1104. [Google Scholar] [CrossRef]
- Arkoub-Djermoune, L.; Boulekbache-Makhlouf, L.; Zeghichi-Hamri, S.; Bellili, S.; Boukhalfa, F.; Madani, K. Influence of the thermal processing on the physicochemical properties and the antioxidant activity of a Solanaceae vegetable: Eggplant. J. Food Qual. 2016, 39, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Villa-Rivera, M.G.; Ochoa-Alejo, N. Chili Pepper Carotenoids: Nutraceutical Properties and Mechanisms of Action. Molecules 2020, 25, 5573. [Google Scholar] [CrossRef] [PubMed]
- Lachman, J.; Hamouz, K.; Orsák, M.; Pivec, V.; Hejtmánková, K.; Pazderů, K.; Dvořák, P.; Čepl, J. Impact of selected factors—Cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 2012, 133, 1107–1116. [Google Scholar] [CrossRef]
- Girard, A.L.; Awika, J.M. Effects of edible plant polyphenols on gluten protein functionality and potential applications of polyphenol–gluten interactions. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2164–2199. [Google Scholar] [CrossRef]
- Van den Berg, H.; Faulks, R.; Fernando Granado, H.; Hirschberg, J.; Olmedilla, B.; Sandmann, G.; Southon, S.; Stahl, W. The potential for the improvement of carotenoid levels in foods and the likely systemic effects. J. Sci. Food Agric. 2000, 80, 880–912. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Y.; Zhang, H.; Mai, Q.; Zhang, B.; Li, H.; Deng, Z. The degradation rules of anthocyanins from eggplant peel and antioxidant capacity in fortified model food system during the thermal treatments. Food Biosci. 2020, 38, 100701. [Google Scholar] [CrossRef]
- Chitchumroonchokchai, C.; Diretto, G.; Parisi, B.; Giuliano, G.; Failla, M.L. Potential of golden potatoes to improve vitamin A and vitamin E status in developing countries. PLoS ONE 2017, 12, e0187102. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Chen, J.; Lv, F.; Chen, S.; Chen, J.; Liu, D.; Ye, X. Domestic cooking methods affect the phytochemical composition and antioxidant activity of purple-fleshed potatoes. Food Chem. 2016, 197, 1264–1270. [Google Scholar] [CrossRef]
- Patras, A.; Brunton, N.; Da Pieve, S.; Butler, F.; Downey, G. Effect of high-pressure processing on antioxidant activity and instrumental colour of tomato and carrot purees. Innov. Food Sci. Emerg. Technol. 2008, 10, 16–22. [Google Scholar] [CrossRef]
- Andres, V.; Villanueva, M.J.; Tenorio, M.D. The effect of high-pressure processing on colour, bioactive compounds, and antioxidant activity in smoothies during refrigerated storage. Food Chem. 2016, 192, 328–335. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Kay, C.D.; Kroon, P.A.; Cassidy, A. The bioactivity of dietary anthocyanins is likely to be mediated by their degradation products. Mol. Nutr. Food Res. 2009, 53 (Suppl. 1), S92–S101. [Google Scholar] [CrossRef]
- Aura, A.M.; Martin-Lopez, P.; O’Leary, K.A.; Williamson, G.; Oksman-Caldentey, K.M.; Poutanen, K.; Santos-Buelga, C. In vitro metabolism of anthocyanins by human gut microflora. Eur. J. Nutr. 2005, 44, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Virgili, F.; Marino, M. Regulation of cellular signals from nutritional molecules, a specific role for phytochemicals, beyond antioxidant activity. Free Radic. Biol. Med. 2008, 45, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Speciale, A.; Anwar, S.; Canali, R.; Chirafisi, J.; Saija, A.; Virgili, F.; Cimino, F. Cyanidin-3-O-glucoside counters the response to tnf-alpha of endothelial cells by activating nrf2 pathway. Mol. Nutr. Food Res. 2013, 57, 1979–1987. [Google Scholar] [CrossRef]
- Speciale, A.; Chirafisi, J.; Saija, A.; Cimino, F. Nutritional antioxidants and adaptive cell responses, an update. Curr. Mol. Med. 2011, 11, 770–789. [Google Scholar] [CrossRef]
- Kim, S.H.; Son, G.H.; Bhattacharjee, S.; Kim, H.J.; Nam, J.C.; Nguyen, P.D.; Hong, J.C.; Gassmann, W. The Arabidopsis immune adaptor SRFR1 interacts with TCP transcription factors that redundantly contribute to effector-triggered immunity. Plant J. 2014, 78, 978–989. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The case for anthocyanin consumption to promote human health: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Tunon, M.J.; Garcia-Mediavilla, M.V.; Sanchez-Campos, S.; Gonzalez-Gallego, J. Potential of flavonoids as anti-inflammatory agents, modulation of pro-inflammatory gene expression and signal transduction pathways. Curr. Drug Metab. 2009, 10, 256–271. [Google Scholar] [CrossRef]
- Vendrame, S.; Klimis-Zacas, D. Anti-inflammatory effect of anthocyanins via modulation of nuclear factor-kB and mitogen-activated protein kinase signaling cascades. Nutr. Rev. 2015, 73, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Hassimotto, N.M.; Moreira, V.; do Nascimento, N.G.; Souto, P.C.; Teixeira, C.; Lajolo, F.M. Inhibition of carrageenan-induced acute inflammation in mice by oral administration of anthocyanin mixture from wild mulberry and cyanidin-3-glucoside. Biomed. Res. Int. 2013, 2013, 146716. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ling, W.; Wang, Q.; Liu, C.; Hu, Y.; Xia, M.; Feng, X.; Xia, X. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. indica) on hyperlipidemia and insulin resistance in fructose-fed rats. Plant Foods Hum. Nutr. 2007, 62, 1–6. [Google Scholar] [CrossRef]
- Salgado, J.M.; Oliveira, A.G.; Mansi, D.N.; Donado-Pestana, C.M.; Bastos, C.R.; Marcondes, F.K. The role of black rice (Oryza sativa L.) in the control of hypercholesterolemia in rats. J. Med. Food 2010, 13, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Andrews, M.C.; Hu, Y.; Wang, D.; Qin, Y.; Zhu, Y.; Ni, H.; Ling, W. Anthocyanin extract from black rice significantly ameliorates platelet hyperactivity and hypertriglyceridemia in dyslipidemic rats induced by high fat diets. J. Agric. Food Chem. 2011, 59, 6759–6764. [Google Scholar] [CrossRef]
- Wang, H.; Liu, D.; Ji, Y.; Liu, Y.; Xu, L.; Guo, Y. Dietary supplementation of black rice anthocyanin extract regulates cholesterol metabolism and improves gut microbiota dysbiosis in C57BL/6J mice fed a high-fat and cholesterol diet. Mol. Nutr. Food Res. 2020, 64, 1900876. [Google Scholar] [CrossRef]
- Ling, W.H.; Cheng, Q.X.; Ma, J.; Wang, T. Red and black rice decrease atherosclerotic plaque formation and increase antioxidant status in rabbits. J. Nutr. 2001, 131, 1421–1426. [Google Scholar] [CrossRef]
- Ling, W.H.; Wang, L.L.; Ma, J. Supplementation of the black rice outer layer fraction to rabbits decreases atherosclerotic plaque formation and increases antioxidant status. J. Nutr. 2002, 132, 20–26. [Google Scholar] [CrossRef]
- Xia, X.; Ling, W.; Ma, J.; Xia, M.; Hou, M.; Wang, Q.; Zhu, H.; Tang, Z. An anthocyanin-rich extract from black rice enhances atherosclerotic plaque stabilization in apolipoprotein e-deficient mice. J. Nutr. 2006, 136, 2220–2225. [Google Scholar] [CrossRef]
- Sharma, S.; Khare, P.; Kumar, A.; Chunduri, V.; Kumar, A.; Kapoor, P.; Mangal, P.; Kondepudi, K.K.; Bishnoi, M.; Garg, M. Anthocyanin-biofortified colored wheat prevents high fat diet–induced alterations in mice, nutrigenomics studies. Mol. Nutr. Food Res. 2020, 64, 1900999. [Google Scholar] [CrossRef]
- Hou, Z.; Qin, P.; Ren, G. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. Japonica) on chronically alcohol-induced liver damage in rats. J. Agric. Food Chem. 2010, 58, 3191–3196. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Guo, H.; Shen, T.; Tang, X.; Yang, Y.; Ling, W. Cyanidin-3-O-β-glucoside purified from black rice protects mice against hepatic fibrosis induced by carbon tetrachloride via inhibiting hepatic stellate cell activation. J. Agric. Food Chem. 2015, 63, 6221–6230. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhou, Y.; Wu, T.; Hao, L. Ameliorative effect of black rice anthocyanin on senescent mice induced by D-galactose. Food Funct. 2014, 5, 2892–2897. [Google Scholar] [CrossRef] [PubMed]
- Pannangrong, W.; Wattanathorn, J.; Muchimapura, S.; Tiamkao, S.; Tong-Un, T. Purple rice berry is neuroprotective and enhances cognition in a rat model of Alzheimer’s disease. J. Med. Food. 2011, 14, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Tikhonova, M.A.; Shoeva, O.Y.; Tenditnik, M.V.; Ovsyukova, M.V.; Akopyan, A.A.; Dubrovina, N.I.; Amstislavskaya, T.G.; Khlestkina, E.K. Evaluating the Effects of Grain of Isogenic Wheat Lines Differing in the Content of Anthocyanins in Mouse Models of Neurodegenerative Disorders. Nutrients 2020, 12, 3877. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Z.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The therapeutic potential of anthocyanins, current approaches based on their molecular mechanism of action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef]
- Toufektsian, M.C.; de Lorgeril, M.; Nagy, N.; Salen, P.; Donati, M.B.; Giordano, L.; Mock, H.P.; Peterek, S.; Matros, A.; Petroni, K.; et al. dietary intake of plant-derived anthocyanins protects the rat heart against ischemia-reperfusion injury. J. Nutr. 2008, 138, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Trinei, M.; Carpi, A.; Menabo’, R.; Storto, M.; Fornari, M.; Marinelli, A.; Minardi, S.; Riboni, M.; Casciaro, F.; DiLisa, F.; et al. Dietary intake of cyanidin-3-glucoside induces a long-lasting cardioprotection from ischemia/reperfusion injury by altering the microbiota. J. Nutr. Biochem. 2022, 101, 108921. [Google Scholar] [CrossRef]
- Petroni, K.; Trinei, M.; Fornari, M.; Calvenzani, V.; Marinelli, A.; Micheli, L.A.; Pilu, R.; Matros, A.; Mock, H.P.; Tonelli, C.; et al. Dietary cyanidin 3-glucoside from purple corn ameliorates doxorubicin-induced cardiotoxicity in mice. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 462–469. [Google Scholar] [CrossRef]
- Li, J.; Kang, M.K.; Kim, J.K.; Kim, J.L.; Kang, S.W.; Lim, S.S.; Kang, Y.H. Purple corn anthocyanins retard diabetes-associated glomerulosclerosis in mesangial cells and db/db mice. Eur. J. Nutr. 2012, 51, 961–973. [Google Scholar] [CrossRef]
- Kang, M.K.; Li, J.; Kim, J.L.; Gong, J.H.; Kwak, S.N.; Park, J.H.; Lee, J.Y.; Lim, S.S.; Kang, Y.H. Purple corn anthocyanins inhibit diabetesassociated glomerular monocyte activation and macrophage infiltration. Am. J. Physiol. Renal. Physiol. 2012, 303, F1060–F1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.K.; Lim, S.S.; Lee, J.Y.; Yeo, K.M.; Kang, Y.H. Anthocyanin-rich purple corn extract inhibit diabetes-associated glomerular angiogenesis. PLoS ONE 2013, 8, e79823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, T.; Horio, F.; Uchida, K.; Aoki, H.; Osawa, T. Dietary cyanidin 3-O-β-D-glucoside rich purple corn color prevents obesity and ameliorates hyperglycemia in mice. J. Nutr. 2003, 133, 2125–2130. [Google Scholar] [CrossRef]
- Luna-Vital, D.A.; de Mejia, E.G. Anthocyanins from purple corn activate free fatty acid-receptor 1 and glucokinase enhancing in vitro insulin secretion and hepatic glucose uptake. PLoS ONE 2018, 13, e0200449. [Google Scholar] [CrossRef] [Green Version]
- Luna-Vital, D.; Luzardo-Ocampo, I.; Cuellar-Nuñez, L.; Loarca-Pina, G.; de Mejia, E.G. Maize extract rich in ferulic acid and anthocyanins prevents high-fat induced obesity in mice by modulating SIRT1, AMPK, and IL-6 associated metabolic and inflammatory pathways. J. Nutr. Biochem. 2020, 79, 108343. [Google Scholar] [CrossRef] [PubMed]
- Magni, G.; Marinelli, A.; Riccio, D.; Lecca, D.; Tonelli, C.; Abbracchio, M.P.; Petroni, K.; Ceruti, S. Purple Corn Extract as Anti-allodynic Treatment for Trigeminal Pain, Role of Microglia. Front. Cell Neurosci. 2018, 12, 378. [Google Scholar] [CrossRef] [PubMed]
- Saclier, M.; Bonfanti, C.; Antonini, S.; Angelini, G.; Mura, G.; Zanaglio, F.; Taglietti, V.; Romanello, V.; Sandri, M.; Tonelli, C.; et al. Nutritional intervention with cyanidin hinders the progression of muscular dystrophy. Cell Death Dis. 2020, 11, 127. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Deng, Z.; Liu, R.; Loewen, S.; Tsao, R. Bioaccessibility, in vitro antioxidant activities and in vivo anti-inflammatory activities of a purple tomato (Solanum lycopersicum L.). Food Chem. 2014, 159, 353–360. [Google Scholar] [CrossRef]
- Tomlinson, M.L.; Butelli, E.; Martin, C.; Carding, S.R. Flavonoids from Engineered Tomatoes Inhibit Gut Barrier Pro-inflammatory Cytokines and Chemokines, via SAPK/JNK and p38 MAPK Pathways. Front. Nutr. 2017, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Liso, M.; De Santis, S.; Scarano, A.; Verna, G.; Dicarlo, M.; Galleggiante, V.; Campiglia, P.; Mastronardi, M.; Lippolis, A.; Vacca, M.; et al. Bronze-tomato enriched diet affects the intestinal microbiome under homeostatic and inflammatory conditions. Nutrients 2018, 10, 1862. [Google Scholar] [CrossRef] [Green Version]
- Sido, A.; Radhakrishnan, S.; Kim, S.W.; Eriksson, E.; Shen, F.; Li, Q.; Bhat, V.; Reddivari, L.; Vanamala, J. A food-based approach that targets interleukin-6, a key regulator of chronic intestinal inflammation and colon carcinogenesis. J. Nutr. Biochem. 2017, 43, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddivari, L.; Wang, T.; Wu, B.; Li, S. Potato, an Anti-Inflammatory Food. Am. J. Potato Res. 2019, 96, 164–169. [Google Scholar] [CrossRef]
- Lee, S.-J.; Shin, J.-S.; Choi, H.-E.; Lee, K.-G.; Cho, Y.-W.; An, H.-J.; Jang, D.S.; Jeong, J.-C.; Kwon, O.-K.; Nam, J.-H.; et al. Chloroform fraction of Solanum tuberosum L. cv Jayoung epidermis suppresses LPS-induced inflammatory responses in macrophages and DSS induced colitis in mice. Food Chem. Toxicol. 2014, 63, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hibasami, H.; Murakami, T.; Terahara, N.; Mori, M.; Tsukui, A. Induction of apoptosis in cultured human stomach cancer cells by potato anthocyanins and its inhibitory effects on growth of stomach cancer in mice. Food Sci. Technol. Res. 2006, 12, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Thompson, M.D.; Thompson, H.J.; McGinley, J.N.; Neil, E.S.; Rush, D.K.; Holm, D.G.; Stushnoff, C. Functional food characteristics of potato cultivars (Solanum tuberosum L.), phytochemical composition and inhibition. J. Food Compos. Anal. 2009, 22, 571–576. [Google Scholar] [CrossRef]
- Yoon, S.S.; Rhee, Y.H.; Lee, H.J.; Lee, E.O.; Lee, M.H.; Ahn, K.S.; Lim, H.T.; Kim, S.H. Uncoupled protein 3 and p38 signal pathways are involved in antiobesity activity of Solanum tuberosum L. cv. Bora Valley. J. Ethnopharmacol. 2008, 118, 396–404. [Google Scholar] [CrossRef]
- Kubow, S.; Hobson, L.; Iskandar, M.M.; Sabally, K.; Donnelly, D.J.; Agellonm, L.B. Extract of Irish potatoes (Solanum tuberosum L.) decreases body weight gain and adiposity and improves glucose control in the mouse model of diet-induced obesity. Mol. Nutr. Food Res. 2014, 58, 2235–2238. [Google Scholar] [CrossRef]
- Singh, N.; Kamath, V.; Rajini, P.S. Attenuation of hyperglycemia and associated biochemical parameters in STZ-induced diabetic rats by dietary supplementation of potato peel powder. Clin. Chim. Acta 2005, 353, 165–175. [Google Scholar] [CrossRef]
- Han, K.H.; Hashimoto, N.; Shimada, K.; Sekikawa, M.; Noda, T.; Yamauchi, H.; Hashimoto, M.; Chiji, H.; Topping, D.L.; Fukushima, M. Hepatoprotective effects of purple potato extract against D-galactosamine-induced liver injury in rats. Biosci. Biotechnol. Biochem. 2006, 70, 1432–1437. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Matsumoto, A.; Shimada, K.; Sekikawa, M.; Fukushima, M. Effects of anthocyanin-rich purple potato flakes on antioxidant status in F344 rats fed a cholesterol-rich diet. Br. J. Nutr. 2007, 98, 914–921. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Shimada, K.; Sekikawa, M.; Fukushima, M. Anthocyanin-rich red potato flakes affect serum lipid peroxidation and hepatic SOD mRNA level in rats. Biosci. Biotechnol. Biochem. 2007, 71, 1356–1359. [Google Scholar] [CrossRef] [PubMed]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Muller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review, vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McRae, M.P. Health Benefits of Dietary Whole Grains: An Umbrella Review of Meta-analyses. J. Chiropr. Med. 2017, 16, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshin, A.; Sur, P.J.; Fay, K.A.; Cornaby, L.; Ferrara, G.; Salama, J.S.; Mullany, E.C.; Abate, K.H.; Abbafati, C.; Abebe, Z.; et al. Health effects of dietary risks in 195 countries, 1990–2017, A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef] [Green Version]
- Pounis, G.; Di Castelnuovo, A.; Bonaccio, M.; Costanzo, S.; Persichillo, M.; Krogh, V.; Donati, M.B.; De Gaetano, G.; Iacoviello, L. Flavonoid and lignan intake in a Mediterranean population, proposal for a holistic approach in polyphenol dietary analysis, the Moli-sani Study. Eur. J. Clin. Nutr. 2016, 70, 338–345. [Google Scholar] [CrossRef]
- Pounis, G.; Bonaccio, M.; Di Castelnuovo, A.; Costanzo, S.; De Curtis, A.; Persichillo, M.; Sieri, S.; Donati, M.B.; Cerletti, C.; De Gaetano, G.; et al. Polyphenol intake is associated with low-grade inflammation, using a novel data analysis from the Moli-sani study. J. Thromb. Haemost. 2016, 115, 344–352. [Google Scholar] [CrossRef]
- Pounis, G.; Costanzo, S.; Bonaccio, M.; Di Castelnuovo, A.; de Curtis, A.; Ruggiero, E.; Persichillo, M.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; et al. Reduced mortality risk by a polyphenol-rich diet, an analysis from the Moli-sani study. Nutrition 2018, 48, 87–95. [Google Scholar] [CrossRef]
- Esposito, S.; Gialluisi, A.; Costanzo, S.; Di Castelnuovo, A.; Ruggiero, E.; De Curtis, A.; Persichillo, M.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; et al. Dietary polyphenol intake is associated with biological aging, a novel predictor of cardiovascular disease, cross-sectional findings from the moli-sani study. Nutrients 2021, 13, 1701. [Google Scholar] [CrossRef]
- Luís, Â.; Domingues, F.; Pereira, L. Association between berries intake and cardiovascular diseases risk factors, a systematic review with meta-analysis and trial sequential analysis of randomized controlled trials. Food Funct. 2018, 9, 740–757. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Willett, W.C.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Seal, C.J.; Courtin, C.M.; Venema, K.; de Vries, J. Health benefits of whole grain, Effects on dietary carbohydrate quality, the gut microbiome and consequences of processing. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2742–2768. [Google Scholar] [CrossRef] [PubMed]
- Lillioja, S.; Neal, A.L.; Tapsell, L.; Jacobs, D.R., Jr. Whole grains, type 2 diabetes, coronary heart disease, and hypertension, links to the aleurone preferred over indigestible fiber. Biofactors 2013, 39, 242–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- U.S. Department of Health and Human Services; U.S. Department of Agriculture. 2015–2020 Dietary Guidelines for Americans, 8th ed.; December 2015. Available online: http://health.gov/dietaryguidelines/2015/guidelines/ (accessed on 3 November 2021).
- EFSA panel on dietetic products, nutrition, and allergies (nda); scientific opinion on dietary reference values for carbohydrates and dietary fibre. EFSA J. 2010, 8, 1462. Available online: www.efsa.europa.eu (accessed on 3 November 2021).
- Gupta, R.; Meghwal, M.; Prabhakar, P.K. Bioactive compounds of pigmented wheat (Triticum aestivum), potential benefits in human health. Trends Food Sci. Technol. 2021, 110, 240–252. [Google Scholar] [CrossRef]
- Chusak, C.; Pasukamonset, P.; Chantarasinlapin, P.; Adisakwattana, S. Postprandial glycemia, insulinemia, and antioxidant status in healthy subjects after ingestion of bread made from anthocyanin-rich riceberry rice. Nutrients 2020, 12, 782. [Google Scholar] [CrossRef] [Green Version]
- Callcott, E.T.; Blanchard, C.L.; Snell, P.; Santhakumar, A.B. The anti-inflammatory and antioxidant effects of acute consumption of pigmented rice in humans. Food Funct. 2019, 10, 8230. [Google Scholar] [CrossRef]
- Liso, M.; Sila, A.; Verna, G.; Scarano, A.; Donghia, R.; Castellana, F.; Cavalcanti, E.; Pesole, P.L.; Sommella, E.M.; Lippolis, A.; et al. Nutritional Regimes Enriched with Antioxidants as an Efficient Adjuvant for IBD Patients under Infliximab Administration, a Pilot Study. Antioxidants 2022, 11, 138. [Google Scholar] [CrossRef]
- Liu, Y.; Qiu, J.; Yue, Y.; Li, K.; Ren, G. Dietary black-grained wheat intake improves glycemic control and inflammatory profile in patients with type 2 diabetes, a randomized controlled trial. Ther. Clin. Risk Manag. 2018, 14, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Gamel, T.H.; Abdel-Aal, E.M.; Tucker, A.J.; Pare, S.M.; Faughnan, K.; O’Brien, C.D.; Dykun, A.; Rabalski, I.; Pickard, M.; Wright, A.J. Consumption of whole purple and regular wheat modestly improves metabolic markers in adults with elevated high-sensitivity C-reactive protein, a randomised, single-blind parallel-arm study. Br. J. Nutr. 2020, 24, 1179–1189. [Google Scholar] [CrossRef]
- Kaspar, K.L.; Park, J.S.; Brown, C.R.; Mathison, B.D.; Navarre, D.A.; Chew, B.P. Pigmented potato consumption alters oxidative stress and inflammatory damage in men. J. Nutr. 2011, 141, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinson, J.A.; Demkosky, C.A.; Navarre, D.A.; Smyda, M.A. High-antioxidant potatoes, Acute in vivo antioxidant source and hypotensive agent in humans after supplementation to hypertensive subjects. J. Agric. Food Chem. 2012, 60, 6749–6754. [Google Scholar] [CrossRef] [PubMed]
- Jokioja, J.; Linderborg, K.M.; Kortesniemi, M.; Nuora, A.; Heinonen, J.; Sainio, T.; Viitanen, M.; Kallio, H.; Yang, B. Anthocyaninrich extract from purple potatoes decreases postprandial glycemic response and affects inflammation markers in healthy men. Food Chem. 2020, 310, 125797. [Google Scholar] [CrossRef] [PubMed]
- Linderborg, K.M.; Salo, J.E.; Kalpio, M.; Vuorinen, A.L.; Kortesniemi, M.; Griinari, M.; Viitanen, M.; Yang, B.; Kallio, H. Comparison of the postprandial effects of purple-fleshed and yellow-fleshed potatoes in healthy males with chemical characterization of the potato meals. Int. J. Food Sci. Nutr. 2016, 67, 581–591. [Google Scholar] [CrossRef]
- Tsang, C.; Smail, N.F.; Almoosawi, S.; McDougall, G.J.M.; Al-Dujaili, E.A.S. Antioxidant Rich Potato Improves Arterial Stiffness in Healthy Adults. Plant Foods Hum. Nutr. 2018, 73, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurbuz, N.; Uluisik, S.; Frary, A.; Frary, A.; Doganlar, S. Health benefits and bioactive compounds of eggplant. Food Chem. 2018, 268, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.J.; Bae, M.S.; Jo, E.K.; Jo, Y.H.; Lee, S.C. Antioxidant activity of different parts of eggplant. J. Med. Plants Res. 2011, 5, 4610–4615. [Google Scholar]
- Casati, L.; Pagani, F.; Braga, P.C.; Scalzo, R.L.; Sibilia, V. Nasunin, a new player in the field of osteoblast protection against oxidative stress. J. Funct. Foods 2016, 23, 474–484. [Google Scholar] [CrossRef]
- Sensoy, I. A review on the relationship between food structure, processing, and bioavailability. Crit. Rev. Food Sci. Nutr. 2014, 54, 902–909. [Google Scholar] [CrossRef]
- Ichiyanagi, T.; Terahara, N.; Rahman, M.M.; Konishi, T. Gastrointestinal uptake of nasunin, acylated anthocyanin in eggplant. J. Agric. Food Chem. 2006, 54, 5306–5312. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, T.; Yin, Y.; Zhang, C.Y.; Zhang, Y.L. Dietary microRNA-A Novel Functional Component of Food. Adv. Nutr. 2019, 10, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Chen, W.L.; Kung, W.H.; Huang, H.D. Plant miRNAs found in human circulating system provide evidences of cross kingdom RNAi. BMC Genom. 2017, 18, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, Y.; Liu, Y.; Liu, H.; Wang, H.; Jin, W.; Zhang, Y.; Zhang, C.; Xu, D. Role of plant MicroRNA in cross-species regulatory networks of humans. BMC Syst. Biol. 2016, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalieri, D.; Rizzetto, L.; Tocci, N.; Rivero, D.; Asquini, E.; Si-Ammour, A.; Bonechi, E.; Ballerini, C.; Viola, R. Plant microRNAs as novel immunomodulatory agents. Sci. Rep. 2016, 6, 25761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, H.; Yuan, Y.; Etheridge, A.; Zhou, Y.; Huang, D.; Wilmes, P.; Galas, D. The complex exogenous RNA spectra in human plasma, An interface with human gut biota? PLoS ONE 2012, 7, e51009. [Google Scholar] [CrossRef]
- Pokoo, R.; Ren, S.; Wang, Q.; Motes, C.M.; Hernandez, T.D.; Ahmadi, S.; Monteros, M.J.; Zheng, Y.; Sunkar, M. Genotype- and tissue-specific miRNA profiles and their targets in three alfalfa (Medicago sativa L) genotypes. BMC Genom. 2018, 19, 913. [Google Scholar] [CrossRef] [PubMed]
- Crisp, P.A.; Hammond, R.; Zhou, P.; Vaillancourt, B.; Lipzen, A.; Daum, C.; Barry, K.; de Leon, N.; Buell, C.R.; Kaeppler, S.M.; et al. Variation and inheritance of small RNAs in maize inbreds and F1 hybrids. Plant Physiol. 2020, 182, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants, implications towards sustainable crop production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- Dad, H.A.; Gu, T.W.; Zhu, A.Q.; Huang, L.Q.; Peng, L.H. Plant exosome-like nanovesicles, emerging therapeutics and drug delivery nanoplatforms. Mol. Ther. 2021, 29, 13–31. [Google Scholar] [CrossRef]
- Bhome, R.; Del Vecchio, F.; Lee, G.H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs), Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef]
- Deng, Z.; Rong, Y.; Teng, Y.; Mu, J.; Zhuang, X.; Tseng, M.; Samykutty, A.; Zhang, L.; Yan, J.; Miller, D.; et al. Broccoli-derived nanoparticle inhibits mouse colitis by activating dendritic cell AMP-activated protein kinase. Mol. Ther. 2017, 25, 1641–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 586564. [Google Scholar] [CrossRef] [PubMed]



| Antioxidant Class | Prevalent Compounds | Main Modifications | Distribution into the Grain | Heritability Range for Total Content | Rice (O. sativa) | Wheat (T. aestivum; T. urum) | Maize (Z. mays) | Sorghum (S. bicolor L.) |
|---|---|---|---|---|---|---|---|---|
| Carotenoids | Lutein, Zeaxanthin, β-carotene, β-cryptoxanthin | endosperm, aleurone and germ (cereals) [66] | 0.7 in pigmented wheat and durum wheats [38] and in RILs of durum wheats [57] | negligible amount [67] | 4–12 μg/g [49] | blue: 0.18 μg/g; yellow: 0.13–60 μg/g [68,69] | 0.02 to 0.85 μg/g [70] | |
| Vitamin E | α and β-tocopherol, α and β-tocotrienols | aleurone, pericarp and germ (maize) | 19.36–63.29 μg/g [71] | 2.81–29.62 μg/g [70] | ||||
| Phenolic acids | ferulic acid, coumaric acid, syringic acid, vanillic acid, caffeic | mainly ester or ether linked to cell wall polimers | aleurone and germ (maize), bran, embryo and endosperm (rice) | 0.63 in tetraploid wheat collection [39] | 78.83–317.4 μg/g [72] | 550–1700 μg/g; 27–45 μg/g [37,39] | blue: 2.60 mg/g; yellow: 3.2 mg/g; white: 2.6 mg/g [73]; purple: 2.66–17.5 mg/g [74] | 1.0–29.6 mg/g [75] |
| Flavonoids | apigenin derivatives | mainly conjugated (glycosides) | pericarp (maize) | 886–2863 (μg rutin equivalent/g) [76] | 70–110 μg/g [36] | red: 27.53 mg/g [77] | * 0–23 mg/g [78] | |
| Anthocyanins | cyanidin 3-O-glucoside | only conjugated (glycosides) | aleurone, pericarp, cob; pericarp (maize) | 0.93 in pigmented wheat and durum wheats [38] | 87.54 mg (Cyanidin-3-glucoside equivalent/100 g rice grain) [72] | purple: 8–50 μg/g; red: 1–25 μg/g; blue: 80–170 μg/g [38] | blue: 0.66 mg/g; purple: 1.64 mg/g [79]; purple cob: 3.1–12.6 mg/g [79,80] | 1–3 mg/g [35] |
| Concentration of Phytochemicals (mg/g) Cereal and Solanaceae Crops Obtained by Breeding or Metabolic Engineering | ||||
|---|---|---|---|---|
| Antioxidant Class | Rice (O. sativa) | Tomato (S. lycopersicum) | Potato (S. tuberosum) | Eggplant (S. melongena) |
| Carotenoids | Golden rice: 5.06 μg/g, Golden Rice 2: up to 37 µg/g DW [89] | SRG1 mutants 5.1× lycopene [126]; “Sun Black” (Aft/Aft atv/atv, peel) 0.2 mg/g DW total carotenoid content [127]; “Bronze” (E8:MYB12, E8:Del/Ros, 35S:StSy) ∼0.55 mg/g DW total carotenoid content [132] | Golden potato (cv. Desiree) 3600-fold increase in beta carotene to 4.7 mg/100 g DW [146]; 4 mg/100 g DW zeaxanthin in S. tuberosum 4n [147]. From traces to 0.33 mg/100 g beta-carotene [148] | eggplant transgenic line EEF48:crtB 0.15 mg g−1 FW of β-carotene [149] |
| Phenolic acids | “Sun Black” (Aft/Aft atv/atv, peel) 0.6 mg/g DW of CGA, 8.6 mg/g DW total phenolic content [127]; “Yellow” E8:MYB12 15 mg g−1 DW CQAs equivalent to 22-fold higher levels, respectively [150] | 3.35-fold increases on average) [151] | ||
| Flavonoids (considered as total content) | “Sun Black” (Aft/Aft atv/atv, peel) 0.8 mg/g DW rutin [132], “Yellow” (E8:MYB12) 72 mg g−1 DW total flavonols equivalent to 65-fold higher levels [150]; “Bronze” (E8:MYB12, E8:Del/Ros1, 35S:StSy), “Indigo” (E8:MYB12, E8:Del/Ros1) ∼15 and 20 mg/g DW total flavonols, respectively [132] | Flavonols (4.50-fold increase on average) [151] | ||
| Flavonoids (Anthocyanins) | “Sun Black” (Aft/Aft atv/atv) 1.2 mg/g DW in fruit peel [127]; Del/Ros1 (E8:Del/Ros1) 14.7 mg g−1; “Indigo” (E8:MYB12, E8:Del/Ros1) ∼5–24 mg g−1 DW [131,132]; “Crimson” (E8:Del/Ros1, E8:AmDFR, f3′5′h) 5.3 ± 1.3 mg/g DW, “Magenta” (E8:Del/Ros1, E8:MYB12, E8:AmDFR, f3′5′h) 7.9 ± 2.3 mg/g DW total anthoyanins [131] | From 0.4 in wt to 3 ug/100 mg Petunidin (7×); from 0.04 in wt to up to 0.3 ug/100 g Pelargonidin (7×) [152] | ||
| Antioxidant-Rich Food | Health Effects in Animal Models | Refs. | |
|---|---|---|---|
| Cereals | Black rice | Improved hyperlipidemia and insulin resistance in rats on high-fructose diet | [219] |
| Reduced hyperlipidemia in rats fed HCD | [220] | ||
| Reduced dyslipidemia, induced optimal platelet function in rats fed HFD | [221] | ||
| Regulated cholesterol metabolism and improved dysbiosis of gut microbiota in mice fed HCD | [222] | ||
| Reduced ethanol-induced liver damage in rats | [227] | ||
| Attenuated liver injury and prevented fibrosis in CCl4-treated mice | [228] | ||
| Ameliorative effect in senescent mice induced by D-galactose | [229] | ||
| Black/red rice | Decreased atherosclerotic plaques, increased antioxidant status in rabbit fed HCD and in apoE-deficient mice | [223,224,225] | |
| Purple rice | Prevented neurodegeneration in a rat model of AD | [230] | |
| Black/purple wheat | Prevented obesity, hyperlipidemia, and insulin resistance in mice fed HFD | [226] | |
| Purple wheat | Preventive effect on cognitive functions in mouse models of AD and PD | [231] | |
| Blue corn | Reduced cardiac infarct size following ischemia-reperfusion in rats | [233] | |
| Purple corn | Long lasting cardioprotection against ischemia-reperfusion mediated by microbiota in mice | [234] | |
| Increased survival and reduced cardiac damages against Doxorubicin-induced cardiotoxic effects in mice | [235] | ||
| Reduced diabetes-associated renal fibrosis, angiogenesis, and mesangial and glomerulal inflammation in db/db mice | [236,237] | ||
| Prevented obesity and ameliorated hyperglycemia in mice fed HFD | [217,218,219] | ||
| Reduced obesity-associated inflammation by reprogramming of ATM in mice fed HFD | [77] | ||
| Reduced trigeminal inflammatory pain in rats | [239] | ||
| Delayed progression of muscular dystrophy reducing inflammation and oxidative stress in Sgca null mice | [240] | ||
| Solanaceae | Purple tomato | Delayed cancer development and increased life span in p53-/- mice | [120] |
| Reduced inflammation and induced antioxidant response in rat model of carrageenan-induced paw oedema | [244] | ||
| Bronze tomato | Reduced inflammation markers, modulated gut microbiota in Winnie mice | [132,246] | |
| Purple potato | Prevented gastrointestinal inflammation/cancers in pig fed HFD | [247] | |
| Reduced chronic intestinal inflammation in DSS-induced colitis in mice | [248,249] | ||
| Prevented obesity, hyperlipidemia, and insulin resistance in rats fed HFD | [252,253] | ||
| Attenuated hyperglycemia in STZ-induced diabetic rats. | [254] | ||
| Reduced obesity-associated oxidative damage in rats fed HCD | [256] | ||
| Suppressed GalN-induced hepatotoxicity via inhibition of lipid peroxidation and/or inflammation in rats | [255] | ||
| Purple/red potato | Reduced proliferation of the benzopyrene-induced stomach cancer in mice | [250] | |
| Red potato | Reduced MNU1-induced breast carcinogenesis in rats | [251] | |
| Inhibited hepatic lipid peroxidation in rats | [257] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bassolino, L.; Petroni, K.; Polito, A.; Marinelli, A.; Azzini, E.; Ferrari, M.; Ficco, D.B.M.; Mazzucotelli, E.; Tondelli, A.; Fricano, A.; et al. Does Plant Breeding for Antioxidant-Rich Foods Have an Impact on Human Health? Antioxidants 2022, 11, 794. https://doi.org/10.3390/antiox11040794
Bassolino L, Petroni K, Polito A, Marinelli A, Azzini E, Ferrari M, Ficco DBM, Mazzucotelli E, Tondelli A, Fricano A, et al. Does Plant Breeding for Antioxidant-Rich Foods Have an Impact on Human Health? Antioxidants. 2022; 11(4):794. https://doi.org/10.3390/antiox11040794
Chicago/Turabian StyleBassolino, Laura, Katia Petroni, Angela Polito, Alessandra Marinelli, Elena Azzini, Marika Ferrari, Donatella B. M. Ficco, Elisabetta Mazzucotelli, Alessandro Tondelli, Agostino Fricano, and et al. 2022. "Does Plant Breeding for Antioxidant-Rich Foods Have an Impact on Human Health?" Antioxidants 11, no. 4: 794. https://doi.org/10.3390/antiox11040794