
Dietary Supplementation of Silybum marianum Seeds Improved Growth Performance and Upregulated Associated Gene Expression of Muscovy Ducklings (Cairina moschata)
 , , , , , ,
, , , , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Ethics for the Used Animals
2.2. Experimental Design and Diets Preparation
2.3. Bird’s Housing and Management
2.4. Growth Performance Parameters
2.5. Biochemical Parameters
2.6. Gene Expression
2.6.1. Total RNA Extraction and Complementary Deoxyribonucleic Acid (cDNA) Synthesis
2.6.2. Differential Expression Analysis of GH, CAT, Cu/Zn-SOD (SOD1), and Mn-SOD (SOD2) Genes by Quantitative Real-Time PCR (qRT-PCR)
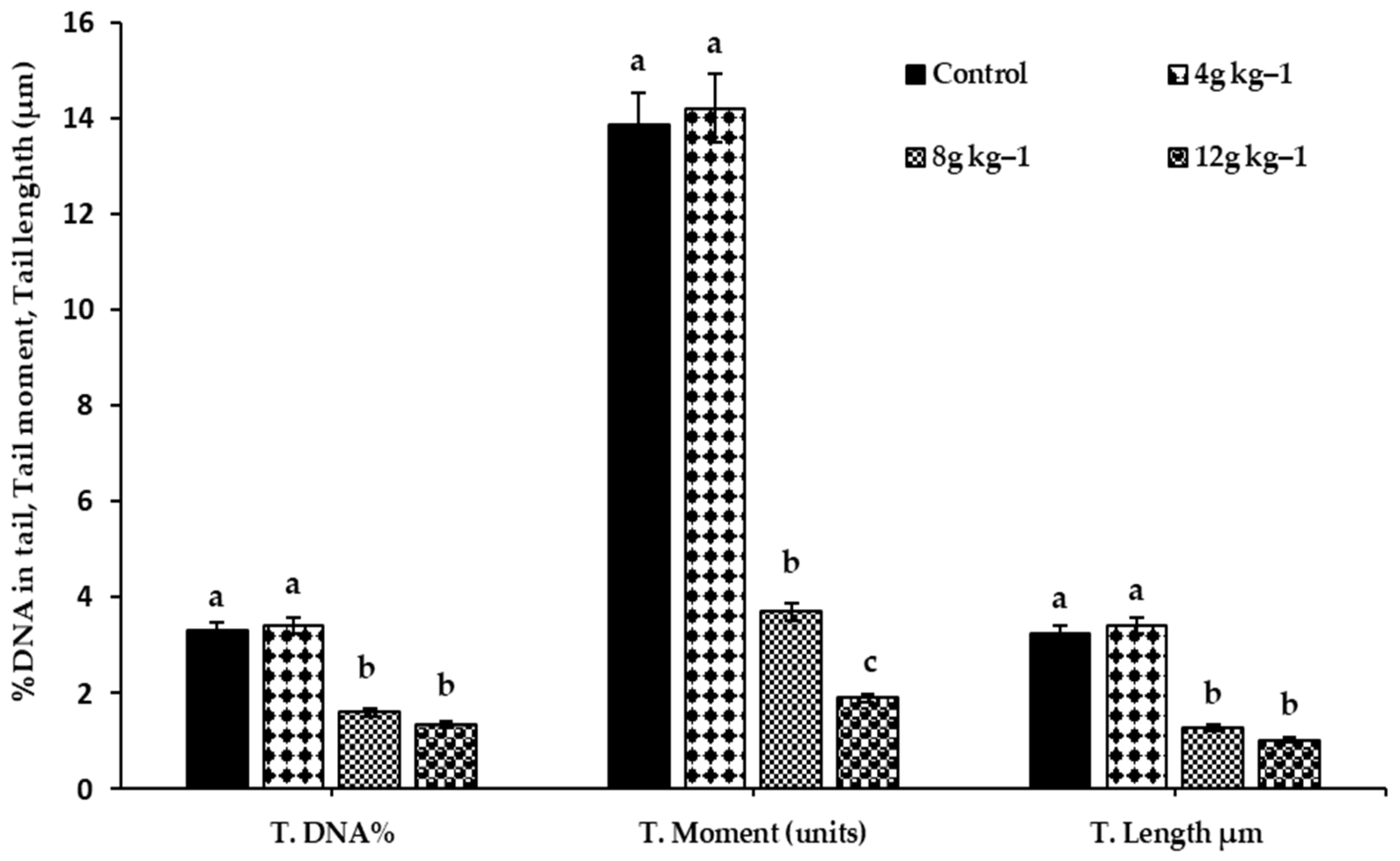
2.7. DNA Comet Assay
2.8. Statistical Analysis
3. Results
3.1. Growth Performance Parameters
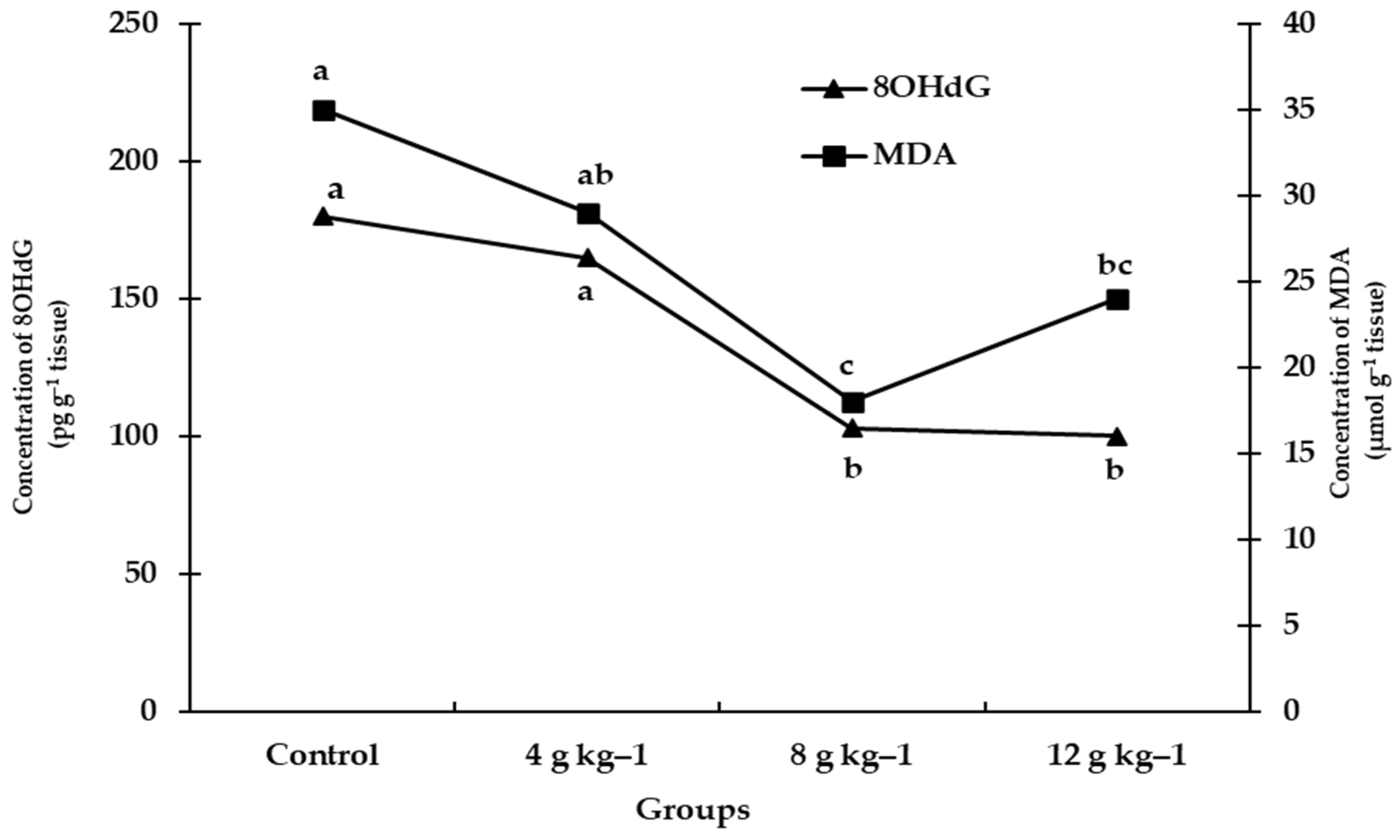
3.2. Biochemical Parameters
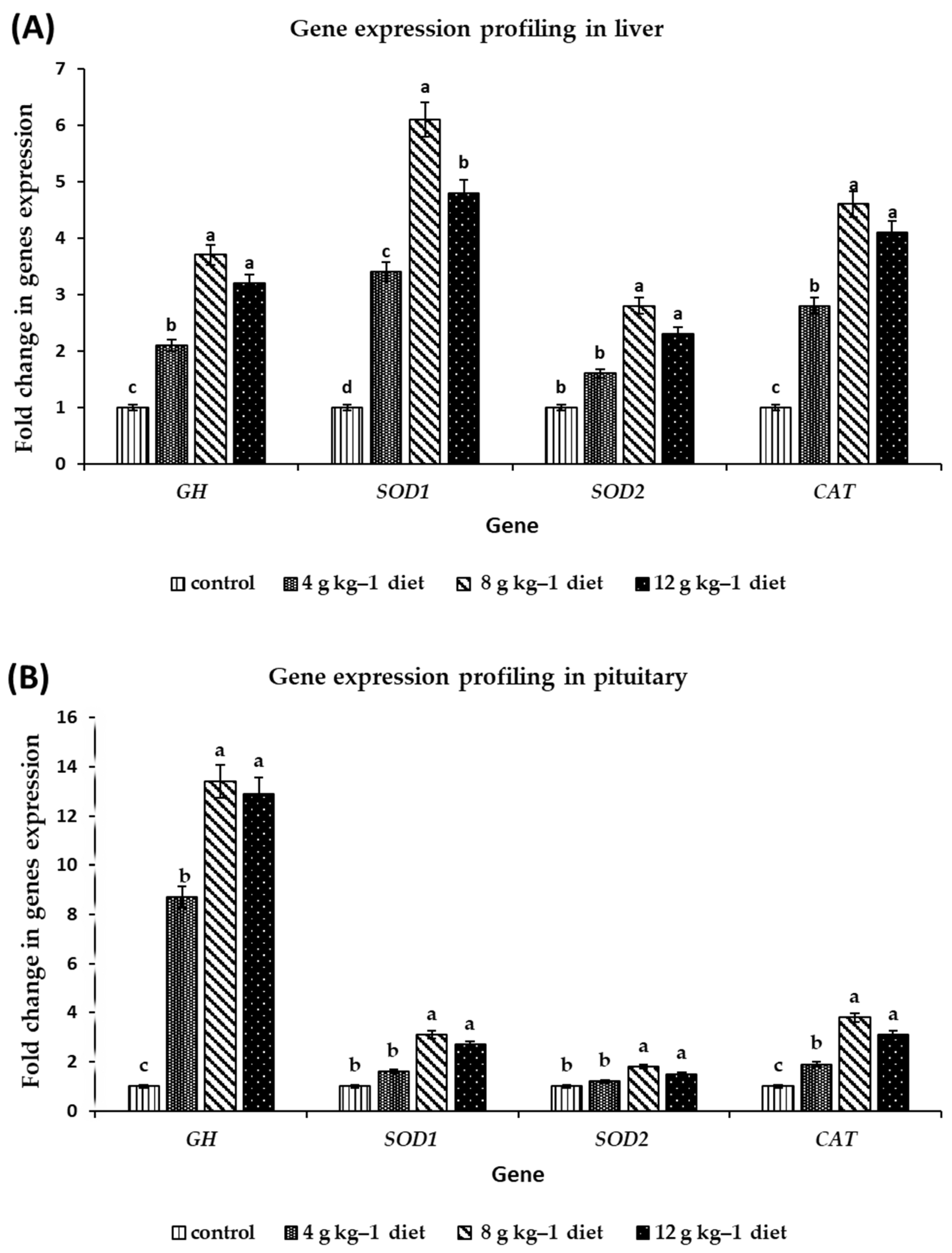
3.3. Gene Expression Profiling
3.4. DNA Comet Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Surai, P.F. Antioxidant Systems in Poultry Biology: Superoxide Dismutase. J. Anim. Res. Nutr. 2016, 1, 8. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant Systems in Poultry Biology: Heat Shock Proteins. J. Sci. 2015, 5, 1188–1222. [Google Scholar]
- Surai, P.F. Natural Antioxidants in Poultry Nutrition: New Developments Natural Antioxidants in Feed Ingredients the Need for Antioxidant Defence. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; World Poultry Science Association: Apeldoorn, The Netherlands, 2007; pp. 26–30. [Google Scholar]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafał, P.; Albrakati, A.; et al. Luteolin Alleviates Aflatoxinb1-induced Apoptosis and Oxidative Stress in the Liver of Mice through Activation of Nrf2 Signaling Pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and Antioxidants in Disease: Oxidative Stress in Farm Animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Shaukat, A.; Rajput, I.R.; Kamboh, A.A.; Iqbal, Z.; Saeed, M.; Akhtar, R.W.; Shah, S.A.H.; Raza, M.A.; El Askary, A.; et al. Ginsenoside Rb1 Prevents Deoxynivalenol-Induced Immune Injury via Alleviating Oxidative Stress and Apoptosis in Mice. Ecotoxicol. Environ. Saf. 2021, 220, 112333. [Google Scholar] [CrossRef]
- Shatskikh, E.V.; Latypova, E.; Fisinin, V.I.; Denev, S.; Surai, P. Molecular Mechanisms and New Strategies to Fight Stresses in Egg-Producing Birds. Agric. Sci. Technol. 2015, 7, 3–10. [Google Scholar]
- Fernández, C.; San Miguel, E.; Fernández-Briera, A. Superoxide Dismutase and Catalase: Tissue Activities and Relation with Age in the Long-Lived Species Margaritifera margaritifera. Biol. Res. 2009, 42, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Zeng, T.; Li, J.J.; Wang, D.Q.; Li, G.Q.; Wang, G.L.; Lu, L.Z. Effects of Heat Stress on Antioxidant Defense System, Inflammatory Injury, and Heat Shock Proteins of Muscovy and Pekin Ducks: Evidence for Differential Thermal Sensitivities. Cell Stress Chaperones 2014, 19, 895–901. [Google Scholar] [CrossRef]
- Comelli, M.C.; Mengs, U.; Schneider, C.; Prosdocimi, M. Toward the Definition of the Mechanism of Action of Silymarin: Activities Related to Cellular Protection from Toxic Damage Induced by Chemotherapy. Integr. Cancer Ther. 2007, 6, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yan, M.J.; Lian, S.Y.; Liu, X.T.; Li, A. GH Gene Polymorphisms and Expression Associated with Egg Laying in Muscovy Ducks (Cairina moschata). Hereditas 2014, 151, 14–19. [Google Scholar] [CrossRef]
- Kosina, P.; Dokoupilová, A.; Janda, K.; Sládková, K.; Silberová, P.; Pivodová, V.; Ulrichová, J. Effect of Silybum marianum Fruit Constituents on the Health Status of Rabbits in Repeated 42-day Fattening Experiment. Anim. Feed Sci. Technol. 2017, 223, 128–140. [Google Scholar] [CrossRef]
- Schiavone, A.; Righi, F.; Quarantelli, A.; Bruni, R.; Serventi, P.; Fusari, A. Use of Silybum marianum Fruit Extract in Broiler Chicken Nutrition: Influence on Performance and Meat Quality. J. Anim. Physiol. Anim. Nutr. 2007, 91, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Hassaan, M.S.M.; Moustafa, M.M.A.; Refaat, M.H. The Influence Of Synbiotic On Growth and Expression Of GH, GHR1 and IGF-I Genes In Oreochromis niloticus L. Fingerlings. J. Fish. Aquac. 2015, 6, 176–182. [Google Scholar]
- NRC (National Research Council). Nutrient Requirements of Poultry; National Academies Press: Washington, DC, USA, 1994.
- Wianowska, D.; Wiśniewski, M. Simplified Procedure of Silymarin Extraction from Silybum marianum L. Gaertner. J. Chromatogr. Sci. 2015, 53, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Sampedro, M.A.; Fernández-Tárrago, J.; Corchete, P. Enhanced Silymarin Accumulation Is Related to Calcium Deprivation in Cell Suspension Cultures of Silybum marianum (L.) Gaertn. J. Plant Physiol. 2005, 162, 1177–1182. [Google Scholar] [CrossRef]
- Broody, S. Biomergetic and Growth; Reinhold Publication Corp.: New York, NY, USA, 1949. [Google Scholar]
- Kar, J.; Barman, T.R.; Sen, A.; Nath, S.K. Management, Growth Performance and Cost Effectiveness of Japanese Quail in Khaza Quail Farm and Hatchery Limited at Chittagong in Bangladesh. Glob. J. Med. Res. 2017, 17, 1–8. [Google Scholar]
- Reitman, S.; Frankel, S. A Colorimetric Method for the Determination of Serum Glutamic Oxalacetic and Glutamic Pyruvic Transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Tietz, N.W.; Rinker, A.D.; Morrison, S.R. When Is a Serum Iron Really a Serum Iron? The Status of Serum Iron Measurements. Clin. Chem. 1994, 40, 546–551. [Google Scholar] [CrossRef]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin Standards and the Measurement of Serum Albumin with Bromcresol Green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef]
- Karatepe, M. Simultaneous Determination of Ascorbic Acid and Free Malondialdehyde in Human Serum by HPLC-UV. LC-GC N. Am. 2004, 22, S104. [Google Scholar]
- Lodovici, M.; Casalini, C.; Briani, C.; Dolara, P. Oxidative Liver DNA Damage in Rats Treated with Pesticide Mixtures. Toxicology 1997, 117, 55–60. [Google Scholar] [CrossRef]
- Jayatilleke, E.; Shaw, S. A High-Performance Liquid Chromatographic Assay for Reduced and Oxidized Glutathione in Biological Samples. Anal. Biochem. 1993, 214, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Papadoyannis, I.N.; Samanidou, V.F.; Nitsos, C.C. Simultaneous Determination of Nitrite and Nitrate in Drinking Water and Human Serum by High Performance Anion-Exchange Chromatography and UV Detection. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 2023–2041. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. [13] Catalase In Vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Teerlink, T.; Hennekes, M.; Bussemaker, J.; Groeneveld, J. Simultaneous Determination of Creatine Compounds and Adenine Nucleotides in Myocardial Tissue by High-Performance Liquid Chromatography. Anal. Biochem. 1993, 214, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Pagel, M.; Barbin, V.; Blanc, P.; Ohnenstetter, D. Cathodoluminescence in Geosciences: An Introduction. In Cathodoluminescence in Geosciences; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–21. [Google Scholar]
- Lucia, M.; Andre, J.M.; Gonzalez, P.; Baudrimont, M.; Gontier, K.; Maury-Brachet, R.; Davail, S. Impact of Cadmium on Aquatic Bird Cairina moschata. BioMetals 2009, 22, 843–853. [Google Scholar] [CrossRef]
- Wu, A.R.; Neff, N.F.; Kalisky, T.; Dalerba, P.; Treutlein, B.; Rothenberg, M.E.; Mburu, F.M.; Mantalas, G.L.; Sim, S.; Clarke, M.F.; et al. Quantitative Assessment of Single-Cell RNA-Sequencing Methods. Nat. Methods 2014, 11, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.R.; Zhu, J.Q.; Ye, T.; Wang, C.L.; Zhu, Y.F.; Dahms, H.U.; Jin, F.; Yang, W.X. Improvement of Single-Cell Gel Electrophoresis (SCGE) Alkaline Comet Assay. Aquat. Biol. 2013, 18, 293–295. [Google Scholar] [CrossRef] [Green Version]
- SAS. User’s Guide: Basics; Institute Statistical Analysis System: Cary, NC, USA, 2000; SMN. 2014. Unidad Del Servicio Meteorológico Nacional, CNA, Coyoacán, Mexico. [Google Scholar]
- Saller, R.; Melzer, J.; Reichling, J.; Brignoli, R.; Meier, R. An Updated Systematic Review of the Pharmacology of Silymarin. Complement. Med. Res. 2007, 14, 70–80. [Google Scholar] [CrossRef]
- Yi, D.; Gu, L.; Ding, B.; Li, M.; Hou, Y.; Wang, L.; Gong, J. Effects of Dietary Silymarin Supplementation on Growth Performance and Oxidative Status in Carassius Auratus Gibelio. J. Anim. Vet. Adv. 2012, 11, 3399–3404. [Google Scholar]
- Alhidary, I.A.; Rehman, Z.; Khan, R.U.; Tahir, M. Anti-Aflatoxin Activities of Milk Thistle (Silybum marianum) in Broiler. World’s Poult. Sci. J. 2017, 73, 559–566. [Google Scholar] [CrossRef]
- Camacho-Luis, A.; Mendoza-Pérez, J.A. La Naturaleza Efímera de los Radicales Libres. Antioxid. Enferme-Dades Crónico Degener. 2009, 9, 27–76. [Google Scholar]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Wen, X.; Li, S.; Zhu, D.; Li, Y. Effects of Dietary Lipid Levels on Growth, Feed Utilization, Body Composition and Antioxidants of Juvenile Mud Crab Scylla paramamosain (Estampador). Aquaculture 2015, 435, 200–206. [Google Scholar] [CrossRef]
- Li, L.; Zeng, J.; Gao, Y.; He, D. Targeting Silibinin in the Antiproliferative Pathway. Expert Opin. Investig. Drugs 2010, 19, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Breschi, M.C.; Martinotti, E.; Apostoliti, F.; Nieri, P. Protective Effect of Silymarin in Antigen Challenge- and Histamine-Induced Bronchoconstriction in In Vivo Guinea-Pigs. Eur. J. Pharmacol. 2002, 437, 91–95. [Google Scholar] [CrossRef]
- Rolo, A.P.; Oliveira, P.J.; Moreno, A.J.M.; Palmeira, C.M. Protection against Post-Ischemic Mitochondrial Injury in Rat Liver by Silymarin or TUDC. Hepatol. Res. 2003, 26, 217–224. [Google Scholar] [CrossRef]
- Zhou, L.; O’Rourke, B. Cardiac Mitochondrial Network Excitability: Insights from Computational Analysis. Am. J. Physiol.-Heart Circ. Physiol. 2012, 302, 2178–2189. [Google Scholar] [CrossRef] [Green Version]
- Ligeret, H.; Brault, A.; Vallerand, D.; Haddad, Y.; Haddad, P.S. Antioxidant and Mitochondrial Protective Effects of Silibinin in Cold Preservation-Warm Reperfusion Liver Injury. J. Ethnopharmacol. 2007, 115, 507–514. [Google Scholar] [CrossRef]
- Serviddio, G.; Bellanti, F.; Giudetti, A.M.; Gnoni, G.V.; Petrella, A.; Tamborra, R.; Romano, A.D.; Rollo, T.; Vendemiale, G.; Altomare, E. A Silybin-Phospholipid Complex Prevents Mitochondrial Dysfunction in a Rodent Model of Nonalcoholic Steatohepatitis. J. Pharmacol. Exp. Ther. 2010, 332, 922–932. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.J.; Liao, J.F.; Lin, D.Y.; Huang, M.C.; Liou, D.Y.; Yang, H.C.; Lee, H.J.; Chen, Y.T.; Chi, C.W.; Huang, W.C.; et al. Silymarin Protects Spinal Cord and Cortical Cells against Oxidative Stress and Lipopolysaccharide Stimulation. Neurochem. Int. 2010, 57, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.A.; Nguyen, J.C.D.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Karimi, G.; Vahabzadeh, M.; Lari, P.; Rashedinia, M.; Moshiri, M. Silymarin, a Promising Pharmacological Agent for Treatment of Diseases. Iran. J. Basic Med. Sci. 2011, 14, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Baluchnejadmojarad, T.; Roghani, M.; Khastehkhodaie, Z. Chronic Treatment of Silymarin Improves Hyperalgesia and Motor Nerve Conduction Velocity in Diabetic Neuropathic Rat. Phyther. Res. 2010, 24, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Best, B.P. Nuclear Dna Damage as a Direct Cause of Aging. Rejuvenat. Res. 2009, 12, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Osuchowski, M.F.; Johnson, V.J.; He, Q.; Sharma, R.P. Alterations in Regional Brain Neurotransmitters by Silymarin, a Natural Antioxidant Flavonoid Mixture, in BALB/c Mice. Pharm. Biol. 2004, 42, 384–389. [Google Scholar] [CrossRef]
- Bansal, N.; Gill, R.; Gupta, G. Das Silymarin: A Flavolignan with Antidepressant Activity. J. Pharm. Innov. 2013, 5, 93–98. [Google Scholar]
- Harvey, S. Growth Hormone and Growth? Gen. Comp. Endocrinol. 2012, 190, 3–9. [Google Scholar] [CrossRef]
- Ji, W.; Sun, G.; Duan, X.; Dong, B.; Bian, Y. Cloning of the Growth Hormone Receptor and Its Muscle-Specific MRNA Expression in Black Muscovy Duck (Cairina moschata). Br. Poult. Sci. 2016, 57, 211–218. [Google Scholar] [CrossRef]
- Harvey, S.; Johnson, C.D.M.; Sanders, E.J. Extra-Pituitary Growth Hormone in Peripheral Tissues of Early Chick Embryos. J. Endocrinol. 2000, 166, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, G.; Tiwari, M.N.; Prakash, O.; Jyoti, A.; Shanker, R.; Singh, M.P. Involvement of Multiple Molecular Events in Pyrogallol-Induced Hepatotoxicity and Silymarin-Mediated Protection: Evidence from Gene Expression Profiles. Food Chem. Toxicol. 2010, 48, 1660–1670. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, S.; Lasagna, E.; Sarti, F.M.; Marelli, S.P.; Cozzi, M.C.; Bernardini, G.; Terova, G. Expression Profile of Six Stress-Related Genes and Productive Performances of Fast and Slow Growing Broiler Strains Reared under Heat Stress Conditions. Meta Gene 2015, 6, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Lucia, M.; Andre, J.M.; Gonzalez, P.; Baudrimont, M.; Bernadet, M.D.; Gontier, K.; Maury-Brachet, R.; Guy, G.; Davail, S. Effects of Dietary Cadmium Contamination on Bird Anas Platyrhynchos-Comparison with Species Cairina moschata. Ecotoxicol. Environ. Saf. 2010, 73, 2010–2016. [Google Scholar] [CrossRef]
- El Mesallamy, H.O.; Metwally, N.S.; Soliman, M.S.; Ahmed, K.A.; Abdel Moaty, M.M. The Chemopreventive Effect of Ginkgo Biloba and Silybum marianum Extracts on Hepatocarcinogenesis in Rats. Cancer Cell Int. 2011, 11, 38. [Google Scholar] [CrossRef] [Green Version]
- Saravanan, R.; Pugalendi, K.V. Assessment of the Pharmacological Effect of Silymarin on Ethanol-Induced DNA Damage by Single-Cell Gel Electrophoresis. Indian J. Pharmacol. 2005, 37, 261–262. [Google Scholar] [CrossRef] [Green Version]
- Al-Rasheed, N.M.; Al-Rasheed, N.M.; Faddah, L.M.; Mohamed, A.M.; Mohammad, R.A.; Al-Amin, M. Potential Impact of Silymarin in Combination with Chlorogenic Acid and/or Melatonin in Combating Cardiomyopathy Induced by Carbon Tetrachloride. Saudi J. Biol. Sci. 2014, 21, 265–274. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients % | Control | 4 g kg−1 | 8 g kg−1 | 12 g kg−1 |
|---|---|---|---|---|
| Yellow corn | 61.00 | 60.60 | 60.2 | 59.8 |
| Soybean meal (44% CP) | 35.50 | 35.50 | 35.50 | 35.50 |
| Dicalcium phosphate | 1.70 | 1.70 | 1.70 | 1.70 |
| Calcium carbonate | 1.10 | 1.10 | 1.10 | 1.10 |
| Sodium chloride | 0.30 | 0.30 | 0.30 | 0.30 |
| Vit. and Mn. premix * | 0.30 | 0.30 | 0.30 | 0.30 |
| DL-Methionine 99% | 0.10 | 0.10 | 0.10 | 0.10 |
| S. marianum dry seeds | 0.00 | 0.40 | 0.80 | 1.20 |
| Chemical analysis % ** | ||||
| Metabolizable Energy (Kcal kg−1) | 2837 | 2840 | 2844 | 2850 |
| Crude protein | 20.26 | 20.30 | 20.37 | 20.43 |
| Crude fat | 2.67 | 2.74 | 2.84 | 2.93 |
| Crude fiber | 2.73 | 2.74 | 2.75 | 2.77 |
| Calcium | 0.93 | 0.93 | 0.93 | 0.93 |
| Available P | 0.43 | 0.43 | 0.43 | 0.43 |
| Methionine | 0.46 | 0.46 | 0.46 | 0.46 |
| Methionine + Cystine | 0.83 | 0.83 | 0.83 | 0.83 |
| Lysine | 1.21 | 1.21 | 1.21 | 1.21 |
| Flavonolignans | Differential Silymarin Profile mg g−1 Dried Powder | ||||||
| Isosilybin A + B | Silybin B | Silybin A | Silydianin | Silychristin | Taxifolin | Total Silymarin | |
| 0.84 | 2.5 | 1.7 | 1 | 2.56 | 2.87 | 12.25 | |
| Diets | The Expected Content of Flavonolignans in the Experimental Diets (mg kg−1) | ||||||
| Iso Sb A + B | Silybin B | Silybin A | Silydianin | Silychristin | Taxifolin | Total Silymarin | |
| 4 g kg−1 | 3.36 | 10.24 | 6.80 | 4.00 | 10.00 | 11.48 | 49 |
| 8 g kg−1 | 6.72 | 20.48 | 13.60 | 8.00 | 20.00 | 22.96 | 98 |
| 12 g kg−1 | 10.08 | 30.72 | 20.40 | 12.00 | 30.00 | 34.44 | 147 |
| Parameters | Method | Company | Reference |
|---|---|---|---|
| Serum AST (U L−1) | Enzymatic-colorimetric | Biodiagnostic (Giza, Egypt) | [20] |
| Serum ALT (U L−1) | Enzymatic-colorimetric | Biodiagnostic (Egypt) | [20] |
| Serum Total protein (g dL−1) | Enzymatic-colorimetric | Biodiagnostic (Egypt) | [21] |
| Serum Albumin (d dL−1) | Enzymatic-colorimetric | Biodiagnostic (Egypt) | [22] |
| Serum Globulin (g dL−1) | Calculated | =TP-Alb | |
| Liver MDA (nmol g−1 tissue) | HPLC | Standard of 1, 1, 3, 3 tetraethoxypropane (Sigma, St. Louis, MO, USA) | [23] |
| Liver 8OHdG (pg g−1 tissue) | HPLC | Standard of 8-hydroxy-2-deoxyguanosine (Sigma). | [24] |
| Liver GSH & GSSG (μmol g−1 tissue) | HPLC | Standard of GSH & GSSG (Sigma) | [25] |
| Liver NO (μmol g−1 tissue) | HPLC | Standard of nitrite and nitrate (Sigma). | [26] |
| Liver SOD (U g−1 tissue) | Colorimetric | Against pyrogallol (Sigma) | [27] |
| Liver Catalase (mmol min−1 g−1 tissue) | Colorimetric | Against H2O2 | [28] |
| Liver ATP, ADP, and AMP (μg g−1 tissue) | HPLC | Sigma | [29] |
| Brain 5HT (μg g−1 tissue) | HPLC | Sigma | [30] |
| Gene | Oligonucleotide Name and Sequence of Qrt-PCR Primers | Reference |
|---|---|---|
| Superoxide dismutase (Cu/Zn) (SOD1) | SOD1 F 5-GCGCACCATGGTGGTCCATG-3 SOD1 R 5-GTCTTCACCAGTTTAACTGATACTCA-3 | [31] |
| Mitochondrial superoxide dismutase (Mn), (SOD2) | SOD2 F 5-CGCCTATGTCAACAACCTCA-3 SOD2 R 5-AGGCGAAAGATTTGTCCAGA-3 | EU598450.1 |
| Catalase (CAT) | CAT F 5-GAGCAGGTGCTTTTGGCTAT-3 CAT R 5-TTTCCCACAAGATCCCAGTT-3 | EU598451.1 |
| Growth hormone (GH) | GH F 5-TGGGGTTGTTTAGCTTGGAG-3 GH R 5-TAAACCTTCCCTGGCACAAC-3 | AB158762.1 |
| β-Actin internal reference gene | β-Actin F 5 GGAAGTTACTCGCCTCTG-3 β-Actin R 5-CGCTCGCTGAACAAATC-3 | [32] |
| Parameter | Groups | |||
|---|---|---|---|---|
| Control | 4 g kg−1 | 8 g kg−1 | 12 g kg−1 | |
| Initial body weight at hatch (g) | 53.5 ± 1.5 | 54.1 ± 1.6 | 54.4 ± 1.7 | 55.1 ± 1.6 |
| Body weight at 4 weeks (g) | 1166 d ± 37.7 | 1310 c ± 39.8 | 1502 a ± 47.6 | 1405 b ± 43.2 |
| Body weight gain at 4 weeks (g) | 1112 d ± 32.6 | 1256 c ± 40.0 | 1448 a ± 43.0 | 1349 b ± 40.0 |
| % Growth rate at 4 weeks | 182 c ± 5.3 | 184 b ± 5.3 | 186 a ± 5.0 | 184 b ± 6.0 |
| Feed consumption (g/bird) | 2742 ± 82.7 | 2748 ± 83.9 | 2745 ± 87.4 | 2739 ± 81.7 |
| Feed conversion (g/bird) | 2.47 a ± 0.07 | 2.19 b ± 0.07 | 1.90 d ± 0.06 | 2.03 c ± 0.06 |
| Percentage of Mortality (%) | 10.0 a ± 0.33 | 3.33 b ± 0.10 | 1.67 c ± 0.05 | 1.67 c ± 0.04 |
| Parameter | Groups | |||
|---|---|---|---|---|
| Control | 4 g kg−1 | 8 g kg−1 | 12 g kg−1 | |
| AST (U/L) | 31.1 a ± 2.64 | 27.2 ab ± 3.51 | 28.0 ab ± 1.00 | 21.6 b ± 2.33 |
| ALT (U/L) | 53.6 a ± 1.85 | 50.3 ab ± 1.76 | 46.0 b ± 2.08 | 40.0 c ± 1.01 |
| TP (g/dL) | 6.37 c ± 0.05 | 6.24 c ± 0.03 | 7.22 b ± 0.22 | 8.04 a ± 0.08 |
| Alb (g/dL) | 3.87 c ± 0.03 | 3.81 c ± 0.04 | 4.24 b ± 0.07 | 4.67 a ± 0.01 |
| Glob (g/dL) | 2.80 bc ± 0.08 | 2.50 c ± 0.06 | 2.98 ab ± 0.23 | 3.36 a ± 0.07 |
| A/G | 1.38 b ± 0.05 | 1.52 a ± 0.06 | 1.42 ab ± 0.12 | 1.39 b ± 0.03 |
| Parameter | Groups | |||
|---|---|---|---|---|
| Control | 4 g kg−1 | 8 g kg−1 | 12 g kg−1 | |
| GSH (nmol g−1 tissue) | 56.2 b ± 5.58 | 52.70 b ± 5.58 | 54.0 b ± 5.58 | 94.2 a ± 5.58 |
| GSSG (nmol g−1 tissue) | 1.61 a ± 0.14 | 1.54 a ± 0.14 | 1.28 ab ± 0.14 | 1.02 b ± 0.14 |
| NO (μmol g−1 tissue) | 22.32 a ± 2.61 | 21.57 ab ± 2.61 | 20.42 b ± 2.61 | 17.39 c ± 2.61 |
| SOD (U g−1 tissue) | 12.0 c ± 0.37 | 13.5 c ± 0.39 | 22.6 a ± 0.65 | 19.6 b ± 0.58 |
| CAT (U g−1 tissue) | 3.62 b ± 0.10 | 4.09 b ± 0.13 | 6.98 a ± 0.20 | 6.19 a ± 0.19 |
| Parameter | Groups | |||
|---|---|---|---|---|
| Control | 4 g kg−1 | 8 g kg−1 | 12 g kg−1 | |
| ATP (μg g−1 tissue) | 55.7 b ± 6.34 | 56.5 b ± 6.34 | 57.7 b ± 6.34 | 76.9 a ± 6.34 |
| ADP (μg g−1 tissue) | 15.3 ab ± 1.16 | 16.1 a ± 1.16 | 14.8 ab ± 1.16 | 14.3 b ± 1.16 |
| AMP (μg g−1 tissue) | 10.5 ab ± 0.50 | 11.3 a ± 0.56 | 9.3 b ± 0.40 | 9.0 b ± 0.26 |
| 5HT (μg g−1 tissue) | 1.80 b ± 0.19 | 2.15 ab ± 0.19 | 2.09 ab ± 0.19 | 2.67 a ± 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Garhy, O.; Soudy, F.A.; Alharbi, Y.M.; Alshanbari, F.A.; Almujaydil, M.S.; Alhomaid, R.M.; Ahmed-Farid, O.A.; Mohamed, S.A.; El-Garhy, H.A.S.; Barakat, H.; et al. Dietary Supplementation of Silybum marianum Seeds Improved Growth Performance and Upregulated Associated Gene Expression of Muscovy Ducklings (Cairina moschata). Antioxidants 2022, 11, 2300. https://doi.org/10.3390/antiox11112300
El-Garhy O, Soudy FA, Alharbi YM, Alshanbari FA, Almujaydil MS, Alhomaid RM, Ahmed-Farid OA, Mohamed SA, El-Garhy HAS, Barakat H, et al. Dietary Supplementation of Silybum marianum Seeds Improved Growth Performance and Upregulated Associated Gene Expression of Muscovy Ducklings (Cairina moschata). Antioxidants. 2022; 11(11):2300. https://doi.org/10.3390/antiox11112300
Chicago/Turabian StyleEl-Garhy, Osama, Fathia A. Soudy, Yousef M. Alharbi, Fahad A. Alshanbari, Mona S. Almujaydil, Raghad M. Alhomaid, Omar A. Ahmed-Farid, Shereen A. Mohamed, Hoda A. S. El-Garhy, Hassan Barakat, and et al. 2022. "Dietary Supplementation of Silybum marianum Seeds Improved Growth Performance and Upregulated Associated Gene Expression of Muscovy Ducklings (Cairina moschata)" Antioxidants 11, no. 11: 2300. https://doi.org/10.3390/antiox11112300