
High Redox Status as the Basis for Heavy Metal Tolerance of Sesuvium portulacastrum L. Inhabiting Contaminated Soil in Jeddah, Saudi Arabia

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area Description
2.2. Sites Localization
2.3. Field Surveys
2.4. Floristic Diversity Analysis
2.5. Collection of S. portulacastrum
2.6. Biological Indices
2.7. Heavy Metal and Mineral Content in Soil and Plant Organs
2.8. Quantification of Organic Acids
2.9. Photosynthesis and Photorespiration Related Parameters
2.10. Quantification of Oxidative Damage Markers
2.11. Quantification of Antioxidant Parameters
2.12. Quantification of Detoxification Related Parameters
2.13. Statistical Analysis
3. Results
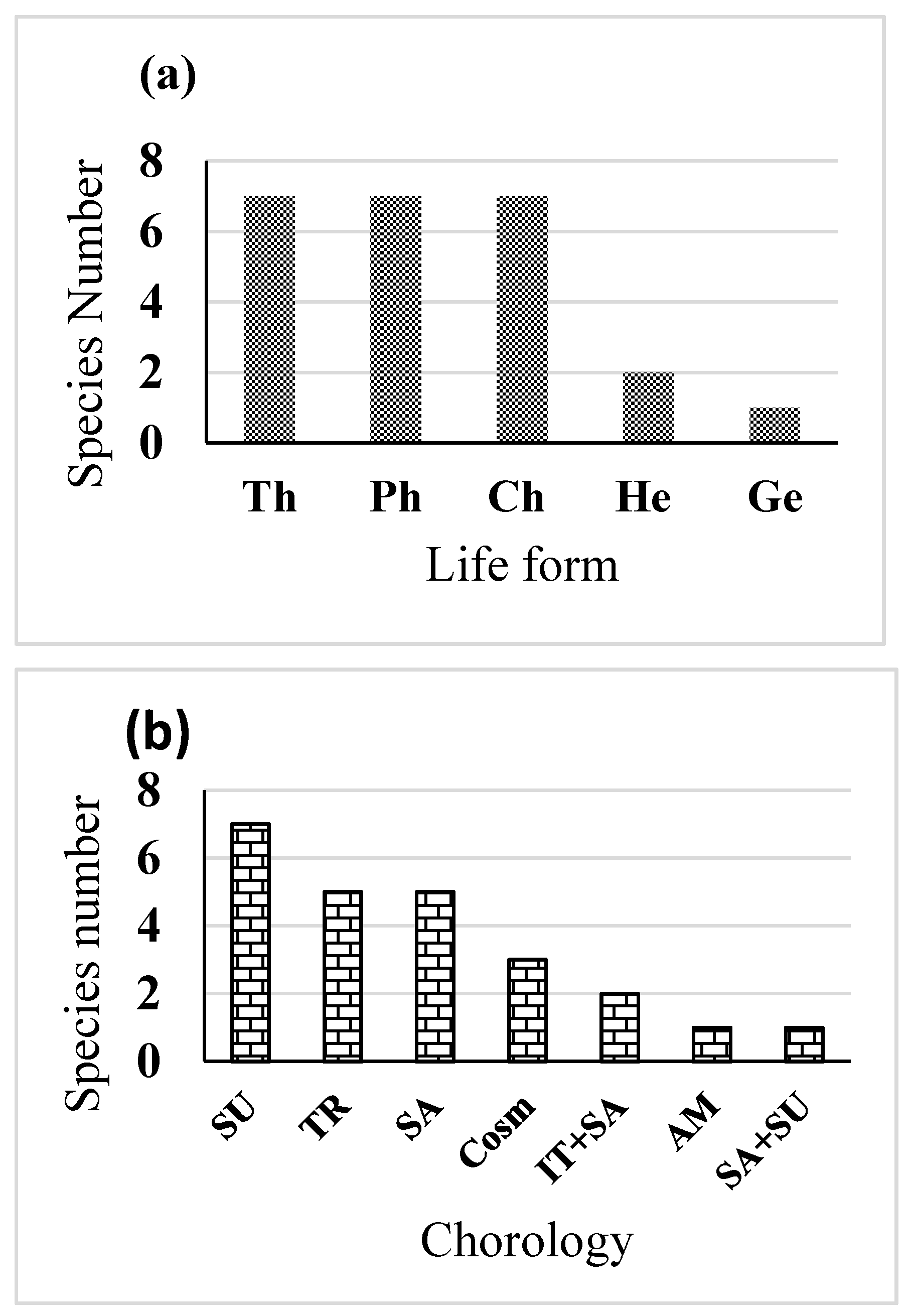
3.1. Effect of Sewage Pollution on Floristic Composition
3.2. Heavy Metals Level in Contaminated Soils and Their Uptake by S. portulacastrum
3.3. Growth Responses to Soil Contamination with Heavy Metals
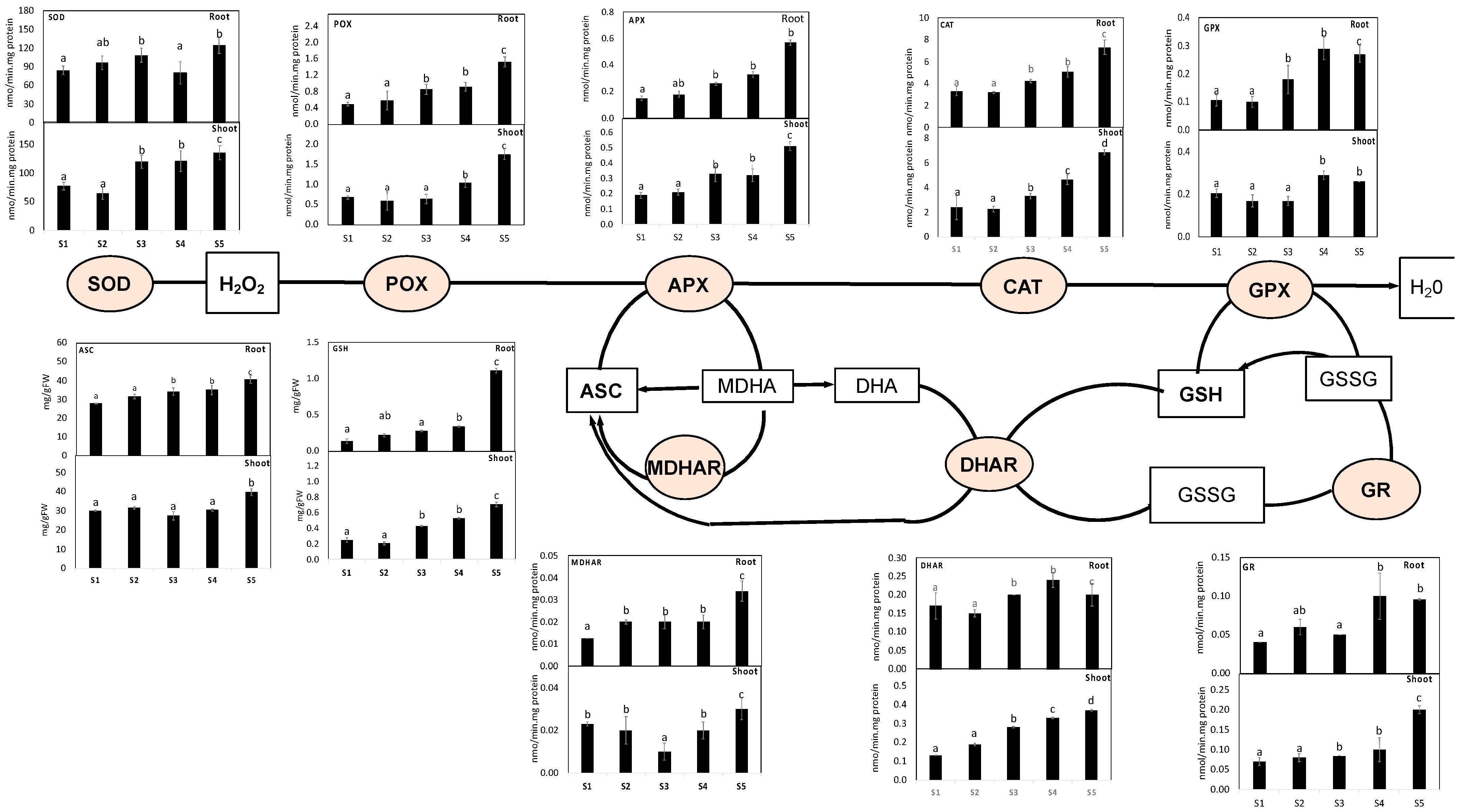
3.4. ROS Production and Oxidative Damage
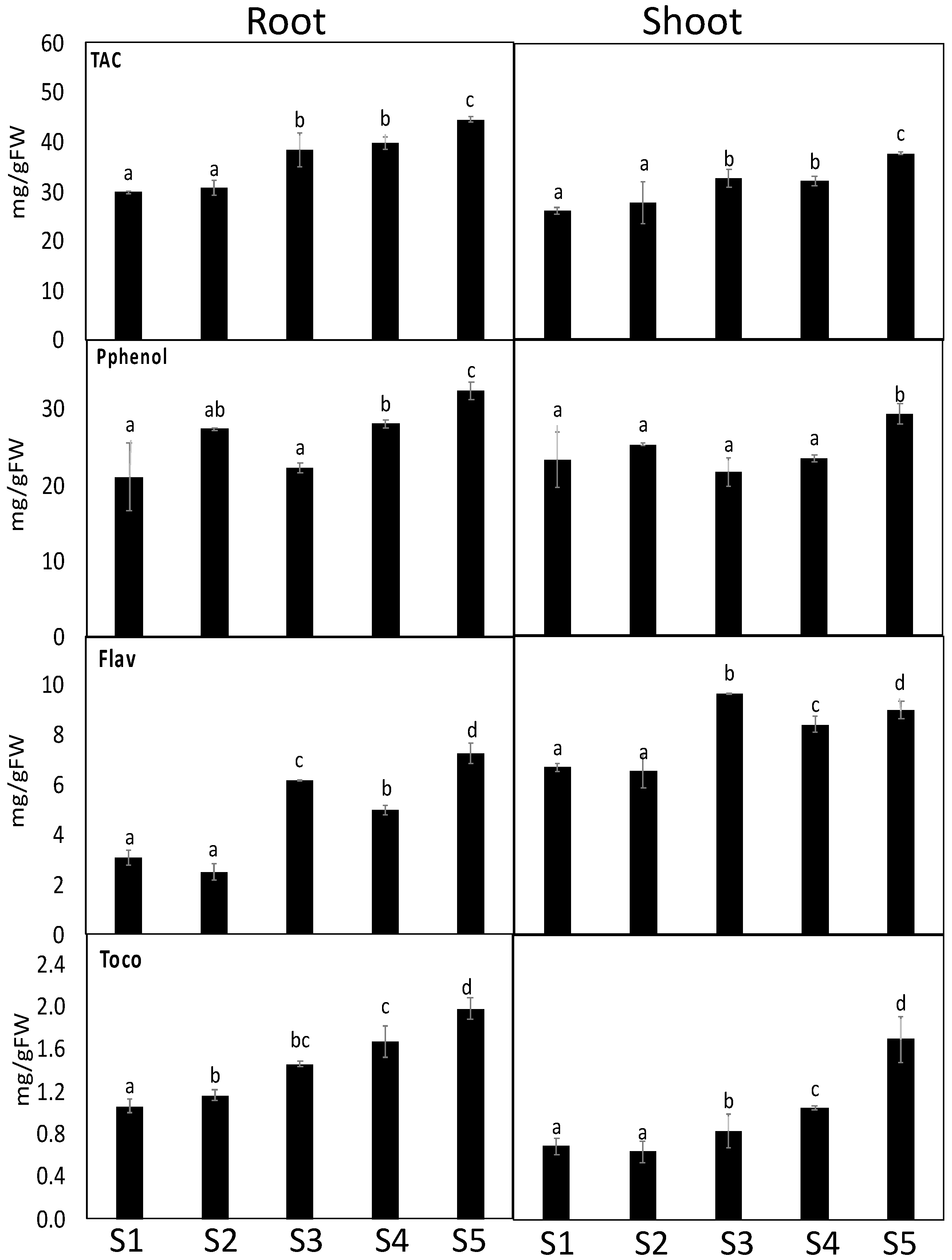
3.5. S. portulacastrum Showed Induced Defense System
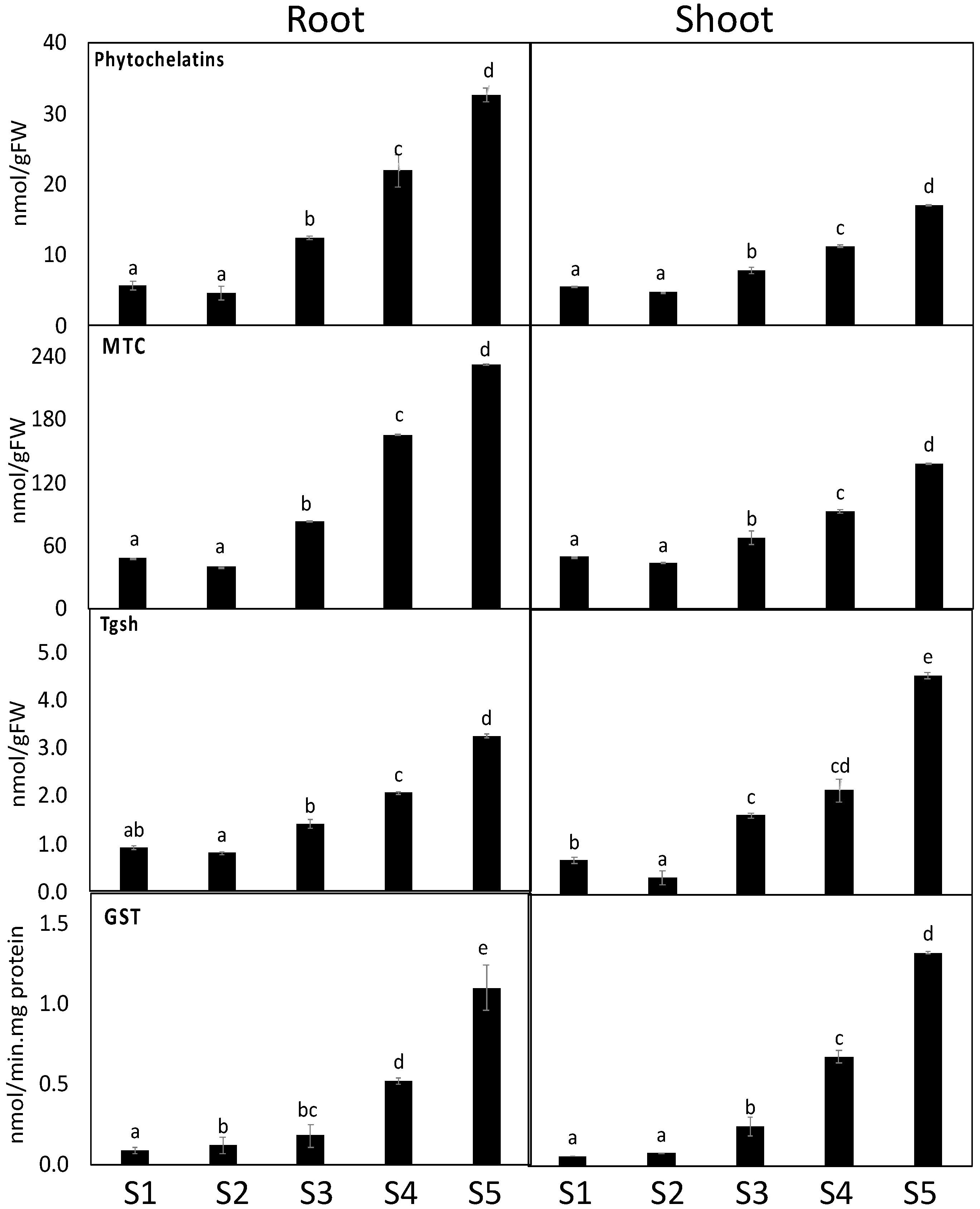
3.6. Heavy Metal Detoxification Was More Pronounced in S. portulacastrum Roots
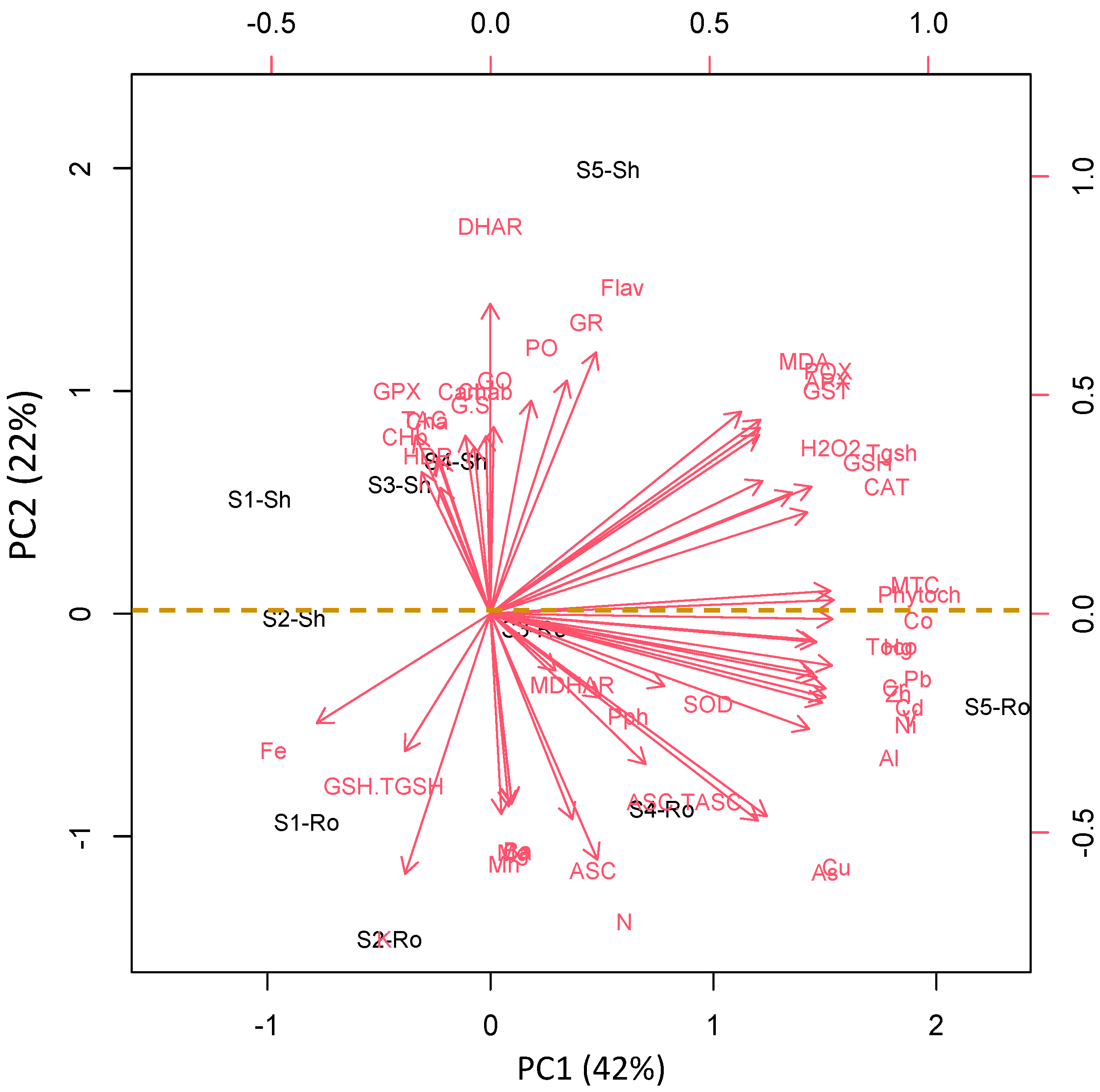
3.7. Organ and Site-Specific Responses Are Supported by PCA Analysis
4. Discussion
4.1. Soil Contamination Impact on the Plant Cover and Biodiversity
4.2. Identification and Selection of Tolerant Species
4.3. S. portulacastrum Accumulated High Levels of Heavy Metals
5. Conclusions
6. Future Perspectives and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, K.K.; Gupta, N.; Kumar, A.; Reece, L.M.; Singh, N.; Rezania, S.; Khan, S.A. Mechanistic understanding and holistic approach of phytoremediation: A review on application and future prospects. Ecol. Eng. 2018, 120, 274–298. [Google Scholar] [CrossRef]
- Igwe, J.C.; Abia, A.A. A bioseparation process for removing heavy metals from waste water using biosorbents. Afr. J. Biotechnol. 2006, 5, 1167–1179. [Google Scholar]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belgacem, A.O.; Tarhouni, M.; Louhaichi, M. Effect of protection on plant community dynamics in the Mediterranean arid zone of southern Tunisia: A case study from BouHedma national park. Land Degrad. Dev. 2011, 24, 57–62. [Google Scholar] [CrossRef]
- Bayouli, I.T.; Bayouli, H.T.; Dell’Oca, A.; Meers, E.; Sun, J. Ecological indicators and bioindicator plant species for biomonitoring industrial pollution: Eco-based environmental assessment. Ecol. Indic. 2021, 125, 107508. [Google Scholar] [CrossRef]
- Fakhry, A.; Migahid, M. Effect of cement dust kiln pollution on the vegetation in Western Mediterranean desert of Egypt. Int. J. Environ. Chem. Ecol. Geol. Geophys. Eng. 2011, 5, 480–486. [Google Scholar]
- Horsfall, M.; Spiff, A. Effect of temperature on the sorption of Pb2+ and Cd2+ from aqueous solution by Caladium bicolor (wild cocoyam) biomass. Electron. J. Biotechnol. 2005, 8, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Moosavi, S.G.; Seghatoleslami, M.J. Phytoremediation: A review. Adv. Agri. Biol. 2013, 1, 5–11. [Google Scholar]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Sébastjen, S.; Zinta, G.; Hassan, Y.H.; Abdel-Mawgoud, M.; Alkhalifah, D.A.; Hozzein, W.N.; Asard, H.; Abuelsoud, W. Soil arsenic toxicity differentially impacts C3 (barley) and C4 (maize) crops. J. Hazard. Mater. 2021, 6, 125331. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; AbdElgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Hamoud, Y.A.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Phytoremediation: Synergistic use of plants and bacteria to clean up the environment. Biotechnol. Adv. 2003, 21, 383–393. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell. Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef] [Green Version]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Ann. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, B.L.; Callahan, D.; Zima, H.P. Parallel programmability and the chapel language. Int. J. High. Perform. Comput. Appl. 2007, 21, 291–312. [Google Scholar] [CrossRef]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557–1563. [Google Scholar] [CrossRef]
- Jaskulak, M.; Rorat, A.; Grobelak, A.; Kacprzak, M. Antioxidative enzymes and expression of rbcL gene as tools to monitor heavy metal-related stress in plants. J. Environ. Manag. 2018, 218, 71–78. [Google Scholar] [CrossRef]
- Lokhande, V.H.; Nikam, T.D.; Patade, V.Y.; Ahire, M.L.; Suprasanna, P. Effects of optimal and supra-optimal salinity stress on antioxidative defence, osmolytes and in vitro growth responses in Sesuvium portulacastrum L. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 41–49. [Google Scholar] [CrossRef]
- Daget, P.; Godron, M. Pastoralisme: Troupeaux, Espaces et Sociétés; Hatier-Aupelf: Uref, France, 1995; p. 510. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. The interrelations of certain analytic and synthetic phytosociological characters. Ecology 1950, 31, 434–455. [Google Scholar] [CrossRef]
- Collenette, S. An Illustrated Guide to the Flowers of Saudi Arabia; Corpion Publishing Ltd.: London, UK, 1985. [Google Scholar]
- Collentette, S. Wild Flowers of Saudi Arabia; National Commission for Wildlife Conservation and Development: Riyadh, Saudi Arabia, 1999. [Google Scholar]
- Miller, A.G.; Cope, T.A. Flora of the Arabian Peninsula. I: 1-586. Mosallam HAM 2007. Comparative Study on the Vegetation of Protected and Nonprotected Areas, Sudera, Taif, Saudi Arabia. Int. J. Agric. Biol. 1996, 9, 202–214. [Google Scholar]
- Chaudhary, S. Flora of the Kingdom o/Saudi Arabia; Ministry of Agriculture and Water: Riyadh, Saudi Arabia, 2001; Volume 2, pp. 1–432. [Google Scholar]
- Raunkiaer, C. Life Forms of Plants and Statistical Geography; Arno Press: Oxford, UK, 1934; 632p. [Google Scholar]
- Zohary, M. Geobotanical Foundations of the Middle East; Gustav Fischer Verlag Press: Jena, Germany; Stuttgart, Swets & Zeitlinger: Amsterdam, The Netherlands, 1973; Volume 2. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Violante, A.; Cozzolino, V.; Perelomov, L.; Caporale, A.G.; Pigna, M. Mobility and bioavailability of heavy metals and metalloids in soil environments. J. Soil Sci. Plant Nutr. 2010, 10, 268–292. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.S.; Kadier, A.; Malyan, S.K.; Ahmad, A.; Bishnoi, N.R. Phytoremediation and Rhizoremediation: Uptake, Mobilization and Sequestration of Heavy Metals by Plants. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Singh, D., Singh, H., Prabha, R., Eds.; Springer: Singapore, 2017; pp. 367–394. [Google Scholar]
- AbdElgawad, H.; Zinta, G.; Hamed, B.A.; Selim, S.; Beemster, G.; Hozzein, W.N.; Wadaan, M.A.M.; Asard, H.; Abuelsoud, W. Maize roots and shoots show distinct profiles of oxidative stress and antioxidant defense under heavy metal toxicity. Environ. Pollut. 2020, 258, 113705. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon dioxide nanoparticles ameliorate the phytotoxic hazards of aluminum in maize grown on acidic soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef] [PubMed]
- Hemphill, J.K.; Venketeswaran, S. Chlorophyll and carotenoid accumulation in three chlorophyllous callus phenotypes of Glycine max. Am. J. Bot. 1978, 65, 1055–1063. [Google Scholar] [CrossRef]
- Feierabend, J.; Beevers, H. Developmental studies on microbodies in wheat leaves: I. Conditions influencing enzyme development. Plant Physiol. 1972, 49, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Novitskaya, N.; Trevanion, S.J.; Driscoll, S.; Foyer, C.H.; Noctor, G. How does photorespiration modulate leaf amino acid contents? A dual approach through modelling and metabolite analysis. Plant Cell Environ. 2002, 25, 821–835. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Y.; Woollard, A.C.; Wolff, S.P. Hydrogen peroxide production during experimental protein glycation. FEBS Lett. 1990, 268, 69–71. [Google Scholar] [CrossRef] [Green Version]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.A.; Barrow, C.J. A simple 96-well microplate method for estimation of total polyphenol content in seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content inpropolis by two complementary colorimetric methods. J. Food Drug Analy. 2002, 10, 178–182. [Google Scholar]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate–glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef]
- Drotar, A.; Phelps, P.; Fall, R. Evidence for glutathione peroxidase activities in cultured plant cells. Plant Sci. 1985, 42, 35–40. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mozer, T.J.; Tiemeier, D.C.; Jaworski, E.G. Purification and characterization of corn glutathione S-transferase. Biochemistry 1983, 22, 1068–1072. [Google Scholar] [CrossRef]
- de Knecht, J.A.; Koevoets, P.L.; Verkleij, J.A.; Ernst, W.H. Evidence against a role for phytochelatins in naturally selected increased cadmium tolerance in Silene vulgaris (Moench) Garcke. New Phytol. 1992, 122, 681–688. [Google Scholar] [CrossRef]
- Al-Sherif, E.A.; Ayesh, A.M.; Rawi, S.M. Floristic composition, life form and chorology of plant life at Khulais region western Saudi Arabia. Pak. J. Bot. 2013, 45, 29–38. [Google Scholar]
- Al-Sherif, E.A.; Fadl, M.A. Floristic study of the Al-Shafa Highlands in Taif, western Saudi Arabia. Flora 2016, 225, 20–29. [Google Scholar] [CrossRef]
- Fadl, M.A.; Al-Yasi, H.M.; Alsherif, E.A. Impact of elevation and slope aspect on floristic composition in wadi Elkor, Sarawat Mountain, Saudi Arabia. Sci. Rep. 2021, 11, 16160. [Google Scholar] [CrossRef]
- Chen, W.; Koide, R.T.; Eissenstat, D.M. Nutrient foraging by mycorrhizas: From species functional traits to ecosystem processes. Funct. Ecol. 2018, 32, 858–869. [Google Scholar] [CrossRef] [Green Version]
- Boutin, C.; Carpenter, D.V. Assessment of wetland/upland vegetation communities and evaluation of soil-plant contamination by polycyclic aromatic hydrocarbons and trace metals in regions near oil sands mining in Alberta. Sci. Total Environ. 2017, 576, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; McLaughlin, N.; Zhang, X.; XU, M.; Liang, A. Effect of tillage and crop residue on soil temperature following planting for a Black soil in Northeast China. Sci. Rep. 2018, 8, 4500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortega-Rosas, C.I.; Enciso-Miranda, C.A.; Macías-Duarte, A.; Morales-Romero, D.; Villarruel-Sahagún, L. Urban vegetation cover correlates with environmental variables in a desert city: Insights of mitigation measures to climate change. Urban Ecosyst. 2020, 23, 1191–1207. [Google Scholar] [CrossRef]
- Edward, E.L.; Bruce, M.G.; Delonay, A.J. Environ. Toxicol. Risk Assess; ASTM: West Conshohocken, PA, USA, 1998; pp. 1–417. [Google Scholar]
- Palacios, G.; Gomez, I.; Carbonell-Barrachina, A.; Pedreño, J.N.; Mataix, J. Effect of nickel concentration on tomato plant nutrition and dry matter yield. J. Plant Nutr. 1998, 21, 2179–2191. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Mocquot, B.; Vangronsveld, J.; Clijsters, H.; Mench, M. Copper toxicity in young maize (Zea mays L.) plants: Effects on growth, mineral and chlorophyll contents, and enzyme activities. Plant Soil. 1996, 182, 287–300. [Google Scholar] [CrossRef]
- Vinit-Dunand, F.; Epron, D.; Alaoui-Sossé, B.; Badot, P.M. Effects of copper on growth and on photosynthesis of mature and expanding leaves in cucumber plants. Plant Sci. 2002, 163, 53–58. [Google Scholar] [CrossRef]
- Rai, P.K. Impacts of particulate matter pollution on plants: Implications for environmental biomonitoring. Ecotoxicol. Environ. Saf. 2016, 129, 120–136. [Google Scholar] [CrossRef]
- Feng, J.; Lin, Y.; Yang, Y.; Shen, Q.; Huang, J.; Wang, S.; Zhu, X.; Li, Z. Tolerance and bioaccumulation of Cd and Cu in Sesuvium portulacastrum. Ecotoxicol. Environ. Saf. 2018, 147, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, P.; Prince, W.S.; Sivakumar, S.; Subbhuraam, C.V. Prosopis juliflora a green solution to decontaminate heavy metal (Cu and Cd) contaminated soils. Chemosphere 2005, 60, 1493–1496. [Google Scholar] [CrossRef]
- Davis, N.E.; Death, C.E.; Coulson, G.; Newby, L.; Hufschmid, J. Interspecific variation in the diets of herbivores in an industrial environment: Implications for exposure to fluoride emissions. Environ. Sci. Pollut. Res. 2016, 23, 10165–10176. [Google Scholar] [CrossRef] [PubMed]
- Ojuederie, O.B.; Babalola, O.O. Microbial and Plant-Assisted Bioremediation of Heavy Metal Polluted Environments: A Review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuypers, A.; Remans, T.; Weyens, N.; Colpaert, J.; Vassilev, A.; Vangronsveld, J. Soil-plant relationships of heavy metals and metalloids. In Heavy Metals in Soils; Springer: Dordrecht, The Netherlands, 2013; pp. 161–193. [Google Scholar]
- Cuypers, A.; Hendrix, S.; Amaral dos Reis, R.; De Smet, S.; Deckers, J.; Gielen, H.; Jozefczak, M.; Loix, C.; Vercampt, H.; Vangronsveld, J.; et al. Hydrogen peroxide, signaling in disguise during metal phytotoxicity. Front. Plant Sci. 2016, 7, 470. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Gill, S.S.; Gill, R.; Hasanuzzaman, M.; Duarte, A.C.; Pereira, E.; Ahmad, I.; Tuteja, R.; Tuteja, N. Metal/metalloid stress tolerance in plants: Role of ascorbate, its redox couple, and associated enzymes. Protoplasma 2014, 251, 1265–1283. [Google Scholar] [CrossRef]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar] [CrossRef] [Green Version]
- Burzyński, M.; Żurek, A. Effects of copper and cadmium on photosynthesis in cucumber cotyledons. Photosynthetica 2007, 45, 239–244. [Google Scholar] [CrossRef]
- Černý, M.; Habánová, H.; Berka, M.; Luklová, M.; Brzobohatý, B. Hydrogen peroxide: Its role in plant biology and crosstalk with signalling networks. Int. J. Molecular Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Wang, H.; Chen, F.; Mao, R.; Liu, H.; Qu, J. Efficient treatment of an electroplating wastewater containing heavy metal ions, cyanide, and organics by H2O2 oxidation followed by the anodic Fenton process. Water Sci. Technol. 2013, 68, 1329–1335. [Google Scholar] [CrossRef]
- Dietz, K.J. Redox regulation of transcription factors in plant stress acclimation and development. Antioxid. Redox Signal. 2014, 21, 1356–1372. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Rhee, J.S.; Jeong, C.B.; Seo, J.S.; Park, G.S.; Lee, Y.M.; Lee, J.S. Heavy metals induce oxidative stress and trigger oxidative stress-mediated heat shock protein (hsp) modulation in the intertidal copepod Tigriopus japonicus. Comp. Biochem. Physiol. 2014, 166, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, M.; Abdel-Farid, I.B.; Choi, Y.H.; Verpoorte, R. Metal ion-inducing metabolite accumulation in Brassica rapa. J. Plant Physiol. 2008, 165, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Skłodowska, M.; Gajewska, E.; Kuźniak, E.; Wielanek, M.; Mikiciński, A.; Sobiczewski, P. Antioxidant profile and polyphenol oxidase activities in apple leaves after Erwinia amylovora infection and pretreatment with a benzothiadiazole-type resistance inducer (BTH). J. Phytopath. 2011, 159, 495–504. [Google Scholar] [CrossRef]
- Cestone, B.; Cuypers, A.; Vangronsveld, J.; Sgherri, C.; Navari-Izzo, F. The influence of EDDS on the metabolic and transcriptional responses induced by copper in hydroponically grown Brassica carinata seedlings. Plant Physiol. Bioch. 2012, 55, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Grúz, J.; Bačkor, M.; Strnad, M.; Repčák, M. Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep. 2009, 28, 135–143. [Google Scholar] [CrossRef]
- Duarte, B.; Delgado, M.; Caçador, I. The role of citric acid in cadmium and nickel uptake and translocation, in Halimione portulacoides. Chemosphere 2007, 69, 836–840. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Swarnavo, C. Cellular and Molecular Phytotoxicity of Lead and Mercury. In Cellular and Molecular Phytotoxicity of Heavy Metals; Springer: Cham, Switzerland, 2020; pp. 373–387. [Google Scholar]
- Zhu, X.; Victor, T.W.; Ambi, A.; Sullivan, J.K.; Hatfield, J.; Xu, F.; Miller, L.M.; Van Nostrand, W.E. Copper accumulation and the effect of chelation treatment on cerebral amyloid angiopathy compared to parenchymal amyloid plaques. Metallomics 2020, 12, 539–546. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M.; Słaba, M.; Mazur, J. Effect of nickel on antioxidative enzyme activities, proline and chlorophyll contents in wheat shoots. Biol. Plant. 2006, 50, 653–659. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1 | S2 | S3 | S4 | S5 | |
|---|---|---|---|---|---|
| Evenness (R) | 0.39 a | 0.36 a | 0.26 b | 0.24 b | 0.22 b |
| Shannon Index | 0.14 a | 0.16 a | 0.18 b | 0.19 c | 0.21 c |
| Species richness | 12 a | 10 b | 10 b | 9 c | 5 d |
| Cover % | 8 a | 9 a | 10 b | 10 b | 16 c |
| S. portulacastrum density | 0 a | 1.2 b | 7.3 c | 9.5 c | 10.33 d |
| S. portulacastrum frequency | 0 a | 15 b | 65 c | 83 d | 100 e |
| Location | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|
| Cd (µg/gm) | 0.43 ± 0.006 a | 1.456 ± 0.30 b | 3.045 ± 0.62 c | 4.962 ± 0.57 cd | 5.19 ± 0.37 d |
| Ni (µg/gm) | 0.441 ± 0.15 a | 0.403 ± 0.18 a | 0.804 ± 0.22 b | 1.115 ± 0.57 bc | 1.26 ± 0.71 c |
| As (µg/gm) | 0.461 ± 0.14 a | 0.336 ± 0.16 a | 0.232 ± 0.08 b | 0.350 ± 0.038 a | 0.45 ± 0.07 a |
| Cu (µg/gm) | 0.863 ± 0.02 a | 1.345 ± 0.14 b | 1.109 ± 0.17 a | 1.349 ± 1.05 b | 1.18 ± 0.14 a |
| Pb (µg/gm) | 0.072 ± 0.001 a | 1.154 ± 0.35 b | 4.734 ± 0.61 c | 7.408 ± 0.59 d | 10.70 ± 0.9 e |
| Co (µg/gm) | 0.108 ± 0.00 a | 0.751 ± 0.12 b | 3.917 ± 0.92 c | 5.767 ± 0.30 d | 8.67 ± 0.84 e |
| Hg (µg/gm) | 0.088 ± 0.01 a | 0.36 ± 0.01 b | 2.76 ± 0.603 c | 5.793 ± 0.45 d | 5.65 ± 0.70 d |
| Al (µg/gm) | 0.046 ± 0.02 a | 0.396 ± 0.17 b | 0.323 ± 0.36 b | 0.629 ± 0.05 c | 0.72 ± 0.21 c |
| V (µg/gm) | 0.046 ± 0.004 a | 0.315 ± 0.13 b | 0.470 ± 0.14 b | 0.682 ± 0.63 c | 0.82 ± 0.17 d |
| Cr (µg/gm) | 0.060 ± 0.02 a | 0.261 ± 0.06 b | 0.617 ± 0.16 c | 0.735 ± 0.00 c | 1.02 ± 0.24 d |
| Zn (µg/gm) | 0.046 ± 0.0 a | 0.096 ± 0.01 b | 0.594 ± 0.05 c | 0.776 ± 0.097 c | 0.89 ± 0.08 d |
| Mn (µg/gm) | 0.079 ± 0.01 a | 0.15 ± 0.02 b | 0.072 ± 0.01 a | 0.178 ± 0.08 b | 0.08 ± 0.010 b |
| Mg (µg/gm) | 0.064 ± 0.007 a | 0.142 ± 0.02 b | 0.062 ± 0.00 a | 0.159 ± 0.07 b | 0.07 ± 0.01 a |
| Ca (µg/gm) | 0.050 ± 0.017 a | 0.104 ± 0.01 b | 0.047 ± 0.00 a | 0.117 ± 0.04 b | 0.05 ± 0.00 a |
| Ba (µg/gm) | 0.028 ± 0.01 a | 0.069 ± 0.02 b | 0.029 ± 0.01 a | 0.076 ± 0.02 b | 0.03 ± 0.01 a |
| Fe (µg/gm) | 0.565 ± 0.08 c | 0.134 ± 0.03 b | 0.060 ± 0.01 a | 0.151 ± 0.07 b | 0.06 ± 0.01 a |
| K (µg/gm) | 1.422 ± 0.13 c | 1.06 ± 0.00ab | 0.871 ± 0.12 a | 1.100 ± 0.8 ab | 0.87 ± 0.07 a |
| N (µg/gm) | 8.56 ± 1.3 b | 7.73 ± 2.17 a | 7.68 ± 0.99 a | 8.14 ± 7.9 ab | 7.77 ± 0.3 a |
| Phenols (µg/gm) | 32.15 ± 0.08 bc | 12.63 ± 3.0 a | 27.02 ± 4.2 b | 43.39 ± 3.7 d | 59.1 ± 5.7 d |
| Citric acid (µg/gm) | 31.50 ± 3.8 c | 4.86 ± 0.86 a | 5.26 ± 1.09 a | 5.75 ± 0.1 a | 7.20 ± 0.64 b |
| pH | 7.3 ± 0.01 a | 7.3 ± 0.01 a | 7.3 ± 0.03 a | 7.4 ± 0.02 a | 7.80 ± 0.09 a |
| O.M (%) | 0.72 ± 0.09 a | 1.38 ± 0.03 a | 1.41 ± 0.02 a | 1.61 ± 0.02 a | 1.94 ± 0.03 a |
| E.C (ds m−1) | 0.71 ± 0.01 b | 0.91 ± 0.01 b | 0.85 ± 0.01 b | 0.99 ± 0.01 a | 1.05 ± 0.01 a |
| Sands (%) | 61.09 ± 1.30 a | 67.74 ± 3.20 a | 61.49 ± 1.60 a | 70.78 ± 1.36 a | 73.99 ± 0.04 a |
| Silts (%) | 21.6 ± 0.89 a | 17.90 ± 1.04 a | 21.2 ± 0.89 a | 16.00 ± 0.68 a | 14.44 ± 0.04 a |
| Clay (%) | 17.31 ± 0.78 a | 14.36 ± 1.05 a | 17.31 ± 0.8/8 a | 13.22 ± 0.74 a | 11.57 ± 0.7 a |
| S1 | S2 | S3 | S4 | S5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | |
| Cd | 1.90 ±0.01 a | 13.39 ± 2.74 a | 21.27 ± 5.1 b | 57.52 ± 6.5 b | 46.5 ± 7.3 c | 127.9 ± 13 c | 76.8 ± 6.5 d | 202.4 ± 22 d | 73.21 ± 8.6 e | 296.63± 64 e |
| Ni | 1.94 ± 0.35 a | 25.18 ± 9.3 a | 11.44 ± 2.68 b | 30.97 ± 3.4 b | 24.5 ± 1.27 c | 68.8 ± 12.5 c | 35. ± 3.4 d | 99.4 ± 14.5 d | 40.9 ± 4.4 e | 134.9 ± 27.1 e |
| As | 2.03 ± 0.3 a | 24.64 ± 4.6 a | 9.54 ± 2.3 bc | 25.78 ± 2.9 a | 7.20 ± 0.4 b | 20.19 ± 3.5 a | 10.54 ± 0.3 c | 29.70 ± 5.9 b | 13.2 ± 0.24 d | 44.50 ± 10.8 c |
| Cu | 3.80 ± 0.06 a | 47.48 ± 11 a | 38.19 ± 2.1 b | 107.16 ± 19 b | 32.4 ± 1.8 b | 91.1 ± 16.2 b | 38.7 ± 1.1 b | 109.2 ± 21 b | 33.33 ± 2.0 b | 166.84 ± 48 c |
| Pb | 0.32 ± 0.01 a | 1.81 ± 0.57 a | 11.34 ± 3.2 b | 57.05 ± 10.0 b | 47.4 ± 7.0 b | 138. ± 50.7 c | 77.0 ± 11 d | 258.2 ± 59 d | 121 ± 8.8 e | 496.69 ± 63 e |
| Co | 0.47 ± 0.00 a | 2.75 ± 0.9 a | 7.72 ± 0.89 b | 22.38 ± 7.4 b | 39.6 ± 3.03 c | 114.2 ± 34 c | 60.1 ± 1.0 d | 177.7 ± 36 d | 159.17 ± 43 c | 490.17 ± 71 e |
| Hg | 0.40 ± 0.03 a | 2.30 ± 0.7 a | 3.83 ± 0.75 b | 16.14 ± 1.4 b | 28.74 ± 4.3 c | 47.65 ± 4.9 c | 63.0±13.1 d | 186. ± 38.3 d | 81.60 ± 5.1 e | 205.21 ± 45 e |
| Al | 0.19 ± 0.03 a | 1.03 ± 0.25 a | 3.91 ± 0.09 b | 11.05 ± 2.2 c | 2.65 ± 1.23 c | 6.84 ± 2.2 b | 5.2 ± 1.03 d | 15.32 ± 6.3 d | 7.08 ± 0.37 e | 24.05 ± 6.8 e |
| V | 0.20 ± 0.01 a | 1.16 ± 0.36 a | 3.14 ± 0.08 b | 8.85 ± 1.82 b | 4.79 ± 0.20 c | 13.69 ± 3.6 c | 6.76 ± 1.4d | 19.97 ± 8.5 d | 8.78 ± 1.08e | 30.22 ± 10.0 e |
| Cr | 0.26 ± 0.05 a | 1.63 ± 0.7 a | 2.62 ± 0.20 b | 7.53 ± 2.2 b | 6.92 ± 1.62 c | 27.93 ± 3.0 c | 8.32 ± 1.8 c | 24.6 ± 10.8 c | 11.39 ± 2.7 d | 40.07 ± 16 d |
| Zn | 0.20 ± 0.01 a | 1.46 ± 0.31 a | 1.01 ± 0.24 a | 3.60 ± 0.7 b | 6.23 ± 1.4 b | 22.33 ± 4.5 c | 8.1 ± 1.9b c | 29.1 ± 5.9 cd | 9.32 ± 2.22 c | 39.65 ± 9.4 d |
| Mn | 2.22 ± 0.08 a | 3.00 ± 0.32 a | 4.58 ± 0.23 b | 6.18 ± 0.75 b | 2.07 ± 0.09 a | 2.79 ± 0.31 a | 5.1 ± 0.26 b | 6.97 ± 0.80b | 2.19 ± 0.11 a | 2.96 ± 0.34 a |
| Mg | 1.77 ± 0.08 a | 2.39 ± 0.27 a | 4.14 ± 0.26 b | 5.59 ± 0.68 b | 1.79 ± 0.10 a | 2.41 ± 0.28 a | 4.6 ± 0.28 b | 6.21 ± 0.75 b | 1.94 ± 0.12 a | 2.62 ± 0.32 a |
| Ca | 1.29 ± 0.11 a | 1.73 ± 0.23 a | 3.08 ± 0.06 b | 4.16 ± 0.43 b | 1.32 ± 0.01 a | 1.78 ± 0.18 a | 3.4 ± 0.04 b | 4.62 ± 0.47 b | 1.46 ± 0.01 a | 1.96 ± 0.20 a |
| Ba | 0.88 ± 0.09 a | 1.19 ± 0.17 a | 2.16 ± 0.14 b | 2.91 ± 0.36 b | 0.92 ± 0.01 a | 1.25 ± 0.13 a | 2.4 ± 0.13 b | 3.24 ± 0.38 b | 1.01 ± 0.04 a | 1.36 ± 0.15 a |
| Fe | 15.32 ± 0.0 a | 20.67 ± 2.1 a | 3.98 ± 0.4 b | 5.36 ± 0.8 b | 1.74 ± 0.15 c | 2.35 ± 0.30 c | 4.4 ± 0.4 b | 5.98 ± 0.8 b | 1.87 ± 0.18 c | 2.53 ± 0.30 c |
| K | 39 ± 0.70 a | 53.10 ± 5.5 a | 30.32 ± 0.1 b | 40.90 ± 4.1 b | 25.62 ± 0.8 c | 34.57± 3.70 c | 31 ± 0.52 b | 42.39 ± 4.3 b | 24.2 ± 0.22 c | 32.68 ± 3.34 c |
| N | 244.6 ± 18 a | 330 ± 42 a | 204.8 ± 16.9 b | 276.35 ± 37 b | 225.8 ± 5 b | 304.6 ± 30 ab | 234 ± 5.2 ab | 316.9 ± 33 ab | 217± 0.46 c | 293.6 ± 29 ab |
| S1 | S2 | S3 | S4 | S5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BCF | TF | BCF | TF | BCF | TF | BCF | TF | BCF | TF | |
| Cd | 30.98 ± 2.10 a | 0.14 ± 0.09 a | 39.51 ±1.05 b | 0.4 ± 0.02 c | 42.0 ± 2.3 c | 0.36 ± 0.02 c | 40.8 ± 1.0 c | 0.38 ± 0.02 c | 57.20 ± 2.1 d | 0.25 ± 0.05 b |
| Ni | 57.05 ± 2.5 a | 0.08 ± 0.01 a | 76.92 ± 2.3 b | 0.37 ±0.13 b | 85.66± 2.7 c | 0.36 ± 0.05 b | 89.18 ± 2.3 c | 0.36 ± 0.01 b | 107.11 ±14 d | 0.30 ± 0.08 b |
| As | 53.44 ± 1.3 a | 0.08 ± 0.02 a | 76.80 ± 2.4 b | 0.36 ± 0.1 b | 87.1 ± 3.1 b | 0.35 ± 0.04 b | 84.95 ± 1.5 b | 0.35 ± 0.05 b | 99.56 ± 11.3 c | 0.29 ± 0.04 c |
| Cu | 55.00 ± 2.6 a | 0.08 ± 0.01 a | 79.69 ± 2.4 b | 0.36 ± 0.12 c | 82.12 ± 2.5 c | 0.36 ± 0.05 c | 80.95 ± 1.2 c | 0.35 ± 0.01 c | 141.31 ± 18 d | 0.20 ± 0.02 b |
| Pb | 25.20 ± 1.03 a | 0.17 ± 0.03 a | 49.45 ± 2.3 c | 0.2 ± 0. 0 a | 29.2 ± 1.2 a | 0.34 ± 0.0 b | 34.8 ± 1.02 b | 0.30 ± 0.01 b | 46.43 ± 2.3 c | 0.24 ± 0.01 a |
| Co | 25.58 ± 1.5 a | 0.17 ± 0.03 a | 29.81 ± 1.02 a | 0.3 ± 0.08 b | 29.1 ± 1.3 b | 0.35 ± 0.05 b | 30.8 ± 1.0 b | 0.34 ± 0.06 b | 56.51 ±11.3 c | 0.32 ± 0.02 b |
| Hg | 26.03 ± 1.6 b | 0.17 ± 0.01 a | 43.98 ± 1.2 d | 0.24 ± 0.0 a | 17.22 ± 0.9 a | 0.60 ± 0.08 d | 32.1 ± 1.04 b | 0.34 ± 0.02 b | 36.33 ± 3.6 c | 0.40 ± 0.04 c |
| Al | 22.44 ± 1.4 a | 0.18 ± 0.01 a | 27.87 ± 1.4 c | 0.35 ± 0.0 c | 21.16 ± 1.5 a | 0.39 ± 0.04 c | 24.3 ± 1.08 b | 0.34 ± 0.0 bc | 33.51 ± 2.5 c | 0.29 ± 0.01 b |
| V | 25.18 ± 1.5 a | 0.17 ± 0.02 a | 28.13 ± 2.0 a | 0.35 ± 0.08 b | 29.10 ± 1.0 a | 0.35 ± 0.03 b | 29.2 ± 1.02 a | 0.34 ± 0.07 b | 36.80 ± 2.1 b | 0.29 ± 0.03 b |
| Cr | 27.26 ± 1.0 a | 0.16 ± 0.00 a | 28.87 ± 1.09 a | 0.35 ± 0.1 c | 45.27 ± 1.5 c | 0.25 ± 0.03 b | 33.4 ± 0.3 b | 0.34 ± 0.1 c | 39.15 ± 3.6 b | 0.28 ± 0.01 b |
| Zn | 31.80 ± 1.3 a | 0.14 ± 0.03 a | 37.68 ± 1.5 ab | 0.28 ± 0.03 c | 37.6 ± 1.0 ab | 0.28 ± 0.0 c | 37.6 ± 0.8 ab | 0.28 ± 0.02 b | 44.60 ± 2.50 c | 0.24 ± 0.01 b |
| Fe | 36.59 ± 2.1 a | 0.74 ± 0.02 a | 39.93 ± 1.04 b | 0.74 ± 0.6 a | 39.12 ± 1 b | 0.74 ± 0.14 a | 39.6 ± 0.25 b | 0.74 ± 0.14 a | 39.51 ± 1.5 b | 0.74 ± 0.01 a |
| S1 | S2 | S3 | S4 | S5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | |
| FW gm | -- | -- | 84.21 ±5.3 a | 385 ± 12.30 a | 78.25 ± 3.4 a | 315 ± 10.90 b | 56.24 ± 4.2 d | 294 ± 14.20 d | 42.36 ± 3.1 a | 285 ± 11.07 c |
| DW gm | -- | -- | 16.25 ±1.3 b | 89.5 ± 6.05 b | 13.25 ± 1.2 a | 82.8 ± 5.62 a | 11.35 ± 1 a | 79.3 ± 5.4 a | 10.58 ± 0.6 d | 77.2 ± 4.81 c |
| CH a | 2.67 ± 0.2 ab | 2.62 ± 0.3 ab | 2.01 ± 0.04 a | 3.4 ± 0.23 c | 3.23 ± 0.6 c | |||||
| CH b | 0.66 ± 0.02 a | 0.74 ± 0.1 b | 0.59 ± 0.06 a | 0.66 ± 0.05 a | 0.73 ± 0.0 b | |||||
| Ch a + Ch b | 1.51 ± 1.3 a | 1.51 ± 1.3 a | 1.51 ± 1.3 a | 3.47 ± 0.2 b | 3.23 ± 0.6 b | |||||
| Carotenoids | 24.23 ± 0.6 a | 24.1 ± 0.31 a | 33.06 ± 1.0 b | 36.40 ± 4.3 b | 42.95 ± 3.1 c | |||||
| S1 | S2 | S3 | S4 | S5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | |
| GO | 0.70 ± 0.04 a | 0.78 ± 0.02 a | 0.90 ± 0.0 ab | 0.85 ± 0.1 ab | 1.60 ± 0.04 b | |||||
| G/S ratio | 21.46 ± 4.28 a | 20.94 ± 4.06 a | 19.09 ± 0.2 a | 26.3 ± 2.00 b | 35.29 ± 3.06 b | |||||
| HDR | 1.80 ± 0.23 a | 2.01 ± 0.45 a | 2.20 ± 0.00 a | 1.83 ± 0.11 a | 3.69 ± 0.69 b | |||||
| H2O2 | 217.2 ± 53.0 a | 235.0 ± 8.5 a | 241.5± 3.3 ab | 254.2 ± 14.2 b | 282.4 ± 14.4 b | 333.0 ± 51.5 c | 305.79 ± 8.9 b | 307.6 ± 9.7 bc | 354.3 ± 18.4 c | 314 ± 16.6 bc |
| MDA | 2.31 ± 0.55 a | 2.88 ± 0.23 a | 3.00 ± 0.06 b | 3.36 ± 0.13 b | 4.69 ± 0.14 a | 3.72 ± 0.45 b | 5.13 ± 0.34 c | 5.01 ± 0.00 c | 5.54 ± 0.17 c | 7.28 ± 0.94 d |
| PO | 1.44 ± 0.01 a | 1.74 ± 0.04 a | 1.50 ± 0.07 a | 1.39 ± 0.18 a | 1.93 ± 0.16 ab | 1.63 ± 0.10 a | 1.42 ± 0.10 a | 1.52 ± 0.04 a | 1.56 ± 0.04 a | 2.88 ± 0.03 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsherif, E.A.; Al-Shaikh, T.M.; Almaghrabi, O.; AbdElgawad, H. High Redox Status as the Basis for Heavy Metal Tolerance of Sesuvium portulacastrum L. Inhabiting Contaminated Soil in Jeddah, Saudi Arabia. Antioxidants 2022, 11, 19. https://doi.org/10.3390/antiox11010019
Alsherif EA, Al-Shaikh TM, Almaghrabi O, AbdElgawad H. High Redox Status as the Basis for Heavy Metal Tolerance of Sesuvium portulacastrum L. Inhabiting Contaminated Soil in Jeddah, Saudi Arabia. Antioxidants. 2022; 11(1):19. https://doi.org/10.3390/antiox11010019
Chicago/Turabian StyleAlsherif, Emad A., Turki M. Al-Shaikh, Omar Almaghrabi, and Hamada AbdElgawad. 2022. "High Redox Status as the Basis for Heavy Metal Tolerance of Sesuvium portulacastrum L. Inhabiting Contaminated Soil in Jeddah, Saudi Arabia" Antioxidants 11, no. 1: 19. https://doi.org/10.3390/antiox11010019