
Elevated CO2 Differentially Mitigated Oxidative Stress Induced by Indium Oxide Nanoparticles in Young and Old Leaves of C3 and C4 Crops

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Pot Experiment
2.2. Elemental Contents in Plants
2.3. Photosynthesis Related Parameters
2.4. Quantification of Oxidative Damage Markers
2.5. Quantification of Antioxidant Parameters
2.6. Quantification of Detoxification Related Parameters
2.7. Determination of Anthocyanins, Phenolics and Flavonoids and the Activity of Related Enzymes
2.8. Statistical Analysis
3. Results
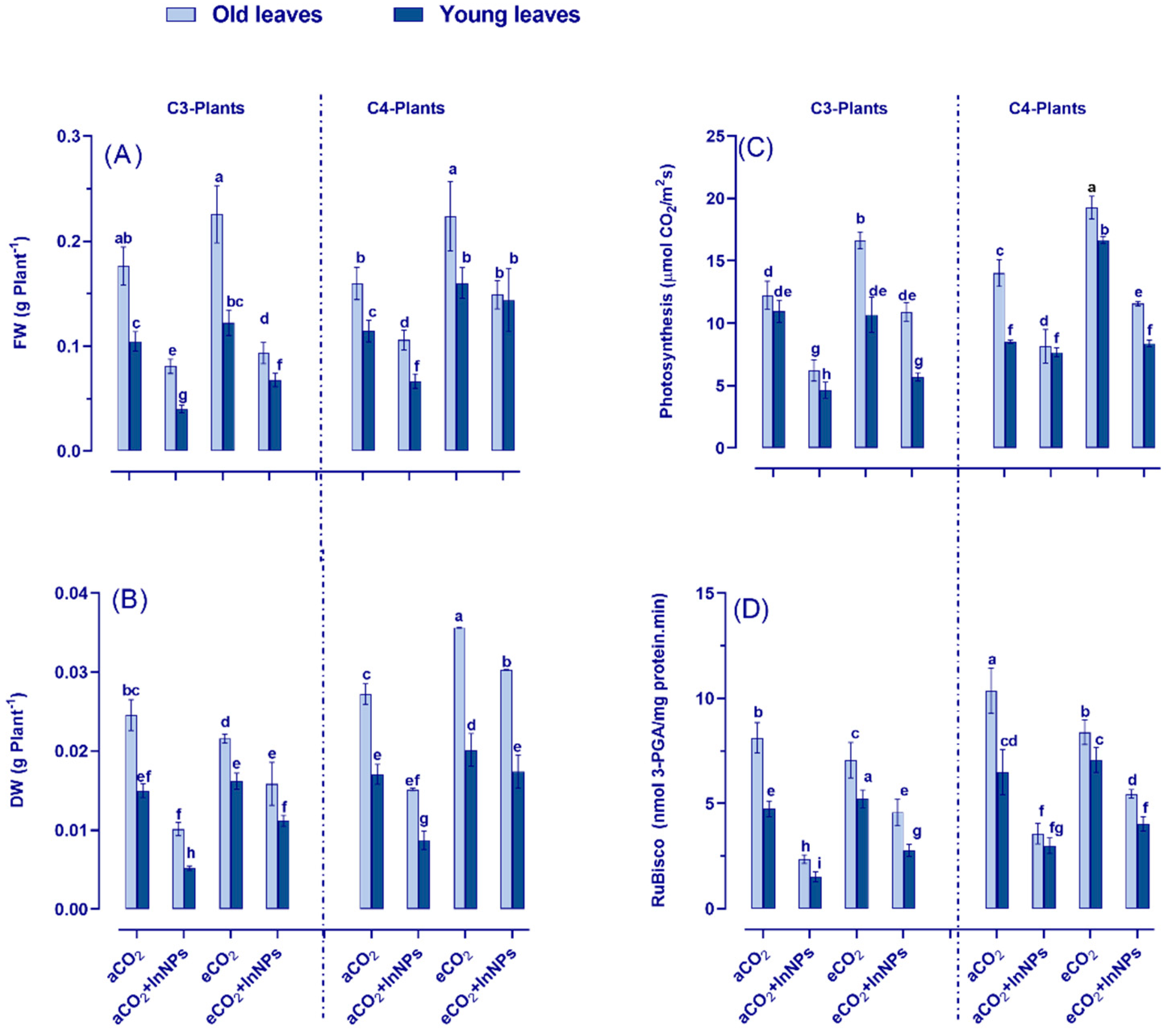
3.1. eCO2 Differentially Enhanced Growth and Photosynthesis of C3- and C4-Plants under the Challenge of In2O3-NPs
3.2. In2O3-NPs Induced in Accumulation, Particularly in C4 Plants and Reduced P and Fe Uptake
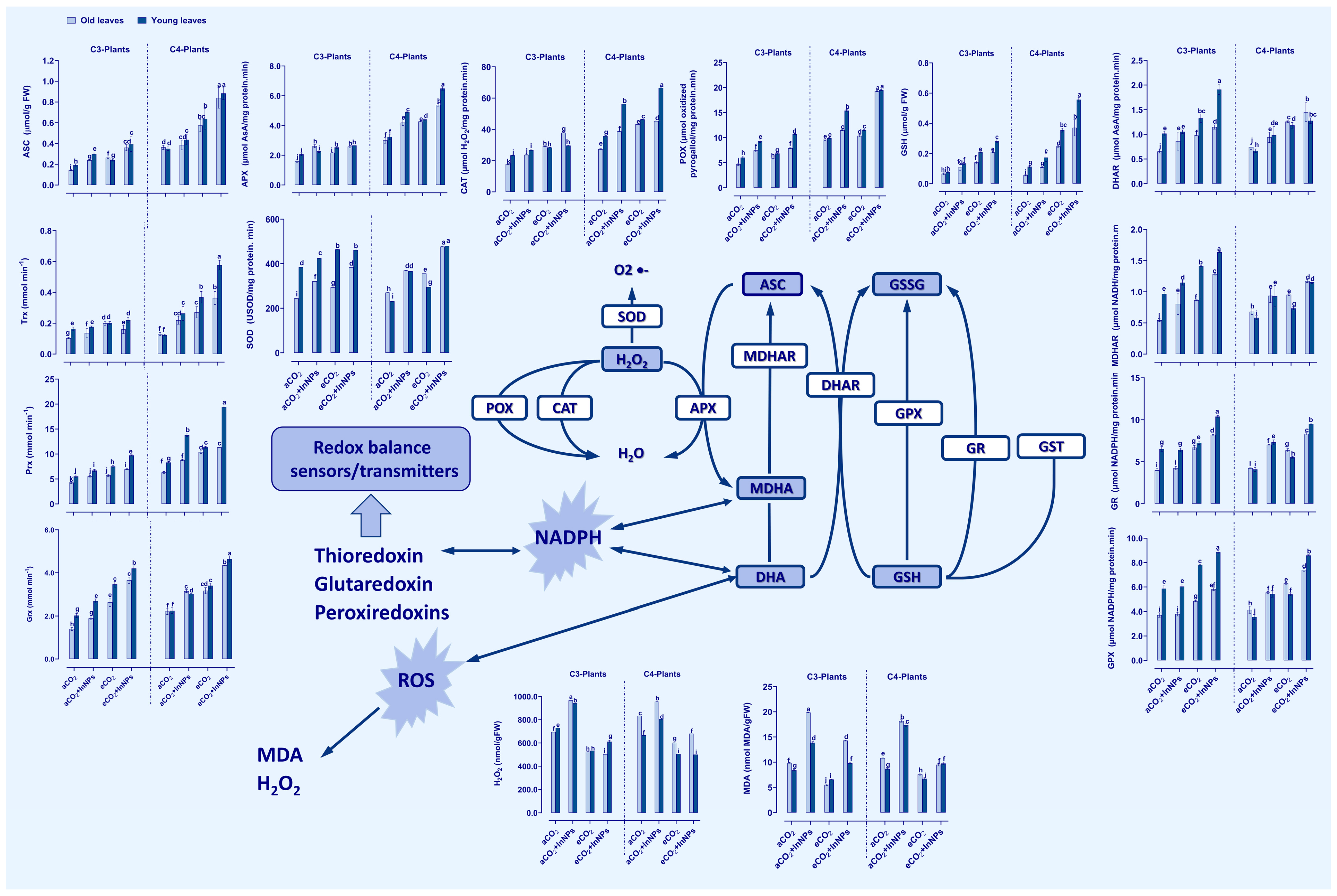
3.3. Increased Antioxidant Defense System in C3- and C4-Plants Grown in In2O3-NP-Polluted Soils Based on Stress Mitigating Impact of eCO2
3.4. In Accumulation under In2O3-NP Stress Induced Differential Oxidative Damage in C3 and C4 Plants, but Not under eCO2 Conditions
3.5. eCO2 Improved the Detoxification System of C3- and C4-Plants under the Challange of In2O3-NPs
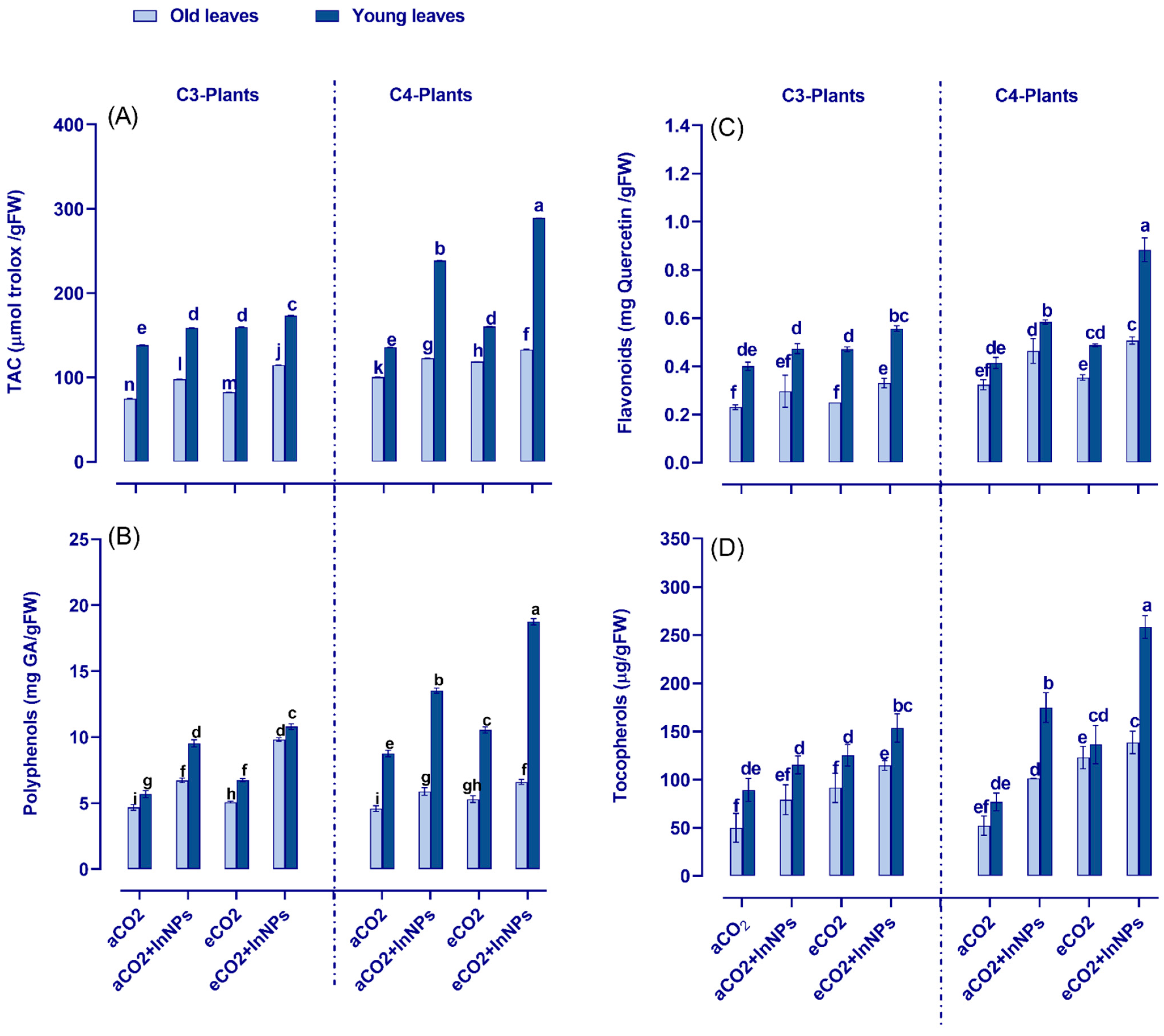
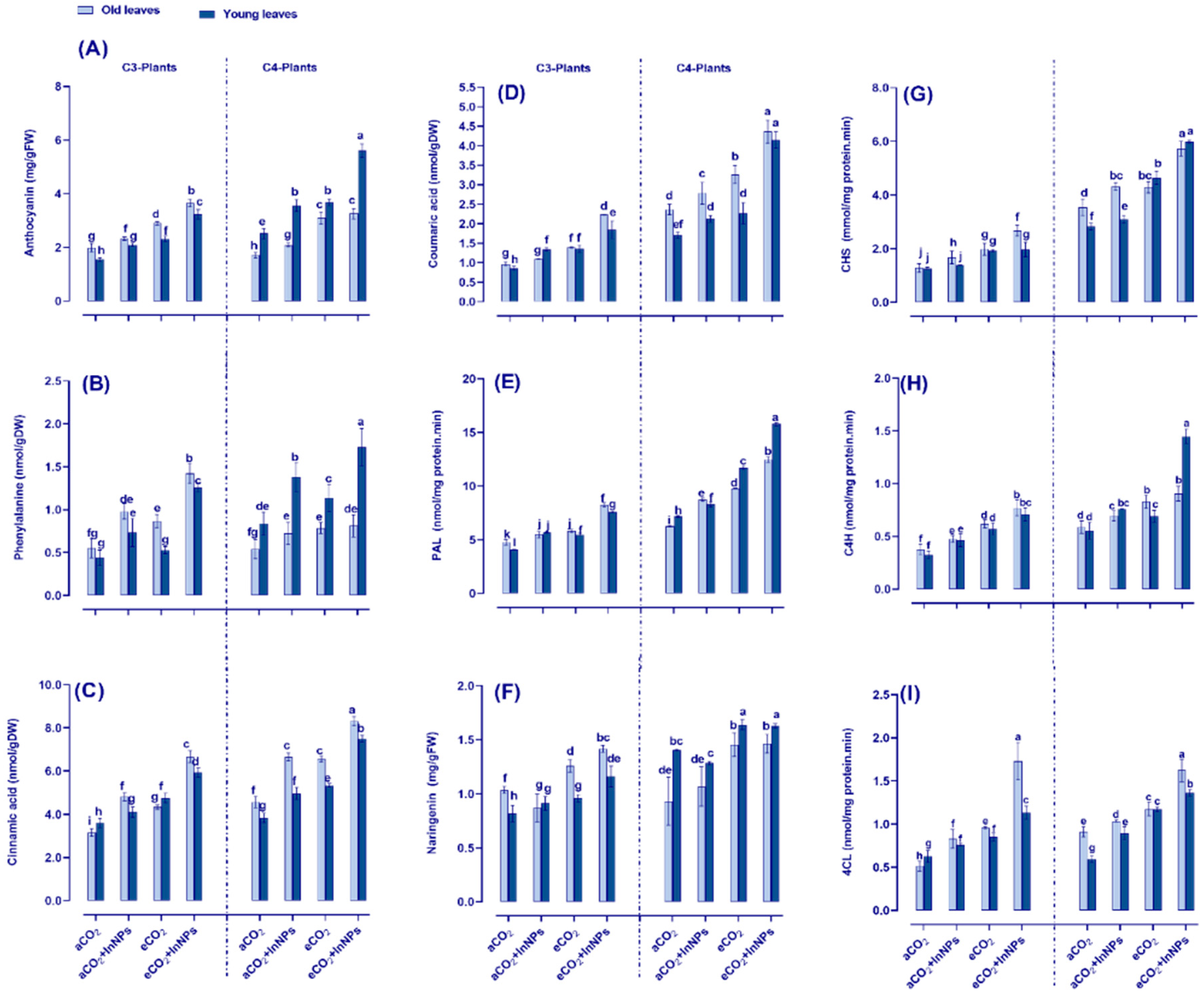
3.6. Anthocyanin Metabolism Greatly Improved by Elevated CO2 in Both C3- and C4-Plants under the Challenge of In2O3-NPs
3.7. SOD Overexpression Increased In-Stress Tolerance in Both C3 and C4 Plants
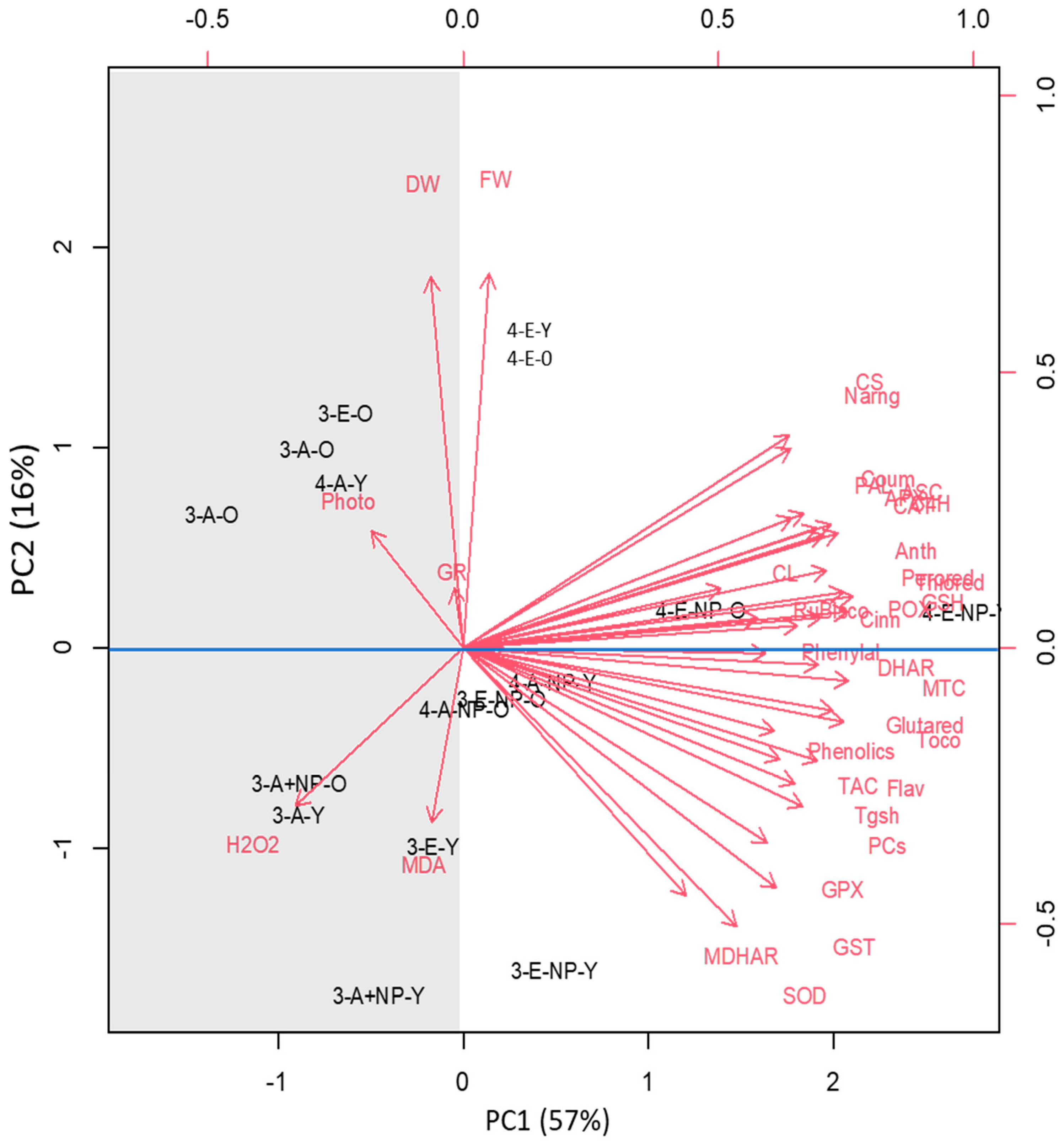
3.8. Species and Developmental Specific Responses
4. Discussion
4.1. eCO2 Alleviated the Growth Reduction and the Oxidative Damage in C3 and C4 Plants Caused by In2O3-NPs
4.2. How Could eCO2 Ameliorat the Oxidative Damage Induced by In2O3-NPs in C3 and C4 Plants?
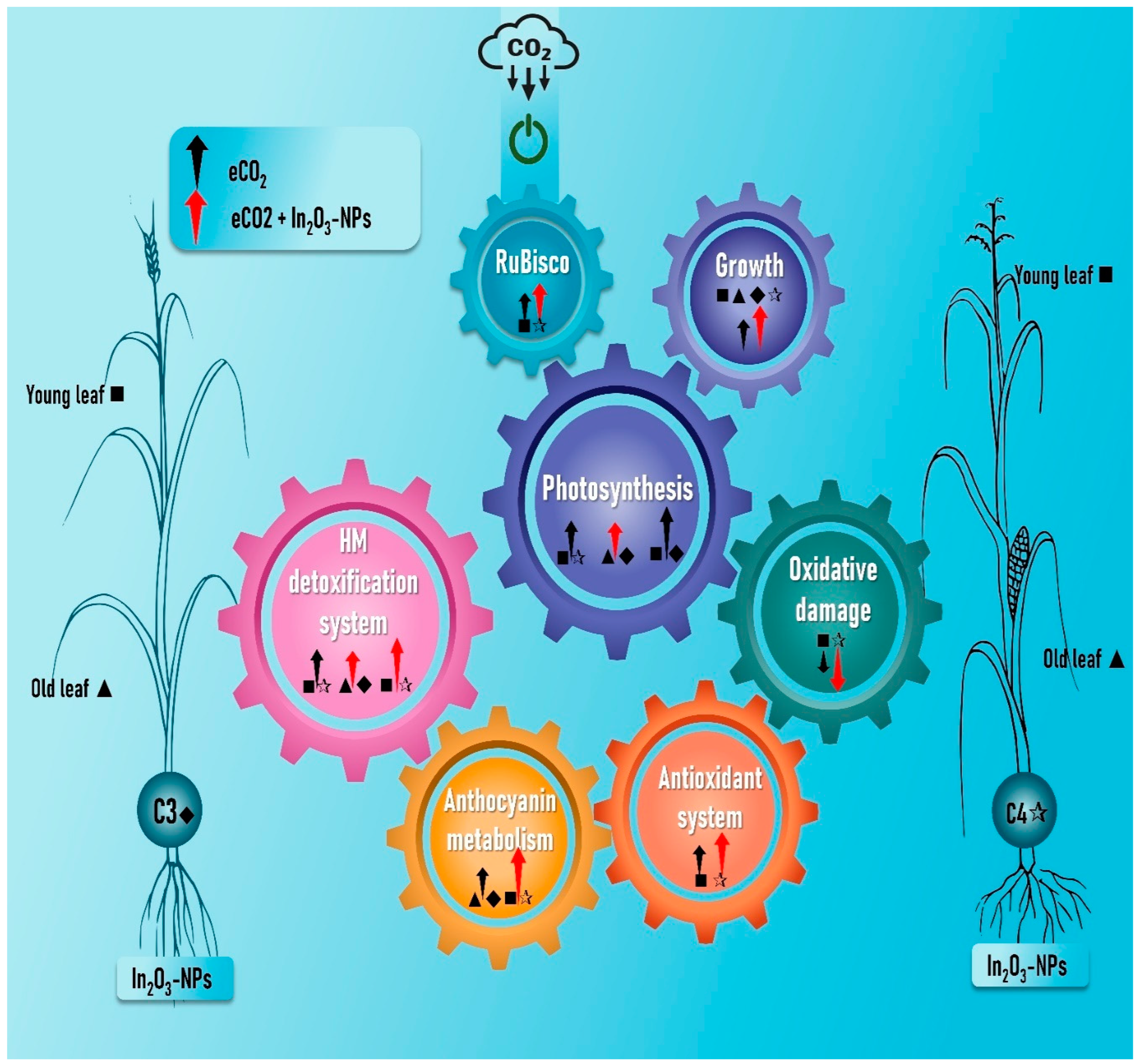
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rajput, V.D.; Minkina, T.; Fedorenko, A.; Tsitsuashvili, V.; Mandzhieva, S.; Sushkova, S.; Azarov, A. Metal oxide nanoparticles: Applications and effects on soil ecosystems. In Soil Contamination: Sources, Assessment and Remediation; Nova Science Publishers: Hauppauge, NY, USA, 2018; pp. 81–106. [Google Scholar]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, K.J.; Virji, M.A.; Park, J.Y.; Stanton, M.L.; Edwards, N.T.; Trapnell, B.C.; Carey, B.; Stefaniak, A.B.; Kreiss, K. Respirable Indium Exposures, Plasma Indium, and Respiratory Health Among Indium-Tin Oxide (ITO) Workers. Am. J. Ind. Med. 2016, 59, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.F.; Wang, S.L.; Lee, D.C.; Hsiao, S.S.Y.; Hashimoto, Y.; Yeh, K.C. Assessment of indium toxicity to the model plant Arabidopsis. J. Hazard. Mater. 2020, 387, 121983. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J. Concentrations of 61 Trace Elements in Sewage Sludge, Farmyard Manure, Mineral Fertiliser, Precipitation and in Oil and Crops; Swedish Environmental Protection Agency: Stockholm, Sweden, 2001. [Google Scholar]
- Kopittke, P.M.; McKenna, B.A.; Blamey, F.P.C.; Wehr, J.B.; Menzies, N.W. Metal-induced cell rupture in elongating roots is associated with metal ion binding strengths. Plant Soil 2009, 322, 303–315. [Google Scholar] [CrossRef]
- Syu, C.H.; Chien, P.H.; Huang, C.C.; Jiang, P.Y.; Juang, K.W.; Lee, D.Y. The growth and uptake of Ga and In of rice (Oryza sative L.) seedlings as affected by Ga and In concentrations in hydroponic cultures. Ecotoxicol. Environ. Saf. 2017, 135, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ahearn, G.A.; Mandal, P.K.; Mandal, A. Mechanisms of heavy-metal sequestration and detoxification in crustaceans: A review. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2004, 174, 439–452. [Google Scholar] [CrossRef]
- USGS. Mineral Commodity Summaries 2020; U.S. Government Publishing Office: Washington, DC, USA, 2020; ISBN 9781411343627. [Google Scholar]
- Jeong, J.; Kim, J.; Seok, S.; Cho, W.S. Indium oxide (In2O3) nanoparticles induce progressive lung injury distinct from lung injuries by copper oxide (CuO) and nickel oxide (NiO) nanoparticles. Arch. Toxicol. 2016, 90, 817–828. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Khan, M.; Alhadlaq, H.; Aldalbahi, A. Nanocubes of indium oxide induce cytotoxicity and apoptosis through oxidative stress in human lung epithelial cells. Colloids Surf. B Biointerfaces 2017, 156, 157–164. [Google Scholar] [CrossRef]
- Landa, P.; Cyrusova, T.; Jerabkova, J.; Drabek, O.; Vanek, T.; Podlipna, R. Effect of Metal Oxides on Plant Germination: Phytotoxicity of Nanoparticles, Bulk Materials, and Metal Ions. Water Air Soil Pollut. 2016, 227, 448. [Google Scholar] [CrossRef]
- Hossain, Z.; Mustafa, G.; Komatsu, S. Plant responses to nanoparticle stress. Int. J. Mol. Sci. 2015, 16, 26644–26653. [Google Scholar] [CrossRef] [Green Version]
- Kamat, J. Reactive oxygen species mediated membrane damage induced by fullerene derivatives and its possible biological implications. Toxicology 2000, 155, 55–61. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant Response to Engineered Metal Oxide Nanoparticles. Nanoscale Res. Lett. 2017, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabbaj, I.; AbdElgawad, H.; Tammar, A.; Alsiary, W.A.; Madany, M.M.Y. Future climate CO2 can harness ROS homeostasis and improve cell wall fortification to alleviate the hazardous effect of Phelipanche infection in pea seedlings. Plant Physiol. Biochem. 2021, 166, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Hassan, Y.M.; Selim, S.; AbdElgawad, H. NiO-nanoparticles induce reduced phytotoxic hazards in wheat (Triticum aestivum L.) grown under future climate CO2. Chemosphere 2019, 220, 1047–1057. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Farfan-Vignolo, E.R.; de Vos, D.; Asard, H. Elevated CO2 mitigates drought and temperature-induced oxidative stress differently in grasses and legumes. Plant Sci. 2015, 231, 1–10. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, U.; Robredo, A.; Lacuesta, M.; Sgherri, C.; Muñoz-Rueda, A.; Navari-Izzo, F.; Mena-Petite, A. The oxidative stress caused by salinity in two barley cultivars is mitigated by elevated CO2. Physiol. Plant. 2009, 135, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Bräutigam, A.; Gowik, U. Photorespiration connects C3 and C4 photosynthesis. J. Exp. Bot. 2016, 67, 2953–2962. [Google Scholar] [CrossRef] [Green Version]
- Voss, I.; Sunil, B.; Scheibe, R.; Raghavendra, A.S. Emerging concept for the role of photorespiration as an important part of abiotic stress response. Plant Biol. 2013, 15, 713–722. [Google Scholar] [CrossRef]
- IPCC. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation—IPCC. Available online: https://www.ipcc.ch/report/managing-the-risks-of-extreme-events-and-disasters-to-advance-climate-change-adaptation/ (accessed on 10 September 2020).
- Van Breusegem, F.; Slooten, L.; Stassart, J.M.; Moens, T.; Botterman, J.; Van Montagu, M.; Inzé, D. Overproduction of Arabidopsis thaliana FeSOD confers oxidative stress tolerance to transgenic maize. Plant Cell Physiol. 1999, 40, 515–523. [Google Scholar] [CrossRef]
- Hemphill, J.K.; Venketeswaran, S. Chlorophyll and Carotenoid Accumulation in Three Chlorophyllous Callus Phenotypes of Glycine max. Am. J. Bot. 1978, 65, 1055. [Google Scholar] [CrossRef]
- Sulpice, R.; Tschoep, H.; Von Korff, M.; Büssis, D.; Usadel, B.; Höhne, M.; Witucka-Wall, H.; Altmann, T.; Stitt, M.; Gibon, Y. Description and applications of a rapid and sensitive non-radioactive microplate-based assay for maximum and initial activity of D-ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Cell Environ. 2007, 30, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.-Y.; Woollard, A.C.S.; Wolff, S.P. Hydrogen peroxide production during experimental protein glycation. FEBS Lett. 1990, 268, 69–71. [Google Scholar] [CrossRef] [Green Version]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1994; Volume 233, pp. 346–357. [Google Scholar]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 76, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirbaş, S.; Acar, O. Superoxide dismutase and peroxidase activities from antioxidative enzymes in Helianthus annuus L. roots during Orobanche cumana Wallr. penetration. Fresenius Environ. Bull. 2008, 17, 1038–1044. [Google Scholar]
- Madany, M.M.Y.; Zinta, G.; Abuelsoud, W.; Hozzein, W.N.; Selim, S.; Asard, H.; Elgawad, H.A. Hormonal seed-priming improves tomato resistance against broomrape infection. J. Plant Physiol. 2020, 250, 153184. [Google Scholar] [CrossRef]
- Zinta, G.; Abdelgawad, H.; Domagalska, M.A.; Vergauwen, L.; Knapen, D.; Nijs, I.; Janssens, I.A.; Beemster, G.T.S.; Asard, H. Physiological, biochemical, and genome-wide transcriptional analysis reveals that elevated CO2 mitigates the impact of combined heat wave and drought stress in Arabidopsis thaliana at multiple organizational levels. Glob. Chang. Biol. 2014, 20, 3670–3685. [Google Scholar] [CrossRef]
- Potters, G.; Horemans, N.; Bellone, S.; Caubergs, R.J.; Trost, P.; Guisez, Y.; Asard, H. Dehydroascorbate influences the plant cell cycle through a glutathione-independent reduction mechanism. Plant Physiol. 2004, 134, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.B.; Khan, P.A. Peroxidase & polyphenol oxidase in excised ragi (Eleusine corocana cv PR 202) leaves during senescence. Indian J. Exp. Biol. 1982, 20, 412–416. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Microplate quantification of enzymes of the plant ascorbate–glutathione cycle. Anal. Biochem. 2008, 383, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Drotar, A.; Phelps, P.; Fall, R. Evidence for glutathione peroxidase activities in cultured plant cells. Plant Sci. 1985, 42, 35–40. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Mozer, T.; Tiemeier, D.; Jaworski, E. Purification and characterization of corn glutathione S-transferase. Biochemistry 1983, 22, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Diopan, V.; Shestivska, V.; Adam, V.; Macek, T.; Mackova, M.; Havel, L.; Kizek, R. Determination of content of metallothionein and low molecular mass stress peptides in transgenic tobacco plants. Plant Cell Tissue Organ Cult. 2008, 94, 291–298. [Google Scholar] [CrossRef]
- De Knecht, J.A.; Koevoets, P.L.M.; Verkleij, J.A.C.; Ernst, W.H.O. Evidence against a role for phytochelatins in naturally selected increased cadmium tolerance in Silene vulgaris (Moench) Garcke. New Phytol. 1992, 122, 681–688. [Google Scholar] [CrossRef]
- Selim, S.; Abuelsoud, W.; Al-Sanea, M.M.; AbdElgawad, H. Elevated CO2 differently suppresses the arsenic oxide nanoparticles-induced stress in C3 (Hordeum vulgare) and C4 (Zea maize) plants via altered homeostasis in metabolites specifically proline and anthocyanin metabolism. Plant Physiol. Biochem. 2021, 166, 235–245. [Google Scholar] [CrossRef]
- Zhao, L.; Peng, B.; Hernandez-Viezcas, J.A.; Rico, C.; Sun, Y.; Peralta-Videa, J.R.; Tang, X.; Niu, G.; Jin, L.; Varela-Ramirez, A.; et al. Stress Response and Tolerance of Zea mays to CeO2 Nanoparticles: Cross Talk among H2O2, Heat Shock Protein, and Lipid Peroxidation. ACS Nano 2012, 6, 9615–9622. [Google Scholar] [CrossRef] [Green Version]
- Naing, A.H.; Kim, C.K. Abiotic stress-induced anthocyanins in plants: Their role in tolerance to abiotic stresses. Physiol. Plant 2021, 172, 1711–1723. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Hassan, Y.M.; Alotaibi, M.O.; Mohammed, A.E.; Saleh, A.M. C3 and C4 plant systems respond differently to the concurrent challenges of mercuric oxide nanoparticles and future climate CO2. Sci. Total Environ. 2020, 749, 142356. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Hassan, Y.M.; Habeeb, T.H.; Alkhalaf, A.A.; Hozzein, W.N.; Selim, S.; AbdElgawad, H. Interactive effects of mercuric oxide nanoparticles and future climate CO2 on maize plant. J. Hazard. Mater. 2021, 401, 123849. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Chhikara, S.; Xing, B.; Musante, C.; White, J.C.; Dhankher, O.P. Physiological and Molecular Response of Arabidopsis thaliana (L.) to Nanoparticle Cerium and Indium Oxide Exposure. ACS Sustain. Chem. Eng. 2013, 1, 768–778. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Moerlein, S.M.; Welch, M.J. The chemistry of gallium and indium as related to radiopharmaceutical production. Int. J. Nucl. Med. Biol. 1981, 8, 277–287. [Google Scholar] [CrossRef]
- Panakkal, H.; Gupta, I.; Bhagat, R.; Ingle, A.P. Effects of Different Metal Oxide Nanoparticles on Plant Growth. In Nanotechnology in Plant Growth Promotion and Protection: Recent Advances and Impacts; John Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 259–282. [Google Scholar] [CrossRef]
- Souri, Z.; Cardoso, A.A.; Da-Silva, C.J.; De Oliveira, L.M.; Dari, B.; Sihi, D.; Karimi, N. Heavy metals and photosynthesis: Recent developments. In Photosynthesis, Productivity and Environmental Stress; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 107–134. [Google Scholar] [CrossRef]
- Thompson, M.; Gamage, D.; Hirotsu, N.; Martin, A.; Seneweera, S. Effects of elevated carbon dioxide on photosynthesis and carbon partitioning: A Perspective on root sugar sensing and hormonal crosstalk. Front. Physiol. 2017, 8, 578. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Trivedi, P.K. Glutathione S-transferases: Role in combating abiotic stresses including arsenic detoxification in plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Bhardwaj, R.; Handa, N.; Gautam, V.; Kohli, S.K.; Bali, S.; Kaur, P.; Thukral, A.K.; Arora, S.; Ohri, P.; et al. Responses of Phytochelatins and Metallothioneins in Alleviation of Heavy Metal Stress in Plants: An Overview. Plant Met. Interact. Emerg. Remediat. Tech. 2016, 263–283. [Google Scholar] [CrossRef]
- Schmoger, M.E.V.; Oven, M.; Grill, E. Detoxification of arsenic by phytochelatins in plants. Plant Physiol. 2000, 122, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Zimeri, A.M.; Dhankher, O.P.; McCaig, B.; Meagher, R.B. The plant MT1 metallothioneins are stabilized by binding cadmium and are required for cadmium tolerance and accumulation. Plant Mol. Biol. 2005, 58, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Sun, S.; Yan, Y.; Jing, X.; Shi, Q. Glutathione Metabolism and Its Function in Higher Plants Adapting to Stress. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 181–205. ISBN 9783319750880. [Google Scholar]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.H.M.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt-and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef] [PubMed]
- Shabbaj, I.; Madany, M.M.Y.; Tammar, A.; Balkhyour, M.A.; AbdElgawad, H. Silicon dioxide nanoparticles orchestrate carbon and nitrogen metabolism in pea seedlings to cope with broomrape infection. Environ. Sci. Nano 2021, 8, 1960–1977. [Google Scholar] [CrossRef]
- Fraser, C.M.; Chapple, C. The Phenylpropanoid Pathway in Arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanoca, A.; Kovinich, N.; Burkel, B.; Stecha, S.; Bohorquez-Restrepo, A.; Ueda, T.; Eliceiri, K.W.; Grotewold, E.; Otegui, M.S. Anthocyanin Vacuolar Inclusions Form by a Microautophagy Mechanism. Plant Cell 2015, 27, 2545–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.P.; Dong, X.J.; Ma, H.H. Antioxidative and Chelating Properties of Anthocyanins in Azolla imbricata Induced by Cadmium. Polish J. Environ. Stud. 2012, 21, 837–844. [Google Scholar]
- Ai, T.N.; Naing, A.H.; Yun, B.-W.; Lim, S.H.; Kim, C.K. Overexpression of RsMYB1 Enhances Anthocyanin Accumulation and Heavy Metal Stress Tolerance in Transgenic Petunia. Front. Plant Sci. 2018, 9, 1388. [Google Scholar] [CrossRef] [Green Version]
- Ghasemzadeh, A.; Jaafar, H.Z.; Karimi, E.; Ibrahim, M.H. Combined effect of CO2 enrichment and foliar application of salicylic acid on the production and antioxidant activities of anthocyanin, flavonoids and isoflavonoids from ginger. BMC Complement. Altern. Med. 2012, 12, 229. [Google Scholar] [CrossRef] [Green Version]
- Van Camp, W.; Capiau, K.; Van Montagu, M.; Inzé, D.; Slooten, L. Enhancement of oxidative stress tolerance in transgenic tobacco plants overproducing Fe-superoxide dismutase in chloroplasts. Plant Physiol. 1996, 112, 1703–1714. [Google Scholar] [CrossRef] [Green Version]
- Slooten, L.; Capiau, K.; Van Camp, W.; Van Montagu, M.; Sybesma, C.; Inzé, D. Factors Affecting the Enhancement of Oxidative Stress Tolerance in Transgenic Tobacco Overexpressing Manganese Superoxide Dismutase in the Chloroplasts. Plant Physiol. 1995, 107, 737–750. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C3-Plants | ||||||||
|---|---|---|---|---|---|---|---|---|
| Old leaves | Young leaves | Old leaves | Young leaves | |||||
| aCO2 | aCO2 + In2O3-NPs | aCO2 | aCO2 + In2O3-NPs | eCO2 | eCO2 + In2O3-NPs | eCO2 | eCO2 + In2O3-NPs | |
| In | 0 ± 0a | 240 ± 12d | 0 ± 0a | 193 ± 5c | 0 ± 0a | 171 ± 7bc | 0 ± 0a | 153 ± 4b |
| P | 4.96 ± 0.6c | 2.0 ± 0.12b | 3.12 ± 0.1c | 1.19 ± 0.06a | 5.67 ± 0.5 | 3.8 ± 0.1bc | 3.67 ± 0.1cd | 2.25 ± 0.06b |
| Fe | 1.26 ± 0.1bc | 0.65 ± 0.02ab | 1.02 ± 0.1c | 0.47 ± 0.06a | 1.27 ± 0.5 | 0.88 ± 0.1bb | 1.17 ± 0.1c | 0.75 ± 0.06b |
| C4-Plants | ||||||||
| Old leaves | Young leaves | Old leaves | Young leaves | |||||
| aCO2 | aCO2 + In2O3-NPs | aCO2 | aCO2 + In2O3-NPs | eCO2 | eCO2 + In2O3-NPs | eCO2 | eCO2 + In2O3-NPs | |
| In | 0 ± 0a | 411 ± 22e | 0 ± 0a | 226 ± 8c | 0 ± 0 | 295 ± 14d | 0 ± 0a | 179 ± 3b |
| P | 5.46 ± 0.19d | 3.14 ± 0.1b | 3.41 ± 0.1c | 1.96 ± 0.06a | 5.86 ± 0.7d | 4 ± 0.3c | 4.03 ± 0.06 | 2.8 ± 0.04ab |
| Fe | 1.51 ± 0.1c | 0.92 ± 0.02ab | 1.31 ± 0.1b | 0.64 ± 0.03a | 1.42 ± 0.7d | 1.2 ± 0.13b | 1.33 ± 0.01c | 0.87 ± 0.04bc |
| C3 Plants | C4 Plants | |||||||
|---|---|---|---|---|---|---|---|---|
| Old Leaves | Young Leaves | Old Leaves | Young Leaves | |||||
| In | 0 ± 0a | 387 ± 9.3d | 0 ± 0a | 215.5 ± 11c | 0 ± 0a | 352 ± 8.5d | 0 ± 0a | 178 ± 3.3b |
| FW | 0.24 ± 0.05b | 0.13 ± 0.01a | 0.17 ± 0.02 | 0.1 ± 0.01a | 0.28 ± 0.01c | 0.18 ± 0.01ab | 0.22 ± 0.01 | 0.12 ± 0.01a |
| DW | 0.032 ± 0.003cd | 0.018 ± 0.003a | 0.022 ± 0.003b | 0.013 ± 0.001a | 0.038 ± 0.0d | 0.024 ± 0.002bc | 0.029 ± 0.002c | 0.017 ± 0b |
| Photo | 16.4 ± 0.5d | 6.8 ± 0.2a | 12.2 ± 0.4c | 4.5 ± 0.1a | 18 ± 0.3d | 8.5 ± 0.1b | 13 ± 0.3 | 5.8 ± 0.1a |
| SOD | 122 ± 3.2a | 157 ± 1.9b | 156 ± 2.6b | 201 ± 5.1c | 145.4 ± 4.2b | 187 ± 2.9c | 179.1 ± 2c | 224 ± 5.9c |
| MDA | 6.4 ± 0.2ab | 9.7 ± 0.9d | 5.6 ± 0.4A | 7.8 ± 0.1c | 5.9 ± 0.2a | 7.1 ± 0.9b | 5.2 ± 0.3A | 6.7 ± 0.9ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shabbaj, I.I.; AbdElgawad, H.; Balkhyour, M.A.; Tammar, A.; Madany, M.M.Y. Elevated CO2 Differentially Mitigated Oxidative Stress Induced by Indium Oxide Nanoparticles in Young and Old Leaves of C3 and C4 Crops. Antioxidants 2022, 11, 308. https://doi.org/10.3390/antiox11020308
Shabbaj II, AbdElgawad H, Balkhyour MA, Tammar A, Madany MMY. Elevated CO2 Differentially Mitigated Oxidative Stress Induced by Indium Oxide Nanoparticles in Young and Old Leaves of C3 and C4 Crops. Antioxidants. 2022; 11(2):308. https://doi.org/10.3390/antiox11020308
Chicago/Turabian StyleShabbaj, Ibrahim I., Hamada AbdElgawad, Mansour A. Balkhyour, Abdurazag Tammar, and Mahmoud M. Y. Madany. 2022. "Elevated CO2 Differentially Mitigated Oxidative Stress Induced by Indium Oxide Nanoparticles in Young and Old Leaves of C3 and C4 Crops" Antioxidants 11, no. 2: 308. https://doi.org/10.3390/antiox11020308