
Lemongrass Essential Oil Components with Antimicrobial and Anticancer Activities

 ,
, 
 ,
,  , , and
, , and
Abstract
:1. Introduction
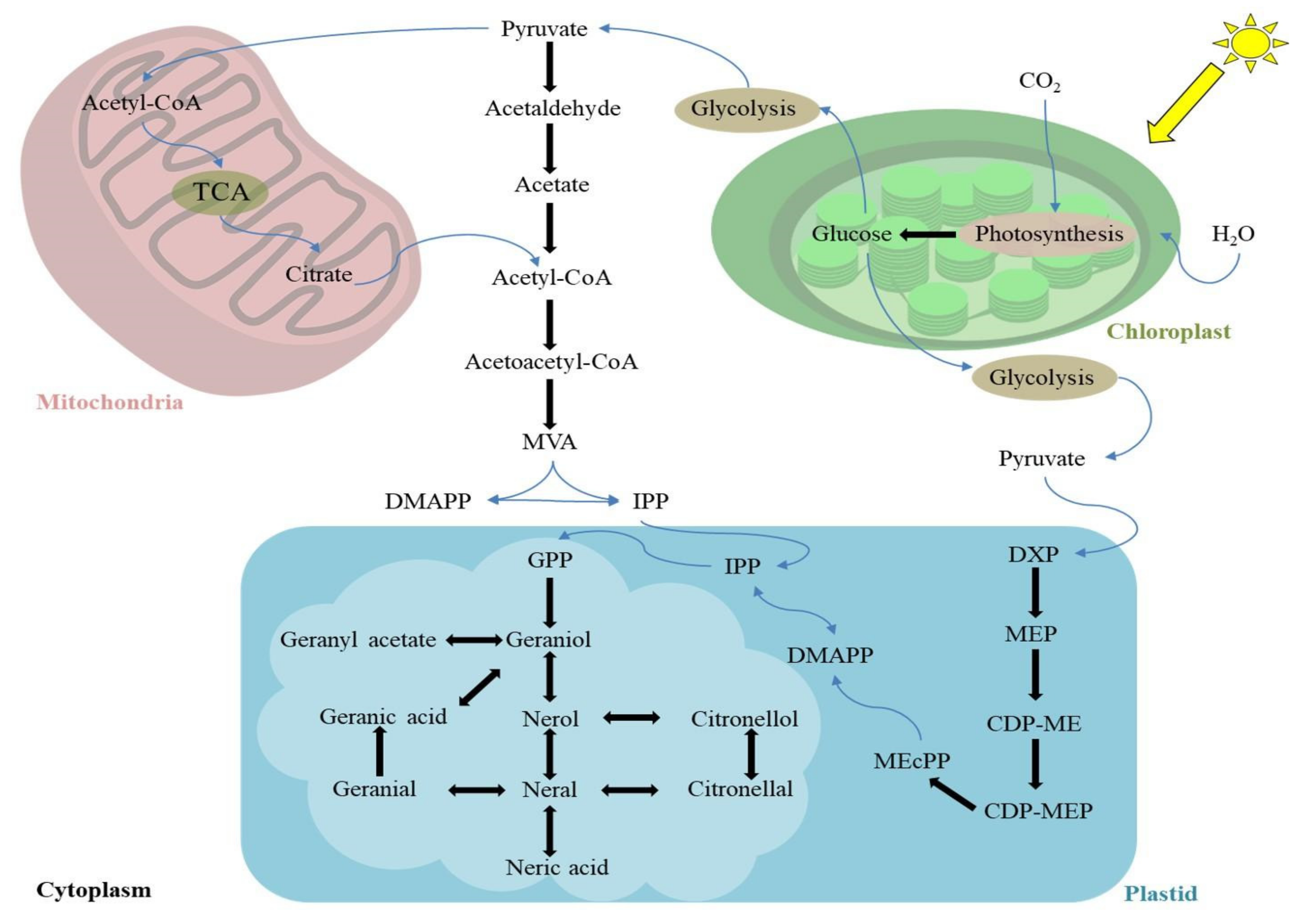
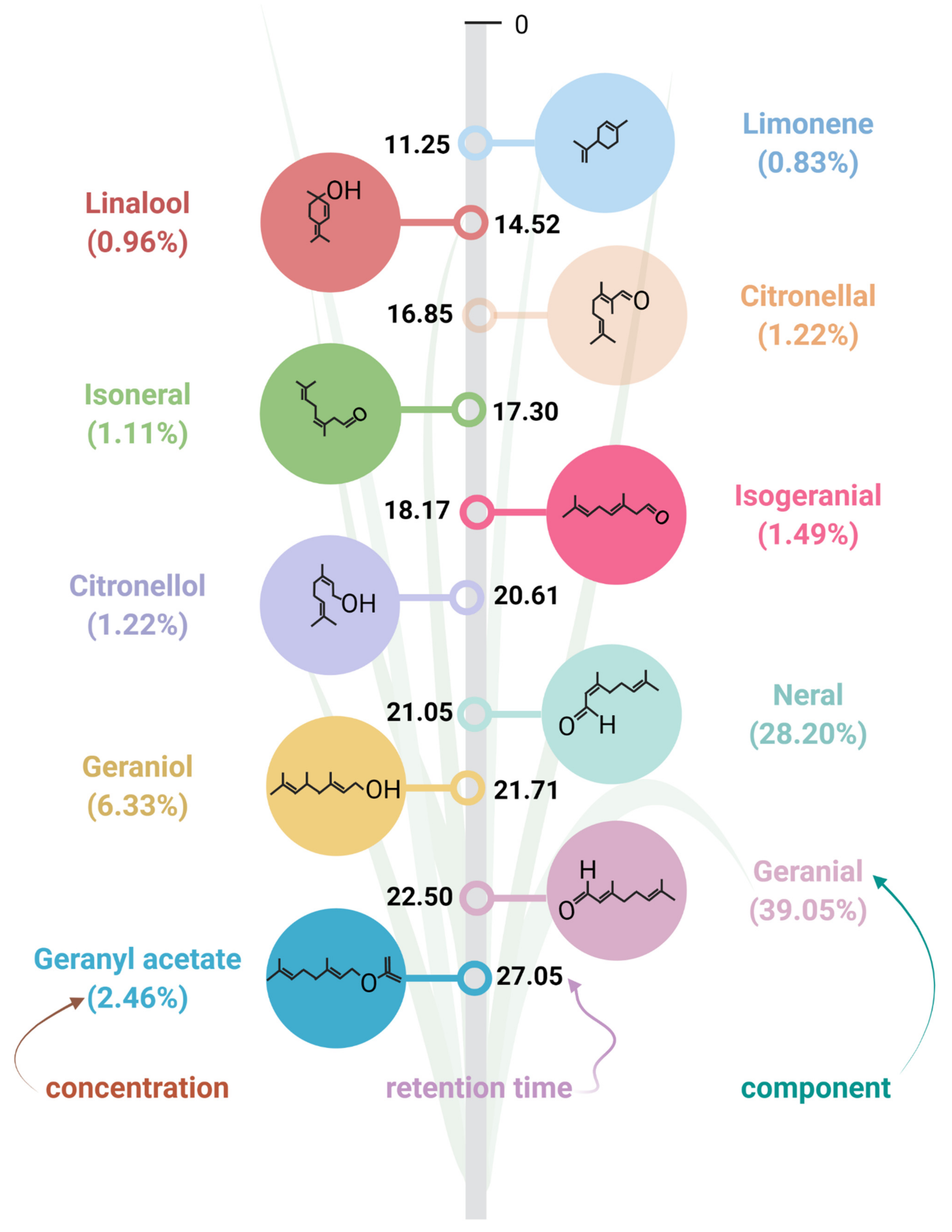
2. LEO: Biosynthesis and Chemical Composition
3. LEO Therapeutics
3.1. Antimicrobial Potential
3.1.1. Antibacterial Activity
3.1.2. Antifungal Activity
3.1.3. Antiviral Activity
3.2. Antioxidant-Related Effects
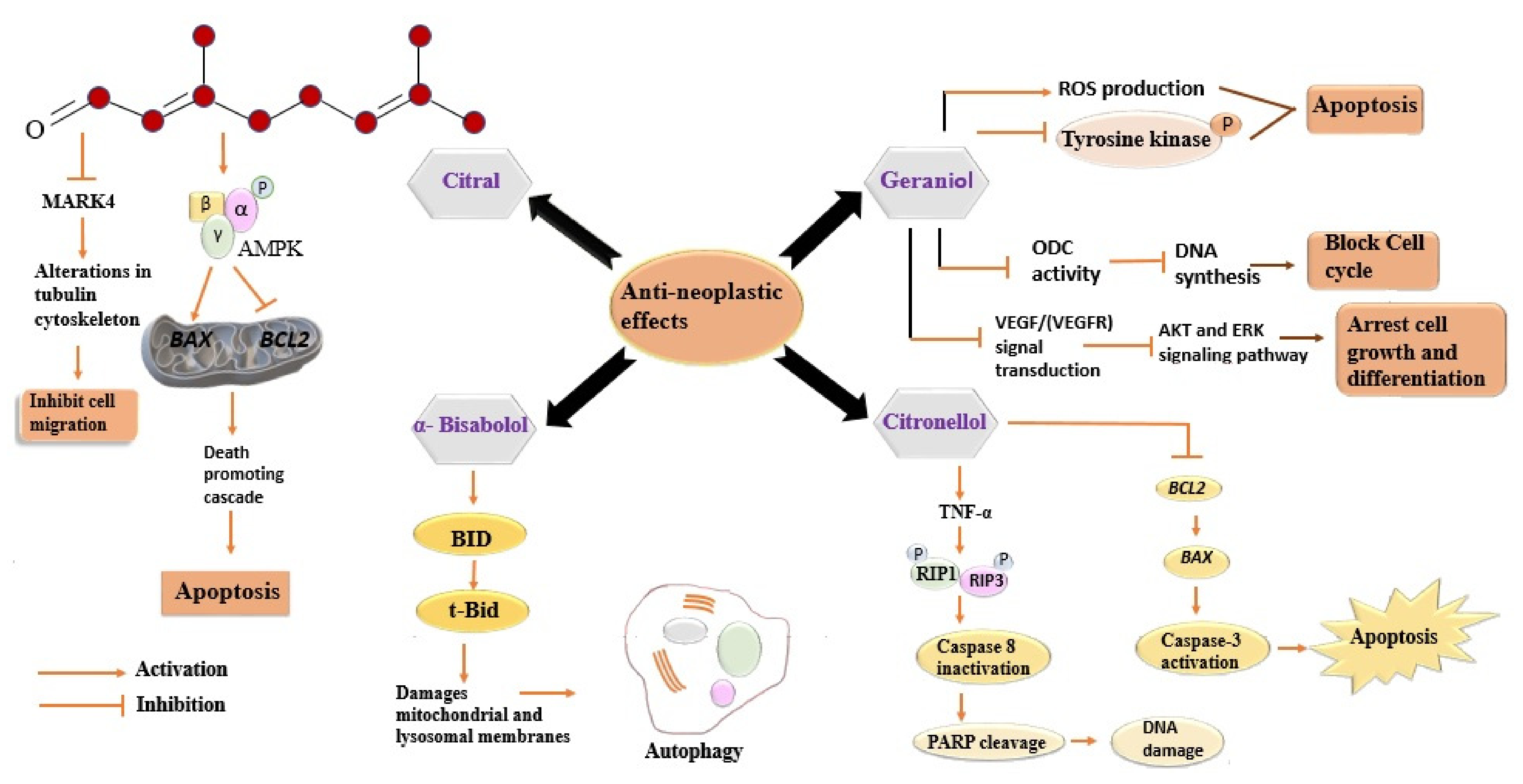
3.3. Anticancer Activity
3.4. Miscellaneous
4. Perspectives
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shah, G.; Shri, R.; Panchal, V.; Sharma, N.; Singh, B.; Mann, S. Scientific basis for the therapeutic use of Cymbopogon citratus, stapf (Lemon grass). J. Adv. Pharm. Technol. Res. 2011, 2, 3–8. [Google Scholar] [CrossRef]
- Srivastava, V.; Dubey, S.; Mishra, A. A Review on Lemon Grass: Agricultural and Medicinal Aspect. Int. Res. J. Pharm. 2013, 4, 42–44. [Google Scholar] [CrossRef]
- Mukarram, M.; Khan, M.M.A.; Corpas, F.J. Silicon nanoparticles elicit an increase in lemongrass (Cymbopogon flexuosus (Steud.) Wats) agronomic parameters with a higher essential oil yield. J. Hazard. Mater. 2021, 412, 125254. [Google Scholar] [CrossRef]
- Mukarram, M.; Khan, M.; Uddin, M.; Corpas, F.J. Irradiated chitosan (ICH): An alternative tool to increase essential oil content in lemongrass (Cymbopogon flexuosus). Acta. Physiol. Plant. 2022, 44, 2. [Google Scholar] [CrossRef]
- Joy, P.P.; Skaria, B.P.; Mathew, S.; Mathew, G.; Joseph, A.; Sreevidya, P.P. Lemongrass. Ind. J. Arecanut Spices Medicin. Plants 2006, 2, 55–64. [Google Scholar]
- Gaba, J.; Bhardwaj, G.; Sharma, A. Lemongrass. In Antioxidants in Vegetables and Nuts—Properties and Health Benefits; Nayik, G.S., Gull, A., Eds.; Springer: Singapore, 2020; pp. 75–103. [Google Scholar]
- Khandro, A.; Dhari, A.M.; Panwar, A.; Naqvi, B.; Hussain, G.; Quaisar, S.; Abbas, S. Determination of diferent trace and essential element in lemon grass samples by x-ray fluorescence spectroscopy technique. Int. Food Res. J. 2011, 18, 265–270. [Google Scholar]
- Haque, A.N.M.A.; Remadevi, R.; Naebe, M. Lemongrass (Cymbopogon): A review on its structure, properties, applications and recent developments. Cellulose 2018, 25, 5455–5477. [Google Scholar] [CrossRef]
- Skaria, B.P.; Joy, P.P.; Mathew, S.; Mathew, G. Lemongrass. In Handbook of Herbs and Spices; Peter, K.V., Ed.; CRC Press: Boca Raton, FL, USA, 2006; Volume 3, pp. 400–419. ISBN 9781845691714. [Google Scholar]
- Singh, K.M.; Jha, A. Medicinal and Aromatic Plants Cultivation in Bihar. India: Economic Potential and Condition for Adoption. 2008. Available online: https://mpra.ub.uni-muenchen.de/47091/ (accessed on 15 December 2021). [CrossRef] [Green Version]
- Gupta, A.K.; Ganjewala, D. A study on biosynthesis of “citral” in lemongrass (Cymbopogon flexuosus) cv. Suarna. Acta Physiol. 2015, 37, 240. [Google Scholar] [CrossRef]
- Mukarram, M.; Khan, M.M.A.; Zehra, A.; Choudhary, S.; Naeem, M.; Aftab, T. Biosynthesis of Lemongrass Essential Oil and the Underlying Mechanism for Its Insecticidal Activity. In Medicinal and Aromatic Plants Healhcare and Indusrial Applications; Aftab, T., Hakeem, K.R., Eds.; Springer: Switzerland, 2021; pp. 429–443. [Google Scholar]
- Ganjewala, D.; Luthra, R. Essential oil biosynthesis and regulation in the genus Cymbopogon. Nat. Prod. Commun. 2010, 5, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S. Lemon grass. Int. J. Pharm. Sci. Rev. Res. 2015, 35, 162–167. [Google Scholar]
- Hanaa, A.M.; Sallam, Y.I.; El-Leithy, A.S.; Aly, S.E. Lemongrass (Cymbopogon citratus) essential oil as affected by drying methods. Ann. Agric. Sci. 2012, 57, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Muturi, E.J.; Selling, G.W.; Doll, K.M.; Hay, W.T.; Ramirez, J.L. Leptospermum scoparium essential oil is a promising source of mosquito larvicide and its toxicity is enhanced by a biobased emulsifier. PLoS ONE 2020, 15, e0229076. [Google Scholar]
- Abdulazeez, M.A.; Abdullahi, A.S.; James, B.D. Lemongrass (Cymbopogon spp.) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: Oxford, UK, 2016; pp. 509–516. [Google Scholar]
- Meena, S.; Kumar, S.R.; Rao, D.K.V.; Dwivedi, V.; Shilpashree, H.B.; Rastogi, S.; Shasany, A.K.; Nagegowda, D.A. De Novo sequencing and analysis of lemongrass transcriptome provide first insights into the essential oil biosynthesis of aromatic grasses. Front. Plant Sci. 2016, 7, 1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, E. The essential oils: History, origin in plants, production. Analysis 1972, 1, 147–151. [Google Scholar]
- Sharma, P.R.; Mondhe, D.M.; Muthiah, S.; Pal, H.C.; Shahi, A.K.; Saxena, A.K.; Qazi, G.N. Anticancer activity of an essential oil from Cymbopogon flexuosus. Chem. Biol. Interact. 2009, 179, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Shendurse, A.M.; Sangwan, R.B.; Amit Kumar, R.V.; Patel, A.C.; Gopikrishna, G.; Roy, S.K. Phytochemical screening and antibacterial activity of lemongrass (Cymbopogon citratus) leaves essential oil. J. Pharmacogn. Phytochem. 2021, 10, 445–449. [Google Scholar]
- Rossi, G.G.; Guterres, K.B.; Bonez, P.C.; Gundel, S.S.; Aggertt, V.A.; Siqueira, F.S.; Iurique, A.F.; Wagnerd, R.; Klein, B.; Santos, R.C.V.; et al. Antibiofilm activity of nanoemulsions of Cymbopogon flexuosus against rapidly growing mycobacteria. Microb. Pathog. 2017, 113, 335–341. [Google Scholar] [CrossRef]
- Silva, N.B.; Rangel, M.L.; Castro, R.D.; Lima, J.M.; Castellano, L.R.C.; Valença, A.M.G.; Cavalcanti, A.L.; Almeida, R.F.C.; Pessoa, J.; Padilha, W.N. Anti-Biofilm and Hemolytic Effects of Cymbopogon citratus (Dc) Stapf Essential Oil. Pesqui. Bras. Odontoped. Clin. Integr. 2020, 19, 5011. [Google Scholar] [CrossRef]
- Gao, S.; Liu, G.; Li, J.; Chen, J.; Li, L.; Li, Z.; Zhang, X.; Zhang, S.; Thorne, R.F.; Zhang, S. Antimicrobial Activity of Lemongrass Essential Oil (Cymbopogon flexuosus) and Its Active Component Citral Against Dual-Species Biofilms of Staphylococcus. Front. Cell. Infect. Microbiol. 2020, 10, 603858. [Google Scholar] [CrossRef] [PubMed]
- Manvitha, K.; Bidya, B. Review on pharmacological activity of Cymbopogon citratus. Int. J. Herb. Med. 2014, 1, 5–7. [Google Scholar]
- Olorunnisola, S.K.; Asiyanbi, H.T.; Hammed, A.M.; Simsek, S. Biological properties of lemongrass: An overview. Int. Food Res. J. 2014, 21, 455–462. [Google Scholar]
- Ekpenyong, C.E.; Akpan, E.; Nyoh, A. Ethnopharmacology, phytochemistry, and biological activities of Cymbopogon citratus (DC.) Stapf extracts. Chin. J. Nat. Med. 2015, 13, 321–337. [Google Scholar] [CrossRef]
- Adukwu, E.C.; Bowles, M.; Edwards-Jones, V.; Bone, H. Antimicrobial activity, cytotoxicity and chemical analysis of lemongrass essential oil (Cymbopogon flexuosus) and pure citral. Appl. Microbiol. Biotechnol. 2016, 100, 9619–9627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjilouka, A.; Mavrogiannis, G.; Mallouchos, A.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Effect of lemongrass essential oil on Listeria monocytogenes gene expression. LWT—Food Sci. Technol. 2017, 77, 510–516. [Google Scholar] [CrossRef]
- Li, M.; Liu, B.; Bernigaud, C.; Fischer, K.; Guillot, J.; Fang, F. Lemongrass (Cymbopogon citratus) Oil: A promising miticidal and ovicidal agent against Sarcoptes scabiei. PLoS Negl. Trop. Dis. 2020, 14, e0008225. [Google Scholar] [CrossRef]
- Hacke, A.C.M.; Miyoshi, E.; Marques, J.A.; Pereira, R.P. Anxiolytic properties of Cymbopogon citratus (DC.) stapf extract, essential oil and its constituents in zebrafish (Danio rerio). J. Ethnopharmacol. 2020, 260, 113036. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Seo, S.M.; Huh, M.J.; Lee, S.C.; Park, I.K. Reactive oxygen species mediated-antifungal activity of cinnamon bark (Cinnamomum verum) and lemongrass (Cymbopogon citratus) essential oils and their constituents against two phytopathogenic fungi. Pestic. Biochem. Phys. 2020, 168, 104644. [Google Scholar] [CrossRef]
- Avoseh, O.; Oyedeji, O.; Rungqu, P.; Nkeh-Chungag, B.; Oyedeji, A. Cymbopogon species; ethnopharmacology, phytochemistry and the pharmacological importance. Molecules 2015, 20, 7438–7453. [Google Scholar] [CrossRef]
- Naik, M.I.; Fomda, B.A.; Jaykumar, E.; Bhat, J.A. Antibacterial activity of lemongrass (Cymbopogon citratus) oil against some selected pathogenic bacterias. Asian Pac. J. Trop. Med. 2010, 3, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Farias, P.K.S.; Lopes Silva, J.C.R.; de Souza, C.N.; da Fonseca, F.S.A.; Brandi, I.V.; Martins, E.R.; Azevedo, A.M.; de Almeida, A.C. Antioxidant activity of essential oils from condiment plants and their effect on lactic cultures and pathogenic bacteria. Cienc. Rural 2019, 49. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zhong, S.; Schwarz, P.; Chen, B.; Rao, J. Physical properties, antifungal and mycotoxin inhibitory activities of five essential oil nanoemulsions: Impact of oil compositions and processing parameters. Food Chem. 2019, 291, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Minami, M.; Kita, M.; Nakaya, T.; Yamamoto, T.; Kuriyama, H.; Imanishi, J. The inhibitory effect of essential oils on herpes simplex virus type-1 replication in vitro. Microbiol. Immunol. 2003, 47, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.R.; Yadav, K.; Khursheed, A.; Rather, M.A. An updated and comprehensive review of the antiviral potential of essential oils and their chemical constituents with special focus on their mechanism of action against various influenza and coronaviruses. Microb. Pathog. 2020, 104620. [Google Scholar] [CrossRef] [PubMed]
- Zulfa, Z.; Chia, C.T.; Rukayadi, Y. In vitro antimicrobial activity of Cymbopogon citratus (lemongrass) extracts against selected foodborne pathogens. Int. Food Res. J. 2016, 23, 1262. [Google Scholar]
- Kalemba, D.; Kunicka, A. Antibacterial and Antifungal Properties of Essential Oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]
- Alagawany, M.; El-Saadony, M.T.; Elnesr, S.S.; Farahat, M.; Attia, G.; Madkour, M.; Reda, F.M. Use of lemongrass essential oil as a feed additive in quail’s nutrition: Its effect on growth, carcass, blood biochemistry, antioxidant and immunological indices digestive enzymes and intestinal microbiota. Poult. Sci. 2021, 100, 101172. [Google Scholar] [CrossRef]
- Sharma, S.; Habib, S.; Sahu, D.; Gupta, J. Chemical properties and therapeutic potential of citral, a monoterpene isolated from lemongrass. Med. Chem. 2019, 17, 2–12. [Google Scholar] [CrossRef]
- Singh, B.R.; Singh, R.K. Antimicrobial activity of lemongrass (Cymbopogon citratus) oil against microbes of environmental, clinical and food origin. Int. Res. J. Pharm. Pharmacol. 2011, 1, 228–236. [Google Scholar]
- Lu, M.; Han, Z.; Xu, Y.; Yao, L. In Vitro and In Vivo Anti-Tobacco Mosaic Virus Activities of Essential Oils and Individual Compounds. J. Microbiol. Biotechnol. 2013, 23, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Negrelle, R.R.B.; Gomes, E.C. Cymbopogon citratus (DC.) Stapf: Chemical composition and biological activities. Rev. Bras. Pl. Med. 2007, 9, 80–92. [Google Scholar]
- Boukhatem, M.N.; Kameli, A.; Ferhat, M.A.; Saidi, F.; Tayebi, K. The food preservative potential of essential oils: Is lemongrass the answer? J. Fur Verbrauch. Leb. 2014, 9, 13–21. [Google Scholar] [CrossRef]
- Gill, A.O.; Holley, R.A. Mechanisms of bactericidal action of cinnamaldehyde against Listeria monocytogenes and of eugenol against L. monocytogenes and Lactobacillus sakei. Appl. Environ. Microbiol. 2004, 70, 5750–5755. [Google Scholar] [CrossRef] [Green Version]
- Viktorová, J.; Stupák, M.; Řehořová, K.; Dobiasová, S.; Hoang, L.; Hajšlová, J.; Thanh, T.V.; Tri, L.V.; Tuan, N.V.; Ruml, T. Lemon Grass Essential Oil Does not Modulate Cancer Cells Multidrug Resistance by Citral-Its Dominant and Strongly Antimicrobial Compound. Foods 2020, 9, 585. [Google Scholar] [CrossRef]
- Vazirian, M.; Kashani, S.T.; Ardekani, M.R.S.; Khanavi, M.; Jamalifar, H.; Fazeli, M.R.; Toosi, A.N. Antimicrobial activity of lemongrass (Cymbopogon citratus (DC) Stapf.) essential oil against food-borne pathogens added to cream-filled cakes and pastries. J. Essent. Oil Res. 2012, 24, 579–582. [Google Scholar] [CrossRef]
- Cai, Z.; Remadevi, R.; Al Faruque, M.; Setty, M.; Fan, L.; Haque, A.N.M.A.; Naebe, M. Fabrication of a cost-effective lemongrass (Cymbopogon citratus) membrane with antibacterial activity for dye removal. RSC Adv. 2019, 9, 34076–34085. [Google Scholar] [CrossRef] [Green Version]
- Raybaudi-Massilia, R.M.; Mosqueda-Melgar, J.; Martin-Belloso, O. Antimicrobial Activity of Essential Oils on Salmonella Enteritidis, Escherichia coli, and Listeria innocua in Fruit Juices. J. Food Prot. 2006. [Google Scholar] [CrossRef]
- Balakrishnan, B.; Paramasivam, S.; Arulkumar, A. Evaluation of the lemongrass plant (Cymbopogon citratus) extracted in different solvents for antioxidant and antibacterial activity against human pathogens. Asian Pac. J. Trop. Dis. 2014, 4. [Google Scholar] [CrossRef]
- Mishra, D.; Khare, P.; Singh, D.K.; Luqman, S.; Kumar, P.V.A.; Yadav, A.; Das, T.; Saikia, B.K. Retention of antibacterial and antioxidant properties of lemongrass oil loaded on cellulose nanofibre-poly ethylene glycol composite. Ind. Crop. Prod. 2018, 114, 68–80. [Google Scholar] [CrossRef]
- Moore-Neibel, K.; Gerber, C.; Patel, J.; Friedman, M.; Ravishankar, S. Antimicrobial activity of lemongrass oil against Salmonella enterica on organic leafy greens. J. Appl. Microbiol. 2012, 112, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Kotzekidou, P.; Giannakidis, P.; Boulamatsis, A. Antimicrobial activity of some plant extracts and essential oils against foodborne pathogens in vitro and on the fate of inoculated pathogens in chocolate. LWT Food Sci. Technol. 2008, 41, 119–127. [Google Scholar] [CrossRef]
- Aiemsaard, J.; Aiumlamai, S.; Aromdee, C.; Taweechaisupapong, S.; Khunkitti, W. The effect of lemongrass oil and its major components on clinical isolate mastitis pathogens and their mechanisms of action on Staphylococcus aureus DMST 4745. Res. Vet. Sci. 2011, 91, 31–37. [Google Scholar] [CrossRef]
- Adukwu, E.C.; Allen, S.C.; Phillips, C.A. The anti-biofilm activity of lemongrass (Cymbopogon flexuosus) and grapefruit (C itrus paradisi) essential oils against five strains of Staphylococcus aureus. J. Appl. Microbiol. 2012, 113, 1217–1227. [Google Scholar] [CrossRef]
- Takaisi-Kikuni, N.; Krüger, D.; Gnann, W.; Wecke, J. Microcalorimetric and electron microscopic investigation on the effects of essential oil from Cymbopogon densiflorus on Staphylococcus aureus. Microbios. 1996, 88, 55–62. [Google Scholar] [PubMed]
- Peichel, C.; Nair, D.V.T.; Dewi, G.; Donoghue, A.M.; Reed, K.M.; Johny, A.K. Effect of lemongrass (Cymbopogon citratus) essential oil on the survival of multidrug-resistant Salmonella enterica serovar Heidelberg in contaminated poultry drinking water. J. Appl. Poult. Res. 2019, 28, 1121–1130. [Google Scholar] [CrossRef]
- Dewi, G.; Nair, D.V.T.; Peichel, C.; Johnson, T.J.; Noll, S.; Johny, A.K. Effect of lemongrass essential oil against multidrug-resistant Salmonella Heidelberg and its attachment to chicken skin and meat. Poult. Sci. 2021, 101116. [Google Scholar] [CrossRef]
- Hassoun, A.; Emir Çoban, Ö. Essential oils for antimicrobial and antioxidant applications in fish and other seafood products. Trends Food Sci. Technol. 2017, 68, 26–36. [Google Scholar] [CrossRef]
- Swamy, M.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Alternat. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef] [PubMed]
- Devi, M.A.; Sahoo, D.; Singh, T.B.; Rajashekar, Y. Toxicity, repellency and chemical composition of essential oils from Cymbopogon species against red flour beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). J. fur Verbraucherschutz Leb. 2020, 15, 181–191. [Google Scholar] [CrossRef]
- Costa, K.A.D.; Moura, R.; Millezi, A.F. Antimicrobial and antibiofilm activity of Cymbopogon flexuosus essential oil microemulsions. Rev. Ceres 2019, 66, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Song, K.; Zhang, X.; Sun, Y.; Sui, Y.; Chen, Y.; Jia, Z.; Sun, H.; Sun, Z.; Xia, X. Antimicrobial Activity and Possible Mechanism of Action of Citral against Cronobacter sakazakii. PLoS ONE 2016, 11, e0159006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassolé, I.H.N.; Lamien-Meda, A.; Bayala, B.; Obame, O.L.C.; Ilboudo, A.J.; Franz, C.; Novak, J.; Nebié, R.C.; Dicko, M.H. Chemical composition and antimicrobial activity of Cymbopogon citratus and Cymbopogon giganteus essential oils alone and in combination. Phytomedicine 2011, 18, 1070–1074. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Application of edible coating with essential oil in food preservation. Crit. Rev. Food Sci. Nutr. 2019, 59, 2467–2480. [Google Scholar] [CrossRef] [PubMed]
- Rkin, R.; Korukluoglu, M. Effectiveness of Cymbopogon citratus l. essential oil to inhibit the growth of some filamentous fungi and yeasts. J. Med. Food. 2009, 12, 193–197. [Google Scholar] [CrossRef]
- Tzortzakis, N.G.; Economakis, C.D. Antifungal activity of lemongrass (Cympopogon citratus L.) essential oil against key postharvest pathogens. Innov. Food Sci. Emerg. Technol. 2007, 8, 253–258. [Google Scholar] [CrossRef]
- Dong, L.M.; Thuy, D.T.K. Evaluation of the synergistic effect of ethanol and lemongrass oil against Aspergillus niger. J. Microbiol. Biotech. Food Sci. 2021, 1312–1316. [Google Scholar] [CrossRef]
- Abd-El Fattah, S.M.; Yahia Hassan, A.; Bayoum, H.M.; Eissa, H.A. The use of lemongrass extracts as antimicrobial and food additive potential in yoghurt. J. Am. Sci. 2010, 6, 582–594. [Google Scholar]
- Júnior, R.C.; Capucho, E.; Garcia, T.M.; Del Valle, T.A.; Campana, M.; Zilio, E.M.C.; Azevedo, E.B.; Morais, J.P.G. Lemongrass essential oil in sugarcane silage: Fermentative profile, losses, chemical composition, and aerobic stability. Anim. Feed Sci. Tech. 2020, 260, 114371. [Google Scholar] [CrossRef]
- Helal, G.A.; Sarhan, M.M.; Shahla, A.N.K.A.; Abou El-Khair, E.K. Effects of Cymbopogon citratus L. essential oil on the growth, morphogenesis and aflatoxin production of Aspergillus flavus ML2-strain. J. Basic Microbiol. 2007, 47, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Alviano, W.S.; Mendonça-Filho, R.R.; Alviano, D.S.; Bizzo, H.R.; Souto-Padrón, T.; Rodrigues, M.L.; Bolognese, A.M.; Alviano, C.S.; Souza, M.M.G. Antimicrobial activity of Croton cajucara Benth linalool-rich essential oil on artificial biofilms and planktonic microorganisms. Oral Microbiol. Immunol. 2005, 20, 101–105. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.d.B.; Guterres, S.S.; Weisheimer, V.; Schapoval, E.E.S. Antifungal activity of the lemongrass oil and citral against Candida spp. Brazilian J. Infect. Dis. 2008, 12, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Tchinda, E.S.; Jazet, P.M.D.; Tatsadjieu, L.N.; Ndongson, B.D.; Amvam, P.H.Z.; Menut, C. Antifungal Activity of the Essential Oil of Cymbopogon citratus (Poaceae) Against Phaeoramularia angolensis. J. Essent. Oil Bear. Plants 2009, 12, 218–224. [Google Scholar] [CrossRef]
- Mishra, A.K.; Dubey, N.K. Evaluation of some essential oils for their toxicity against fungi causing deterioration of stored food commodities. Appl. Environ. Microbiol. 1994, 60, 1101–1105. [Google Scholar] [CrossRef] [Green Version]
- Masniyom, P.; Benjama, O.; Maneesri, J. Effect of turmeric and lemongrass essential oils and their mixture on quality changes of refrigerated green mussel (Perna viridis). Int. J. Food Sci. Technol. 2012, 47, 1079–1085. [Google Scholar] [CrossRef]
- Jayasena, D.D.; Jo, C. Essential oils as potential antimicrobial agents in meat and meat products: A review. Trends Food Sci. Technol. 2013, 34, 96–108. [Google Scholar] [CrossRef]
- Oh, Y.A.; Oh, Y.J.; Song, A.Y.; Won, J.S.; Song, K.B.; Min, S.C. Comparison of effectiveness of edible coatings using emulsions containing lemongrass oil of different size droplets on grape berry safety and preservation. LWT 2017, 75, 742–750. [Google Scholar] [CrossRef]
- Muhammad, I.; Riffat, T.; Asif, J.; Atif, J.; Raja, M.U.; Anjum, M. Lemongrass essential oil as an alternate approach to manage seed associated fungi of wheat and rice. Int. J. Agric. Biol. 2017, 19, 1301–1306. [Google Scholar]
- Artiga-Artigas, M.; Acevedo-Fani, A.; Martin-Belloso, O. Improving the shelf life of low-fat cut cheese using nanoemulsion-based edible coatings containing oregano essential oil and mandarin fiber. Food Control 2017, 76, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Frazão, G.G.S.; Blank, A.F.; de Aquino Santana, L.C.L. Optimisation of edible chitosan coatings formulations incorporating Myrcia ovata Cambessedes essential oil with antimicrobial potential against foodborne bacteria and natural microflora of mangaba fruits. LWT Food Sci.Technol. 2017, 79, 1–10. [Google Scholar] [CrossRef]
- Silveira, S.M.D.; Júnior, A.C.; Scheuerman, G.N.; Secchi, F.L.; Vieira, C.R.W. Chemical composition and antimicrobial activity of essential oils from selected herbs cultivated in the South of Brazil against food spoilage and foodborne pathogens. Cienc. Rural. 2012, 42, 1300–1306. [Google Scholar] [CrossRef] [Green Version]
- Belewu, M.; Ahmed El-Imam, A.M.; Adeyemi, K.; Belewu, M.A.; Ahmed El-Imam, A.M.; Adeyemi, K.D.; Oladunjoye, S.A. Eucalyptus Oil and Lemon Grass Oil: Effect on Chemical Composition and Shelf-Life of Soft Cheese. Environ. Nat. Resour. Res. 2012, 2, 114–118. [Google Scholar] [CrossRef]
- Yousuf, B.; Srivastava, A.K. Flaxseed gum in combination with lemongrass essential oil as an effective edible coating for ready-to-eat pomegranate arils. Int. J. Biol. Macromol. 2017, 104, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Khan, A.L.; Waqas, M.; Lee, I.; Lee, I. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Feriotto, G.; Marchetti, N.; Costa, V.; Beninati, S.; Tagliati, F.; Mischiati, C. Chemical Composition of Essential Oils from Thymus vulgaris, Cymbopogon citratus, and Rosmarinus officinalis, and Their Effects on the HIV-1 Tat Protein Function. Chem. Biodivers. 2018, 15, e1700436. [Google Scholar] [CrossRef]
- Chiamenti, L.; Silva, F.P.D.; Schallemberger, K.; Demoliner, M.; Rigotto, C.; Fleck, J.D. Cytotoxicity and antiviral activity evaluation of Cymbopogon spp hydroethanolic extracts. Braz. J. Pharm. Sci. 2019, 55, e18063. [Google Scholar] [CrossRef]
- Asif, M.; Saleem, M.; Saadullah, M.; Yaseen, H.S.; Al Zarzour, R. COVID-19 and therapy with essential oils having antiviral, anti-inflammatory and immunomodulatory properties. Inflammopharmacology 2020, 28, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Mukarram, M.; Choudhary, S.; Kurjak, D.; Petek, A.; Khan, M.M.A. Drought: Sensing, signalling, effects and tolerance in higher plants. Physiol. Plant. 2021, 172, 1291–1300. [Google Scholar] [CrossRef]
- Anggraeni, N.I.; Hidayat, I.W.; Saadah, D.; Rachman, E. Bioactivity of essential oil from lemongrass (Cymbopogon citratus Stapf) as antioxidant agent. AIP Conf. Proc. 2018, 1927, 30007. [Google Scholar] [CrossRef]
- Karpagam, G.N.; Gayathri, R.; Vishnupriya, V. Bioactivity analysis of lemongrass oil. Asian J. Res. Chem. 2016, 9, 903. [Google Scholar] [CrossRef]
- Selim, S.A. Chemical composition, antioxidant and antimicrobial activity of the essential oil and methanol extract of the Egyptian lemongrass Cymbopogon proximus Stapf. Grasas Y Aceites 2011, 62, 55–61. [Google Scholar] [CrossRef]
- Mirghani, M.E.S.; Liyana, Y.; Parveen, J. Bioactivity analysis of lemongrass (Cymbopogan citratus) essential oil. Int. Food Res. J. 2012, 19, 569–575. [Google Scholar]
- Tiwari, M.; Dwivedi, U.N.; Kakkar, P. Suppression of oxidative stress and pro-inflammatory mediators by Cymbopogon citratus D. Stapf extract in lipopolysaccharide stimulated murine alveolar macrophages. Food Chem. Toxicol. 2010, 48, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Fayyaz, S.; Poltronieri, P.; Calin, G.; Mallardo, M. Epigenetic deregulation in cancer: Enzyme players and non-coding RNAs. Semin. Cancer Biol. 30 July epub ahead of print. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kapur, A.; Felder, M.; Fass, L.; Kaur, J.; Czarnecki, A.; Rathi, K.; Zeng, S.; Osowski, K.; Howell, C.; Xiong, M.; et al. Modulation of oxidative stress and subsequent induction of apoptosis and endoplasmic reticulum stress allows citral to decrease cancer cell proliferation. Sci. Rep. 2016, 6, 1–14. [Google Scholar]
- Ndhlala, A.R.; Moyo, M.; van Staden, J. Natural Antioxidants: Fascinating or Mythical Biomolecules? Molecules 2010, 15, 6905–6930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartatie, E.S.; Prihartini, I.; Widodo, W.; Wahyudi, A. Bioactive Compounds of Lemongrass (Cymbopogon citratus) essential oil from different parts of the plant and distillation methods as natural antioxidant in broiler meat. A. IOP Conf. Ser. Mater. Sci. Eng. 2019, 532, 012018. [Google Scholar] [CrossRef] [Green Version]
- Hedges, L.J.; Lister, C.E. Nutritional Attributes of Herbs. Crop & Food Research Confidential Report No. 1891; New Zealand Institute for Crop & Food Research Limited: Christchruch, New Zealand, 2007; pp. 1–85. [Google Scholar]
- Soares, M.O.; Vinha, A.F.; Barreira, S.V.P.; Coutinho, F.; Aires-Gonçalves, S.; Oliveira, M.B.P.P.; Pires, P.C.; Castro, A. Evaluation of Antioxidant and Antimicrobial Properties of the Angolan Cymbopogon Citratus Essential Oil With a View to Its Utilization as Food Biopreservative. J. Agric. Sci. 2013, 5, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Nguefack, J.; Nguikwie, S.K.; Fotio, D.; Dongmo, B.; Zollo, P.H.A.; Leth, V.; Nkengfack, A.E.; Poll, L. Fungicidal potential of essential oils and fractions from Cymbopogon citratus, Ocimum gratissimum and Thymus vulgaris to control Alternaria padwickii and Bipolaris oryzae, two seed-borne fungi of rice (Oryza sativa L.). J. Essent. Oil Res. 2007, 19, 581–587. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.V. Anticancer Activity of Natural Health Products (Dandelion Root, Lemongrass, and Hibiscus Extracts); A Study of Efficacy, Interaction, and Mechanism of Action. Master Thesis, Windsor University, Ontario, CA, USA, 2019. [Google Scholar]
- Parveen, A.; Akash, M.S.H.; Rehman, K.; Kyun, W.W. Anticancer activities of medicinal plants: Modulation of p53 expression and induction of apoptosis. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 257–271. [Google Scholar] [CrossRef]
- Manosroi, J.; Dhumtanom, P.; Manosroi, A. Anti-proliferative activity of essential oil extracted from Thai medicinal plants on KB and P388 cell lines. Cancer Lett. 2006, 235, 114–120. [Google Scholar] [CrossRef]
- Bailly, C. Targets and pathways involved in the antitumor activity of citral and its stereo-isomers. Eur. J. Pharmacol. 2020, 871, 172945. [Google Scholar] [CrossRef]
- Liu, Y.; Whelan, R.J.; Pattnaik, B.R.; Ludwig, K.; Subudhi, E.; Rowland, H.; Claussen, N.; Zucker, N.; Uppal, S.; Kushner, D.M.; et al. Terpenoids from Zingiber officinale (Ginger) Induce Apoptosis in Endometrial Cancer Cells through the Activation of p53. PLoS ONE 2012, 7, e53178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idrees, M.; Hakkim, F.L.; Naikoo, G.A.; Hassan, I.U. Recent advances in extraction, characterization, and potential use of citral. In Natural Bio-active Compounds; Aktar, M.S., Swami, M.K., Eds.; Springer Nature: Switzerland, 2019; Volume 3. [Google Scholar] [CrossRef]
- Halabi, M.; Sheikh, B. Anti-proliferative effect and phytochemical analysis of Cymbopogon citratus extract. Biomed. Res. Int. 2014, 2014, 906239. [Google Scholar] [CrossRef] [Green Version]
- Najar, B.; Shortrede, J.E.; Pistelli, L.; Buhagiar, J. Chemical Composition and in Vitro Cytotoxic Screening of Sixteen Commercial Essential Oils on Five Cancer Cell Lines. Chem. Biodivers. 2020, 17, e1900478. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.B.; Thakkar, V.R.; Patel, J.S. Cellular effect of curcumin and citral combination on breast cancer cells: Induction of apoptosis and cell cycle arrest. J. Breast Cancer 2015, 18, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudai, N.; Weinstein, Y.; Krup, M.; Rabinski, T.; Ofir, R. Citral is a new inducer of caspase-3 in tumor cell lines. Planta Med. 2005, 71, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Trang, D.T.; Hoang, T.K.V.; Nguyen, T.T.M.; Van Cuong, P.; Dang, N.H.; Dang, H.D.; Quang, T.N.; Dat, N.T. Essential Oils of Lemongrass (Cymbopogon citratus Stapf) Induces Apoptosis and Cell Cycle Arrest in A549 Lung Cancer Cells. Biomed. Res. Int. 2020, 2020, 5924856. [Google Scholar] [CrossRef] [Green Version]
- Harada, D.; Takigawa, N.; Kiura, K. The Role of STAT3 in Non-Small Cell Lung Cancer. Cancers 2014, 6, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.A. STAT3 as a central mediator of neoplastic cellular transformation. Cancer Lett. 2007, 251, 199–210. [Google Scholar] [CrossRef]
- Maruoka, T.; Kitanaka, A.; Kubota, Y.; Yamaoka, G.; Kameda, T.; Imataki, O.; Dobashi, H.; Bandoh, S.; Kadowaki, N.; Tanaka, T. Lemongrass essential oil and citral inhibit Src/Stat3 activity and suppress the proliferation/survival of small-cell lung cancer cells, alone or in combination with chemotherapeutic agents. Int. J. Oncol. 2018, 52, 1738–1748. [Google Scholar] [CrossRef] [Green Version]
- Balusamy, S.R.; Perumalsamy, H.; Veerappan, K.; Huq, M.; Rajeshkumar, S.; Lakshmi, T.; Kim, Y.J. Citral induced apoptosis through modulation of key genes involved in fatty acid biosynthesis in human prostate cancer cells: In silico and in vitro study. BioMed Res. Int. 2020, 2020, 6040727. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.L.; De Antueno, R.; Coyle, K.M.; Sultan, M.; Cruickshank, B.M.; Giacomantonia, M.A.; Giacomantonia, C.A.; Duncan, R.; Marcato, P. Citral reduces breast tumor growth by inhibiting the cancer stem cell marker ALDH1A3. Mol. Oncol. 2016, 10, 1485–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Gan, L.; Chen, Y.; Luo, D.; Zhang, Z.; Cao, W.; Zhou, Z.; Lin, X.; Sun, C. Mark4 promotes oxidative stress and inflammation via binding to PPARγ and activating NF-κB pathway in mice adipocytes. Sci. Rep. 2016, 6, 21382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Tian, L.; Gan, L.; Liu, Z.; Sun, C. Mark4 promotes adipogenesis and triggers apoptosis in 3T3-L1 adipocytes by activating JNK1 and inhibiting p38MAPK pathways. Biol. Cell 2014, 106, 294–307. [Google Scholar] [CrossRef]
- Naz, F.; Khan, F.I.; Mohammad, T.; Khan, P.; Manzoor, S.; Hasan, G.M.; Lobb, K.A.; Luqman, S.; Islam, A.; Ahmad, F.; et al. Investigation of molecular mechanism of recognition between citral and MARK4: A newer therapeutic approach to attenuate cancer cell progression. Int. J. Biol. Macromol. 2018, 107, 2580–2589. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, B.Y.; Sarker, M.M.R.; Kamarudin, M.N.A.; Mohan, G. Antiproliferative and apoptosis inducing effects of citral via p53 and ROS-induced mitochondrial-mediated apoptosis in human colorectal HCT116 and HT29 cell lines. Biomed. Pharmacother. 2017, 96, 834–846. [Google Scholar] [CrossRef]
- Sanches, L.J.; Marinello, P.C.; Panis, C.; Fagundes, T.R.; Morgada-Diaz, J.A.; de-Freitas-Junior, J.C.M.; Cecchini, R.; Cecchini, A.L.; Luiz, R.C. Cytotoxicity of citral against melanoma cells: The involvement of oxidative stress generation and cell growth protein reduction. Tumor Biol. 2017, 39, 1010428317695914. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Park, E.J.; Lee, C.R.; Chun, J.N.; Cho, N.H.; Lee, S.; Kim, T.W.; Park, H.H.; So, I.; Jeon, J.H. Geraniol induces cooperative interaction of apoptosis and autophagy to elicit cell death in PC-3 prostate cancer cells. Int. J. Oncol. 2012, 40, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Galle, M.; Crespo, R.; Kladniew, B.R.; Villegas, S.M.; Polo, M.; De Bravo, M.G. Suppression by Geraniol of the Growth of A549 Human Lung Adenocarcinoma Cells and Inhibition of the Mevalonate Pathway in Culture and In Vivo: Potential Use in Cancer Chemotherapy. Nutr. Cancer 2014, 66, 888–895. [Google Scholar] [CrossRef]
- Wittig, C.; Scheuer, C.; Parakenings, J.; Menger, M.D.; Laschke, M.W. Geraniol suppresses angiogenesis by downregulating vascular endothelial growth factor (VEGF)/VEGFR-2 signaling. PLoS ONE 2015, 10, e0131946. [Google Scholar] [CrossRef] [Green Version]
- Crespo, R.; Rodenak-Kladniew, B.E.; Castro, M.A.; Soberon, M.V.; Lavarias, S.M. Induction of oxidative stress as a possible mechanism by which geraniol affects the proliferation of human A549 and HepG2 tumor cells. Chem. Biol. Interact. 2020, 320, 109029. [Google Scholar] [CrossRef]
- Maczka, W.; Wínska, K.; Grabarczyk, M. One Hundred Faces of Geraniol. Molecules 2020, 25, 3303. [Google Scholar] [CrossRef] [PubMed]
- Murray-Stewart, T.R.; Woster, P.M.; Casero Jr, R.A. Targeting polyamine metabolism for cancer therapy and prevention. Biochem. J. 2016, 473, 2937–2953. [Google Scholar] [CrossRef] [Green Version]
- Fatima, K.; Wani, Z.A.; Meena, A.; Luqman, S. Geraniol exerts its antiproliferative action by modulating molecular targets in lung and skin carcinoma cells. Phytother. Res. 2021, 35, 3861–3874. [Google Scholar] [CrossRef] [PubMed]
- Madan, K.; Devaki, T. Geraniol, a component of plant essential oils-a review of its pharmacological activities. Int. J. Pharm. Pharm. Sci. 2015, 7, 67–70. [Google Scholar]
- Shen, X.; Cui, X.; Cui, H.; Jin, Y.; Jin, W.; Sun, H. Geraniol and lupeol inhibit growth and promote apoptosis in human hepatocarcinoma cells through the MAPK signaling pathway. J. Cell. Biochem. 2018, 120, 5033–5041. [Google Scholar] [CrossRef]
- Kuzu, B.; Cüce, G.; Çınar Ayan, İ.; Gültekin, B.; Tuba Canbaz, H.; Gül Dursun, H.; Şahin, Z.; Keskin, İ.; Serpil Kalkan, S.S. Evaluation of Apoptosis Pathway of Geraniol on Ishikawa Cells. Nutr. Cancer 2020. 22 September epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Shanmugapriya, S.; Subramanian, P.; Kanimozhi, S. Geraniol Inhibits Endometrial Carcinoma via Downregulating Oncogenes and Upregulating Tumour Suppressor Genes. Indian, J. Clin. Biochem. 2017, 32, 214–219. [Google Scholar] [CrossRef]
- Cho, M.; So, I.; Chun, J.N.; Jeon, J.H. The antitumor effects of geraniol: Modulation of cancer hallmark pathways (Review). Int. J. Oncol. 2016, 48, 1772–1782. [Google Scholar] [CrossRef] [Green Version]
- Hajizadeh, M.; Maleki, H.; Barani, M.; Fahmindehkar, M.A.; Mahmoodi, M.; Torkzadeh-Mahani, M. In vitro cytotoxicity assay of D-limonene niosomes: An efficient nano-carrier for enhancing solubility of plant-extracted agents. Res. Pharma. Sci. 2019, 14, 448. [Google Scholar]
- Jia, S.S.; Xi, G.P.; Zhang, M.; Chen, Y.B.; Lei, B.; Dong, X.S.; Yang, Y.M. Induction of apoptosis by D-limonene is mediated by inactivation of Akt in LS174T human colon cancer cells. Oncol. Rep. 2013, 29, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Liang, Z.; Mi, Q.; Guo, Y. Limonene terpenoid obstructs human bladder cancer cell (T24 cell line) growth by inducing cellular apoptosis, caspase activation, G2/M phase cell cycle arrest and stops cancer metastasis. J. BUON./Off. J. Balk. Union. Oncol. 2020, 25, 280–285. [Google Scholar]
- Jayaganesh, R.; Pugalendhi, P.; Murali, R. Effect of citronellol on NF-kB inflammatory signaling molecules in chemical carcinogen-induced mammary cancer in the rat model. J. Biochem. Mol. Toxicol. 2020, 34, e22441. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Lai, Y.J.; Ma, J.; Ho, C.T.; Hung, S.W.; Chen, Y.H.; Chen, C.T.; Kao, J.Y.; Wat, T.D. Citronellol induces necroptosis of human lung cancer cells via TNF-α pathway and reactive oxygen species accumulation. In Vivo 2019, 33, 1193–1201. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.; Suphrom, N.; Daowtak, K.; Potup, P.; Thongsri, Y.; Usuwanthim, K. Anticancer Effect of Citrus hystrix DC. Leaf Extract and Its Bioactive Constituents Citronellol and, Citronellal on the Triple Negative Breast Cancer MDA-MB-231 Cell Line. Pharmaceuticals 2020, 13, 476. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, J.; Pachaiappan, P.; Thangarasu, R. Citronellol, an Acyclic Monoterpene Induces Mitochondrial-Mediated Apoptosis through Activation of Proapoptotic Factors in MCF-7 and MDA-MB-231 Human Mammary Tumor Cells. Nutr. Cancer 2020, 73, 1448–1458. [Google Scholar] [CrossRef]
- Cavalieri, E.; Mariotto, S.; Fabrizi, C.; de Prati, A.C.; Gottardo, R.; Leone, S.; Berra, L.V.; Lauro, G.M.; Ciampa, A.R.; Suzuki, H. α-Bisabolol, a nontoxic natural compound, strongly induces apoptosis in glioma cells. Biochem. Biophys. Res. Commun. 2004, 315, 589–594. [Google Scholar] [CrossRef]
- Rigo, A.; Ferrarini, I.; Lorenzetto, E.; Darra, E.; Liparulo, I.; Bergamini, C.; Sissa, C.; Cavalieri, E.; Vinante, F. BID and the α-bisabolol-triggered cell death program: Converging on mitochondria and lysosomes. Cell. Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rigo, A.; Vinante, F. The antineoplastic agent α-bisabolol promotes cell death by inducing pores in mitochondria and lysosomes. Apoptosis 2016, 21, 917–927. [Google Scholar] [CrossRef]
- Uno, M.; Kokuryo, T.; Yokoyama, Y.; Senga, T.; Nagino, M. α-Bisabolol inhibits invasiveness and motility in pancreatic cancer through KISS1R activation. Anticancer Res. 2016, 36, 583–589. [Google Scholar]
- Fang, D.; Wang, H.; Li, M.; Wei, W. α-bisabolol enhances radiotherapy-induced apoptosis in endometrial cancer cells by reducing the effect of XIAP on inhibiting caspase-3. Biosci. Rep. 2019, 39, BSR20190696. [Google Scholar] [CrossRef] [Green Version]
- Balusamy, S.R.; Ramani, S.; Natarajan, S.; Kim, Y.J.; Perumalsamy, H. Integrated transcriptome and in vitro analysis revealed anti-proliferative effect of citral in human stomach cancer through apoptosis. Sci. Rep. 2019, 9, 4883. [Google Scholar] [CrossRef]
- Izham, M.; Hussin, Y.; Aziz, M.; Yeap, S.; Rahman, H.S.; Masarudin, M.J.; Mohamad, N.E.; Abdullah, R.; Alitheen, N.B. Preparation and Characterization of Self Nano-Emulsifying Drug Delivery System Loaded with Citral and Its Antiproliferative Effect on Colorectal Cells In Vitro. Nanomaterials 2019, 9, 1028. [Google Scholar] [CrossRef] [Green Version]
- Sabo, A.A.; Dudau, M.; Constantin, G.L.; Cop, T.C.; Geifus, C.M.; Naccarati, A.; Dragomir, M.P. Two Worlds Colliding: The Interplay Between Natural Compounds and Non-Coding Transcripts in Cancer Therapy. Front. Pharmacol. 2021, 12, 652074. [Google Scholar] [CrossRef]
- Cherng, J.M.; Shieh, D.E.; Chiang, W.; Chang, M.Y.; Chiang, L.C. Chemopreventive effects of minor dietary constituents in common foods on human cancer cells. Biosci. Biotechnol. Biochem. 2007, 71, 1500–1504. [Google Scholar] [CrossRef]
- Lee, H.Y.; Son, S.W.; Moeng, S.; Choi, S.Y.; Park, J.K. The role of noncoding RNAs in the regulation of anoikis and anchorage-independent growth in cancer. Int. J. Mol. Sci. 2021, 22, 627. [Google Scholar] [CrossRef]
- Xu, Q.; Li, M.; Yang, M.; Yang, J.; Xie, J.; Lu, X.; Wang, F.; Chen, W. α-pinene regulates miR-221 and induces G2/M phase cell cycle arrest in human hepatocellular carcinoma cells. Biosci. Rep. 2018, 38, BSR20180980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poltronieri, P.; Xu, B.; Giovinazzo, G. Resveratrol and other Stilbenes: Effects on Dysregulated Gene Expression in Cancers and Novel Delivery Systems. Anticancer Agents Med. Chem. 2021, 21, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Fuentes-Mattei, E.; Fayyaz, S.; Raj, P.; Goblirsch, M.; Poltronieri, P.; Calin, G.A. Interplay between epigenetic abnormalities and deregulated expression of microRNAs in cancer. Semin. Cancer Biol. 2019, 58, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Chen, R.; Lu, G.; Li, C.; Lian, S.; Kang, T.W.; Do Jung, Y. Natural phytochemicals in bladder cancer prevention and therapy. Front. Oncol. 2021, 11, 652033. [Google Scholar] [CrossRef] [PubMed]
- Ruvinov, I.; Nguyen, C.; Scaria, B.; Vegh, C.; Zaitoon, O.; Baskaran, K.; Mehaidli, A.; Nunes, M.; Pandey, S. Lemongrass extract possesses potent anticancer activity against human colon cancers, inhibits tumorigenesis, enhances efficacy of FOLFOX, and reduces its adverse effects. Integr. Cancer. Ther. 2019, 18, 1534735419889150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, N.; Zainah, A.; Arapoc, D.J.; Mohamed, Z.A.M.A.; Shafii, K. Anticancer Effect and Apoptosis Induction of Cymbopogon citratus Plant on Head and Neck HTB43 Cancer Cell Lines. R&D Seminar 2016: Research and Development Seminar, Bangi, Malaysia, 8–10 November 2016. [Google Scholar]
- De Lima, D.A.N.; Pelegrini, B.B.; Uechi, F.A.A.; Varago, R.C.; Pimenta, B.B.; de Souza Kaneshima, A.M.; Kaneshima, E.N.; da Costa Souza, P.; Pedroso, R.B.; Silveira, T.G.V.; et al. Evaluation of Antineoplasic Activity of Zingiber Officinale Essential Oil in the Colorectal Region of Wistar Rats. Asian Pac. J. Cancer. Prevent. 2020, 21, 2141–2147. [Google Scholar] [CrossRef]
- Philion, C.; Ma, D.; Ruvinov, I.; Mansour, F.; Pignanelli, C.; Noel, M.; Saleem, A.; Arnason, J.; Rodrigues, M.; Singh, I.; et al. Cymbopogon citratus and Camellia sinensis extracts selectively induce apoptosis in cancer cells and reduce growth of lymphoma xenografts in vivo. Oncotarget 2017, 8, 110756–110773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.; Bhat, S.; Kamath, S.; Aggarwall, M.; Bhandarkar, M.; Mahima, B.; Sukreeth, S. A novel anti-oxidant lemongrass oil mouthwash-a clinical trial. J. Exp. Biol. Sci. 2011, 3, 14–18. [Google Scholar]
- Kim, I.; Oh, Y.A.; Lee, H.; Song, K.B.; Min, S.C. Grape berry coatings of lemongrass oil-incorporating nanoemulsion. LWT-Food Sci. Technol. 2014, 58, 1–10. [Google Scholar] [CrossRef]
- Tchoumbougnang, F.; Amvam Zollo, P.H.; Dagne, E.; Mekonnen, Y. In vivo antimalarial activity of essential oils from Cymbopogon citratus and Ocimum gratissimum on mice infected with Plasmodium berghei. Planta Med. 2005, 71, 20–23. [Google Scholar] [CrossRef]
- Ntonga, P.; Baldovini, N.; Mouray, E.; Mambu, L.; Belong, P.; Grellier, P. Activity of Ocimum basilicum, Ocimum canum, and Cymbopogon citratus essential oils against Plasmodium falciparum and mature-stage larvae of Anopheles funestus s.s. Parasite 2014, 21, 33. [Google Scholar] [CrossRef] [Green Version]
- Oladeji, O.S.; Adelowo, F.E.; Ayodele, D.T.; Odelade, K.A. Phytochemistry and pharmacological activities of Cymbopogon citratus: A review. Sci. African 2019, 6, e00137. [Google Scholar] [CrossRef]
- Aboelhadid, S.M.; Youssef, I.M.I. Control of red flour beetle (Tribolium castaneum) in feeds and commercial poultry diets via using a blend of clove and lemongrass extracts. Environ. Sci. Pollut. Res. 2021, 28, 30111–30120. [Google Scholar] [CrossRef] [PubMed]
- Solomon, B.; Gebre-Mariam, T.; Asres, K. Mosquito repellent actions of the essential oils of Cymbopogon citratus, Cymbopogon nardus and Eucalyptus citriodora: Evaluation and formulation studies. J. Essent. Oil-Bearing Plants 2012, 15, 766–773. [Google Scholar] [CrossRef]
- Eden, W.T.; Alighiri, D.; Supardi, K.I.; Cahyono, E. The Mosquito Repellent Activity of the Active Component of Air Freshener Gel from Java Citronella Oil (Cymbopogon winterianus). J. Parasitol. Res. 2020, 2020, 9053741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tak, J.; Isman, M. Metabolism of citral, the major constituent of lemongrass oil, in the cabbage looper, Trichoplusia ni, and effects of enzyme inhibitors on toxicity and metabolism. Pestic. Biochem. Physiol. 2016, 133, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, J.; Krnjajić, S.; Ćirković, J.; Radojkovic, A.; Popovic, T.; Brankovic, G.; Brankovic, Z. Effect of encapsulated lemongrass (Cymbopogon citratus L.) essential oil against potato tuber moth Phthorimaea operculella. Crop Prot. 2020, 132, 105109. [Google Scholar]
- Plata-Rueda, A.; Rolim, G.D.S.; Wilcken, C.F.; Zanuncio, J.C.; Serrão, J.E.; Martínez, L.C. Acute Toxicity and Sublethal Effects of Lemongrass Essential Oil and Their Components against the Granary Weevil, Sitophilus Granarius. Insects 2020, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Manh, H.D.; Hue, D.T.; Hieu, N.T.T.; Tuyen, D.T.T.; Tuyet, O.T. The mosquito larvicidal activity of essential oils from Cymbopogon and Eucalyptus species in vietnam. Insects 2020, 11, 128. [Google Scholar] [CrossRef] [Green Version]
- Feroz, A. Efficacy and cytotoxic potential of deltamethrin, essential oils of Cymbopogon citratus and Cinnamonum camphora and their synergistic combinations against stored product pest, Trogoderma granarium (Everts). J. Stored Prod. Res. 2020, 87, 101614. [Google Scholar] [CrossRef]
- El Mokhtari, K.; EL Kouali, M.; Talbi, M.; Hajji, L.; El Brouzi, A. Chemical composition and insecticidal activity of Anacyclus pyrethrum essential oil from the Bensliman area against Culex pipiens. Mediterr. J. Chem. 2020, 10, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Afify, A.; Betz, J.F.; Riabinina, O.; Lahondère, C.; Potter, C.J. Commonly Used Insect Repellents Hide Human Odors from Anopheles Mosquitoes. Curr. Biol. 2019, 29, 3669–3680. [Google Scholar] [CrossRef]
- Nordin, N.; Yeap, S.K.; Zamberi, N.R.; Abu, N.; Mohamad, N.E.; Rahman, H.S.; How, C.W.; Masarudin, M.J.; Abdullah, R.; Aitheen, N.B. Characterization and toxicity of citral incorporated with nanostructured lipid carrier. PeerJ. 2018, 6, 3916. [Google Scholar] [CrossRef] [Green Version]
- Madeira, P.L.; Carvalho, L.T.; Paschoal, M.A.; de Sousa, E.M.; Moffa, E.B.; da Silva, M.A.; Tavarez Rde, J.; Goncalves, L.M. In vitro Effects of Lemongrass Extract on Candida albicans Biofilms, Human Cells Viability, and Denture Surface. Front. Cell Infect. Microbiol. 2016, 6, 71. [Google Scholar] [CrossRef] [Green Version]
- Mat-Rani, S.; Chotprasert, N.; Srimaneekarn, N.; Choonharuangdej, S. Fungicidal Effect of Lemongrass Essential Oil on Candida albicans Biofilm Pre-established on Maxillofacial Silicone Specimens. J. Int. Soc. Prev. Community Dent. 2021, 11, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Bakrudeen, A.A.A. (Ed.) Microbial Biofilms. Properties and Applications in the Environment, Agriculture, and Medicine; CRC press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Martins, W.D.S.; de Arajo, J.S.F.; Feitosa, B.F.; Oliveira, J.R.; Kotzebue, L.R.V.; Agostini, D.L.D.S.; de Oliveira, D.L.V.; Mazzetto, S.E.; Cavalcanti, M.T.; da Silva, A.L. Lemongrass (Cymbopogon citratus DC. Stapf) essential oil microparticles: Development, characterization, and antioxidant potential. Food Chem. 2021, 355, 129644. [Google Scholar] [CrossRef] [PubMed]
- Choonharuangdej, S.; Srithavaj, T.; Thummawanit, S. Fungicidal and inhibitory efficacy of cinnamon and lemongrass essential oils on Candida albicans biofilm established on acrylic resin: An in vitro study. J. Prosthet. Dent. 2021, 125, 707.e1–707.e6. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.S.; Othman, H.H.; Hammadi, N.I.; Yeap, S.K.; Amin, K.M.; Abdul Samad, N.; Alitheen, N.B. Novel Drug Delivery Systems for Loading of Natural Plant Extracts and Their Biomedical Applications. Int. J. Nanomed. 2020, 15, 2439–2483. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Microbe | Family/ Kingdom | Cymbopogon citratus Essential Oil | Citral | ||
|---|---|---|---|---|---|---|
| IC50 [µL/L] | MIC [µL/L] | IC50 [µL/L] | MIC [µL/L] | |||
| Candida famata | fungi | Saccharomycetaceae | 177 ± 19 | 3684 ± 271 | 37 ± 7 | 142 ± 19 |
| Cryptococcus albidus | fungi | Tremellaceae | 199 ± 25 | 265 ± 31 | 2 ± 0 | 20 ± 6 |
| Candida albicans | fungi | Saccharomycetaceae | 571 ± 109 | 2734 ± 250 | 83 ± 8 | 110 ± 15 |
| Mycobacterium smegmatis | bacteria | Mycobateriaceae (Gram positive) | 860 ± 89 | 3409 ± 775 | 109 ± 12 | 137 ± 19 |
| Proteus vulgaris | bacteria | Enterobacteriaceae (Gram negative) | 992 ± 37 | 1453 ± 40 | 97 ± 12 | 163 ± 34 |
| Staphylococcus aureus | bacteria | Staphylococcaceaea (Gram positive) | 1841 ±199 | 5830 ± 198 | 77 ± 2 | 92 ± 2 |
| Pseudomonas aeruginosa | bacteria | Pseudomonadaceae (Gram negative) | 2385 ± 162 | 5308 ± 339 | 41 ± 2 | 93 ± 8 |
| Salmonella enterica | bacteria | Enterobacteriaceae (Gram negative) | 2626 ± 301 | 4693 ± 634 | 66 ± 8 | 97 ± 3 |
| Test Organisms | Family | Lemongrass Essential Oil | References | ||
|---|---|---|---|---|---|
| Species | MIC | MBC | |||
| Staphylococcus aureus | Staphylococcaceae | C. citratus | 0.06 * | 0.06 * | [34] |
| Bacillus cereus | Bacillaceae | C. citratus | 0.06 * | 0.06 * | |
| Bacillus subtilis | Bacillaceae | C. citratus | 0.06 * | 0.12 * | |
| Escherichia coli | Enterobacteriaceae | C. citratus | 0.12 * | 0.12 * | |
| Klebsiella pneumoniae | Enterobacteriaceae | C. citratus | 0.50* | 0.50 * | |
| Staphylococcus aureus | Staphylococcaceae | C. flexuosus | 0.0781 * | - | [24] |
| Enterococcus faecalis | Enterococcaceae | C. giganteous and C. citratus | 1 ** | - | [66] |
| Staphylococcus aureus | Staphylococcaceae | C. giganteous and C. citratus | 2.5 ** | - | |
| Listeria monocytogenes | Listeriaceae | C. giganteous and C. citratus | 8.3 ** | - | |
| Enterobacter aerogenes | Enterobacteriaceae | C. giganteous and C. citratus | 13.3 ** | - | |
| Escherichia coli | Enterobacteriaceae | C. giganteous and C. citratus | 10 ** | - | |
| Pseudomonas aeruginosa | Pseudomonadaceae | C. giganteous | >80 ** | - | |
| Salmonella enterica | Enterobacteriaceae | C. giganteous and C. citratus | 2.1 ** | - | |
| Salmonella typhimurium | Enterobacteriaceae | C. giganteous and C. citratus | 2.5 ** | - | |
| Shigella dysenteriae | Enterobacteriaceae | C. giganteous and C. citratus | 8.3 ** | - | |
| Escherichia coli | Enterobacteriaceae | C. khasianus | 20 # | 20 | [43] |
| Staphylococcus aureus | Staphylococcaceae | C. khasianus | 30 # | – | |
| Pseudomonas aeruginosa | Pseudomonadaceae | C. khasianus | 20 # | 30 | |
| S. enterica typhimurium | Staphylococcaceae | C. khasianus | 30 # | – | |
| Bacillus subtilis | Bacillaceae | C. khasianus | 25 # | – | |
| Klebsiellapneumoniae | Enterobacteriaceae | C. khasianus | 20 # | – | |
| Fungal Species | Family/Kingdom | Lemongrass Essential Oil | References | |
|---|---|---|---|---|
| Species | MIC | |||
| Candida albicans | Saccharomycetaceae | C. flexuosus | 0.0781 * | [26,77] |
| Candida tropicalis | Saccharomycetaceae | C. flexuosus | 0.039 * | |
| Candida albicans | Saccharomycetaceae | C. khasianus | 100 | [43] |
| Aspergillus flavus | Trichocomaceae | C. citratus | 50 ## | [45] |
| Aspergillus niger | Trichocomaceae | C. citratus | 30 ## | |
| Alternaria alternata | Pleosporaceae | C. citratus | 30 ## | |
| Aspergillus fumigatus | Trichocomaceae | C. citratus | 30 ## | |
| Fusarium moniliforme | Nectriaceae | C. citratus | 30 ## | |
| Cochliobolus lunatus | Pleosporaceae | C. citratus | 30 ## | |
| Drosera indica | Droseraceae | C. citratus | 20 ## | |
| Tissue | Cell Lines | Lemongrass Essential Oil | References | |
|---|---|---|---|---|
| Species | IC50 (μg/mL) | |||
| colon | HT-29 | C. flexuosus | 42.4 | [107] |
| HCT-152 | C. flexuosus | 60.2 | ||
| SW-620 | C. flexuosus | 28.1 | ||
| 502713 | C. flexuosus | 4.2 | ||
| lung | H-226 | C. flexuosus | 61.4 | |
| A-549 | C. flexuosus | 49.7 | ||
| Hop-62 | C. flexuosus | 79 | ||
| liver | Hep-2 | C. flexuosus | 4.8 | |
| cervix | SiHa | C. flexuosus | 6.5 | |
| prostate | DU-145 | C. flexuosus | 41.5 | |
| oral | KB | C. flexuosus | 50.8 | |
| neuroblastoma | IMR-32 | C. flexuosus | 4.7 | |
| lung | A549 | C. citratus | 1.73 | [107] |
| H1975 | C. citratus | 4.01 | ||
| Components | Experimental Model | Mechanism of Action | References |
|---|---|---|---|
| citral | A549 (human lung carcinoma) | Growth arrest of cell cycle at sub G1 phase | [115] |
| NCI-H1975 (human lung adenocarcinoma) | Up-regulation of procaspase-3 | ||
| NCI-H1650 (human lung adenocarcinoma) | Decrease of Bcl-2 and increase of expression of Bax | ||
| NCI-H1299 (human lung large cell carcinoma) | |||
| citral | Prostate cancer cells PC3 and PC3M (metastatic) | Inhibition of colony formation, suppression of expression of AMPK pathway genes SREBP1, ACC, HMGR | [119,150] |
| Colony forming assay 10, 15, 25, 50, 100 μg/Ml | 216 upregulated genes 396 downregulated genes | ||
| ATCC-CRL-1739/ | Apoptosis, block of colony formation and migration | ||
| AGS stomach cancer cells | |||
| 5, 10, 20 μg/mL | |||
| citral | Human colorectal cancer HCT116 and HT29 cells | Induction of phosphorylation of p53, triggering ROS mediated mitochondrial intrinsic apoptosis | [124,151] |
| HT29, SW620 lines | Cytotoxicity | ||
| geraniol | A549 human lung adenocarcinoma cells in culture and in vivo in nude mice | Decreased the level of membrane-bound Ras protein, decreased the level of cholesterol and HMGCR protein | [127] |
| geraniol | In vitro murine endothelial-like eEND2 cells and HDMEC (dermal microvascular endothelial cells), | Blocked VEGF/ VEGFR signal transduction and suppression of cAKT and ERK signalling pathways | [128] |
| In vivo, CT26 cell lines from undifferentiated colon carcinoma of the BALB/ c mouse | |||
| geraniol | Human hepatoma (HepG2) and human lung adenocarcinoma (A549) cell lines | Growth arrest in G0/G1 interphase of the cell cycle, increased the production of ROS | [129] |
| citronellol | Invitro, non-small lung cancer cell (NCI-H1299); | Arrest of cell cycle at G1 phase, down-regulation of expression of cyclin E, and cyclin D, increase in expression of TNF-α, and activation of RIP1/RIP | [129] |
| In vivo, injected NCI-H1299 into BALB/c nude mice | |||
| citronellol | Triple-negative breast cancer MDA-MB-231 cell line | Decreased the expression of Bcl-2 gene and protein and increased Bax expression. | [141] |
| citronellol | DMBA(7,12-dimethylbenz(a) anthracene) induced mammary cancer in rats | Down-regulation of expression of NF-kB, IL-6, and TNF-α. Suppression of activity of COX-2. | [142] |
| α-bisabolol | CML-T1, Jurkat, HeLa cell lines | Cytotoxicity via mitochondria and lysosome-initiated caspase cascade and induction of autophagy and apoptosis | [143] |
| α-bisabolol | KLM1, KP4, and Panc 1 human pancreatic cancer cell lines | Up-regulation of KISS1R | [146] |
| α-bisabolol | Endometrial cancer cell lines RL95–2, ECC001, ECC003 | Decreased activity of COX-2, induction in PARP cleavage, increased apoptosis via XIAP/ caspase 3 pathway | [148,149] |
| Ishikawa cell line | |||
| ECC E6/E7 cell line | |||
| limonene | Bladder cancer line T24; | Arrest of cell cycle in the G2/M phase; block of cancer cell migration; apoptosis; inhibition of PI3K/AKT pathway induces cell cycle G2/M, suppressing migration. Induces chromatin concentration, nuclear fragmentation, increases Bax, caspase 3, decreases Bcl-2 | [135,149] |
| colon cancer LS174T line; | |||
| Bladder cancer cells | |||
| pinene | HepG2, HCC cells | CDKN1C/p57 and p27/CDKN1B upregulation, miR-221 downregulation | [152,153,154] |
| linalool | HeLa, | antiproliferative | [150] |
| H520 lung cancer line, | |||
| BCC-1/KMC skin cancer |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukarram, M.; Choudhary, S.; Khan, M.A.; Poltronieri, P.; Khan, M.M.A.; Ali, J.; Kurjak, D.; Shahid, M. Lemongrass Essential Oil Components with Antimicrobial and Anticancer Activities. Antioxidants 2022, 11, 20. https://doi.org/10.3390/antiox11010020
Mukarram M, Choudhary S, Khan MA, Poltronieri P, Khan MMA, Ali J, Kurjak D, Shahid M. Lemongrass Essential Oil Components with Antimicrobial and Anticancer Activities. Antioxidants. 2022; 11(1):20. https://doi.org/10.3390/antiox11010020
Chicago/Turabian StyleMukarram, Mohammad, Sadaf Choudhary, Mo Ahamad Khan, Palmiro Poltronieri, M. Masroor A. Khan, Jamin Ali, Daniel Kurjak, and Mohd Shahid. 2022. "Lemongrass Essential Oil Components with Antimicrobial and Anticancer Activities" Antioxidants 11, no. 1: 20. https://doi.org/10.3390/antiox11010020