
Biosynthesis, Quantification and Genetic Diseases of the Smallest Signaling Thiol Metabolite: Hydrogen Sulfide

 , and
, and
Abstract
:
1. Introduction
1.1. Biological Chemistry of H2S
1.2. Concentration, Signaling and Cellular Response to H2S
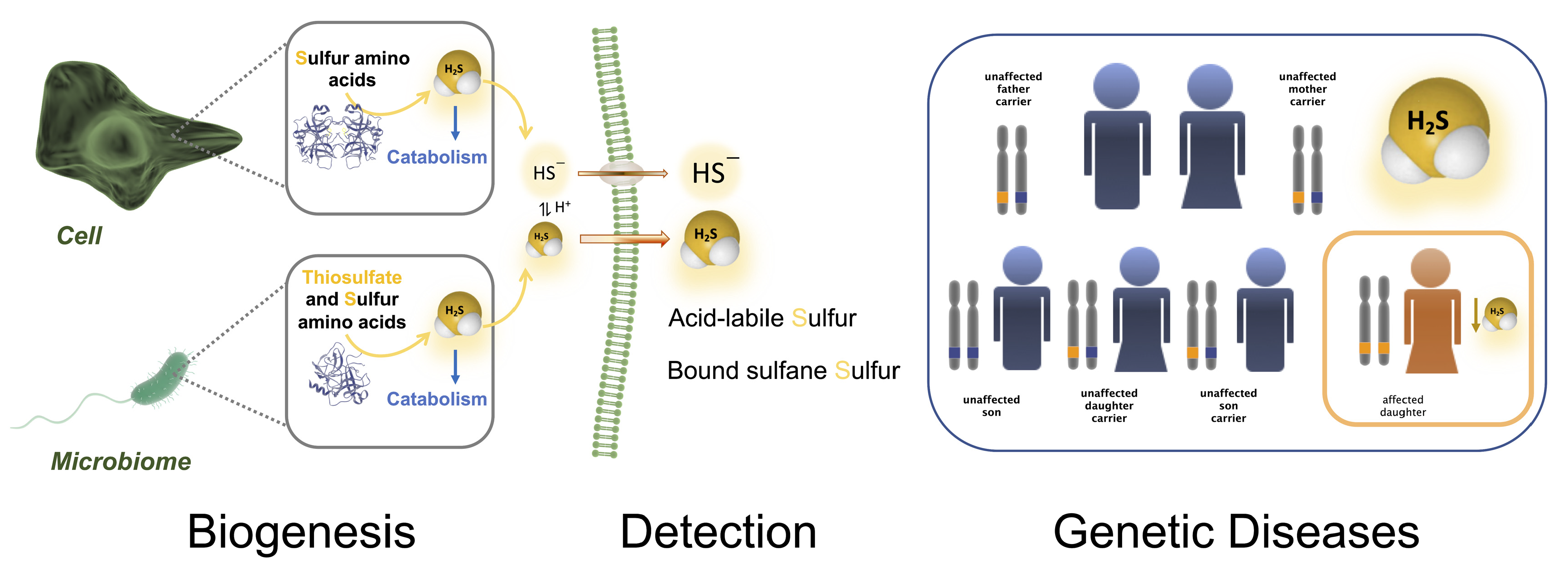
2. Biogenesis, Catabolism and Reservoir of Hydrogen Sulfide in Humans
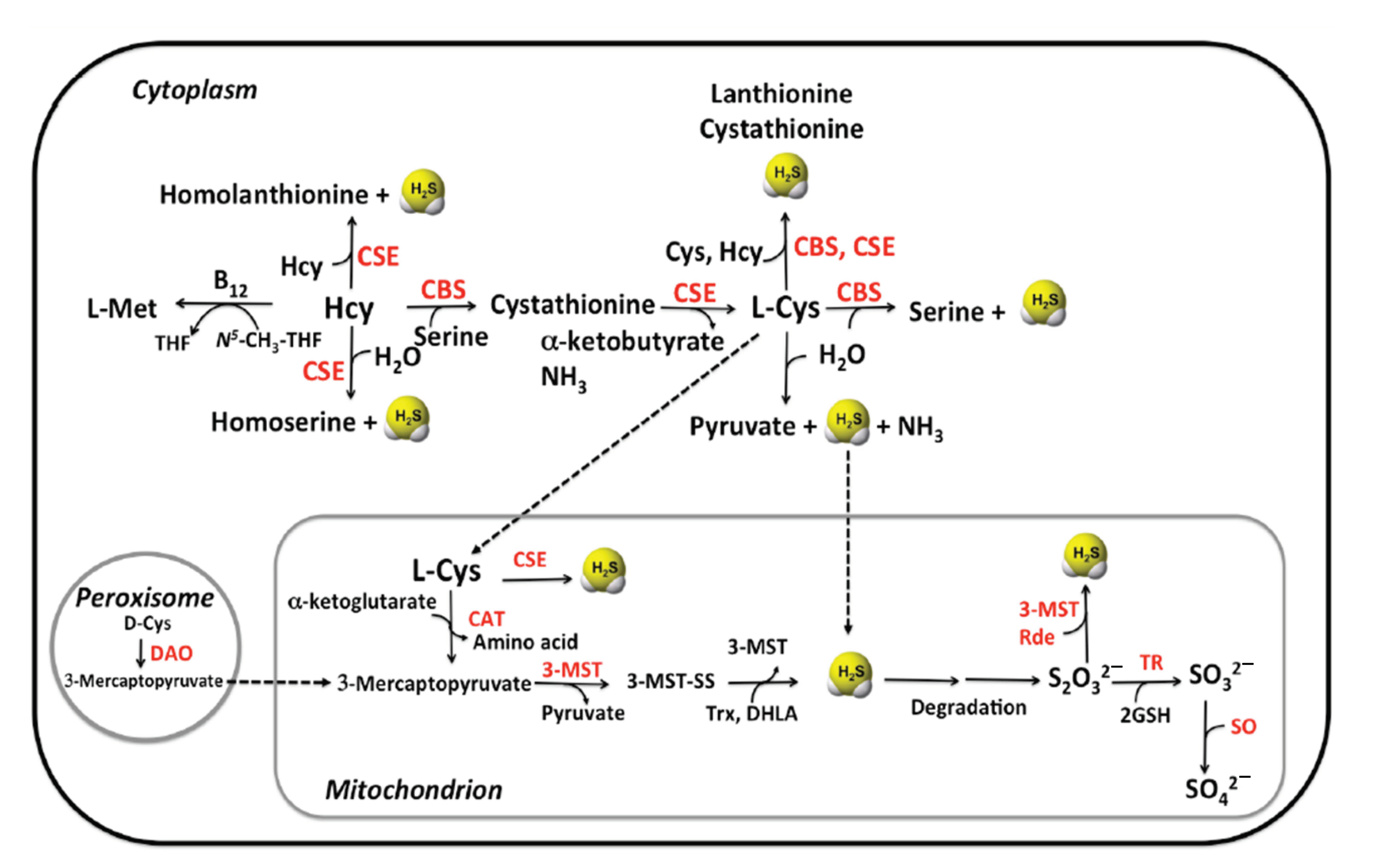
2.1. Enzymatic Production of Hydrogen Sulfide
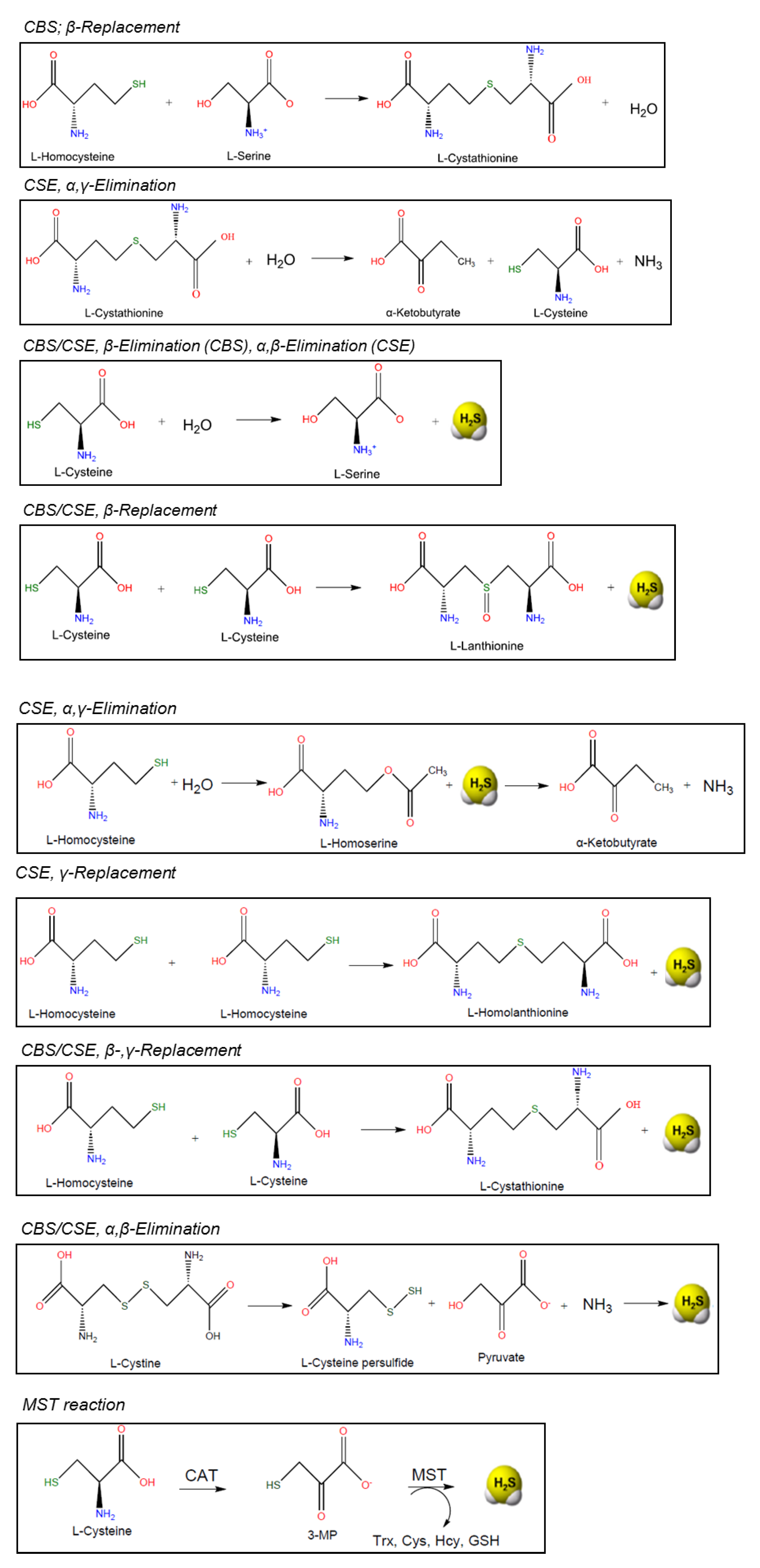
2.1.1. CBS
2.1.2. CSE
2.1.3. MST
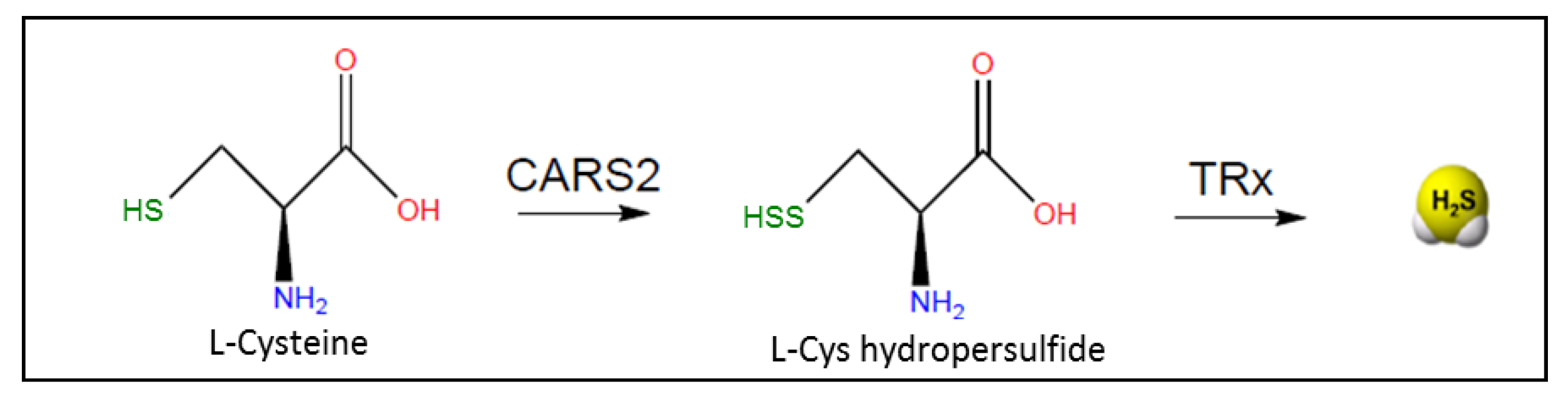
2.2. Non-Canonical Biogenesis of H2S
2.3. Hydrogen Sulfide Derived from the Microbiome
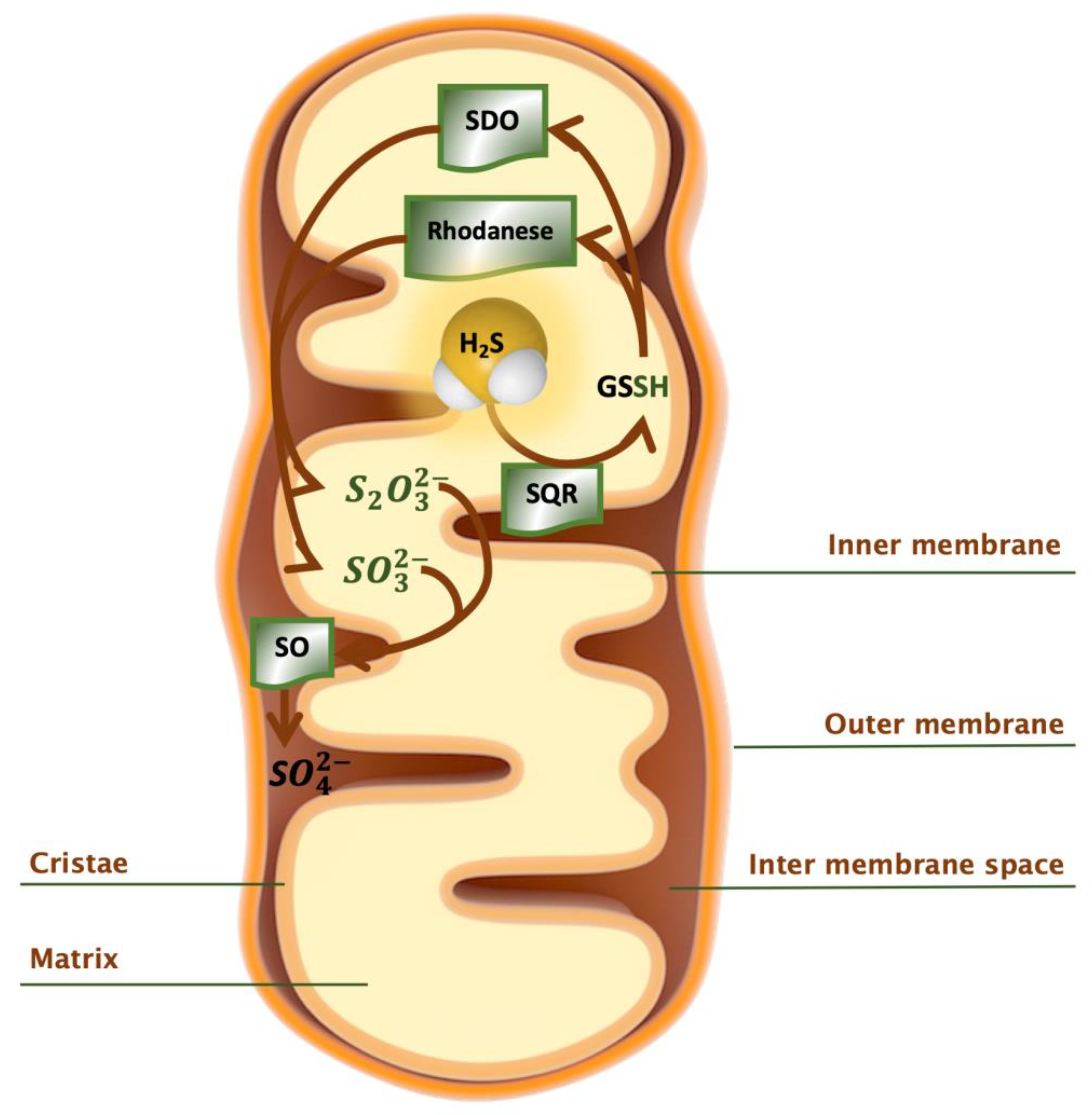
2.4. Catabolism of H2S
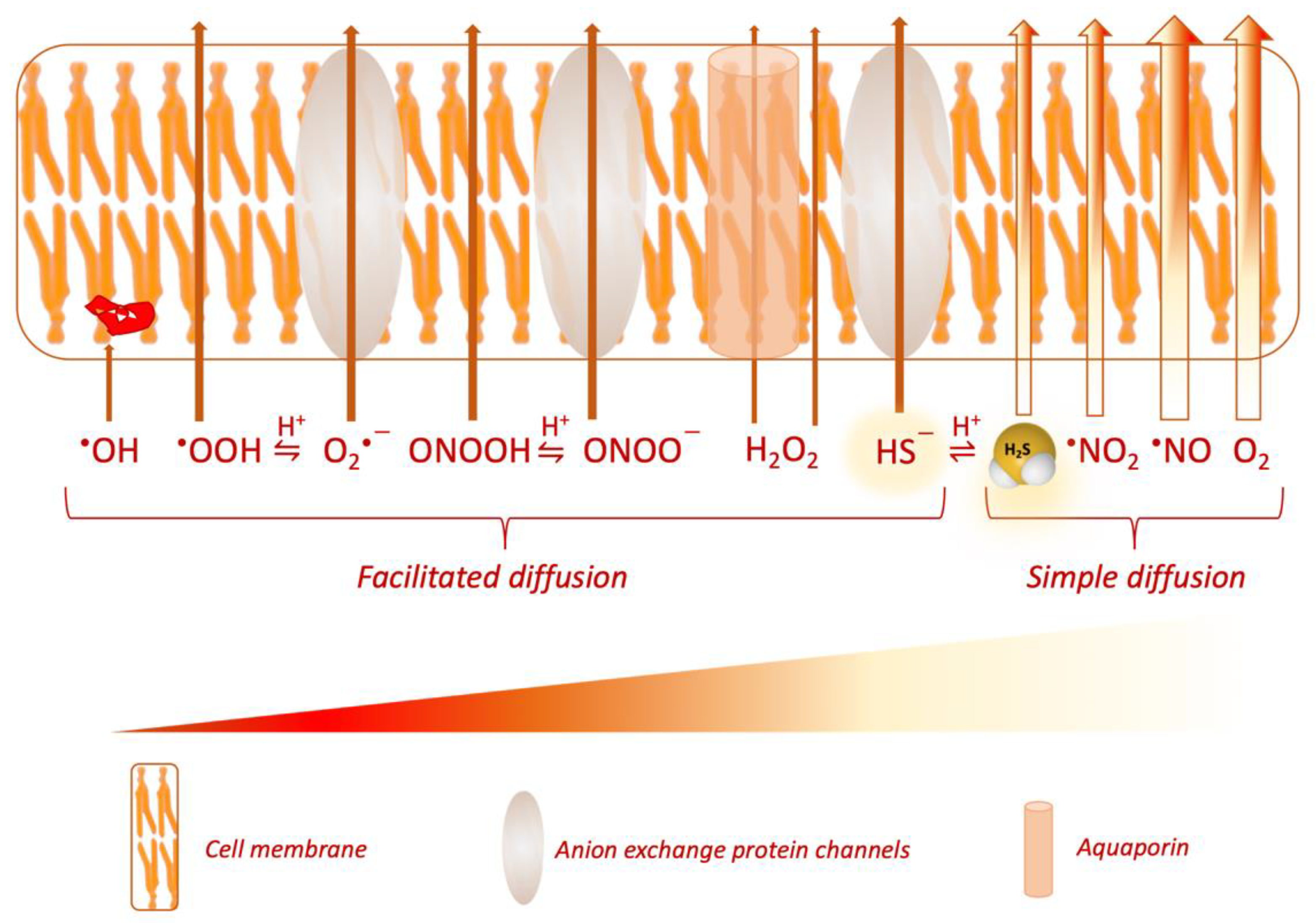
2.5. Reservoir of H2S
2.6. Antioxidant Functions of H2S
3. Methods of Detection of Hydrogen Sulfide
3.1. Direct UV-Visible Spectrophotometric Measurement
3.2. Chromatography
3.2.1. Gas Chromatography/Ion Chromatography
3.2.2. Flow Gas Dialysis/Ion Chromatography
3.2.3. Gas Chromatography/Chemiluminescence Sulfur Detector
3.2.4. Gas Chromatography with Silver Particles
3.2.5. High Pressure Liquid Chromatography (HPLC)
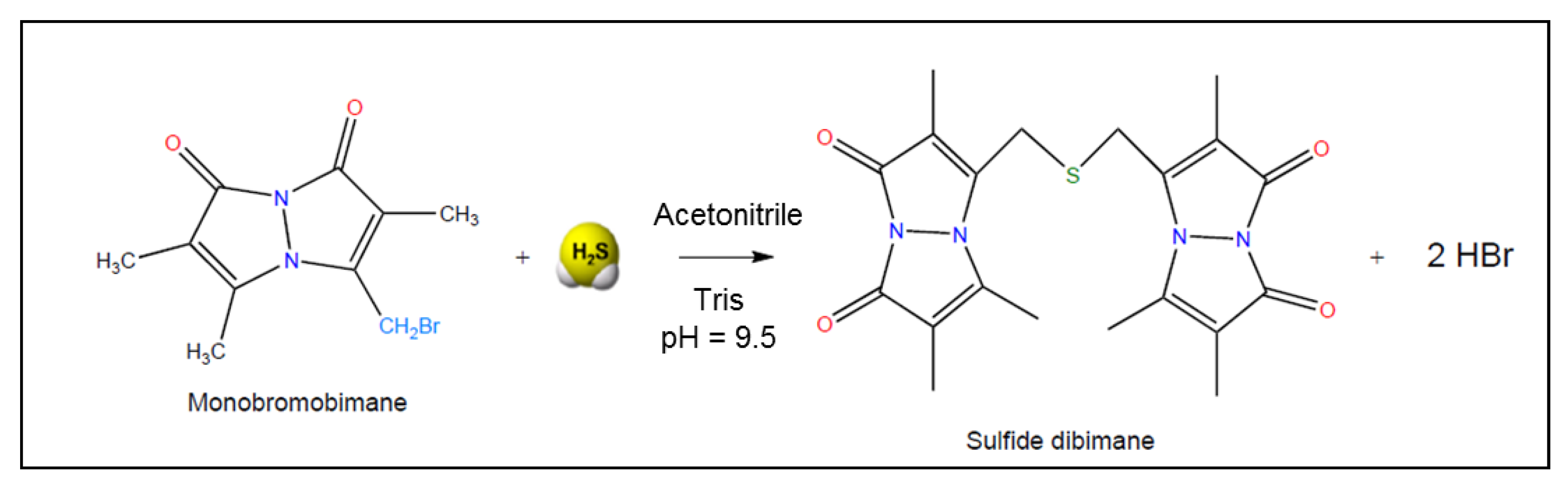
3.3. Monobromobimane Assay
3.4. Other Fluorescent Probes for H2S Detection
3.5. Sulfide Specific Ion-Selective Electrodes (ISE)
3.6. Polarographic Electrodes

{kind=link}
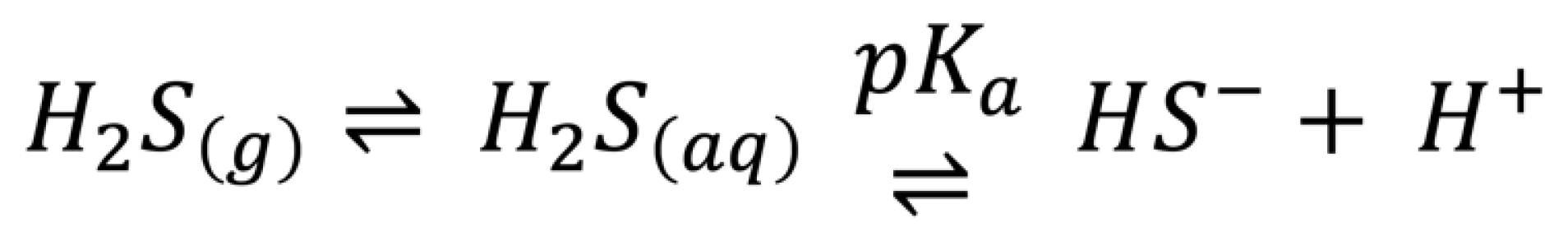{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Sample Pre-Analytics | Limit of Detection | Type of Sample | Literature |
|---|---|---|---|---|
| 1. Direct spectrophotometric measurement | ||||
| 1.1. Direct ultraviolet detection of the HS− ion, absorption measurement from 214 to 300 nm (peak around 230 nm) | Sulfide dissolved in natural water, pH ~8.0 | <1 µM | Dissolved sulfide in natural waters | [157] |
| 1.2. Zinc trap/methylene blue: Trapping sulfide with a metal (e.g., zinc acetate) with subsequent acidification and reaction with DMPD, formation of methylene blue by a ferric chloride catalyzed reaction, spectrophotometric measurement of methylene blue, improvement by replacing the spectrophotometer detector with a mass spectrometer | No pre-treatment of sulfide is necessary | 1 μM 50 ng L− | Aquatic sample PSI particles | [163,164,165,166,167] |
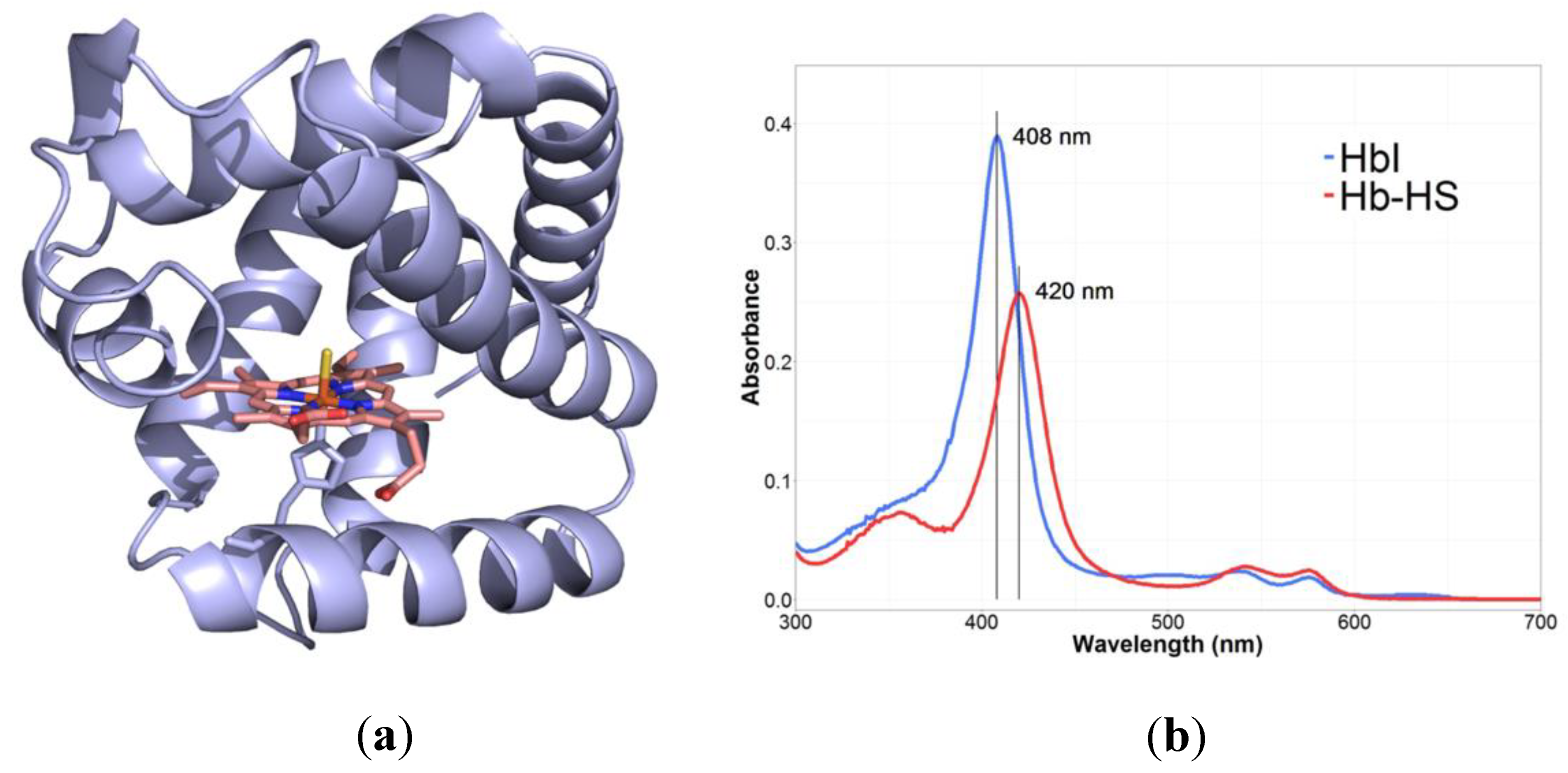
| 1.3. Hemoglobin I. Based on the 1:1 reaction between H2S and the ferric species of hemoglobin I (HbI),simple and fast, measuring of absorbance at 408 nm and 429 nm, determination of the fractional saturation of HbI-H2S complex | Purification of ferric L. pectinata HbI from the clam, determination of HbI concentration, H2S of analytical grade was used without further purification, calculation of solubility of H2S | Up to nM | Ferric L. pectinata HbI, dissolved H2S of analytical grade | [158,169] |
| 2. Chromatography | ||||
| 2.1. Gas chromatography/ion chromatography: Determination of H2S using an A7 column equipped with a flame photometric detector (GC-FPD)/quantitation with electrochemical detection | Tissues are treated with acid to liberate H2S from ALS, sulfane sulfur is liberated as thiocyanate by treating tissues with cyanide, homogenization with sodium chloride/oxidation of hydrogen sulfide by hydrogen peroxide | >100 nm/g | Rat liver and heart tissues | [37] |
| 2.2. Flow gas dialysis/ion chromatography: Separation of sulfide from the homogenate matrix enables a pre-concentration/electrochemical detection and quantitation of H2S through a silver working electrode, a silver/silver chloride reference electrode and a stainless-steel counter electrode | Fatal intoxication of the brain by intraperitoneal injection of NaHS, homogenization with NaOH, reaction of the homogenate with HCl to release H2S | 100 μM | Human and rat brain tissue | [41] |
| 2.3. Gas chromatography/chemiluminescence: Injection of a gas sample onto a Teflon column, determination of H2S concentration via comparison of peak area of the unknown to that of known standards | Incubation of homogenized tissue in media, homogenization in ice-cold potassium phosphate buffer, addition of aliquots of the tissue homogenate to pre-warmed polypropylene syringes | 17 nM 14 nM 100 pM | Liver tissue Brain tissue of mice Human blood | [32] |
| 2.4. Gas chromatography/silver particles: Adsorption of H2S as Ag2S through silver particles from the sample, application of thiourea and H2SO4 to protein-free silver particles to release H2S from silver sulfide produced on the surface of the particles, measurement of H2S concentration at various pH values with GC | Alkalinization of the cytoplasm for H2S release from bound sulfur, acid-labile sulfur: homogenization of rat tissues with NaOH, neutralization of cleared supernatant with phosphoric acid, bound sulfur: homogenization of rat or mouse tissue with a polytron homogenizer in ice-cold buffer, addition of DTT to supernatant | 15 nM 9 μM 6 pM | Mouse blood Rat brain Rat homogenate tissue | [34,38] |
| 2.5. Liquid chromatography–mass spectrometry: Measurement of CBS activity, using the stable isotope substrate 2,3,3-2H serine, monitoring of 3,3-2H-cystathionine, cystathionine and 3,3,4,4-2H-labeled cystathionine formation | Addition of pyridoxine to all plasma samples at the time of sampling, addition of plasma or serum to a solution containing Tris- pyridoxal 5′-phosphate and 2,3,3-2H-labeled serine, activation of CBS by SAM | Up to nM | Human plasma or serum | [186] |
| 2.6. Modification of liquid chromatography–mass spectrometry: Optimization of liquid chromatography with a commercially available Symmetry C18 column and MS/MS detection | No difference | 0.1 μM | Human plasma or serum | [187] |
| 2.7. HPLC:(2.7.1) Derivatization of sulfide via reversed-phase HPLC (RP-HPLC) separation, determination of sulfide via spectrophotometric detection of methylene blue, trapping of H2S with zinc acetate, measuring of H2S concentration under acidic conditions | Addition of zinc acetate to each rumen fluid sample, homogenization of cerebrocortical gray matter in aqueous zinc acetate, treatment of human whole blood with heparin, preparation of spiked samples by adding sulfide standard solutions, conversion into the fluorescent derivative thionine, under acidic conditions | 0.123 to 0.189 pmol dm−3 | Bovine brain tissue | [174] |
| (2.7.2) Separation of sulfide on a reversed-phase column and fluorimetrical detection, after pre-column fluorescence derivatization, conversion of low sulfide concentration into thionine, formation of methylene blue | Human red blood cells | [175] | ||
| 3. Monobromobimane assay (gold standard) | ||||
| 3.1. Reaction with the thiol-specific derivatization agent monobromobimane to sulfide-dibimane, detection of sulfide-dibimane concentration by reversed-phase fluorescence HPLC and/or mass spectrometry | Reaction of sulfide with 2.0 equivalents of monobromobimane, formation of sulfide-dibimane | 0.7 μM | Rat bloodHuman plasma | [35] |
| 3.2. Monobromobimane method coupled with RP-HPLC and fluorescence detection, measurement of sulfide by methylene blue | Liberation of acid-labile H2S by incubation of the sample in an acidic solution (pH 2.6) with diethylenetriaminepentaacetic acid (DTPA), incubation of the sample with TCEP | 0.7 μM | Mouse plasma | [33] |
| 4. Fluorescent probes | ||||
| 4.1. Conductometric gas nanosensors for H2S, which are synthesized by electrodepositing gold nanoparticles on single-walled carbon nanotube (SWNT) networks, detection of H2S is based on conductivity changes of thin films upon exposure to H2S gas | Hydrogen sulfide (purity: 99.998%) diluted in dry air at a gas flow of 200 std. cm~3 min−1 | 3 ppb | H2S gas | [188] |
| 4.2. Measurement of H2S plasma concentration by assessing the fluorescence signal from benzodithiolone product formation. Benzodithiolone has a bis-electrophilic center with specificity for H2S detection | Preparation of the probe with thiosalicylic acid, preparation of bovineplasma containing NaHS at different concentrations (0, 50, 100 and 500 mm) | Reactive disulfide-containing probeBovineplasma | [177] | |
| 4.3. Dansyl azide (DNS-Az) is a reduction-sensitive, stable, non-fluorescent chemoprobe, it becomes fluorescent upon reacting with sulfide | No sample pre-treatment | 1 μM 5 μM | Aqueous solution Bovine serumMouse blood | [178] |
| 4.4. 7-azido-4-methylcoumarin (AzMC) is a fluorogenic probe selective for H2S; it provides a facile, direct, continuous and sensitive assay for activity monitoring of PLP-dependent enzymes | Synthesis and purification of hCBS | 0.1 µM | Truncated version of human CBS (without the regulatory domain) | [179] |
| 4.5. 4-methyl-2-oxo-2H-chromen-7-yl 5-azidopentanoate, a fluorescent probe for H2S detection, two characteristic reactions of H2S: reduction of an azido group to an amine, intramolecular lactamization with simultaneous release of highly fluorescent 7-hydroxy-4-methylcoumarin | Saliva samples were tested instantly, measurements do not require any chemical pre-treatment, aqueous medium at pH = 7.4 | 1.641–7.124 µM | Sodium phosphate buffer at physiological pHSaliva | [40] |
| 4.6. N-[2(2-hydroxyethoxy)ethyl]-6,8-difluoro- 7-hydroxycoumarin-3-carboxamide is a turn-on fluorescent sensor to detect H2S, the sensor rHbI functions as a filter at the excitation wavelength of fluorophore in absence of H2S, but not in its presence, permits end-point measurement of H2S concentration, reversible and selective | Expression of recombinant rHbI in E. coli BLi5 strain, purification of recombinant rHbI, UV−vis spectrum to confirm the formation of rHbI-H2S, adding of potassium cyanide to the reaction mixture, monitoring of displacement reaction by UV−vis spectroscopy | ∼200 nM at pH 7.4 | Recombinant hemoglobin I from Lucina pectinata Human plasma | [156,180] |
| 5. Sulfide specific ion-selective electrodes | ||||
| Measurement against a glass pH electrode, ISEs measure the HS− form of sulfide in an alkaline environment to favor HS− formation | Samples placed in alkaline buffer, saturated solutions of sparingly soluble metal salts | 1 fM–1 μM | Bovine serum albumin (BSA) | [25,26,183,184] |
| 6. Polarographic electrodes | ||||
| 6.1. PHSS records the concentration in a respirometer chamber/vessel, selective for H2S, responds rapidly to varying H2S concentrations, contains a H2S-permeable polymer membrane | Injection of supernatant into the respirometer chamber containing PBS, addition of the substrate L-cysteine and the cofactor PLP to support the activity of enzymes CBS and CSE | 10 nM | Rat aorta Smooth muscle cellsIntact rat thoracicaorta | [36] |
| 6.2. Polydimethylsiloxane (PDMS) membranes are permeable to H2S, a continuous measurement of H2S is possible, not permeable for other thiols | Purification of CSE | ∼0.51 μM | Free H2S | [185] |
4. Determination of Hydrogen Sulfide Production in Genetic Diseases of Sulfur Metabolism
4.1. H2S Metabolism in Human Studies
4.2. H2S Metabolism in Animal Models of Human Disease
4.2.1. Cystathione β-Synthase (CBS)
4.2.2. Cystathione γ-Lyase (CSE)
4.2.3. Cysteine Dioxygenase
4.2.4. Rhodanese
| Mutation | H2S Formation | Enzyme Activity | Biological Sample | Method | Literature |
|---|---|---|---|---|---|
| Human Samples | |||||
| Pyridoxine non-responsive CBSD | Plasma sulfide concentration was similar to the controls | Absent activity of CBS→ Hcy elevation in plasma and urine, grossly decreased Cystat in plasma and urine, synthesis of H2S from grossly elevated Hcy by CSE | Plasma samples | Monobromobimane (MBB) derivatization followed by HPLC separation and fluorescent quantitation | [42] |
| Thiosulfate was increased three times | Urine | ||||
| RMD due to cblG, cblE, cblJ | Thiosulfate was increased 1.7 and 2.4 times | Decreased enzyme activity to 53% and 64% of the median in healthy controls | Plasma sample, urine | Monobromobimane (MBB) derivatization followed by HPLC separation and fluorescent quantitation | [42] |
| ETHE1 | Plasma sulfide increased 5-fold with respect to reference range | Absent enzyme activity | Human plasma | Monobromobimane (MBB) derivatization followed by HPLC separation and HPLC-MS/MS measurements | [27] |
| T152I ETHE1 mutation | Marked increase in H2S and thiosulfate concentration in both mutations | 3-fold lower activity of the enzyme, 4-fold decrease in Vmax, Km for GSSHunaffected | Recombinant human ETHE1 missing the N-terminal mitochondrial leader peptide | Monobromobimane, reverse phase HPLC with a multi-signalfluorescence detector, mass spectroscopy | [127] |
| D196N ETHE1 mutation | 2-fold higher Km for the substrate glutathione persulfide, around 15% decrease in Vmax, whereas 2-fold higher Km for GSSH | ||||
| ETHE1−/− | H2S concentration in all tissues much higher compared to WT: brain: 5-fold higher, muscle: 10-fold higher, liver: 10-fold higherthiosulfate in urine several-fold higher, sulfate lower, thiosulfate markedly increased in kidney, liver, muscle and brain | Total lack of ETHE1 activity | Ethe−/− mice: liver, brain, kidney, external muscle, layers of colon, urine | Zinc acetate trap, gas chromatography/sulfur chemiluminescence detector | [197,205] |
| human (with ethylmalonic encephalopathy) blood and urine | |||||
| CBS p.P49L | 3-fold lower H2S formation in PLP-untreated CBS, similar formation by SAM stimulation | Structural changes affect heme reactivity, enhanced affinity for CO, poor responsiveness to SAM | Human cultured fibroblasts | Amperometry with H2S -selective electrode/coulometry (acetate method), UV-spectrophotometry | [193,206] |
| H2S formation similar to WT in PLP-treated CBS, impaired formation by SAM stimulation (1.1-fold vs. 1.9-fold in WT)CO inhibits the H2S producing activity | |||||
| Vitamin B6 insufficiency | Impaired H2S production capacity because of the sensitivity of CSE to inactivation by loss of the PLP coenzyme | Little or no effect on CBS activity in the presence or absence of stimulation by SAM, CSE exhibited 70% lower activity, lower production of H2S-specific biomarkers | Human lysates of cultured hepatoma cells | Gas chromatography with sulfur chemiluminescence detector | [194] |
| c.637G > A, a Glu213Lys variant of SQORc.446delT | Episodic accumulation of H2S, at high concentrationsH2S can coordinate to the iron in the heme a group of complex IV resulting in strong inhibition | Compared to WT, mutants exhibited: reduced SQOR protein and enzyme activity, unchanged sulfide generating enzyme concentration, reduced fibroblast SQOR enzyme activity and protein concentration, reduced SQOR protein and enzyme activity, unchanged sulfide generating enzyme concentration, decreased complex IV activity, but normal complex IV protein concentration and complex formation | Isolated mitochondrial membrane fractions from human liver, lysates derived from peripheral blood mononuclear cells and from fibroblasts | UV-spectrophotometry, monitoring of the decrease in absorbance at 278 nm due to reduction of CoQ1 | [131,195] |
| Animal models | |||||
| Cysteine dioxygenase knockout CDO−/− | 4.5-fold elevated concentration of urinary thiosulfate, lower abundances of COX4 and COX5b in liver, pancreas and kidney, slight elevation in plasma sulfate concentration, excess H2S/HS− production | Complete lack of CDO activity | Mouse liver, kidney, brain, pancreas | Indirect measurement of H2S (i.e., thiosulfate excretion, cytochrome c oxidase (COX) inhibition): taurine and hypotaurine were measured by HPLC, COX4 and COX5 by Western blotting | [204] |
| Homozygous (CSE−/−) and heterozygous (CSE−/+) mutant mice | Homozygous mice compared to WT exhibited: endogenous H2S concentration in aorta and heart decreased by about 80%, serum H2S concentration by about 50% Heterozygous mice compared to WT exhibited: H2S concentration in aorta and heart decreased by about 50%, serum H2S concentration decreased by about 20% | CBS−/−: Complete lack of enzyme activityCSE−/+: around 50% lower CBS activity versus wild type | Mouse heart, aorta, vascular system, | Ion-selective electrode on a Fisher Accumet Model 10 pH meter | [96] |
| CBS heterozygous knockout (CBS−/+) mice | Significantly reduced H2S concentration by 30% and 46% compared to wild type in male and female CBS−/+ | Around 50% CBS activity compared to WT | Mouse liver | Methylene blue method, HPLC and quantification by fluorescence detector (measurement of thiol metabolites) | [200] |
| CBS treated with the Ad-lacZ virus (Ad-hCBS) | Increased H2S concentration in plasma by 1.8-fold | Increased CBS activity by 1.4-fold, reduced homocysteine concentration significantly by 5.3-fold | |||
| E102D variant of rhodanese P285 A variant of rhodanese | H2S production in murine liver lysate is low | More stable than wild type, exhibition of 4- and 2.7-fold higher specific activity, lower Kmfor cyanide for the E102D, but higher for the P285A, higher Km for thiosulfate for E102D, but lower for P285A, 17- and 1.6-fold higher catalytic efficiency (kcat/Km(CN)) in the cyanide detoxification assay, 1.6 and 4-fold lower sulfur transfer reaction from GSSH to sulfite, similar kcat/Km GSSH values | Murine liver, human blood (wild type human rhodanese and recombinant variants) | Turbidimetric lead acetate assay, UV-spectrophotometry, monobromobimane, reverse phase HPLC column, MS (detection of thiosulfate) thiosulfate-dependent H2S generation: gas chromatography | [134] |
| CSE−/− mice on a C57BL/6 background (in hypoxia for a 72 h period)injection of either saline | Reduced H2S concentration in mutant compared to WT | Lower hemoglobin concentration, higher EPO concentration than wild type mice during normoxia, but lower during hypoxia, hypoxia downregulates CBS expression | Mouse blood, kidney, human urine | sulfide/H2S -sensitive microelectrode system (ion electrode)HPLC (thiosulfate concentration) | [203] |
| or Na2S | Comparable blood H2S concentration in mutants and wild type | Rescue of hemoglobin concentration, rescued EPO concentration during hypoxia, increased CBS protein concentration in hypoxic CSE−/− | |||
| CBS+/− (B6.129P2-CBStm1Unc/J 002853) | H2S added exogenously via GYY4137, a slow H2S donor | Decreased CBS, CSE and MTHFR expressions, GYY4137 treatment rescues their concentration, GYY4137 reduces oxidative stress marker concentration and total GSH concentration, it normalizes overall glutamate concentration, it reduces ocular pressure back to the baseline levels and enables less vessel density and permeability in the eye | Mouse blood, retinal tissue | GYY4137 (crystalline solid in nature) dissolved in phosphate buffered saline (PBS) and injected intraperitoneally to mice | [201] |
| CBS-knockout (KO) mice | ~25% of H2S -forming activity when no activity in presence of CSE inhibitor PAG | Total lack of CBS activity | Mouse liver, kidney, brainMonkey plasma (WT) | LC-MS/MS, HPLC | [202] |
5. Current Limitations and Future Studies
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Szabo, C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef]
- Turell, L.; Radi, R.; Alvarez, B. The thiol pool in human plasma: The central contribution of albumin to redox processes. Free Radic. Biol. Med. 2013, 65, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Hydrogen sulfide and persulfides oxidation by biologically relevant oxidizing species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevasanta, E.; Möller, M.N.; Alvarez, B. Biological chemistry of hydrogen sulfide and persulfides. Arch. Biochem. Biophys. 2017, 617, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M.; Fernandez, B.O.; Kelm, M.; Butler, A.R.; Feelisch, M. On the chemical biology of the nitrite/sulfide interaction. Nitric Oxide Biol. Chem. 2015, 46, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, P.M.; Batka, D.; Hefter, G.; Königsberger, E.; Rowland, D. Goodbye to S2- in aqueous solution. Chem. Commun. 2018, 54, 1980–1983. [Google Scholar] [CrossRef]
- Morse, J.W.; Millero, F.J.; Cornwell, J.C.; Rickard, D. The chemistry of the hydrogen sulfide and iron sulfide systems in natural waters. Earth Sci. Rev. 1987, 24, 1–42. [Google Scholar] [CrossRef]
- Li, Q.; Lancaster, J.R. Chemical foundations of hydrogen sulfide biology. Nitric Oxide Biol. Chem. 2013, 35, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Polhemus, D.J.; Lefer, D.J. Emergence of hydrogen sulfide as an endogenous gaseous signaling molecule in cardiovascular disease. Circ. Res. 2014, 114, 730–737. [Google Scholar] [CrossRef]
- Giuffrè, A.; Vicente, J.B. Hydrogen sulfide biochemistry and interplay with other gaseous mediators in mammalian physiology. Oxidative Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aroca, A.; Gotor, C.; Bassham, D.C. Hydrogen Sulfide: From a Toxic Molecule to a Key Molecule of Cell Life. Antioxidants 2020, 621. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, D.W.; Hannibal, L. Redox signaling in inherited diseases of metabolism. Curr. Opin. Physiol. 2019, 9, 48–55. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Kim, E. Cysteine-derived hydrogen sulfide and gut health: A matter of endogenous or bacterial origin. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Beard, R.S.; Bearden, S.E. Vascular complications of cystathionine β-synthase deficiency: Future directions for homocysteine-to-hydrogen sulfide research. Am. J. Physiol. Heart Circ. Physiol. 2011, 300. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.L.; Ritz, N.L.; Fauque, G.D.; Lin, H.C. Sulfur Cycling and the Intestinal Microbiome. Dig. Dis. Sci. 2017, 62, 2241–2257. [Google Scholar] [CrossRef]
- Dias-junior, C.A.; Sandrim, V.; Valeria, C. Circulating levels of hydrogen sulphide negatively correlate to nitrite levels in gestational hypertensive and preeclamptic pregnant women. Clin. Exp. Pharmacol. Physiol. 2021. [Google Scholar] [CrossRef]
- Suzuki, K.; Sagara, M.; Aoki, C.; Tanaka, S.; Aso, Y. Clinical implication of plasma hydrogen sulfide levels in Japanese patients with type 2 diabetes. Intern. Med. 2017, 56, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Kuang, Q.; Xue, N.; Chen, J.; Shen, Z.; Cui, X.; Fang, Y.; Ding, X. Low Plasma Hydrogen Sulfide Is Associated with Impaired Renal Function and Cardiac Dysfunction. Am. J. Nephrol. 2018, 47, 361–371. [Google Scholar] [CrossRef]
- Karunya, R.; Jayaprakash, K.S.; Gaikwad, R.; Sajeesh, P.; Ramshad, K.; Muraleedharan, K.M.; Dixit, M.; Thangaraj, P.R.; Sen, A.K. Rapid measurement of hydrogen sulphide in human blood plasma using a microfluidic method. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Du, S.X.; Xiao, J.; Guan, F.; Sun, L.M.; Wu, W.S.; Tang, H.; Du, J.B.; Tang, C.S.; Jin, H.F. Predictive role of cerebrospinal fluid hydrogen sulfide in central nervous system leukemia. Chin. Med. J. 2011, 124, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Neri, C.; Pieragostino, D.; Spalloni, A.; Persichilli, S.; Gastaldi, M.; Mercuri, N.B.; Longone, P.; Urbani, A. Investigating different forms of hydrogen sulfide in cerebrospinal fluid of various neurological disorders. Metabolites 2021, 11, 152. [Google Scholar] [CrossRef]
- Olson, K.R. Is hydrogen sulfide a circulating “gasotransmitter” in vertebrate blood? Biochim. Biophys. Acta Bioenerg. 2009, 1787, 856–863. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R.; Deleon, E.R.; Liu, F. Controversies and conundrums in hydrogen sulfide biology. Nitric Oxide Biol. Chem. 2014, 41, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Kolluru, G.K.; Shen, X.; Bir, S.C.; Kevil, C.G. Hydrogen sulfide chemical biology: Pathophysiological roles and detection. Nitric Oxide Biol. Chem. 2013, 35, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, P.; Pálinkás, Z.; Nagy, A.; Budai, B.; Tóth, I.; Vasas, A. Chemical aspects of hydrogen sulfide measurements in physiological samples. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Ditrói, T.; Nagy, A.; Martinelli, D.; Rosta, A.; Kožich, V.; Nagy, P. Comprehensive analysis of how experimental parameters affect H 2 S measurements by the monobromobimane method. Free Radic. Biol. Med. 2019, 136, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Cuevasanta, E.; Denicola, A.; Alvarez, B.; Möller, M.N. Solubility and permeation of hydrogen sulfide in lipid membranes. PLoS ONE 2012, 7, 3–8. [Google Scholar] [CrossRef]
- Möller, M.N.; Cuevasanta, E.; Orrico, F.; Lopez, A.C.; Thomson, L.; Denicola, A. Diffusion and transport of reactive species across cell membranes. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2019; Volume 1127, pp. 3–19. [Google Scholar]
- Olson, K.R. A theoretical examination of hydrogen sulfide metabolism and its potential in autocrine/paracrine oxygen sensing. Respir. Physiol. Neurobiol. 2013, 186, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.L. Transport of H2S and HS- across the human red blood cell membrane: Rapid H2S diffusion and AE1-mediated Cl-/HS- exchange. Am. J. Physiol. Cell Physiol. 2013, 305. [Google Scholar] [CrossRef] [Green Version]
- Furne, J.; Saeed, A.; Levitt, M.D. Whole tissue hydrogen sulfide concentrations are orders of magnitude lower than presently accepted values. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Peter, E.A.; Bir, S.; Wang, R.; Kevil, C.G. Analytical measurement of discrete hydrogen sulfide pools in biological specimens. Free Radic. Biol. Med. 2012, 52, 2276–2283. [Google Scholar] [CrossRef] [Green Version]
- Levitt, M.D.; Abdel-Rehim, M.S.; Furne, J. Free and acid-labile hydrogen sulfide concentrations in mouse tissues: Anomalously high free hydrogen sulfide in aortic tissue. Antioxid. Redox Signal. 2011, 15, 373–378. [Google Scholar] [CrossRef]
- Wintner, E.A.; Deckwerth, T.L.; Langston, W.; Bengtsson, A.; Leviten, D.; Hill, P.; Insko, M.A.; Dumpit, R.; Vandenekart, E.; Toombs, C.F.; et al. A monobromobimane-based assay to measure the pharmacokinetic profile of reactive sulphide species in blood. Br. J. Pharmacol. 2010, 160, 941–957. [Google Scholar] [CrossRef] [Green Version]
- Doeller, J.E.; Isbell, T.S.; Benavides, G.; Koenitzer, J.; Patel, H.; Patel, R.P.; Lancaster, J.R.; Darley-Usmar, V.M.; Kraus, D.W. Polarographic measurement of hydrogen sulfide production and consumption by mammalian tissues. Anal. Biochem. 2005, 341, 40–51. [Google Scholar] [CrossRef]
- Ubuka, T.; Abe, T.; Kajikawa, R.; Morino, K. Determination of hydrogen sulfide and acid-labile sulfur in animal tissues by gas chromatography and ion chromatography. J. Chromatogr. B Biomed. Sci. Appl. 2001, 757, 31–37. [Google Scholar] [CrossRef]
- Ishigami, M.; Hiraki, K.; Umemura, K.; Ogasawara, Y.; Ishii, K.; Kimura, H. A source of hydrogen sulfide and a mechanism of its release in the brain. Antioxid. Redox Signal. 2009, 11, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangerman, A.; Meuwese-Arends, M.T.; Van Tongeren, J.H.M. New methods for the release of volatile sulfur compounds from human serum: Its determination by Tenax trapping and gas chromatography and its application in liver diseases. J. Lab. Clin. Med. 1985, 106, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Zaorska, E.; Konop, M.; Ostaszewski, R.; Koszelewski, D.; Ufnal, M. Salivary hydrogen sulfide measured with a new highly sensitive self-immolative coumarin-based fluorescent probe. Molecules 2018, 23, 2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, L.R.; Francom, D.; Dieken, F.P.; Taylor, J.D.; Warenycia, M.W.; Reiffenstein, R.J.; Dowling, G. Determination of Sulfide in Brain Tissue by Gas Dialysis/Ion Chromatography: Postmortem Studies and Two Case Reports. J. Anal. Toxicol. 1989, 13, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Kožich, V.; Ditrói, T.; Sokolová, J.; Křížková, M.; Krijt, J.; Ješina, P.; Nagy, P. Metabolism of sulfur compounds in homocystinurias. Br. J. Pharmacol. 2019, 176, 594–606. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.H.; Wong, P.T.; Bian, J. Neurochemistry International Hydrogen sulfide: A novel signaling molecule in the central nervous system. Neurochem. Int. 2010, 56, 3–10. [Google Scholar] [CrossRef]
- Attene-Ramos, M.S.; Wagner, E.D.; Plewa, M.J.; Gaskins, H.R. Evidence that hydrogen sulfide is a genotoxic agent. Mol. Cancer Res. 2006, 4, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Fiorucci, S.; Antonelli, E.; Distrutti, E.; Rizzo, G.; Mencarelli, A.; Orlandi, S.; Zanardo, R.; Renga, B.; Di Sante, M.; Morelli, A.; et al. Inhibition of hydrogen sulfide generation contributes to gastric injury caused by anti-inflammatory nonsteroidal drugs. Gastroenterology 2005, 129, 1210–1224. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Dargusch, R.; Schubert, D.; Kimura, H. Hydrogen sulfide protects HT22 neuronal cells from oxidative stress. Antioxid. Redox Signal. 2006, 8, 661–670. [Google Scholar] [CrossRef]
- Szabo, C. Hydrogen Sulfide, an Endogenous Stimulator of Mitochondrial Function in Cancer Cells. Cells 2021, 10, 220. [Google Scholar] [CrossRef]
- Kimura, Y.; Kimura, H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 2004, 18, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Q.; Xin, H.; Zhu, Y.Z. Hydrogen sulfide: Third gaseous transmitter, but with great pharmacological potential. Acta Pharmacol. Sin. 2007, 28, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Sun, X.; Wang, R. Hydrogen sulfide-induced apoptosis of human aorta smooth muscle cells via the activation of mitogen-activated protein kinases and caspase-3. FASEB J. 2004, 18, 1782–1784. [Google Scholar] [CrossRef] [PubMed]
- Renga, B. Hydrogen sulfide generation in mammals: The molecular biology of cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE). Inflamm. Allergy Drug Targets 2011, 10, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Le Trionnaire, S.L.; Chopra, M.; Fox, B.; Whatmore, J. Emerging role of hydrogen sulfide in health and disease: Critical appraisal of biomarkers and pharmacological tools. Clin. Sci. 2011, 121, 459–488. [Google Scholar] [CrossRef]
- Perry, M.M.; Hui, C.K.; Whiteman, M.; Wood, M.E.; Adcock, I.; Kirkham, P.; Michaeloudes, C.; Chung, K.F. Hydrogen sulfide inhibits proliferation and release of IL-8 from human airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2011, 45, 746–752. [Google Scholar] [CrossRef]
- Yang, W.; Yang, G.; Jia, X.; Wu, L.; Wang, R. Activation of KATP channels by H2S in rat insulin-secreting cells and the underlying mechanisms. J. Physiol. 2005, 569, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, M.; Huat, B.T.K.; Hsu, A.; Whiteman, M.; Bhatia, M.; Moore, P.K. Streptozotocin-induced diabetes in the rat is associated with enhanced tissue hydrogen sulfide biosynthesis. Biochem. Biophys. Res. Commun. 2005, 333, 1146–1152. [Google Scholar] [CrossRef]
- Medeiros, J.V.R.; Bezerra, V.H.; Gomes, A.S.; Barbosa, A.L.R.; Lima, R.C.P.; Soares, P.M.G.; Brito, G.A.C.; Ribeiro, R.A.; Cunha, F.Q.; De Souza, M.H.L.P. Hydrogen sulfide prevents ethanol-induced gastric damage in mice: Role of ATP-sensitive potassium channels and capsaicin-sensitive primary afferent neurons. J. Pharmacol. Exp. Ther. 2009, 330, 764–770. [Google Scholar] [CrossRef]
- Deplancke, B.; Gaskins, H.R. Hydrogen sulfide induces serum-independent cell cycle entry in nontransformed rat intestinal epithelial cells. FASEB J. 2003, 17, 1310–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.; Ellis, C.J.; Furne, J.K.; Springfield, J.; Levitt, M.D. Fecal hydrogen sulfide production in ulcerative colitis. Am. J. Gastroenterol. 1998, 93, 83–87. [Google Scholar] [CrossRef]
- Bogdándi, V.; Ida, T.; Sutton, T.R.; Bianco, C.; Ditrói, T.; Koster, G.; Henthorn, H.A.; Minnion, M.; Toscano, J.P.; van der Vliet, A.; et al. Speciation of reactive sulfur species and their reactions with alkylating agents: Do we have any clue about what is present inside the cell? Br. J. Pharmacol. 2019, 176, 646–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, K.R. H 2 S and polysulfide metabolism: Conventional and unconventional pathways. Biochem. Pharmacol. 2018, 149, 77–90. [Google Scholar] [CrossRef]
- Beus, M.D.D.E.; Chung, J. Modification of cysteine 111 in Cu / Zn superoxide dismutase results in altered spectroscopic and biophysical properties. Protein Sci. 2004, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Ogasawara, Y.; Shibuya, N.; Kimura, H.; Ishii, K. Polysulfide exerts a protective effect against cytotoxicity caused by t -buthylhydroperoxide through Nrf2 signaling in neuroblastoma cells. FEBS Lett. 2013, 587, 3548–3555. [Google Scholar] [CrossRef] [Green Version]
- Marcia, M.; Ermler, U.; Peng, G.; Michel, H. The structure of Aquifex aeolicus sulfide: Quinone oxidoreductase, a basis to understand sulfide detoxification and respiration. Proc. Natl. Acad. Sci. USA 2009, 106, 9625–9630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, T.M.; Grieshaber, M.K. Three enzymatic activities catalyze the oxidation of sulfide to thiosulfate in mammalian and invertebrate mitochondria. FEBS J. 2008, 275, 3352–3361. [Google Scholar] [CrossRef]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the human mitochondrial hydrogen sulfide oxidation pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, A.P.; Ballou, D.P.; Banerjee, R. Hydrogen Sulfide Oxidation by Sulfide Quinone Oxidoreductase. ChemBioChem 2020, cbic.202000661. [Google Scholar] [CrossRef] [PubMed]
- Makarov, S.V.; Horváth, A.K.; Makarova, A.S. Reactivity of Small Oxoacids of Sulfur. Molecules 2019, 24, 2768. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.R.; Farmer, P.J. Chemical trapping and characterization of small oxoacids of sulfur (SOS) generated in aqueous oxidations of H2S. Redox Biol. 2018, 14, 485–491. [Google Scholar] [CrossRef]
- Makarov, S.V.; Makarova, A.S.; Silaghi-Dumitrescu, R. Sulfoxylic and Thiosulfurous Acids and their Dialkoxy Derivatives. In Patai’s Chemistry of Functional Groups, Peroxides; Liebman, J., Greer, A., Eds.; Wiley: Hoboken, NJ, USA, 2014; Volume 3, Part I; pp. 265–305. [Google Scholar]
- Kabil, O.; Banerjee, R. Enzymology of H2S biogenesis, decay and signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Hannibal, L.; Blom, H.J. Homocysteine and disease: Causal associations or epiphenomenons? Mol. Asp. Med. 2017, 53, 36–42. [Google Scholar] [CrossRef]
- Gregory, J.F.; DeRatt, B.N.; Rios-Avila, L.; Ralat, M.; Stacpoole, P.W. Vitamin B6 nutritional status and cellular availability of pyridoxal 5′-phosphate govern the function of the transsulfuration pathway’s canonical reactions and hydrogen sulfide production via side reactions. Biochimie 2016, 126, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Kožich, V.; Krijt, J.; Sokolová, J.; Melenovská, P.; Ješina, P.; Vozdek, R.; Majtán, T.; Kraus, J.P. Thioethers as markers of hydrogen sulfide production in homocystinurias. Biochimie 2016, 126, 14–20. [Google Scholar] [CrossRef]
- Chen, X.; Jhee, K.H.; Kruger, W.D. Production of the neuromodulator H2S by cystathionine β-synthase via the condensation of cysteine and homocysteine. J. Biol. Chem. 2004, 279, 52082–52086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, M.; Janosik, M.; Kery, V.; Kraus, J.P.; Burkhard, P. Structure of human cystathionine beta-synthase: A unique pyridoxal 5’-phosphate-dependent heme protein. EMBO J. 2001, 20, 3910–3916. [Google Scholar] [CrossRef] [Green Version]
- Kabil, O.; Zhou, Y.; Banerjee, R. Human cystathionine β-synthase is a target for sumoylation. Biochemistry 2006, 45, 13528–13536. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Saha, S.; Giri, K.; Lanza, I.R.; Nair, K.S.; Jennings, N.B.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Basal, E.; Weaver, A.L.; et al. Cystathionine Beta-Synthase (CBS) Contributes to Advanced Ovarian Cancer Progression and Drug Resistance. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative contributions of cystathionine β-synthase and γ-cystathionase to H2S biogenesis via alternative trans-sulfuration reactions. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanishi, M.; Kabil, O.; Sen, S.; Banerjee, R. Structural insights into pathogenic mutations in heme-dependent cystathionine-β-synthase. J. Inorg. Biochem. 2006, 100, 1988–1995. [Google Scholar] [CrossRef]
- Ereño-Orbea, J.; Majtan, T.; Oyenarte, I.; Kraus, J.P.; Martínez-Cruz, L.A. Structural basis of regulation and oligomerization of human cystathionine β-synthase, the central enzyme of transsulfuration. Proc. Natl. Acad. Sci. USA 2013, 110, E3790–E3799. [Google Scholar] [CrossRef] [Green Version]
- Majtan, T.; Singh, L.R.; Wang, L.; Kruger, W.D.; Kraus, J.P. Active cystathionine β-synthase can be expressed in heme-free systems in the presence of metal-substituted porphyrins or a chemical chaperone. J. Biol. Chem. 2008, 283, 34588–34595. [Google Scholar] [CrossRef] [Green Version]
- Taoka, S.; Banerjee, R. Characterization of NO binding to human cystathionine β-synthase: Possible implications of the effects of CO and NO binding to the human enzyme. J. Inorg. Biochem. 2001, 87, 245–251. [Google Scholar] [CrossRef]
- Jhee, K.H.; McPhie, P.; Miles, E.W. Domain architecture of the heme-independent yeast cystathionine β-synthase provides insights into mechanisms of catalysis and regulation. Biochemistry 2000, 39, 10548–10556. [Google Scholar] [CrossRef] [PubMed]
- Hayes, L.C.; Plapp, F.V.; Tilzer, L.L. Activation of cystathionine synthase by adenosylmethionine and adenosylethionine. Biochem. Biophys. Res. Commun. 1975, 65, 258–264. [Google Scholar] [CrossRef]
- Shafqat, N.; Muniz, J.R.C.; Pilka, E.S.; Papagrigoriou, E.; Von Delft, F.; Oppermann, U.; Yue, W.W. Insight into S-Adenosylmethionine biosynthesis from the crystal structures of the human methionine adenosyltransferase catalytic and regulatory subunits. Biochem. J. 2013, 452, 27–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudova, A.; Bauman, Z.; Braun, A.; Vitvitsky, V.; Lu, S.C.; Banerjee, R. S-adenosylmethionine stabilizes cystathionine β-synthase and modulates redox capacity. Proc. Natl. Acad. Sci. USA 2006, 103, 6489–6494. [Google Scholar] [CrossRef] [Green Version]
- Majtan, T.; Liu, L.; Carpenter, J.F.; Kraus, J.P. Rescue of cystathionine β-synthase (CBS) mutants with chemical chaperones: Purification and characterization of eight CBS mutant enzymes. J. Biol. Chem. 2010, 285, 15866–15873. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.B.; Colaço, H.G.; Sarti, P.; Leandro, P.; Giuffrè, A. S-adenosyl-L-methionine modulates CO and NO· binding to the human H2S-generating enzyme cystathionine β-synthase. J. Biol. Chem. 2016, 291, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, N.; Banerjee, R. Human polycomb 2 protein is a SUMO E3 ligase and alleviates substrate-induced inhibition of cystathionine β-synthase sumoylation. PLoS ONE 2008, 3, 1–7. [Google Scholar] [CrossRef]
- Diwakar, L.; Kenchappa, R.S.; Annepu, J.; Ravindranath, V. Downregulation of glutaredoxin but not glutathione loss leads to mitochondrial dysfunction in female mice CNS: Implications in excitotoxicity. Neurochem. Int. 2007, 51, 37–46. [Google Scholar] [CrossRef]
- Hosoki, R.; Matsuki, N.; Kimura, H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 1997, 237, 527–531. [Google Scholar] [CrossRef]
- Kaneko, Y.; Kimura, Y.; Kimura, H.; Niki, I. L-cysteine inhibits insulin release from the pancreatic β-cell: Possible involvement of metabolic production of hydrogen sulfide, a novel gasotransmitter. Diabetes 2006, 55, 1391–1397. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Vatish, M.; Heptinstall, J.; Wang, R.; Carson, R.J. The endogenous production of hydrogen sulphide in intrauterine tissues. Reprod. Biol. Endocrinol. 2009, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitvitsky, V.; Thomas, M.; Ghorpade, A.; Gendelman, H.E.; Banerjee, R. A functional transsulfuration pathway in the brain links to glutathione homeostasis. J. Biol. Chem. 2006, 281, 35785–35793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Bhatia, M.; Moore, P.K. Hydrogen sulphide—A novel mediator of inflammation? Curr. Opin. Pharmacol. 2006, 6, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine γ-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiku, T.; Padovani, D.; Zhu, W.; Singh, S.; Vitvitsky, V.; Banerjee, R. H2S biogenesis by human cystathionine γ-lyase leads to the novel sulfur metabolites lanthionine and homolanthionine and is responsive to the grade of hyperhomocysteinemia. J. Biol. Chem. 2009, 284, 11601–11612. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; He, G.W. Imbalance of Homocysteine and H2S: Significance, Mechanisms, and Therapeutic Promise in Vascular Injury. Oxid. Med. Cell. Longev. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Banerjee, R. PLP-dependent H 2S biogenesis. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Nagahara, N.; Nishino, T. Role of amino acid residues in the active site of rat liver mercaptopyruvate sulfurtransferase: cDNA cloning, overexpression, and site- directed mutagenesis. J. Biol. Chem. 1996, 271, 27395–27401. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, N.; Tanaka, M.; Yoshida, M.; Ogasawara, Y.; Togawa, T.; Ishii, K.; Kimura, H. 3-Mercaptopyruvate sulfurtransferase produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid. Redox Signal. 2009, 11, 703–714. [Google Scholar] [CrossRef]
- Vitvitsky, V.; Yadav, P.K.; Kurthen, A.; Banerjee, R. Sulfide oxidation by a noncanonical pathway in red blood cells generates thiosulfate and polysulfides. J. Biol. Chem. 2015, 290, 8310–8320. [Google Scholar] [CrossRef] [Green Version]
- Fräsdorf, B.; Radon, C.; Leimkühler, S. Characterization and interaction studies of two isoforms of the dual localized 3-mercaptopyruvate sulfurtransferase TUM1 from humans. J. Biol. Chem. 2014, 289, 34543–34556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Goto, Y.I.; Kimura, H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxid. Redox Signal. 2010, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide: Its production, release and functions. Amino Acids 2011, 41, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.K.; Vitvitsky, V.; Carballal, S.; Seravalli, J.; Banerjee, R. Thioredoxin regulates human mercaptopyruvate sulfurtransferase at physiologically-relevant concentrations. J. Biol. Chem. 2020, 295, 6299–6310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
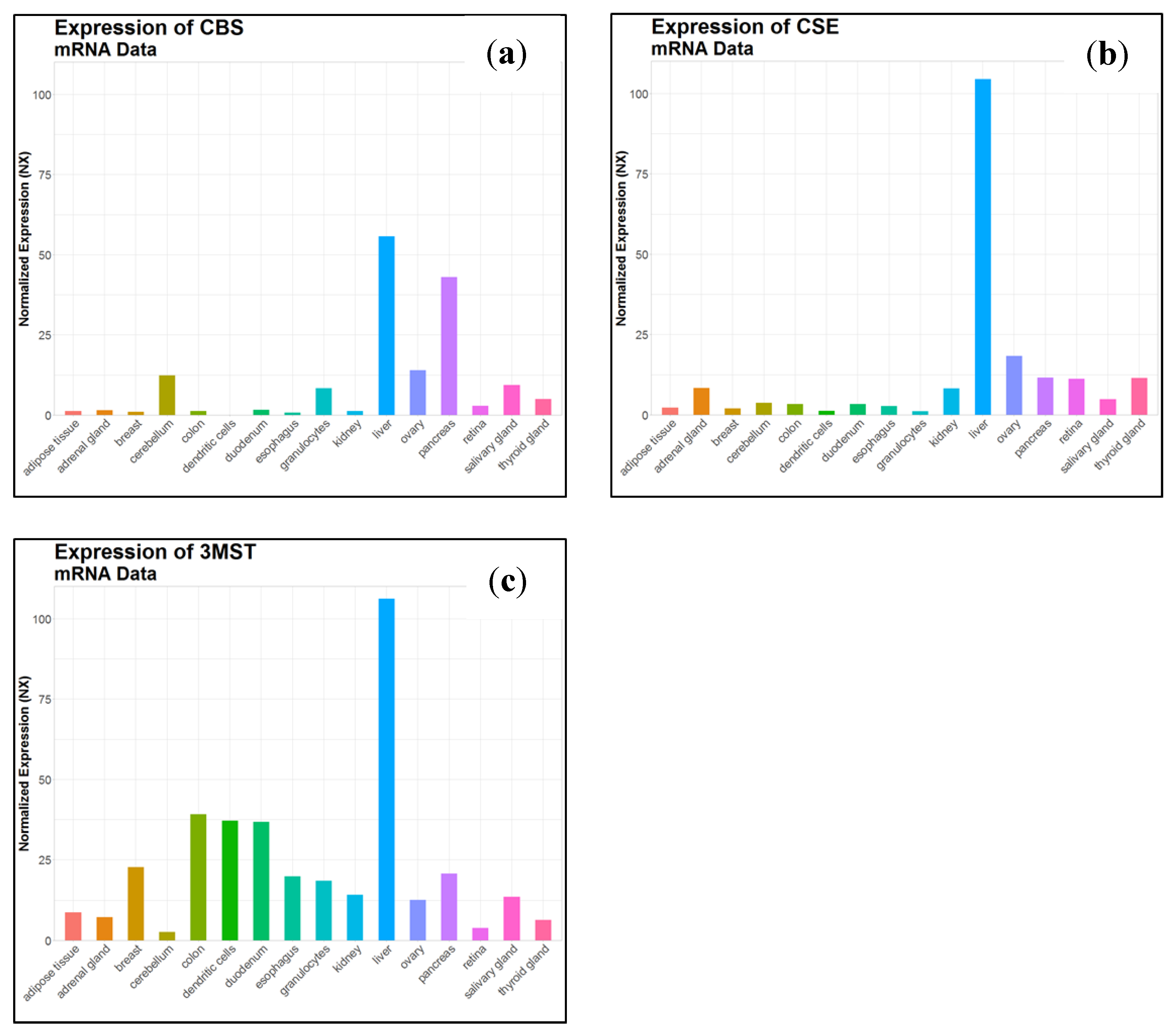
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347. [Google Scholar] [CrossRef]
- Kohl, J.B.; Mellis, A.; Schwarz, G. Homeostatic impact of sulfite and hydrogen sulfide on cysteine catabolism. Br. J. Pharmacol. 2019, 554–570. [Google Scholar] [CrossRef] [Green Version]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [Green Version]
- Searcy, D.G.; Lee, S.H. Sulfur reduction by human erythrocytes. J. Exp. Zool. 1998, 282, 310–322. [Google Scholar] [CrossRef]
- Masi, A.D.; Ascenzi, P. H2S: A “Double face” molecule in health and disease. BioFactors 2013, 39, 186–196. [Google Scholar] [CrossRef]
- Akaike, T.; Ida, T.; Wei, F.Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S.; et al. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dóka, É.; Pader, I.; Bíró, A.; Johansson, K.; Cheng, Q.; Ballagó, K.; Prigge, J.R.; Pastor-Flores, D.; Dick, T.P.; Schmidt, E.E.; et al. A novel persulfide detection method reveals protein persulfide- and polysulfide-reducing functions of thioredoxin and glutathione systems. Sci. Adv. 2016, 2, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, C.L.; Savitsky, A.; Feelisch, M.; Cortese-Krott, M.M. Investigations on the role of hemoglobin in sulfide metabolism by intact human red blood cells. Biochem. Pharmacol. 2018, 149, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Cortese-Krott, M.M. Red blood cells as a “central hub” for sulfide bioactivity: Scavenging, metabolism, transport, and cross-talk with nitric oxide. Antioxid. Redox Signal. 2020, 33, 1332–1349. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.F.; Yu, T.C.; Hong, J.; Fang, J.Y. Emerging roles of hydrogen sulfide in inflammatory and neoplastic colonic diseases. Front. Physiol. 2016, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hale, V.L.; Jeraldo, P.; Mundy, M.; Yao, J.; Keeney, G.; Cheek, E.H.; Davidson, J.; Green, M.; Martinez, C.; Pettry, C.; et al. Synthesis of multi-omic data and community metabolic models reveals insights into the role of hydrogen sulfide in colon cancer. HHS Public Access 2018, 68. [Google Scholar] [CrossRef]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut bacteria and hydrogen sulfide: The new old players in circulatory system homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef]
- Seiflein, T.A.; Lawrence, J.G. Methionine-to-cysteine recycling in Klebsiella aerogenes. J. Bacteriol. 2001, 183, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Basic, A.; Blomqvist, M.; Dahlén, G.; Svensäter, G. The proteins of Fusobacterium spp. involved in hydrogen sulfide production from L-cysteine. BMC Microbiol. 2017, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Carlström, M.; Borniquel, S.; Jädert, C.; Kevil, C.G.; Lundberg, J.O. Microbial regulation of host hydrogen sulfide bioavailability and metabolism. Free Radic. Biol. Med. 2013, 60, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Ritz, N.L.; Burnett, B.J.; Setty, P.; Reinhart, K.M.; Wilson, M.R.; Alcock, J.; Singh, S.B.; Barton, L.L.; Lin, H.C. Sulfate-reducing bacteria impairs working memory in mice. Physiol. Behav. 2016, 157, 281–287. [Google Scholar] [CrossRef]
- Linden, D.R. Hydrogen sulfide signaling in the gastrointestinal tract. Antioxid. Redox Signal. 2014, 20, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Linden, D.R.; Furne, J.; Stoltz, G.J.; Abdel-Rehim, M.S.; Levitt, M.D.; Szurszewski, J.H. Sulphide quinone reductase contributes to hydrogen sulphide metabolism in murine peripheral tissues but not in the CNS. Br. J. Pharmacol. 2012, 165, 2178–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Human sulfide:Quinone oxidoreductase catalyzes the first step in hydrogen sulfide metabolism and produces a sulfane sulfur metabolite. Biochemistry 2012, 51, 6804–6815. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Characterization of patient mutations in human persulfide dioxygenase (ETHE1) involved in H2S catabolism. J. Biol. Chem. 2012, 287, 44561–44667. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R. Mitochondrial adaptations to utilize hydrogen sulfide for energy and signaling. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2012, 182, 881–897. [Google Scholar] [CrossRef] [PubMed]
- Schrader, N.; Fischer, K.; Theis, K.; Mendel, R.R.; Schwarz, G.; Kisker, C. The crystal structure of plant sulfite oxidase provides insights into sulfite oxidation in plants and animals. Structure 2003, 11, 1251–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, G.; Mendel, R.R.; Ribbe, M.W. Molybdenum cofactors, enzymes and pathways. Nature 2009, 460, 839–847. [Google Scholar] [CrossRef]
- Friederich, M.W.; Elias, A.F.; Kuster, A.; Laugwitz, L.; Larson, A.A.; Landry, A.P.; Ellwood-Digel, L.; Mirsky, D.M.; Dimmock, D.; Haven, J.; et al. Pathogenic variants in SQOR encoding sulfide:quinone oxidoreductase are a potentially treatable cause of Leigh disease. J. Inherit. Metab. Dis. 2020, 43, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aminlari, M.; Gilanpour, H. Comparative studies on the distribution of rhodanese in different tissues of domestic animals. Comp. Biochem. Physiol. Part B Biochem. 1991, 99, 673–677. [Google Scholar] [CrossRef]
- Furne, J.; Springfield, J.; Koenig, T.; DeMaster, E.; Levitt, M.D. Oxidation of hydrogen sulfide and methanethiol to thiosulfate by rat tissues: A specialized function of the colonic mucosa. Biochem. Pharmacol. 2001, 62, 255–259. [Google Scholar] [CrossRef]
- Libiad, M.; Sriraman, A.; Banerjee, R. Polymorphic variants of human rhodanese exhibit differences in thermal stability and sulfur transfer kinetics. J. Biol. Chem. 2015, 290, 23579–23588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzi, M.; Wittenberg, J.B.; Coda, A.; Fasano, M.; Ascenzi, P.; Bolognesi, M. Structure of the sulfide-reactive hemoglobin from the clam Lucina pectinata: Crystallographic analysis at 1.5 Å resolution. J. Mol. Biol. 1994, 244, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bellelli, A.; Coletta, M.; Colosimo, A.; Falcioni, G.; Giacometti, G.M.; Ippoliti, R.; Zolla, L.; Giardina, B. Multiple strategies for O2 transport: From simplicity to complexity. IUBMB Life 2007, 59, 600–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, R.O.; Bus, J.S.; Popp, J.A.; Boreiko, C.J.; Andjelkovich, D.A.; Leber, P. A critical review of the literature on hydrogen sulfide toxicity. Crit. Rev. Toxicol. 1984, 13, 25–97. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Isoda, S.; Tanabe, S. Tissue and Subcellular Distribution of Bound and Acid-Labile Sulfur, and the Enzymic Capacity for Sulfide Production in the Rat. Biol. Pharm. Bull. 1994, 17, 1535–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toohey, J.I. Sulphane sulphur in biological systems: A possible regulatory role. Biochem. J. 1989, 264, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Zhao, K.; Ju, Y.; Mani, S.; Cao, Q.; Puukila, S.; Khaper, N.; Wu, L.; Wang, R. Hydrogen sulfide protects against cellular senescence via s-sulfhydration of keap1 and activation of Nrf2. Antioxid. Redox Signal. 2013, 18, 1906–1919. [Google Scholar] [CrossRef]
- Hourihan, J.M.; Kenna, J.G.; Hayes, J.D. The gasotransmitter hydrogen sulfide induces Nrf2-target genes by inactivating the keap1 ubiquitin ligase substrate adaptor through formation of a disulfide bond between Cys-226 and Cys-613. Antioxid. Redox Signal. 2013, 19, 465–481. [Google Scholar] [CrossRef]
- Xie, L.; Gu, Y.; Wen, M.; Zhao, S.; Wang, W.; Ma, Y.; Meng, G.; Han, Y.; Wang, Y.; Liu, G.; et al. Hydrogen sulfide induces Keap1 S-sulfhydration and suppresses diabetes-accelerated atherosclerosis via Nrf2 activation. Diabetes 2016, 65, 3171–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Liu, K.; He, J.; Tian, C.; Yu, X.; Yang, J. Direct Proteomic Mapping of Cysteine Persulfidation. Antioxid. Redox Signal. 2020, 33, 1061–1076. [Google Scholar] [CrossRef] [PubMed]
- Benchoam, D.; Semelak, J.A.; Cuevasanta, E.; Mastrogiovanni, M.; Grassano, J.S.; Ferrer-Sueta, G.; Zeida, A.; Trujillo, M.; Möller, M.N.; Estrin, D.A.; et al. Acidity and nucleophilic reactivity of glutathione persulfide. J. Biol. Chem. 2020, 295, 15466–15481. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Zivanovic, J.; Alvarez, B.; Banerjee, R. Chemical Biology of H2S Signaling through Persulfidation. Chem. Rev. 2018, 118, 1253–1337. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H.; Kashfi, K. Effects of hydrogen sulfide on mitochondrial function and cellular bioenergetics. Redox Biol. 2021, 38. [Google Scholar] [CrossRef] [PubMed]
- Fukuto, J.M.; Vega, V.S.; Works, C.; Lin, J. The chemical biology of hydrogen sulfide and related hydropersulfides: Interactions with biologically relevant metals and metalloproteins. Curr. Opin. Chem. Biol. 2020, 55, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, P.; Kim, J.K. Sulphide as an inhibitor and electron donor for the cytochrome c oxidase system. Can. J. Biochem. 1982, 60, 613–623. [Google Scholar] [CrossRef]
- Scammahorn, J.J.; Nguyen, I.T.N.; Bos, E.M.; van Goor, H.; Joles, J.A. Fighting oxidative stress with sulfur: Hydrogen sulfide in the renal and cardiovascular systems. Antioxidants 2021, 10, 373. [Google Scholar] [CrossRef]
- Olson, K.R.; Gao, Y.; Briggs, A.; Devireddy, M.; Iovino, N.A.; Licursi, M.; Skora, N.C.; Whelan, J.; Villa, B.P.; Straub, K.D. ‘Antioxidant’ berries, anthocyanins, resveratrol and rosmarinic acid oxidize hydrogen sulfide to polysulfides and thiosulfate: A novel mechanism underlying their biological actions. Free Radic. Biol. Med. 2021, 165, 67–78. [Google Scholar] [CrossRef]
- Xie, Z.Z.; Liu, Y.; Bian, J.S. Hydrogen Sulfide and Cellular Redox Homeostasis. Oxidative Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Casin, K.M.; Calvert, J.W. Harnessing the benefits of endogenous hydrogen sulfide to reduce cardiovascular disease. Antioxidants 2021, 10, 383. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, A.H.; Ahmad, F.U.D.; Furwa, K.; Madni, A. Role of oxidative stress and reduced endogenous hydrogen sulfide in diabetic nephropathy. Drug Des. Dev. Ther. 2021, 15, 1031–1043. [Google Scholar] [CrossRef]
- Dillon, K.M.; Matson, J.B. A Review of Chemical Tools for Studying Small Molecule Persulfides: Detection and Delivery. ACS Chem. Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Dulac, M.; Melet, A.; Galardon, E. Reversible Detection and Quantification of Hydrogen Sulfide by Fluorescence Using the Hemoglobin i from Lucina pectinata. ACS Sens. 2018, 3, 2138–2144. [Google Scholar] [CrossRef]
- Guenther, E.A.; Johnson, K.S.; Coale, K.H. Direct ultraviolet spectrophotometric determination of total sulfide and iodide in natural waters. Anal. Chem. 2001, 73, 3481–3487. [Google Scholar] [CrossRef] [PubMed]
- Boffi, A.; Rizzi, M.; Monacelli, F.; Ascenzi, P. Determination of H2S solubility via the reaction with ferric hemoglobin I from the bivalve mollusc Lucina pectinata. Biochim. Biophys. Acta Gen. Subj. 2000, 1523, 206–208. [Google Scholar] [CrossRef]
- Budd, M.S.; Bewick, H.A. Photometric Determination of Sulfide and Reducible Sulfur in Alkalies. Anal. Chem. 1952, 24, 1536–1540. [Google Scholar] [CrossRef]
- Sheppard, S.E.; Geddes, A.L. Effect of Solvents upon the Absorption Spectra of Dyes. V. Water as Solvent: Quantitative Examination of the Dimerization Hypothesis. J. Am. Chem. Soc. 1944, 66, 2003–2009. [Google Scholar] [CrossRef]
- Pomeroy, R. The determination of sulphides in sewage. Sew. Work. J. 1936, 8, 572–591. [Google Scholar]
- Kovatsis, A.; Tsougas, M. Determination of hydrogen sulfide (H2S) in environment by indirect atomic absorption spectroscopy. Bull. Environ. Contam. Toxicol. 1976, 15, 412–420. [Google Scholar] [CrossRef]
- Fischer, E. Bildung von Methylenblau als Reaktion auf Schwefelwasserstoff. Ber. Dtsch. Chem. Ges. 1883, 16, 2234–2236. [Google Scholar] [CrossRef]
- Sakurai, H.; Lien, S.; Pietro, A.S. Determination of acid-labile sulfide and zero-valence sulfur in subchloroplast particles in the presence of sodium dodecyl sulfate. Anal. Biochem. 1982, 119, 372–377. [Google Scholar] [CrossRef]
- Mylon, S.E.; Benoit, G. Subnanomolar detection of acid-labile sulfides by the classical methylene blue method coupled to HPLC. Environ. Sci. Technol. 2001, 35, 4544–4548. [Google Scholar] [CrossRef] [PubMed]
- Haddad, P.R.; Heckenberg, A.L. Trace determination of sulfide by reversed-phase ion- interaction chromatography using pre-column derivatization. J. Chromatogr. 1988, 447, 415–420. [Google Scholar] [CrossRef]
- Small, J.M.; Hintelmann, H. Methylene blue derivatization then LC-MS analysis for measurement of trace levels of sulfide in aquatic samples. Anal. Bioanal. Chem. 2007, 387, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Pattillo, C.B.; Pardue, S.; Bir, S.C.; Wang, R.; Kevil, C.G. Measurement of plasma hydrogen sulfide in vivo and in vitro. Free Radic. Biol. Med. 2011, 50, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Kraus, D.; Wittenberg, J.B. Hemoglobins of the Lucina pectinatu/Bacteria Symbiosis. J. Biol. Chem. 1990, 265, 16043–16053. [Google Scholar] [CrossRef]
- Collazo, E.; Pietri, R.; De Jesús, W.; Ramos, C.; Del Toro, A.; León, R.G.; Cadilla, C.L.; López-Garriga, J. Functional Characterization of the Purified Holo Form of Hemoglobin I from Lucina pectinata Overexpressed in Escherichia coli. Protein J. 2004, 23, 239–245. [Google Scholar] [CrossRef]
- Ramos-Alvarez, C.; Yoo, B.K.; Pietri, R.; Lamarre, I.; Martin, J.L.; Lopez-Garriga, J.; Negrerie, M. Reactivity and dynamics of H2S, NO, and O2 interacting with hemoglobins from Lucina pectinata. Biochemistry 2013, 52, 7007–7021. [Google Scholar] [CrossRef]
- León, R.G.; Munier-Lehmann, H.; Barzu, O.; Baudin-Creuza, V.; Pietri, R.; López-Garriga, J.; Cadilla, C.L. High-level production of recombinant sulfide-reactive hemoglobin I from Lucina pectinata in Escherichia coli: High yields of fully functional holoprotein synthesis in the BLi5 E. coli strain. Protein Expr. Purif. 2004, 38, 184–195. [Google Scholar] [CrossRef]
- Westley, A.M.; Westley, J. Biological sulfane sulfur. Anal. Biochem. 1991, 195, 63–67. [Google Scholar] [CrossRef]
- Savage, J.C.; Gould, D.H. Determination of sulfide in brain tissue and rumen fluid by ion-interaction reversed-phase high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1990, 526, 540–545. [Google Scholar] [CrossRef]
- Ogasawara, Y.; Ishii, K.; Togawa, T.; Tanabe, S. Determination of trace amounts of sulphide in human red blood cells by high-performance liquid chromatography with fluorimetric detection after derivatization with p-phenylenediamine and iron(III). Analyst 1991, 116, 1359–1363. [Google Scholar] [CrossRef]
- Sutton, T.R.; Minnion, M.; Barbarino, F.; Koster, G.; Fernandez, B.O.; Cumpstey, A.F.; Wischmann, P.; Madhani, M.; Frenneaux, M.P.; Postle, A.D.; et al. A robust and versatile mass spectrometry platform for comprehensive assessment of the thiol redox metabolome. Redox Biol. 2018, 16, 359–380. [Google Scholar] [CrossRef]
- Liu, C.; Pan, J.; Li, S.; Zhao, Y.; Wu, L.Y.; Berkman, C.E.; Whorton, A.R.; Xian, M. Capture and visualization of hydrogen sulfide by a fluorescent probe. Angew. Chem. Int. Ed. 2011, 50, 10327–10329. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Cheng, Y.; Dai, C.; King, A.L.; Predmore, B.L.; Lefer, D.J.; Wang, B. A fluorescent probe for fast and quantitative detection of hydrogen sulfide in blood. Angew. Chem. Int. Ed. 2011, 50, 9672–9675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorson, M.K.; Majtan, T.; Kraus, J.P.; Barrios, A.M. Identification of Cystathionine b -Synthase Inhibitors Using a Hydrogen Sulfide Selective Probe. Angew. Chem. 2013, 52, 4641–4644. [Google Scholar] [CrossRef] [PubMed]
- Kerkovius, J.K.; Menard, F. A Practical Synthesis of 6,8-Difluoro-7-hydroxycoumarin Derivatives for Fluorescence Applications. Synth. 2016, 48, 1622–1629. [Google Scholar] [CrossRef]
- Tóth, I.; Solymosi, P.; Szabó, Z. Application of a sulphide-selective electrode in the absence of a pH-buffer. Talanta 1988, 35, 783–788. [Google Scholar] [CrossRef]
- Whitfield, N.L.; Kreimier, E.L.; Verdial, F.C.; Skovgaard, N.; Olson, K.R. Reappraisal of H2S/sulfide concentration in vertebrate blood and its potential significance in ischemic preconditioning and vascular signaling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, 1930–1937. [Google Scholar] [CrossRef] [Green Version]
- Searcy, D.G.; Peterson, M.A. Hydrogen sulfide consumption measured at low steady state concentrations using a sulfidostat. Anal. Biochem. 2004, 324, 269–275. [Google Scholar] [CrossRef]
- Koenitzer, J.R.; Isbell, T.S.; Patel, H.D.; Benavides, G.A.; Dickinson, D.A.; Patel, R.P.; Darley-Usmar, V.M.; Lancaster, J.R.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates vasoactivity in an O2-dependent manner. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, 1953–1960. [Google Scholar] [CrossRef] [Green Version]
- Faccenda, A.; Wang, J.; Mutus, B. Polydimethylsiloxane permeability-based method for the continuous and specific detection of hydrogen sulfide. Anal. Chem. 2012, 84, 5243–5249. [Google Scholar] [CrossRef] [PubMed]
- Krijt, J.; Kopecká, J.; Hnízda, A.; Moat, S.; Kluijtmans, L.A.J.; Mayne, P.; Kožich, V. Determination of cystathionine beta-synthase activity in human plasma by LC-MS/MS: Potential use in diagnosis of CBS deficiency. J. Inherit. Metab. Dis. 2011, 34, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Alcaide, P.; Krijt, J.; Ruiz-Sala, P.; Ješina, P.; Ugarte, M.; Kožich, V.; Merinero, B. Enzymatic diagnosis of homocystinuria by determination of cystathionine-ß-synthase activity in plasma using LC-MS/MS. Clin. Chim. Acta 2015, 438, 261–265. [Google Scholar] [CrossRef]
- Mubeen, S.; Zhang, T.; Chartuprayoon, N.; Rheem, Y.; Mulchandani, A.; Myung, N.V.; Deshusses, M.A. Sensitive detection of H2S using gold nanoparticle decorated single-walled carbon nanotubes. Anal. Chem. 2010, 82, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Morava, E.; Rahman, S.; Peters, V.; Baumgartner, M.R.; Patterson, M.; Zschocke, J. Quo vadis: The re-definition of “inborn metabolic diseases”. J. Inherit. Metab. Dis. 2015, 38, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Zschocke, J. SSIEM Classification of Inborn Errors of Metabolism. In Physician’s Guide to the Diagnosis, Treatment, and Follow-Up of Inherited Metabolic Diseases; Springer: Berlin/Heidelberg, Germany, 2014; pp. 817–830. [Google Scholar]
- CBS[gene]—ClinVar—NCBI. Available online: https://www.ncbi.nlm.nih.gov/clinvar/?term=CBS%5Bgene%5D (accessed on 12 May 2020).
- Kraus, J.P.; Janosǐk, M.; Kožich, V.; Mandell, R.; Shih, V.; Sperandeo, M.P.; Sebastio, G.; De Franchis, R.; Andria, G.; Kluijtmans, L.A.J.; et al. Cystathionine β-synthase mutations in homocystinuria. Hum. Mutat. 1999, 13, 362–375. [Google Scholar] [CrossRef]
- Vicente, J.B.; Colaço, H.G.; Malagrinò, F.; Santo, P.E.; Gutierres, A.; Bandeiras, T.M.; Leandro, P.; Brito, J.A.; Giuffrè, A. A Clinically Relevant Variant of the Human Hydrogen Sulfide-Synthesizing Enzyme Cystathionine β -Synthase: Increased CO Reactivity as a Novel Molecular Mechanism of Pathogenicity? Oxidative Med. Cell. Longev. 2017, 20178. [Google Scholar] [CrossRef] [Green Version]
- DeRatt, B.N.; Ralat, M.A.; Kabil, O.; Chi, Y.-Y.; Banerjee, R.; Gregory, J.F. Vitamin B-6 Restriction Reduces the Production of Hydrogen Sulfide and its Biomarkers by the Transsulfuration Pathway in Cultured Human Hepatoma Cells. J. Nutr. 2014, 144, 1501–1508. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Liu, D.; Qiu, J.; Yuan, H.; Hu, Q.; Xue, H.; Li, T.; Ma, W.W.; Zhang, Q.; Li, G.; et al. Evaluation of H2S-producing enzymes in cerebrospinal fluid and its relationship with interleukin-6 and neurologic deficits in subarachnoid hemorrhage. Biomed. Pharmacother. 2020, 123, 109722. [Google Scholar] [CrossRef]
- Banerjee, R.; Chiku, T.; Kabil, O.; Libiad, M.; Motl, N.; Yadav, P.K. Assay Methods for H2S Biogenesis and Catabolism Enzymes. Methods Enzym. 2015, 554, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Tiranti, V.; Viscomi, C.; Hildebrandt, T.; Di Meo, I.; Mineri, R.; Tiveron, C.; Levitt, M.D.; Prelle, A.; Fagiolari, G.; Rimoldi, M.; et al. Loss of ETHE1, a mitochondrial dioxygenase, causes fatal sulfide toxicity in ethylmalonic encephalopathy. Nat. Med. 2009, 15, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice deficient in cystathionine β-synthase: Animal models for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. USA 1995, 92, 1585–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Wang, L.; Slifker, M.J.; Cai, K.Q.; Maclean, K.N.; Wasek, B.; Bottiglieri, T.; Kruger, W.D. Analysis of differential neonatal lethality in cystathionine β-synthase deficient mouse models using metabolic profiling. FASEB J. 2021, 35. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.K.; Geoghagen, N.S.; Jin, L.; Holt, T.G.; Luo, Q.; Malkowitz, L.; Ni, W.; Quan, S.; Waters, M.G.; Zhang, A.; et al. Pharmacological activation and genetic manipulation of cystathionine beta-synthase alter circulating levels of homocysteine and hydrogen sulfide in mice. Eur. J. Pharmacol. 2011, 650, 86–93. [Google Scholar] [CrossRef]
- George, A.K.; Homme, R.P.; Majumder, A.; Laha, A.; Metreveli, N.; Sandhu, H.S.; Tyagi, S.C.; Singh, M. Hydrogen sulfide intervention in cystathionine-β-synthase mutant mouse helps restore ocular homeostasis. Int. J. Ophthalmol. 2019, 12, 754–764. [Google Scholar] [CrossRef]
- Majtan, T.; Krijt, J.; Sokolová, J.; Křížková, M.; Ralat, M.A.; Kent, J.; Gregory, J.F.; Kožich, V.; Kraus, J.P. Biogenesis of Hydrogen Sulfide and Thioethers by Cystathionine Beta-Synthase. Antioxid. Redox Signal. 2018, 28, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.; Juriasingani, S.; Akbari, M.; Shao, P.; Saha, M.N.; Lobb, I.; Bachtler, M.; Fernandez, B.; Qian, Z.; Van Goor, H.; et al. Endogenous H2S production deficiencies lead to impaired renal erythropoietin production. Can. Urol. Assoc. J. 2019, 13, E210–E219. [Google Scholar] [CrossRef] [PubMed]
- Roman, H.B.; Hirschberger, L.L.; Krijt, J.; Valli, A.; Kožich, V.; Stipanuk, M.H. The cysteine dioxgenase knockout mouse: Altered cysteine metabolism in nonhepatic tissues leads to excess H2S/HS- production and evidence of pancreatic and lung toxicity. Antioxid. Redox Signal. 2013, 19, 1321–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linden, D.R.; Sha, L.; Mazzone, A.; Stoltz, G.J.; Bernard, C.E.; Furne, J.K.; Levitt, M.D.; Farrugia, G.; Szurszewski, J.H. Production of the gaseous signal molecule hydrogen sulfide in mouse tissues. J. Neurochem. 2008, 106, 1577–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, M.I.S.; Colaço, H.G.; Smith, D.E.C.; Ramos, R.J.J.F.; Pop, A.; Van Dooren, S.J.M.; Tavares De Almeida, I.; Kluijtmans, L.A.J.; Janssen, M.C.H.; Rivera, I.; et al. Reduced response of Cystathionine Beta-Synthase (CBS) to S-Adenosylmethionine (SAM): Identification and functional analysis of CBS gene mutations in Homocystinuria patients. J. Inherit. Metab. Dis. 2014, 37, 245–254. [Google Scholar] [CrossRef] [PubMed]









| Specimen Type | [H2S] | Methods of Detection | Reference |
|---|---|---|---|
| (1) Animal | |||
| Mouse brain | 14 nM | Gas chromatography/chemiluminescence sulfur detector | [32] |
| Mouse liver | 17 nM | Gas chromatography/chemiluminescence sulfur detector | [32] |
| Mouse plasma | 0.7 μM | Monobromobimane assay | [33] |
| Mouse whole blood | 15 nM | GC method and chemiluminescence detection | [34] |
| Rat whole blood | 0.7 μM | Monobromobimane assay | [35] |
| Rat aorta | 10 nM | Polarographic electrode PHSS | [36] |
| Rat liver and heart | >100 nM/g | Photometric detector and ion chromatography | [37] |
| Rat brain | 9 µM | Gas chromatography/silver particles | [38] |
| Rat homogenate tissue | 6 pM | Gas chromatography/silver particles | [38] |
| (2) Human | |||
| Human breath | 0.2 ng/L (0.1 ppb) | Chromatography | [39] |
| Human saliva | 1.641–7.124 µM | Fluorescent probe | [40] |
| Human brain tissue | 100 µM | Gas dialysis/ion chromatography | [41] |
| Human cerebrospinal fluid | 3.48 ± 1.68 mg/L (102 ± 49 µM) | Selective electrochemical detection with ion chromatography | [22] |
| Human cerebrospinal fluid (pediatric patients) | 8.71 ± 1.25 µM | Electrode (PXS-270, Shanghai Leici, China) | [21] |
| Human whole blood | 100 pM | Gas chromatography/chemiluminescence sulfur detector | [32] |
| Human plasma (pediatric patients) | 45.55 ± 6.55 µM | Electrode (PXS-270, Shanghai Leici, China) | [21] |
| Human plasma | 300–500 nM | Monobromobimane (MMB) assay | [25] |
| 400–1000 nM | Monobromobimane assay | [42] | |
| 70–125 μM | Microfluidic device with 2,6-dansyl azide (DNS-Az) probe | [20] | |
| c.a 5–100 µM | Methylene blue/UV-visible spectrophotometry | [17] | |
| 54.0 ± 26.4 μM | Methylene blue/UV-visible spectrophotometry | [18] | |
| 14.11 ± 5.24 μM | Methylene blue/UV-visible spectrophotometry | [19] |
| Cell Type | H2S Dosage | Mechanism | Response | Reference |
|---|---|---|---|---|
| Neurons, glia | Supraphysiological | Stimulation of NMDA receptors, induced by an increase of cAMP concentration | Causes/contributes to tissue damage, neuroinflammatory | [43] |
| Physiological | Messenger in response to specific stimuli (usually noxious) | [43] | ||
| Physiological | Increasing the concentration of glutathione, activating K+ATP and Cl− channels | Protection from oxidative stress | [46,48] | |
| Vascular endothelial cells | Release of H2S | Membrane hyperpolarization (opening of the VSMC K+ATP channels), reduction of extracellular Ca2+ entry | Vasorelaxant effect | [49] |
| Exogenous H2S | Suppression of VSMC proliferation through protein kinase (MAPK) signaling pathway | Attenuation of cell proliferation rate, increase of cell apoptosis, regulation of cell growth, structural remodeling of vessel tissues | [49,50] | |
| Reduced (e.g., due to CSE deficiency) | Loss of regulatory effects | Hypertension, atherosclerosis, myocardial injury | [51] | |
| Pulmonary artery cells | Endogenous release | Inhibition of proliferation and interleukin- (IL)-8 secretion by impairing extracellular signal-regulated kinase 1/2 (ERK1/2) and p38-dependent signaling pathways | Lung remodeling, inhibiting collagen accumulation in the wall of the pulmonary artery in hypoxia | [52] |
| Inhibition of endogenous H2S release | Signal-regulated kinase 1/2 (ERK1/2) and p38-dependent signaling pathways active | Increased human airway, smooth muscle proliferation, IL-8 secretion | [53] | |
| Pancreatic islets cells | Physiological | Stimulation of K+ATP channels in β-cells, possible inhibition of insulin release by activation of K+ATP channels | Inhibition of insulin release, induction of apoptosis | [54,55] |
| Colonocytes | Release of H2S | Activation of K+ATP channels in the vascular tissue, stimulation of vanilloid type I receptor | Inhibition of jejunum and colon motility, regulation of the gastric mucosal blood flow, secretion and increased spike activity in the afferent neurons of the enteric nervous system | [56,57] |
| Supraphysiological | Inhibits cytochrome c oxidase and synthesis of ATP | Pro-inflammatory, suppresses mitochondrial energy production, destabilizes the protective intestinal mucus layer, potentially participates in the etiology of bowel disorders, occurs as genotoxic agent in colorectal cancer | [14,44,58] | |
| Low concentration | Energy substrate | Anti-inflammatory | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Myszkowska, J.; Derevenkov, I.; Makarov, S.V.; Spiekerkoetter, U.; Hannibal, L. Biosynthesis, Quantification and Genetic Diseases of the Smallest Signaling Thiol Metabolite: Hydrogen Sulfide. Antioxidants 2021, 10, 1065. https://doi.org/10.3390/antiox10071065
Myszkowska J, Derevenkov I, Makarov SV, Spiekerkoetter U, Hannibal L. Biosynthesis, Quantification and Genetic Diseases of the Smallest Signaling Thiol Metabolite: Hydrogen Sulfide. Antioxidants. 2021; 10(7):1065. https://doi.org/10.3390/antiox10071065
Chicago/Turabian StyleMyszkowska, Joanna, Ilia Derevenkov, Sergei V. Makarov, Ute Spiekerkoetter, and Luciana Hannibal. 2021. "Biosynthesis, Quantification and Genetic Diseases of the Smallest Signaling Thiol Metabolite: Hydrogen Sulfide" Antioxidants 10, no. 7: 1065. https://doi.org/10.3390/antiox10071065