
An Insight into Giant Cell Arteritis Pathogenesis: Evidence for Oxidative Stress and SIRT1 Downregulation

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Sample Collection
2.3. Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
2.4. Blood Leukocytes Intracellular ROS Levels
2.5. Plasma Lipid Peroxidation Estimation (Thiobarbituric Acid Reactive Substances Assay; TBARS Assay)
2.6. Plasma Total Antioxidant Capacity Assay (Oxygen Radical Absorbance Capacity Assay; ORAC Assay)
2.7. Oxidative Stress Index (OSI)
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Subjects
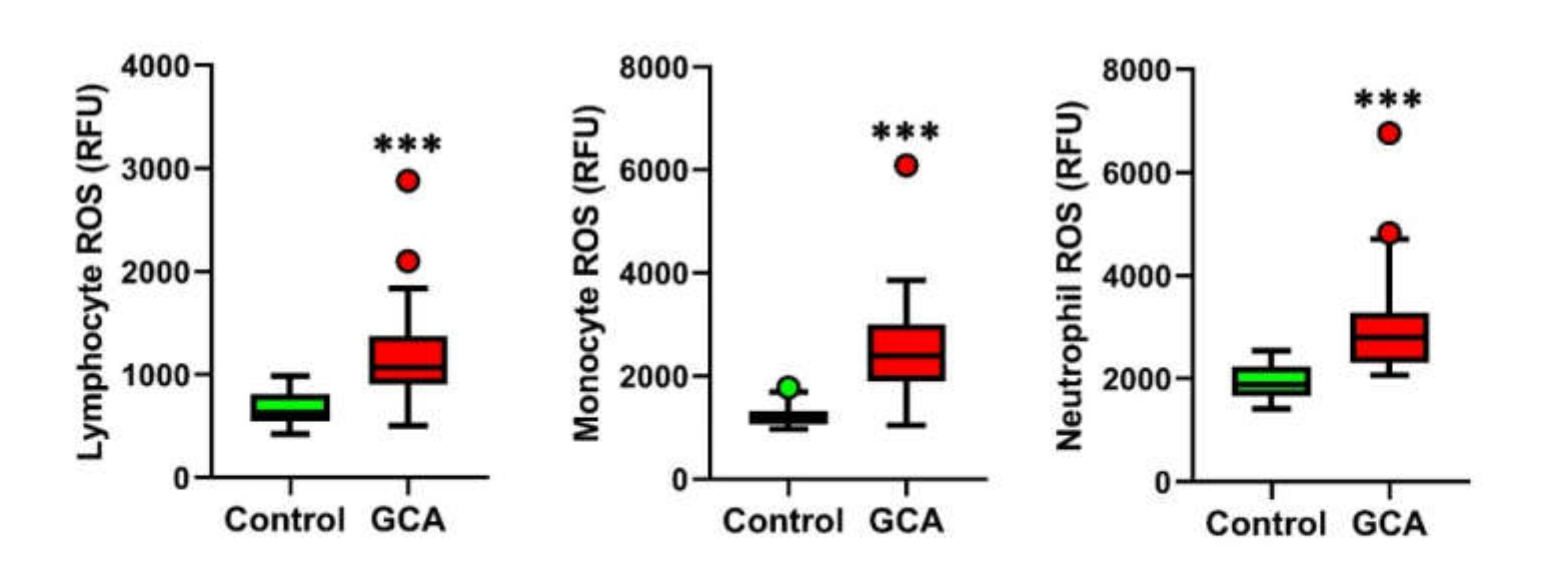
3.2. Blood Leukocytes from GCA Display Increased Intracellular ROS Levels
3.3. Signs of Plasma Oxidative Stress in GCA
3.4. PBMCs Derived from GCA Patients Display a Significant Decrease in SIRT1 Expression
3.5. Blood Oxidative Stress Parameters and Sirtuin Expression in GCA Patients with and without Diabetes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weyand, C.M.; Liao, Y.J.; Goronzy, J.J. The immunopathology of giant cell arteritis: Diagnostic and therapeutic implications. J Neuroophthalmol. 2012, 32, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Azhar, S.S.; Tang, R.A.; Dorotheo, E.U. Giant cell arteritis: Diagnosing and treating inflammatory disease in older adults. Geriatrics 2005, 60, 26–30. [Google Scholar] [PubMed]
- Aviña-Zubieta, J.A.; Bhole, V.M.; Amiri, N.; Sayre, E.C.; Choi, H.K. The risk of deep venous thrombosis and pulmonary embolism in giant cell arteritis: A general population-based study. Ann. Rheum. Dis. 2016, 75, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ai, Z.; Khoyratty, T.; Zec, K.; Eames, H.L.; Van Grinsven, E.; Hudak, A.; Morris, S.; Ahern, D.; Monaco, C.; et al. ROS-producing immature neutrophils in giant cell arteritis are linked to vascular pathologies. JCI Insight 2020, 5, e139163. [Google Scholar] [CrossRef] [PubMed]
- Hunder, G.G.; Bloch, D.A.; Michel, B.A.; Stevens, M.B.; Arend, W.P.; Calabrese, L.H.; Edworthy, S.M.; Fauci, A.S.; Leavitt, R.Y.; Lie, J.T.; et al. The American College of Rheumatology 1990 criteria for the classification of giant cell arteritis. Arthritis Rheum. 1990, 33, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Rittner, H.L.; Hafner, V.; Klimiuk, P.A.; Szweda, L.I.; Goronzy, J.J.; Weyand, C.M. Aldose reductase functions as a detoxification system for lipid peroxidation products in vasculitis. J. Clin. Investig. 1999, 103, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becatti, M.; Emmi, G.; Silvestri, E.; Bruschi, G.; Ciucciarelli, L.; Squatrito, D.; Vaglio, A.; Taddei, N.; Abbate, R.; Emmi, L.; et al. Neutrophil Activation Promotes Fibrinogen Oxidation and Thrombus Formation in Behçet Disease. Circulation 2016, 133, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Yuan, X.; Bober, E.; Braun, T. Sirtuins in the Cardiovascular System: Potential Targets in Pediatric Cardiology. Pediatr. Cardiol. 2018, 39, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Wu, D.; Li, Y.; Zhu, K.S.; Wang, H.; Zhu, W.G. Advances in Cellular Characterization of the Sirtuin Isoform, SIRT7. Front. Endocrinol. 2018, 9, 652. [Google Scholar] [CrossRef] [Green Version]
- Santos-Barriopedro, I.; Vaquero, A. Complex role of SIRT6 in NF-κB pathway regulation. Mol. Cell Oncol. 2018, 5, e1445942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Zhang, X.; Zhang, L. Negative regulation of inflammation by SIRT1. Pharmacol. Res. 2013, 67, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Schenk, S.; Imamura, T.; Babendure, J.L.; Sonoda, N.; Bae, E.J.; Oh, D.Y.; Lu, M.; Milne, J.C.; Westphal, C.; et al. SIRT1 inhibits inflammatory pathways in macrophages and modulates insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E419–E428. [Google Scholar] [CrossRef] [Green Version]
- Schug, T.T.; Xu, Q.; Gao, H.; Peres-Da-Silva, A.; Draper, D.W.; Fessler, M.B.; Purushotham, A.; Li, X. Myeloid deletion of SIRT1 induces inflammatory signaling in response to environmental stress. Mol. Cell. Biol. 2010, 30, 4712–4721. [Google Scholar] [CrossRef] [Green Version]
- Colley, T.; Mercado, N.; Kunori, Y.; Brightling, C.; Bhavsar, P.K.; Barnes, P.J.; Ito, K. Defective sirtuin-1 increases IL-4 expression through acetylation of GATA-3 in patients with severe asthma. J. Allergy Clin. Immunol. 2016, 137, 1595–1597. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, A.; Sofi, F.; Luisi, M.L.E.; Rafanelli, E.; Fiorillo, C.; Becatti, M.; Abbate, R.; Casini, A.; Gensini, G.F.; Benedettelli, S. An organic khorasan wheat-based replacement diet improves risk profile of patients with acute coronary syndrome: A randomized crossover trial. Nutrients 2015, 7, 3401–3415. [Google Scholar] [CrossRef] [Green Version]
- Emmi, G.; Bettiol, A.; Niccolai, E.; Ramazzotti, M.; Amedei, A.; Pagliai, G.; Taddei, N.; Sofi, F.; Fiorillo, C.; Prisco, D.; et al. Butyrate-Rich Diets Improve Redox Status and Fibrin Lysis in Behçet’s Syndrome. Circ. Res. 2021, 128, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Becatti, M.; Fucci, R.; Mannucci, A.; Barygina, V.; Mugnaini, M.; Criscuoli, L.; Giachini, C.; Bertocci, F.; Picone, R.; Emmi, G.; et al. A Biochemical Approach to Detect Oxidative Stress in Infertile Women Undergoing Assisted Reproductive Technology Procedures. Int. J. Mol. Sci. 2018, 19, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, A.; Dinu, M.; Cesari, F.; Gori, A.M.; Fiorillo, C.; Becatti, M.; Casini, A.; Marcucci, R.; Benedettelli, S.; Sofi, F. A khorasan wheat-based replacement diet improves risk profile of patients with type 2 diabetes mellitus (T2DM): A randomized crossover trial. Eur. J. Nutr. 2017, 56, 1191–1200. [Google Scholar] [CrossRef] [Green Version]
- Cito, G.; Becatti, M.; Natali, A.; Fucci, R.; Picone, R.; Cocci, A.; Falcone, P.; Criscuoli, L.; Mannucci, A.; Argento, F.R.; et al. Redox status assessment in infertile patients with non-obstructive azoospermia undergoing testicular sperm extraction: A prospective study. Andrology 2020, 8, 364–371. [Google Scholar]
- Ianni, A.; Kumari, P.; Tarighi, S.; Simonet, N.G.; Popescu, D.; Guenther, S.; Hölper, S.; Schmidt, A.; Smolka, C.; Yue, S.; et al. SIRT7-dependent deacetylation of NPM promotes p53 stabilization following UV-induced genotoxic stress. Proc. Natl. Acad. Sci. USA 2021, 118, e2015339118. [Google Scholar] [CrossRef]
- Kitada, M.; Ogura, Y.; Monno, I.; Koya, D. Sirtuins and Type 2 Diabetes: Role in Inflammation, Oxidative Stress, and Mitochondrial Function. Front. Endocrinol. 2019, 10, 187. [Google Scholar] [CrossRef] [Green Version]
- De Smit, E.; Palmer, A.J.; Hewitt, A.W. Projected worldwide disease burden from giant cell arteritis by 2050. J. Rheumatol. 2015, 42, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.V.; Liao, Y.J.; Kim, J.W.; Goronzy, J.J.; Weyand, C.M. Giant cell arteritis: Immune and vascular aging as disease risk factors. Arthritis Res. Ther. 2011, 13, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginzales-Gay, M.A. Genetic epidemiology. Giant cell arteritis and polymyalgia rheumatica. Arthritis Res. 2001, 3, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Pang, J.; Han, Y.; Dai, Y.; Dai, D.; Cai, J.; Zhang, T.-M. Age-dependent tissue expression patterns of Sirt1 in senescence-accelerated mice. Mol. Med. Rep. 2014, 10, 3296–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.R.; Wright, J.; Bauter, M.; Seweryniak, K.; Kode, A.; Rahman, I. Sirtuin regulates cigarette smoke-induced proinflammatory mediator release via RelA/p65 NF-kappaB in macrophages in vitro and in rat lungs in vivo: Implications for chronic inflammation and aging. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L567–L576. [Google Scholar]
- Conti, V.; Corbi, G.; Manzo, V.; Malangone, P.; Vitale, C.; Maglio, A.; Cotugno, R.; Capaccio, D.; Marino, L.; Selleri, C.; et al. SIRT1 Activity in Peripheral Blood Mononuclear Cells Correlates with Altered Lung Function in Patients with Chronic Obstructive Pulmonary Disease. Oxid. Med. Cell. Longev. 2018, 2018, 9391261. [Google Scholar] [CrossRef]
- Kong, S.; McBurney, M.W.; Fang, D. Sirtuin 1 in immune regulation and autoimmunity. Immunol Cell Biol. 2012, 90, 6–13. [Google Scholar]
- De Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.A.; Rutter, G.A. The role of oxidative stress and hypoxia in pancreatic beta-cell dysfunction in diabetes mellitus. Antioxid. Redox Signal. 2017, 26, 501–518. [Google Scholar] [CrossRef] [Green Version]
- Kitada, M.; Kume, S.; Kanasaki, K.; Takeda-Watanabe, A.; Koya, D. Sirtuins as possible drug targets in type 2 diabetes. Curr. Drug Targets 2013, 14, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Koya, D. SIRT1 in type 2 diabetes: Mechanisms and therapeutic potential. Diabetes Metab. J. 2013, 37, 315–325. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, G.; Tao, R.; Wu, P.; Kono, T.; Li, K.; Ding, W.-X.; Tong, X.; Tersey, S.A.; Harris, R.A.; et al. Sirtuin 6 regulates glucose-stimulated insulin secretion in mouse pancreatic beta cells. Diabetologia 2016, 59, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, J.H.; Tuckwell, K.; Dimonaco, S.; Klearman, M.; Aringer, M.; Blockmans, D.E.; Brouwer, E.; Cid, M.C.; Dasgupta, B.; Rech, J.; et al. Trial of Tocilizumab in Giant-Cell Arteritis. N. Engl. J. Med. 2017, 377, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Villiger, P.M.; Adler, S.; Kuchen, S.; Wermelinger, F.; Dan, D.; Fiege, V.; Bütikofer, L.; Seitz, M.; Reichenbach, S. Tocilizumab for induction and maintenance of remission in giant cell arteritis: A phase 2, randomised, double-blind, placebo-controlled trial. Lancet 2016, 387, 1921–1927. [Google Scholar] [CrossRef]
- Unizony, S.; McCulley, T.J.; Spiera, R.; Pei, J.; Sidiropoulos, P.N.; Best, J.H.; Birchwood, C.; Pavlov, A.; Stone, J.H. Clinical outcomes of patients with giant cell arteritis treated with tocilizumab in real-world clinical practice: Decreased incidence of new visual manifestations. Arthritis Res. Ther. 2021, 23, 8. [Google Scholar] [CrossRef] [PubMed]
- Milne, J.C.; Lambert, P.D.; Schenk, S.; Carney, D.P.; Smith, J.J.; Gagne, D.J.; Jin, L.; Boss, O.; Perni, R.B.; Vu, C.B. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature 2007, 450, 712–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, H.; Sinclair, D.A.; Ellis, J.L.; Steegborn, C. Sirtuin activators and inhibitors: Promises, achievements, and challenges. Pharmacol. Ther. 2018, 188, 140–154. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients Features | GCA Cases (% out of 30) | Healthy Controls (% out of 30) |
|---|---|---|
| Age at diagnosis, years (median, IQR) | 74 (69–80) | |
| Age at inclusion, years (median, IQR) | 76 (71–81) | 75 (71–82) |
| Disease duration, years (median, IQR) | 1 (1–2) | - |
| Gender | ||
| Female | 21 (70%) | 21 (70%) |
| Male | 9 (30%) | 9 (30%) |
| Disease Manifestations at Diagnosis | ||
| Headache | 21 (70%) | - |
| Arthralgia/myalgia | 21 (70%) | |
| Jaw claudication | 12 (40%) | |
| Weight loss | 11 (36.7%) | |
| Cutaneous symptoms | 9 (30%) | |
| Fever | 8 (26.7%) | |
| Visual loss | 6 (20%) | |
| Cardiovascular events | 3 (10%) | |
| Polymyalgia rheumatica | 11 (36.7%) | |
| Cardiovascular Risk Factors | ||
| Hypertension | 17 (56.7%) | 0 (0%) |
| Dyslipidemia | 12 (40%) | 0 (0%) |
| Diabetes | 7 (23.3%) | 0 (0%) |
| Cardiopathy | 3 (10%) | 0 (0%) |
| Obesity | 1 (3.3%) | 0 (0%) |
| Ongoing Therapy | ||
| Corticosteroids | 26 (86.7%) | |
| Median prednisone daily dose, mg | 10 (5–17.5) | |
| Antiplatelet therapy | 12 (40%) |
| GCA Patients with Diabetes (n = 7) | GCA Patients without Diabetes (n = 23) | p-Value * | |
|---|---|---|---|
| Oxidative Stress Parameters | |||
| Lymphocytes ROS (RFU) | 1079 (984–1635) | 1065 (877–1330) | 0.447 |
| Monocytes ROS (RFU) | 2703 (2169–3321) | 2369 (1881–2923) | 0.447 |
| Neutrophil ROS (RFU) | 3029 (2362–3421) | 2753 (2237–3217) | 0.270 |
| Plasma lipid peroxidation (MDA nmol/mL) | 2.0 (1.5–2.3) | 1.6 (1.4–2.1) | 0.390 |
| Plasma antioxidant capacity (nmol/mL Trolox Equivalent) | 18.5 (15.5–23.8) | 16.7 (15.5–19.6) | 0.508 |
| OSI (MDA/ORAC * 100) | 11.3 (9.5–12.0) | 9.1 (7.9–12.6) | 0.391 |
| SIRT Relative Expression | |||
| SIRT1 | 52 (42–58) | 42 (39–50) | 0.134 |
| SIRT6 | 134 (82–158) | 85 (58–113) | 0.292 |
| SIRT7 | 111 (50–164) | 76 (36–136) | 0.249 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ianni, A.; Kumari, P.; Tarighi, S.; Argento, F.R.; Fini, E.; Emmi, G.; Bettiol, A.; Braun, T.; Prisco, D.; Fiorillo, C.; et al. An Insight into Giant Cell Arteritis Pathogenesis: Evidence for Oxidative Stress and SIRT1 Downregulation. Antioxidants 2021, 10, 885. https://doi.org/10.3390/antiox10060885
Ianni A, Kumari P, Tarighi S, Argento FR, Fini E, Emmi G, Bettiol A, Braun T, Prisco D, Fiorillo C, et al. An Insight into Giant Cell Arteritis Pathogenesis: Evidence for Oxidative Stress and SIRT1 Downregulation. Antioxidants. 2021; 10(6):885. https://doi.org/10.3390/antiox10060885
Chicago/Turabian StyleIanni, Alessandro, Poonam Kumari, Shahriar Tarighi, Flavia Rita Argento, Eleonora Fini, Giacomo Emmi, Alessandra Bettiol, Thomas Braun, Domenico Prisco, Claudia Fiorillo, and et al. 2021. "An Insight into Giant Cell Arteritis Pathogenesis: Evidence for Oxidative Stress and SIRT1 Downregulation" Antioxidants 10, no. 6: 885. https://doi.org/10.3390/antiox10060885