
Tocotrienols: Dietary Supplements for Chronic Obstructive Pulmonary Disease


Abstract
:1. Introduction
2. Pathogenesis, Diagnosis, and Risk Factors of COPD
3. Oxidative Stress in the Lung
4. Biological Mechanisms of COPD: Free Radicals, Inflammation, and the NFκB Pathway
5. Role of Vitamin E in COPD: Introduction to Tocopherols and Tocotrienols
6. Antioxidant and Anti-Inflammatory Effects of Tocotrienol: Cell Culture Models
7. Antioxidant and Anti-Inflammatory Effects of Tocotrienol: Animal Models
8. Antioxidant and Anti-Inflammatory Effects of Tocotrienol in COPD: Human Studies
9. Availability, Metabolism, and Safety of Tocotrienols
10. Limitation
11. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| G-CSF | granulocyte-colony-stimulating factor; |
| hs-CRP | high-sensitivity C-reaction protein; |
| IL-1β | interleukin-1β; |
| IL-6 | interleukin-6; |
| iNOS | inducible nitric oxide synthase; |
| LPS | lipopolysaccharide; |
| MDA | malondialdehyde; |
| MCP-1 | monocyte chemoattractant protein-1; |
| NO | nitric oxide; |
| NF-kB | nuclear factor kappa B; |
| Nrf2 | nuclear factor erythroid 2; |
| ROS | reactive oxygen species; |
| STAT3 | signal transducer and activator of transcription 3; |
| TNF-α | tumor necrosis factor-α; |
| VCAM-1 | vascular adhesion protein. |
References
- Quaderi, S.A.; Hurst, J.R. The unmet global burden of COPD. Glob. Health Epidemiol. Genom. 2018, 3, e4. [Google Scholar] [CrossRef] [Green Version]
- Burkes, R.M.; Donohue, J.F. An Update on the Global Initiative for Chronic Obstructive Lung Disease 2017 Guidelines with a Focus on Classification and Management of Stable COPD. Respir. Care 2018, 63, 749–758. [Google Scholar] [CrossRef]
- Matera, M.G.; Page, C.P.; Cazzola, M. Novel bronchodilators for the treatment of chronic obstructive pulmonary disease. Trends Pharm. Sci. 2011, 32, 495–506. [Google Scholar] [CrossRef]
- Hizawa, N. LAMA/LABA vs ICS/LABA in the treatment of COPD in Japan based on the disease phenotypes. Int. J. Chron. Obstr. Pulmon. Dis. 2015, 10, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Wedzicha, J.A.; Banerji, D.; Chapman, K.R.; Vestbo, J.; Roche, N.; Ayers, R.T.; Thach, C.; Fogel, R.; Patalano, F.; Vogelmeier, C.F.; et al. Indacaterol-Glycopyrronium versus Salmeterol-Fluticasone for COPD. N. Engl. J. Med. 2016, 374, 2222–2234. [Google Scholar] [CrossRef] [Green Version]
- Keatings, V.M.; Jatakanon, A.; Worsdell, Y.M.; Barnes, P.J. Effects of inhaled and oral glucocorticoids on inflammatory indices in asthma and COPD. Am. J. Respir. Crit. Care Med. 1997, 155, 542–548. [Google Scholar] [CrossRef]
- Loppow, D.; Schleiss, M.B.; Kanniess, F.; Taube, C.; Jorres, R.A.; Magnussen, H. In patients with chronic bronchitis a four week trial with inhaled steroids does not attenuate airway inflammation. Respir. Med. 2001, 95, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Price, D.; Yawn, B.; Brusselle, G.; Rossi, A. Risk-to-benefit ratio of inhaled corticosteroids in patients with COPD. Prim. Care Respir. J. 2013, 22, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, F.L.; Krahnke, J.S.; Kim, V. Clinical issues of mucus accumulation in COPD. Int. J. Chron. Obstr. Pulmon. Dis. 2014, 9, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Saetta, M.; Turato, G.; Baraldo, S.; Zanin, A.; Braccioni, F.; Mapp, C.E.; Maestrelli, P.; Cavallesco, G.; Papi, A.; Fabbri, L.M. Goblet cell hyperplasia and epithelial inflammation in peripheral airways of smokers with both symptoms of chronic bronchitis and chronic airflow limitation. Am. J. Respir. Crit. Care Med. 2000, 161, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.L. The importance of the assessment of pulmonary function in COPD. Med. Clin. N. Am. 2012, 96, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanney, M.P.; Ruppel, G.; Enright, P.L.; Pedersen, O.F.; Crapo, R.O.; Miller, M.R.; Jensen, R.L.; Falaschetti, E.; Schouten, J.P.; Hankinson, J.L.; et al. Using the lower limit of normal for the FEV1/FVC ratio reduces the misclassification of airway obstruction. Thorax 2008, 63, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Qaseem, A.; Wilt, T.J.; Weinberger, S.E.; Hanania, N.A.; Criner, G.; van der Molen, T.; Marciniuk, D.D.; Denberg, T.; Schunemann, H.; Wedzicha, W.; et al. Diagnosis and management of stable chronic obstructive pulmonary disease: A clinical practice guideline update from the American College of Physicians, American College of Chest Physicians, American Thoracic Society, and European Respiratory Society. Ann. Intern. Med. 2011, 155, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Agusti, A.; Anzueto, A.; Barnes, P.J.; Bourbeau, J.; Celli, B.R.; Criner, G.J.; Frith, P.; Halpin, D.M.G.; Han, M.; et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease: The GOLD science committee report 2019. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [PubMed]
- Foreman, M.G.; Campos, M.; Celedon, J.C. Genes and chronic obstructive pulmonary disease. Med. Clin. N. Am. 2012, 96, 699–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eklund, B.M.; Nilsson, S.; Hedman, L.; Lindberg, I. Why do smokers diagnosed with COPD not quit smoking?—A qualitative study. Tob. Induc. Dis. 2012, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Lundback, B.; Lindberg, A.; Lindstrom, M.; Ronmark, E.; Jonsson, A.C.; Jonsson, E.; Larsson, L.G.; Andersson, S.; Sandstrom, T.; Larsson, K.; et al. Not 15 but 50% of smokers develop COPD?—Report from the Obstructive Lung Disease in Northern Sweden Studies. Respir. Med. 2003, 97, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H.; Sood, A.; Meek, P.M.; Shen, X.; Cheng, Y.; Belinsky, S.A.; Owen, C.A.; Washko, G.; Pinto-Plata, V.; Kelly, E.; et al. Rapid lung function decline in smokers is a risk factor for COPD and is attenuated by angiotensin-converting enzyme inhibitor use. Chest 2014, 145, 695–703. [Google Scholar] [CrossRef] [Green Version]
- Pelkonen, M.K.; Notkola, I.K.; Laatikainen, T.K.; Jousilahti, P. Chronic bronchitis in relation to hospitalization and mortality over three decades. Respir. Med. 2017, 123, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Louhelainen, N.; Rytila, P.; Haahtela, T.; Kinnula, V.L.; Djukanovic, R. Persistence of oxidant and protease burden in the airways after smoking cessation. BMC Pulm. Med. 2009, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Schaberg, T.; Klein, U.; Rau, M.; Eller, J.; Lode, H. Subpopulations of alveolar macrophages in smokers and nonsmokers: Relation to the expression of CD11/CD18 molecules and superoxide anion production. Am. J. Respir. Crit. Care Med. 1995, 151, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.; Caramori, G.; Oates, T.; Capelli, A.; Lusuardi, M.; Gnemmi, I.; Ioli, F.; Chung, K.F.; Donner, C.F.; Barnes, P.J.; et al. Increased expression of nuclear factor-kappaB in bronchial biopsies from smokers and patients with COPD. Eur. Respir. J. 2002, 20, 556–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, S.S. Tobacco carcinogens, their biomarkers and tobacco-induced cancer. Nat. Rev. Cancer 2003, 3, 733–744. [Google Scholar] [CrossRef]
- Bluhm, A.L.; Weinstein, J.; Sousa, J.A. Free radicals in tobacco smoke. Nature 1971, 229, 500. [Google Scholar] [CrossRef] [PubMed]
- Goel, R.; Bitzer, Z.; Reilly, S.M.; Trushin, N.; Foulds, J.; Muscat, J.; Liao, J.; Elias, R.J.; Richie, J.P., Jr. Variation in Free Radical Yields from U.S. Marketed Cigarettes. Chem. Res. Toxicol. 2017, 30, 1038–1045. [Google Scholar] [CrossRef] [Green Version]
- McGowan, S.E.; Hunninghake, G.W. Neutrophils and emphysema. N. Engl. J. Med. 1989, 321, 968–970. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.E.; Thompson, R.J. Extracellular matrix proteoglycan degradation by human alveolar macrophages and neutrophils. J. Appl. Physiol. 1989, 66, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; MacNee, W. Oxidant/antioxidant imbalance in smokers and chronic obstructive pulmonary disease. Thorax 1996, 51, 348–350. [Google Scholar] [CrossRef] [Green Version]
- Heijink, I.H.; Brandenburg, S.M.; Postma, D.S.; van Oosterhout, A.J. Cigarette smoke impairs airway epithelial barrier function and cell-cell contact recovery. Eur. Respir. J. 2012, 39, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Lo Tam Loi, A.T.; Hoonhorst, S.J.; Franciosi, L.; Bischoff, R.; Hoffmann, R.F.; Heijink, I.; van Oosterhout, A.J.; Boezen, H.M.; Timens, W.; Postma, D.S.; et al. Acute and chronic inflammatory responses induced by smoking in individuals susceptible and non-susceptible to development of COPD: From specific disease phenotyping towards novel therapy. Protocol of a cross-sectional study. BMJ Open 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Bernard, A.; Ku, J.M.; Vlahos, R.; Miller, A.A. Cigarette smoke extract exacerbates hyperpermeability of cerebral endothelial cells after oxygen glucose deprivation and reoxygenation. Sci. Rep. 2019, 9, 15573. [Google Scholar] [CrossRef]
- Sargeant, L.A.; Jaeckel, A.; Wareham, N.J. Interaction of vitamin C with the relation between smoking and obstructive airways disease in EPIC Norfolk. European Prospective Investigation into Cancer and Nutrition. Eur. Respir. J. 2000, 16, 397–403. [Google Scholar] [CrossRef] [PubMed]
- West, X.Z.; Malinin, N.L.; Merkulova, A.A.; Tischenko, M.; Kerr, B.A.; Borden, E.C.; Podrez, E.A.; Salomon, R.G.; Byzova, T.V. Oxidative stress induces angiogenesis by activating TLR2 with novel endogenous ligands. Nature 2010, 467, 972–976. [Google Scholar] [CrossRef]
- Pace, E.; Ferraro, M.; Siena, L.; Melis, M.; Montalbano, A.M.; Johnson, M.; Bonsignore, M.R.; Bonsignore, G.; Gjomarkaj, M. Cigarette smoke increases Toll-like receptor 4 and modifies lipopolysaccharide-mediated responses in airway epithelial cells. Immunology 2008, 124, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Adcock, I.M.; Caramori, G.; Barnes, P.J. Chronic obstructive pulmonary disease and lung cancer: New molecular insights. Respiration 2011, 81, 265–284. [Google Scholar] [CrossRef]
- Kumar, R.; Clermont, G.; Vodovotz, Y.; Chow, C.C. The dynamics of acute inflammation. J. Theor. Biol. 2004, 230, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Taneja, V.; Vassallo, R. Cigarette smoking and inflammation: Cellular and molecular mechanisms. J. Dent. Res. 2012, 91, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Eeden, S.F.V. Lung Macrophage Phenotypes and Functional Responses: Role in the Pathogenesis of COPD. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewhurst, J.A.; Lea, S.; Hardaker, E.; Dungwa, J.V.; Ravi, A.K.; Singh, D. Characterisation of lung macrophage subpopulations in COPD patients and controls. Sci. Rep. 2017, 7, 7143. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.M.J.; Ji, X.; Zimmerman, L.J.; Li, M.; Harris, B.K.; Hoeksema, M.D.; Trenary, I.A.; Zou, Y.; Qian, J.; Slebos, R.J.; et al. The airway epithelium undergoes metabolic reprogramming in individuals at high risk for lung cancer. JCI Insight 2016, 1, e88814. [Google Scholar] [CrossRef] [Green Version]
- Semlali, A.; Witoled, C.; Alanazi, M.; Rouabhia, M. Whole cigarette smoke increased the expression of TLRs, HBDs, and proinflammory cytokines by human gingival epithelial cells through different signaling pathways. PLoS ONE 2012, 7, e52614. [Google Scholar] [CrossRef] [PubMed]
- Kolsum, U.; Roy, K.; Starkey, C.; Borrill, Z.; Truman, N.; Vestbo, J.; Singh, D. The repeatability of interleukin-6, tumor necrosis factor-alpha, and C-reactive protein in COPD patients over one year. Int. J. Chron. Obstr. Pulmon. Dis. 2009, 4, 149–156. [Google Scholar] [CrossRef] [Green Version]
- MacNee, W. Systemic inflammatory biomarkers and co-morbidities of chronic obstructive pulmonary disease. Ann. Med. 2013, 45, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Caramori, G.; Adcock, I.M.; Di Stefano, A.; Chung, K.F. Cytokine inhibition in the treatment of COPD. Int. J. Chron. Obstr. Pulmon. Dis. 2014, 9, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Caramori, G.; Casolari, P.; Cavallesco, G.N.; Giuffre, S.; Adcock, I.; Papi, A. Mechanisms involved in lung cancer development in COPD. Int. J. Biochem. Cell Biol. 2011, 43, 1030–1044. [Google Scholar] [CrossRef]
- Peh, H.Y.; Tan, W.S.D.; Chan, T.K.; Pow, C.W.; Foster, P.S.; Wong, W.S.F. Vitamin E isoform gamma-tocotrienol protects against emphysema in cigarette smoke-induced COPD. Free Radic. Biol. Med. 2017, 110, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Tavilani, H.; Nadi, E.; Karimi, J.; Goodarzi, M.T. Oxidative stress in COPD patients, smokers, and non-smokers. Respir. Care 2012, 57, 2090–2094. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.J.; Celli, B.R. Systemic manifestations and comorbidities of COPD. Eur. Respir. J. 2009, 33, 1165–1185. [Google Scholar] [CrossRef] [Green Version]
- Arja, C.; Surapaneni, K.M.; Raya, P.; Adimoolam, C.; Balisetty, B.; Kanala, K.R. Oxidative stress and antioxidant enzyme activity in South Indian male smokers with chronic obstructive pulmonary disease. Respirology 2013, 18, 1069–1075. [Google Scholar] [CrossRef]
- Ahmad, A.; Shameem, M.; Husain, Q. Altered oxidant-antioxidant levels in the disease prognosis of chronic obstructive pulmonary disease. Int. J. Tuberc. Lung Dis. 2013, 17, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Ochs-Balcom, H.M.; Grant, B.J.; Muti, P.; Sempos, C.T.; Freudenheim, J.L.; Browne, R.W.; McCann, S.E.; Trevisan, M.; Cassano, P.A.; Iacoviello, L.; et al. Antioxidants, oxidative stress, and pulmonary function in individuals diagnosed with asthma or COPD. Eur. J. Clin. Nutr. 2006, 60, 991–999. [Google Scholar] [CrossRef]
- Guenegou, A.; Leynaert, B.; Pin, I.; Le Moel, G.; Zureik, M.; Neukirch, F. Serum carotenoids, vitamins A and E, and 8 year lung function decline in a general population. Thorax 2006, 61, 320–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeever, T.M.; Lewis, S.A.; Smit, H.A.; Burney, P.; Cassano, P.A.; Britton, J. A multivariate analysis of serum nutrient levels and lung function. Respir. Res. 2008, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.C.; Wu, T.C.; Chen, P.Y.; Hsieh, L.Y.; Yeh, S.L. Comparison of plasma and intake levels of antioxidant nutrients in patients with chronic obstructive pulmonary disease and healthy people in Taiwan: A case-control study. Asia Pac. J. Clin. Nutr. 2010, 19, 393–401. [Google Scholar] [PubMed]
- Bartoli, M.L.; Novelli, F.; Costa, F.; Malagrino, L.; Melosini, L.; Bacci, E.; Cianchetti, S.; Dente, F.L.; Di Franco, A.; Vagaggini, B.; et al. Malondialdehyde in exhaled breath condensate as a marker of oxidative stress in different pulmonary diseases. Mediat. Inflamm. 2011, 2011, 891752. [Google Scholar] [CrossRef] [PubMed]
- Cheeseman, K.H.; Burton, G.W.; Ingold, K.U.; Slater, T.F. Lipid peroxidation and lipid antioxidants in normal and tumor cells. Toxicol. Pathol. 1984, 12, 235–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peh, H.Y.; Tan, W.S.; Liao, W.; Wong, W.S. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Brigelius-Flohe, R.; Traber, M.G. Vitamin E: Function and metabolism. FASEB J. 1999, 13, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, H.; Ahad, A.; Iqbal, J.; Siddiqui, W.A. Pharmacological potential of tocotrienols: A review. Nutr. Metab. 2014, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Gensler, H.L.; Magdaleno, M. Topical vitamin E inhibition of immunosuppression and tumorigenesis induced by ultraviolet irradiation. Nutr. Cancer 1991, 15, 97–106. [Google Scholar] [CrossRef]
- Dorgan, J.F.; Schatzkin, A. Antioxidant micronutrients in cancer prevention. Hematol. Oncol. Clin. N. Am. 1991, 5, 43–68. [Google Scholar] [CrossRef]
- Greenberg, E.R.; Baron, J.A.; Tosteson, T.D.; Freeman, D.H., Jr.; Beck, G.J.; Bond, J.H.; Colacchio, T.A.; Coller, J.A.; Frankl, H.D.; Haile, R.W.; et al. A clinical trial of antioxidant vitamins to prevent colorectal adenoma. Polyp Prevention Study Group. N. Engl. J. Med. 1994, 331, 141–147. [Google Scholar] [CrossRef]
- Albanes, D.; Heinonen, O.P.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; Freedman, L.S.; Haapakoski, J.; et al. Alpha-Tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: Effects of base-line characteristics and study compliance. J. Natl. Cancer Inst. 1996, 88, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Rautalahti, M.; Virtamo, J.; Haukka, J.; Heinonen, O.P.; Sundvall, J.; Albanes, D.; Huttunen, J.K. The effect of alpha-tocopherol and beta-carotene supplementation on COPD symptoms. Am. J. Respir. Crit. Care Med. 1997, 156, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.Y.; Ward, L.C.; Fong, C.W.; Yap, W.N.; Brown, L. Anti-inflammatory gamma- and delta-tocotrienols improve cardiovascular, liver and metabolic function in diet-induced obese rats. Eur. J. Nutr. 2017, 56, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Serbinova, E.; Kagan, V.; Han, D.; Packer, L. Free radical recycling and intramembrane mobility in the antioxidant properties of alpha-tocopherol and alpha-tocotrienol. Free Radic. Biol. Med. 1991, 10, 263–275. [Google Scholar] [CrossRef]
- Parker, R.A.; Pearce, B.C.; Clark, R.W.; Gordon, D.A.; Wright, J.J. Tocotrienols regulate cholesterol production in mammalian cells by post-transcriptional suppression of 3-hydroxy-3-methylglutaryl-coenzyme A reductase. J. Biol. Chem. 1993, 268, 11230–11238. [Google Scholar] [CrossRef]
- McIntyre, B.S.; Briski, K.P.; Tirmenstein, M.A.; Fariss, M.W.; Gapor, A.; Sylvester, P.W. Antiproliferative and apoptotic effects of tocopherols and tocotrienols on normal mouse mammary epithelial cells. Lipids 2000, 35, 171–180. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S.; Packer, L. Molecular basis of vitamin E action. Tocotrienol potently inhibits glutamate-induced pp60(c-Src) kinase activation and death of HT4 neuronal cells. J. Biol. Chem. 2000, 275, 13049–13055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.; Mo, H. Potential of tocotrienols in the prevention and therapy of Alzheimer’s disease. J. Nutr. Biochem. 2016, 31, 1–9. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishio, K.; Horie, M.; Akazawa, Y.; Shichiri, M.; Iwahashi, H.; Hagihara, Y.; Yoshida, Y.; Niki, E. Attenuation of lipopolysaccharide (LPS)-induced cytotoxicity by tocopherols and tocotrienols. Redox Biol. 2013, 1, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Krishnan, K.; Aggarwal, B.B. γ-Tocotrienol inhibits nuclear factor-κB signaling pathway through inhibition of receptor-interacting protein and TAK1 leading to suppression of antiapoptotic gene products and potentiation of apoptosis. J. Biol. Chem. 2007, 282, 809–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walda, I.C.; Tabak, C.; Smit, H.A.; Rasanen, L.; Fidanza, F.; Menotti, A.; Nissinen, A.; Feskens, E.J.; Kromhout, D. Diet and 20-year chronic obstructive pulmonary disease mortality in middle-aged men from three European countries. Eur. J. Clin. Nutr. 2002, 56, 638–643. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Mo, H.; Packer, L.; Peterson, D.M. Isolation and identification of novel tocotrienols from rice bran with hypocholesterolemic, antioxidant, and antitumor properties. J. Agric. Food Chem. 2000, 48, 3130–3140. [Google Scholar] [CrossRef]
- Ji, X.; Wang, Z.; Geamanu, A.; Sarkar, F.H.; Gupta, S.V. Inhibition of cell growth and induction of apoptosis in non-small cell lung cancer cells by delta-tocotrienol is associated with notch-1 down-regulation. J. Cell Biochem. 2011, 112, 2773–2783. [Google Scholar] [CrossRef] [PubMed]
- Muid, S.; Froemming, G.R.; Rahman, T.; Ali, A.M.; Nawawi, H.M. Delta- and gamma-tocotrienol isomers are potent in inhibiting inflammation and endothelial activation in stimulated human endothelial cells. Food Nutr. Res. 2016, 60, 31526. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Reis, J.C.; Papasian, C.J.; Morrison, D.C.; Qureshi, N. Tocotrienols inhibit lipopolysaccharide-induced pro-inflammatory cytokines in macrophages of female mice. Lipids Health Dis. 2010, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Buckner, T.; Fan, R.; Kim, Y.; Kim, J.; Chung, S. Annatto tocotrienol attenuates NLRP3 inflammasome activation in macrophages. Curr. Dev. Nutr. 2017, 1, e000760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Park, N.Y.; Jang, Y.; Ma, A.; Jiang, Q. Vitamin e gamma-tocotrienol inhibits cytokine-stimulated NF-κB activation by induction of anti-inflammatory a20 via stress adaptive response due to modulation of sphingolipids. J. Immunol. 2015, 195, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Gromovsky, A.D.; Brown, J.M.; Chung, S. Gamma-tocotrienol attenuates the aberrant lipid mediator production in NLRP3 inflammasome-stimulated macrophages. J. Nutr. Biochem. 2018, 58, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Kang, I.; Fang, X.; Wang, W.; Lee, M.A.; Hollins, R.R.; Marshall, M.R.; Chung, S. Gamma-tocotrienol attenuates high-fat diet-induced obesity and insulin resistance by inhibiting adipose inflammation and M1 macrophage recruitment. Int. J. Obes. 2015, 39, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, Q. gamma-Tocotrienol inhibits lipopolysaccharide-induced interlukin-6 and granulocyte colony-stimulating factor by suppressing C/EBPbeta and NF-kappaB in macrophages. J. Nutr. Biochem. 2013, 24, 1146–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yam, M.L.; Abdul Hafid, S.R.; Cheng, H.M.; Nesaretnam, K. Tocotrienols suppress proinflammatory markers and cyclooxygenase-2 expression in RAW264.7 macrophages. Lipids 2009, 44, 787–797. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, Q. Vitamin E delta-tocotrienol inhibits TNF-alpha-stimulated NF-kappaB activation by up-regulation of anti-inflammatory A20 via modulation of sphingolipid including elevation of intracellular dihydroceramides. J. Nutr. Biochem. 2019, 64, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Casati, L.; Pagani, F.; Limonta, P.; Vanetti, C.; Stancari, G.; Sibilia, V. Beneficial effects of delta-tocotrienol against oxidative stress in osteoblastic cells: Studies on the mechanisms of action. Eur. J. Nutr. 2020, 59, 1975–1987. [Google Scholar] [CrossRef] [Green Version]
- Harlan, L.; Mena, L.T.; Ramalingam, L.; Jayarathne, S.; Shen, C.L.; Moustaid-Moussa, N. Mechanisms Mediating Anti-Inflammatory Effects of Delta-Tocotrienol and Tart Cherry Anthocyanins in 3T3-L1 Adipocytes. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Matsunaga, T.; Shoji, A.; Gu, N.; Joo, E.; Li, S.; Adachi, T.; Yamazaki, H.; Yasuda, K.; Kondoh, T.; Tsuda, K. gamma-Tocotrienol attenuates TNF-alpha-induced changes in secretion and gene expression of MCP-1, IL-6 and adiponectin in 3T3-L1 adipocytes. Mol. Med. Rep. 2012, 5, 905–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Yagiz, Y.; Xu, C.; Lu, J.; Chung, S.; Marshall, M.R. Muscadine grape seed oil as a novel source of tocotrienols to reduce adipogenesis and adipocyte inflammation. Food Funct. 2015, 6, 2293–2302. [Google Scholar] [CrossRef]
- Shamaan, N.A.; Wan Ngah, W.Z.; Ibrahim, R.; Jarien, Z.; Top, A.G.; Abdul Kadir, K. Effect of tocotrienol on the activities of cytosolic glutathione-dependent enzymes in rats treated with 2-acetylaminofluorene. Biochem. Pharm. 1993, 45, 1517–1519. [Google Scholar] [CrossRef]
- Allen, L.; Ramalingam, L.; Menikdiwela, K.; Scoggin, S.; Shen, C.L.; Tomison, M.D.; Kaur, G.; Dufour, J.M.; Chung, E.; Kalupahana, N.S.; et al. Effects of delta-tocotrienol on obesity-related adipocyte hypertrophy, inflammation and hepatic steatosis in high-fat-fed mice. J. Nutr. Biochem. 2017, 48, 128–137. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Tan, X.; Reis, J.C.; Badr, M.Z.; Papasian, C.J.; Morrison, D.C.; Qureshi, N. Suppression of nitric oxide induction and pro-inflammatory cytokines by novel proteasome inhibitors in various experimental models. Lipids Health Dis. 2011, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Peh, H.Y.; Ho, W.E.; Cheng, C.; Chan, T.K.; Seow, A.C.; Lim, A.Y.; Fong, C.W.; Seng, K.Y.; Ong, C.N.; Wong, W.S. Vitamin E Isoform gamma-Tocotrienol Downregulates House Dust Mite-Induced Asthma. J. Immunol. 2015, 195, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Natarajan, S.K.; Chung, S. Gamma-Tocotrienol Attenuates the Hepatic Inflammation and Fibrosis by Suppressing Endoplasmic Reticulum Stress in Mice. Mol. Nutr. Food Res. 2018, 62, e1800519. [Google Scholar] [CrossRef]
- Zainal, Z.; Abdul Rahim, A.; Khaza’ai, H.; Chang, S.K. Effects of Palm Oil Tocotrienol-Rich Fraction (TRF) and Carotenes in Ovalbumin (OVA)-Challenged Asthmatic Brown Norway Rats. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agler, A.H.; Kurth, T.; Gaziano, J.M.; Buring, J.E.; Cassano, P.A. Randomised vitamin E supplementation and risk of chronic lung disease in the Women’s Health Study. Thorax 2011, 66, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Watson, L.; Margetts, B.; Howarth, P.; Dorward, M.; Thompson, R.; Little, P. The association between diet and chronic obstructive pulmonary disease in subjects selected from general practice. Eur. Respir. J. 2002, 20, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Tabak, C.; Smit, H.A.; Heederik, D.; Ocke, M.C.; Kromhout, D. Diet and chronic obstructive pulmonary disease: Independent beneficial effects of fruits, whole grains, and alcohol (the MORGEN study). Clin. Exp. Allergy 2001, 31, 747–755. [Google Scholar] [CrossRef]
- Celik, F.; Topcu, F. Nutritional risk factors for the development of chronic obstructive pulmonary disease (COPD) in male smokers. Clin. Nutr. 2006, 25, 955–961. [Google Scholar] [CrossRef] [PubMed]
- De Batlle, J.; Barreiro, E.; Romieu, I.; Mendez, M.; Gomez, F.P.; Balcells, E.; Ferrer, J.; Orozco-Levi, M.; Gea, J.; Anto, J.M.; et al. Dietary modulation of oxidative stress in chronic obstructive pulmonary disease patients. Free Radic. Res. 2010, 44, 1296–1303. [Google Scholar] [CrossRef] [PubMed]
- Bolton-Smith, C.; Casey, C.E.; Gey, K.F.; Smith, W.C.; Tunstall-Pedoe, H. Antioxidant vitamin intakes assessed using a food-frequency questionnaire: Correlation with biochemical status in smokers and non-smokers. Br. J. Nutr. 1991, 65, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardestani, M.E.; Onvani, S.; Esmailzadeh, A.; Feizi, A.; Azadbakht, L. Adherence to Dietary Approaches to Stop Hypertension (DASH) Dietary Pattern in Relation to Chronic Obstructive Pulmonary Disease (COPD): A Case-Control Study. J. Am. Coll. Nutr. 2017, 36, 549–555. [Google Scholar] [CrossRef]
- Guertin, K.A.; Grant, R.K.; Arnold, K.B.; Burwell, L.; Hartline, J.; Goodman, P.J.; Minasian, L.M.; Lippman, S.M.; Klein, E.; Cassano, P.A. Effect of long-term vitamin E and selenium supplementation on urine F2-isoprostanes, a biomarker of oxidative stress. Free Radic. Biol. Med. 2016, 95, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grievink, L.; Smit, H.A.; Ocke, M.C.; van’t Veer, P.; Kromhout, D. Dietary intake of antioxidant (pro)-vitamins, respiratory symptoms and pulmonary function: The MORGEN study. Thorax 1998, 53, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, H.A.; Grievink, L.; Tabak, C. Dietary influences on chronic obstructive lung disease and asthma: A review of the epidemiological evidence. Proc. Nutr. Soc. 1999, 58, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Hackney, J.D.; Linn, W.S.; Buckley, R.D.; Jones, M.P.; Wightman, L.H.; Karuza, S.K.; Blessey, R.L.; Hislop, H.J. Vitamin E supplementation and respiratory effects of ozone in humans. J. Toxicol. Environ. Health 1981, 7, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Raj, H.G.; Chhabra, S.K. Increased oxidative stress and altered levels of antioxidants in chronic obstructive pulmonary disease. Inflammation 2005, 29, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Pavord, I.D.; Richards, K.A.; Knox, A.J.; Wisniewski, A.F.; Lewis, S.A.; Tattersfield, A.E.; Weiss, S.T. Dietary antioxidant vitamin intake and lung function in the general population. Am. J. Respir. Crit. Care Med. 1995, 151, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Daga, M.K.; Chhabra, R.; Sharma, B.; Mishra, T.K. Effects of exogenous vitamin E supplementation on the levels of oxidants and antioxidants in chronic obstructive pulmonary disease. J. Biosci. 2003, 28, 7–11. [Google Scholar] [CrossRef]
- Pervez, M.A.; Khan, D.A.; Ijaz, A.; Khan, S. Effects of Delta-tocotrienol Supplementation on Liver Enzymes, Inflammation, Oxidative stress and Hepatic Steatosis in Patients with Nonalcoholic Fatty Liver Disease. Turk. J. Gastroenterol. 2018, 29, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Zuvanich, E.G.; Khan, D.A.; Mushtaq, S.; Silswal, N.; Qureshi, N. Proteasome inhibitors modulate anticancer and anti-proliferative properties via NF-kB signaling, and ubiquitin-proteasome pathways in cancer cell lines of different organs. Lipids Health Dis. 2018, 17, 62. [Google Scholar] [CrossRef] [Green Version]
- Muid, S.; Hamid, Z.; Nawawi, H. Tocotrienol rich fraction supplement reduces oxidative stress in non familial hypercholesterolaemia: Beyond the lipid lowering capability. Int. Food Res. J. 2018, 25, 9. [Google Scholar]
- Goon, J.A.; Azman, N.H.E.N.; Ghani, S.M.A.; Hamid, Z.; Ngah, W.Z.W. Comparing palm oil tocotrienol rich fraction with a-tocopherol supplementation on oxidative stress in healthy older adults. Clin. Nutr. Espen. 2017, 21, 12. [Google Scholar] [CrossRef]
- Tan, B.; Watson, R.R.; Preedy, V.R. Tocotrienols Vitamin E Beyond Tocopherols, 2nd ed.; Taylor & Frances: Oxford, UK, 2016. [Google Scholar]
- Black, T.M.; Wang, P.; Maeda, N.; Coleman, R.A. Palm tocotrienols protect ApoE +/− mice from diet-induced atheroma formation. J. Nutr. 2000, 130, 2420–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traber, M.G.; Burton, G.W.; Hamilton, R.L. Vitamin E trafficking. Ann. N. Y. Acad. Sci. 2004, 1031, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mahipal, A.; Klapman, J.; Vignesh, S.; Yang, C.S.; Neuger, A.; Chen, D.T.; Malafa, M.P. Pharmacokinetics and safety of vitamin E delta-tocotrienol after single and multiple doses in healthy subjects with measurement of vitamin E metabolites. Cancer Chemother. Pharmacol. 2016, 78, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, A.A.; Khan, D.A.; Silswal, N.; Saleem, S.; Qureshi, N. Evaluation of Pharmacokinetics, and Bioavailability of Higher Doses of Tocotrienols in Healthy Fed Humans. J. Clin. Exp. Cardiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Fairus, S.; Nor, R.M.; Cheng, H.M.; Sundram, K. Postprandial metabolic fate of tocotrienol-rich vitamin E differs significantly from that of alpha-tocopherol. Am. J. Clin. Nutr. 2006, 84, 835–842. [Google Scholar] [CrossRef] [Green Version]
- Khosla, P.; Patel, V.; Whinter, J.M.; Khanna, S.; Rakhkovskaya, M.; Roy, S.; Sen, C.K. Postprandial levels of the natural vitamin E tocotrienol in human circulation. Antioxid. Redox Signal. 2006, 8, 1059–1068. [Google Scholar] [CrossRef]
- Hor, C.P.; Fung, W.Y.; Ang, H.A.; Lim, S.C.; Kam, L.Y.; Sim, S.W.; Lim, L.H.; Choon, W.Y.; Wong, J.W.; Yuen, K.H.; et al. Efficacy of Oral Mixed Tocotrienols in Diabetic Peripheral Neuropathy: A Randomized Clinical Trial. JAMA Neurol. 2018, 75, 444–452. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Khan, D.A.; Mahjabeen, W.; Trias, A.M.; Silswal, N.; Qureshi, N. Impact of δ-Tocotrienol on Inflammatory Biomarkers and Oxidative Stress in. Hypercholesterolemic Subjects. J. Clin. Exp. Cardiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, C.B.; Andersen, R.F.; Steffensen, K.D.; Adimi, P.; Jakobsen, A. Delta tocotrienol in recurrent ovarian cancer. A phase II trial. Pharm. Res. 2019, 141, 392–396. [Google Scholar] [CrossRef]
- Khanna, S.; Patel, V.; Rink, C.; Roy, S.; Sen, C.K. Delivery of orally supplemented alpha-tocotrienol to vital organs of rats and tocopherol-transport protein deficient mice. Free Radic. Biol. Med. 2005, 39, 1310–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, M.T.; Becker, E.M.; Skibsted, L.H. Molecular mechanism of antioxidant synergism of tocotrienols and carotenoids in palm oil. J. Agric. Food Chem. 2006, 54, 3445–3453. [Google Scholar] [CrossRef]
- Panfili, G.; Fratianni, A.; Irano, M. Normal phase high-performance liquid chromatography method for the determination of tocopherols and tocotrienols in cereals. J. Agric. Food Chem. 2003, 51, 3940–3944. [Google Scholar] [CrossRef] [PubMed]
- Yap, S.P.; Yuen, K.H.; Wong, J.W. Pharmacokinetics and bioavailability of alpha-, gamma- and delta-tocotrienols under different food status. J. Pharm. Pharmacol. 2001, 53, 67–71. [Google Scholar] [CrossRef]
- Panagabko, C.; Morley, S.; Hernandez, M.; Cassolato, P.; Gordon, H.; Parsons, R.; Manor, D.; Atkinson, J. Ligand specificity in the CRAL-TRIO protein family. Biochemistry 2003, 42, 6467–6474. [Google Scholar] [CrossRef]
- Shibata, A.; Nakagawa, K.; Shirakawa, H.; Kobayashi, T.; Kawakami, Y.; Takashima, R.; Ohashi, A.; Sato, S.; Ohsaki, Y.; Kimura, F.; et al. Physiological effects and tissue distribution from large doses of tocotrienol in rats. Biosci. Biotechnol. Biochem. 2012, 76, 1805–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, Y.; Tsuzuki, T.; Nakagawa, K.; Miyazawa, T. Distribution of tocotrienols in rats fed a rice bran tocotrienol concentrate. Biosci. Biotechnol. Biochem. 2007, 71, 464–471. [Google Scholar] [CrossRef]
- Alqahtani, S.; Alayoubi, A.; Nazzal, S.; Sylvester, P.W.; Kaddoumi, A. Enhanced solubility and oral bioavailability of gamma-tocotrienol using a self-emulsifying drug delivery system (SEDDS). Lipids 2014, 49, 819–829. [Google Scholar] [CrossRef]
- Alqahtani, S.; Simon, L.; Astete, C.E.; Alayoubi, A.; Sylvester, P.W.; Nazzal, S.; Shen, Y.; Xu, Z.; Kaddoumi, A.; Sabliov, C.M. Cellular uptake, antioxidant and antiproliferative activity of entrapped alpha-tocopherol and gamma-tocotrienol in poly (lactic-co-glycolic) acid (PLGA) and chitosan covered PLGA nanoparticles (PLGA-Chi). J. Colloid Interface Sci. 2015, 445, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Ananthula, S.; Parajuli, P.; Behery, F.A.; Alayoubi, A.Y.; El Sayed, K.A.; Nazzal, S.; Sylvester, P.W. Oxazine derivatives of gamma- and delta-tocotrienol display enhanced anticancer activity in vivo. Anticancer Res. 2014, 34, 2715–2726. [Google Scholar] [PubMed]
- Allen-Ramey, F.C.; Gupta, S.; DiBonaventura, M.D. Patient characteristics, treatment patterns, and health outcomes among COPD phenotypes. Int. J. Chron. Obstr. Pulmon. Dis. 2012, 7, 779–787. [Google Scholar] [CrossRef] [Green Version]


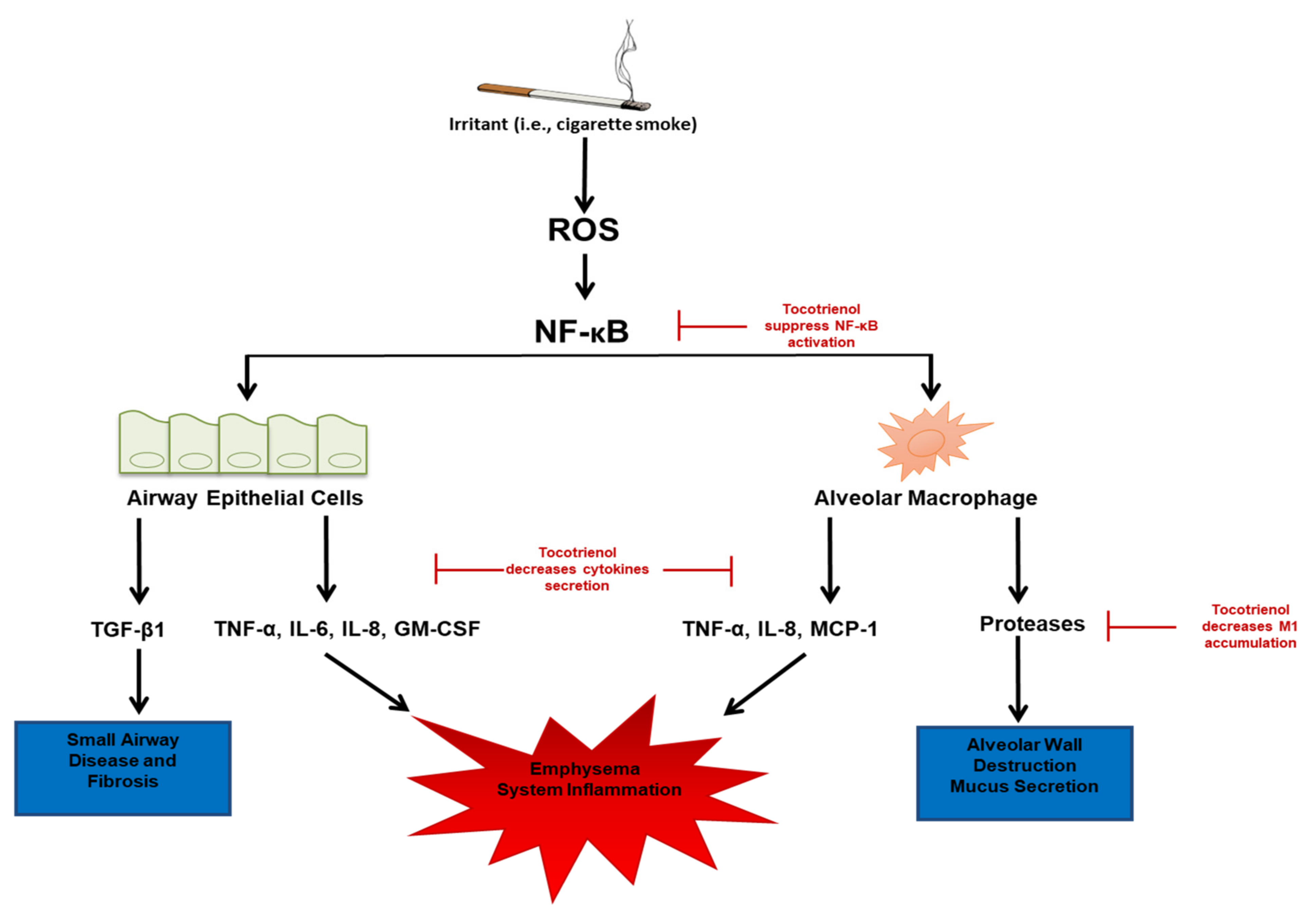{kind=link}
| Model | Vitamin E Types | Mechanism | Reference |
|---|---|---|---|
| Cell Culture | |||
| rat liver microsomes | α-tocotrienol | α-Tocotrienol has 40–60 times higher antioxidant activity than α-tocopherol. | [66] |
| human lung carcinoma A549 cell | α- and γ-tocotrienol | Both isoforms reduce ROS formation, lipid peroxidation, cytokine production, and apoptosis; γ-tocotrienol ameliorates the LPS-induced reduction in cell viability. | [72] |
| human myeloid KBM-5 cells, lung adenocarcinoma cells H1299, breast cancer MCF7, multiple myeloma U266, squamous cell carcinoma SCC4 | γ-tocotrienol | 🠗 NFκB activation by different stimulants (LPS, EGF, TNF-α) | [73] |
| Human lung cancer cell lines A549 and | δ -tocotrienol | 🠗 cancer cells proliferation, migration and invasion | [74,75,76] |
| LPS-induced inflammation in human umbilical vein endothelial cells (HUVECs) | δ- and γ-tocotrienol | 🠗 IL-6, ICAM-1, VCAM-1 and NFκB; 🠗 e-selectin and eNOs; | [77] |
| RAW 264.7 macrophages and peritoneal macrophages isolated from LPS-treated BALB/c mice | α-, δ-, and γ-tocotrienol | 🠗 TNF-α dose-dependently; 🠗 gene expression of TNF-α, IL-1β, IL-6, iNOS; δ -tocotrienol is most effective | [78] |
| iJ774 macrophages | δ-tocotrienol | 🠗 ROS, IL-1β, NLRP3 inflammasome | [79] |
| murine RAW 264.7 macrophages and primary bone marrow-derived macrophages | γ-tocotrienol | 🠕 inhibitors of NFκB (A20, Cezanne) and de novo synthesis of sphingolipid | [80] |
| bone marrow derived macrophage | γ-tocotrienol | 🠗 glucose intake 🠗 LPS-induced NFκB activation | [81] |
| bone marrow derived macrophage | γ-tocotrienol | 🠗 CD11b, TNF-α, IL-6, and IL-1β 🠗 LPS-induced M1 macrophage polarization and NFκB activation | [82] |
| LPS-induced inflammation in murine RAW 264.7 macrophages | γ-tocotrienol | 🠗 IL-6 and G-CSF production through the inhibition of NFκB and C/EBPβ pathways. | [83] |
| murine RAW 264.7 macrophages | tocotrienol rich fraction; α-, δ-, and γ-tocotrienol | 🠗 production of inflammatory products (TNF-α, IL-6, NO, COX-2) | [84] |
| murine RAW 264.7 macrophages. | δ-tocotrienol | 🠗 NFκB activation 🠗 phosphorylation of TAK1 🠕 up-regulation of A20 and CYLD | [85] |
| osteoblastic cells | δ-tocotrienol | 🠗 NFκB downstream target | [86] |
| murine 3T3-L1 adipocytes | δ-tocotrienol | 🠗 secretion of adipokines IL-6 🠗 JNK inflammation pathway 🠗 NFκB activation | [87] |
| murine 3T3-L1 adipocytes | γ-tocotrienol | 🠗 TNF-α in adipocytes; 🠗 secretion of adipokines IL-6, MCP-1 🠗 NFκB activation | [88] |
| human adipose-derived stem cells | muscadine grape seed oil containing α- and γ-tocotrienol | 🠗 LPS-induced IL-6, IL-8, and MCP-1 | [89] |
| Animal Study | |||
| cigarette-smoke induced model of COPD in a rat model | γ-tocotrienol | 🠗 reduces cigarette smoke-induced BAL fluid neutrophil counts and inflammatory cytokine concentrations dose-dependently | [47] |
| metabolic syndrome and bone loss in rats | palm tocotrienol | 🠕 skeletal-promoting benefit by modulating the levels of osteocytes- derived bone-related peptides | [65] |
| BALB/c mice | α-, δ-, and γ-tocotrienol | 🠗 TNF-α, IL-1β, IL-6 and iNOS | [78] |
| obesity induced-inflammation and insulin resistance in C57BL/6J mice | γ-tocotrienol | 🠗 body weight gain, improved insulin signaling 🠗 MCP-1 and macrophage recruitment into adipose tissue | [82] |
| obesity in mice | δ-tocotrienol | 🠗 fat cell hypertrophy and inflammation in both liver and adipose tissue | [90,91] |
| C57BL/6, BALB/c, LMP7/MECL-1-/-, and PPARα-/- mice | δ-tocotrienol | 🠗 TNF-α, iNOS induction, and NO production. | [92] |
| house dust-mite- mediated asthma model in BALB/c mice | γ-tocotrienol | 🠗 productions of free radicals, cytokines, chemokines, ROS, oxidative damage biomarkers, NFκB 🠕 nuclear Nrf2, endogenous antioxidant activity | [93] |
| C57BL/6 male mice with high fat diet inducing nonalcoholic fatty liver disease (NAFLD) | γ-tocotrienol | 🠗 productions reduce the diet-induced hepatic ER stress and fibrosis | [94] |
| C57BL/6 male mice ovalbumin (OVA)-challenged asthmatic brown Norway rats | γ-tocotrienol palm oil tocotrienol-rich fraction (TRF) | 🠗 hepatic inflammation and the expression of MCP-1, CDE11c, TNF-α, NLRP3, and IL-1β 🠗 reduces edema and inflammatory cell infiltration in the bronchial wall | [94,95] |
| Human Chronic and Intervention Studies | |||
| 2917 men aged 50–69 y | Dietary Vitamin E | Vitamin E intake prevent the development of COPD | [74] |
| Total 38,597 women without chronic lung disease in the Women’s Health Study (WHS) | randomised double-blind placebo-controlled factorial trial of vitamin E (600 IU every other day) | 600 IU vitamin E led to a 10% reduction in the risk of chronic lung disease in women | [96] |
| 115 COPD patients and 115 controls | Spirometry and food questionnair were used | fruit and vegetable consumption is inversely associated with chronic obstructive pulmonary disease | [97] |
| MORGEN study 13 651 men and women aged 20–59 years for 20 years | fruit, vegetable, fish, alcohol, and whole grain consumption | independent beneficial effects of fruits, whole grains on COPD | [98] |
| 40 male smokers with clinical diagnosis of COPD (Group-I (GI)) and 36 healthy smokers without COPD | 65 food items from five main food groups (grain, meat and alternatives, dairy products, vegetables-fruits, and fat) and 25 dietary habits. | Dietary intake of black tea and vegetables-fruits consumptions may be protecting male smokers from developing COPD | [99] |
| A total of 267 patients with COPD | Dietary data of the last 2 years was assessed using a validated food questionnaire | dietary vitamin E intake prevents systemic oxidative stress in COPD patients, particularly in those that continue smoking. | [100] |
| 196 Scottish Men in smoker and non-smokers | Food Frequency Questionnaire and validated by serum samples | Both dietary and serum values of vitamin E were lower in smokers than non-smokers | [101] |
| 84 patients with COPD and 80 controls | Dietary Approaches to Stop Hypertension (DASH) diet | DASH dietary pattern among patients with COPD was significantly lower compared to the control group. Cough was significantly decreased by increments in adherence to a DASH dietary | [102] |
| Total 35,533 subjects | randomized vitamin E supplementation | Vitamin E supplementation decreases urine 8-iso-PGF2α among male cigarette smokers | [103] |
| Dutch population on 6555 adults during 1994 and 1995 | Asemi-quantitative food frequency questionnaire and respiratory symptoms were assessed by a self-administered questionnaire. | Vitamin E intake showed no association with most symptoms and lung function, but had a positive association with productive cough. | [104] |
| Subjects data were collected in the 1960s in Finland (1248), Italy (1386), and the Netherlands (691) | The cross-check dietary history method was used and lung function was measured by the Spirometry | Associations of vitamin E with pulmonary function were not consistent across countries | [105] |
| young healthy adult volunteers exposed for 2 h periods to 0.5 ppm O3, with secondary stresses of heat and intermittent light exercise. | Subjects received 800 or 1600 IU vitamin E per day for 9 or more wk | There is not significantly difference between vitamin E and placebo groups | [106] |
| 82 patients with COPD and 22 healthy non-smoking controls | Lung function was measured by spirometry. | systemic oxidant-antioxidant imbalance in the patients with COPD. | [107] |
| 2633 subjects 18 to 70 yr | Vitamin E by semiquantitative food frequency questionnair | higher intake of vitamin E was associated with better lung function | [108] |
| Thirty patients with COPD with 12 weeks of supplementation with 400 IU of vitamin E daily | spirometry, plasma malondialdehyde (MDA), SOD levels were measured | Vitamin E supplementation does not have any significant effect on lung function but it lowers the levels of MDA | [109] |
| 71 patients with non-alcoholic fatty liver disease (NAFLD) | oral supplementation of δ-tocotrienol | 🠗 hs-CRP, MDA, and FLI score in comparison to the placebo group | [110] |
| 14 patients with hepatitis C | δ-tocotrienol (500 mg/d) for 6 weeks | 🠗 δ-tocotrienol inhibits multiple-signaling pathways such as TNF-α, LMP2, 7 and 10, IFN-γ, ICAM1, VCAM1 without any side- | [111] |
| non-familial hypercholesterolmia patients | 60 mg/d TRF | 🠗 MDA, F2-isoprostanes, ox-LDL | [112] |
| 71 subjects both male and female aged between 50 and 55 years | plasma MDA, protein carbonyl, total DNA damage, vitamin D concentration and vitamin E isomers were measured | 🠗 Tocotrienol supplementation effects were different from α-tocopherol in reducing oxidative stress markers | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.; Yao, H.; Meister, M.; Gardenhire, D.S.; Mo, H. Tocotrienols: Dietary Supplements for Chronic Obstructive Pulmonary Disease. Antioxidants 2021, 10, 883. https://doi.org/10.3390/antiox10060883
Ji X, Yao H, Meister M, Gardenhire DS, Mo H. Tocotrienols: Dietary Supplements for Chronic Obstructive Pulmonary Disease. Antioxidants. 2021; 10(6):883. https://doi.org/10.3390/antiox10060883
Chicago/Turabian StyleJi, Xiangming, Hongwei Yao, Maureen Meister, Douglas S. Gardenhire, and Huanbiao Mo. 2021. "Tocotrienols: Dietary Supplements for Chronic Obstructive Pulmonary Disease" Antioxidants 10, no. 6: 883. https://doi.org/10.3390/antiox10060883