
Metabolomics of Chlorophylls and Carotenoids: Analytical Methods and Metabolome-Based Studies


Abstract
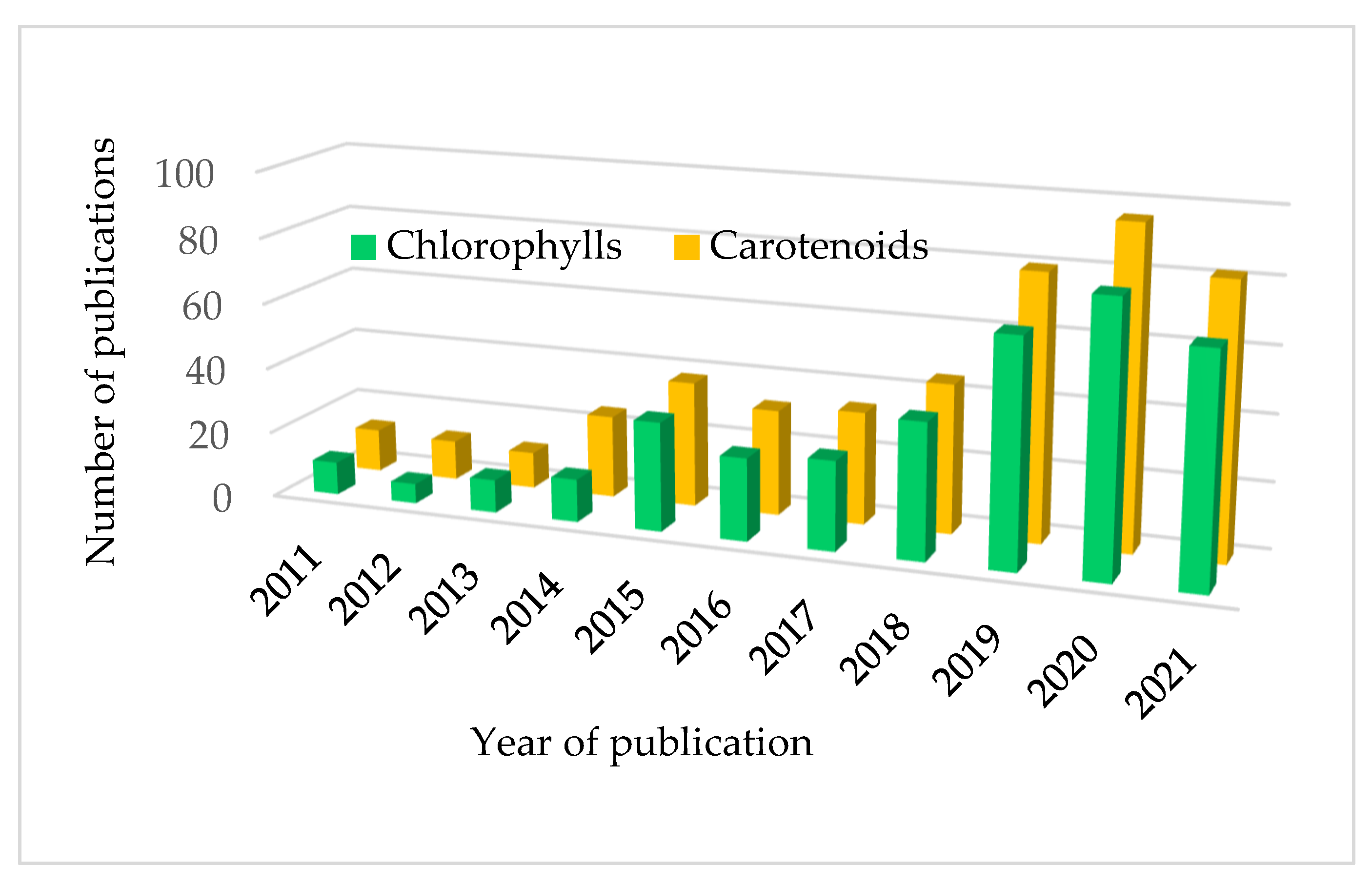
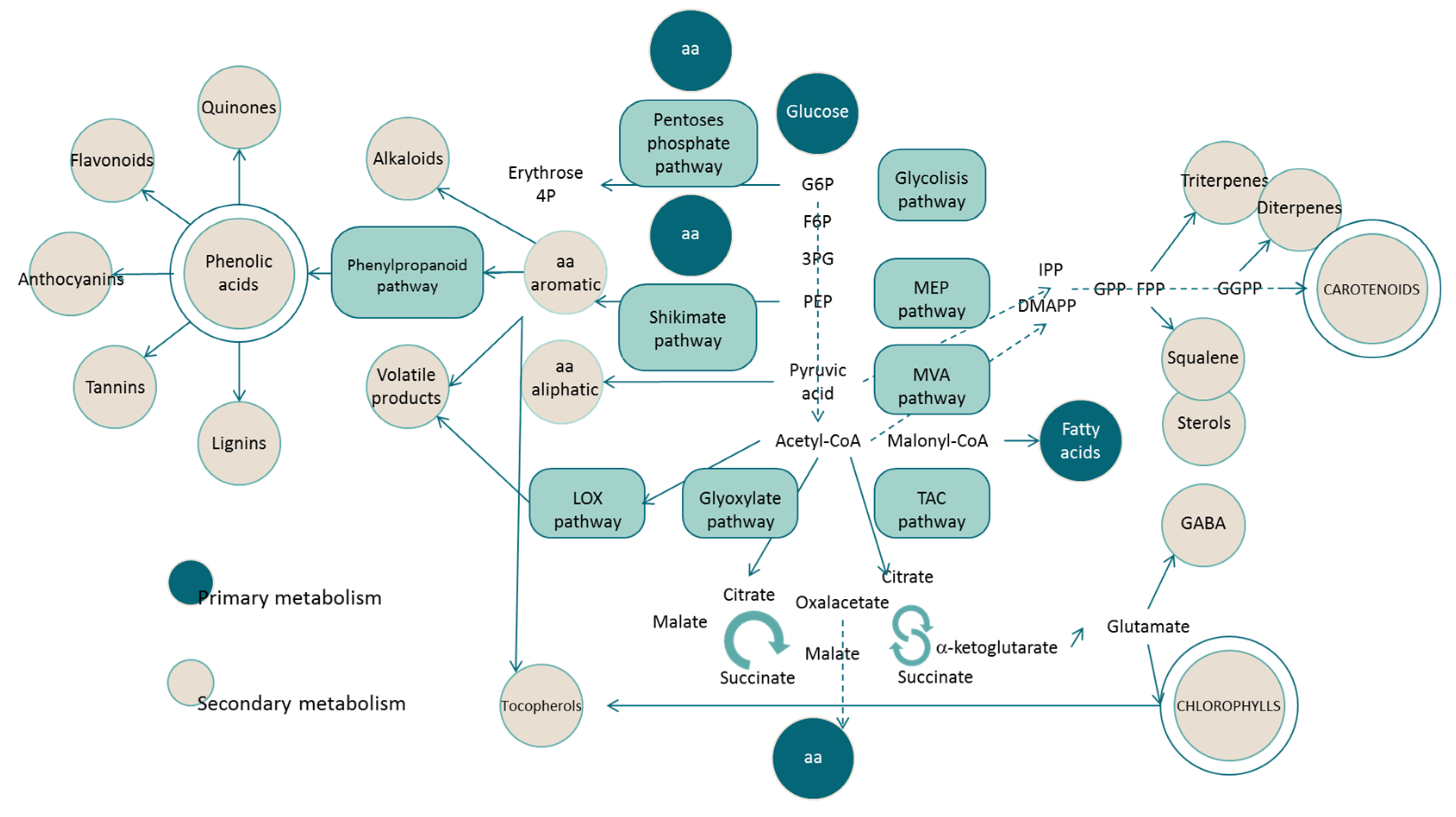
:1. Introduction
2. Biochemistry of Chlorophylls and Carotenoids
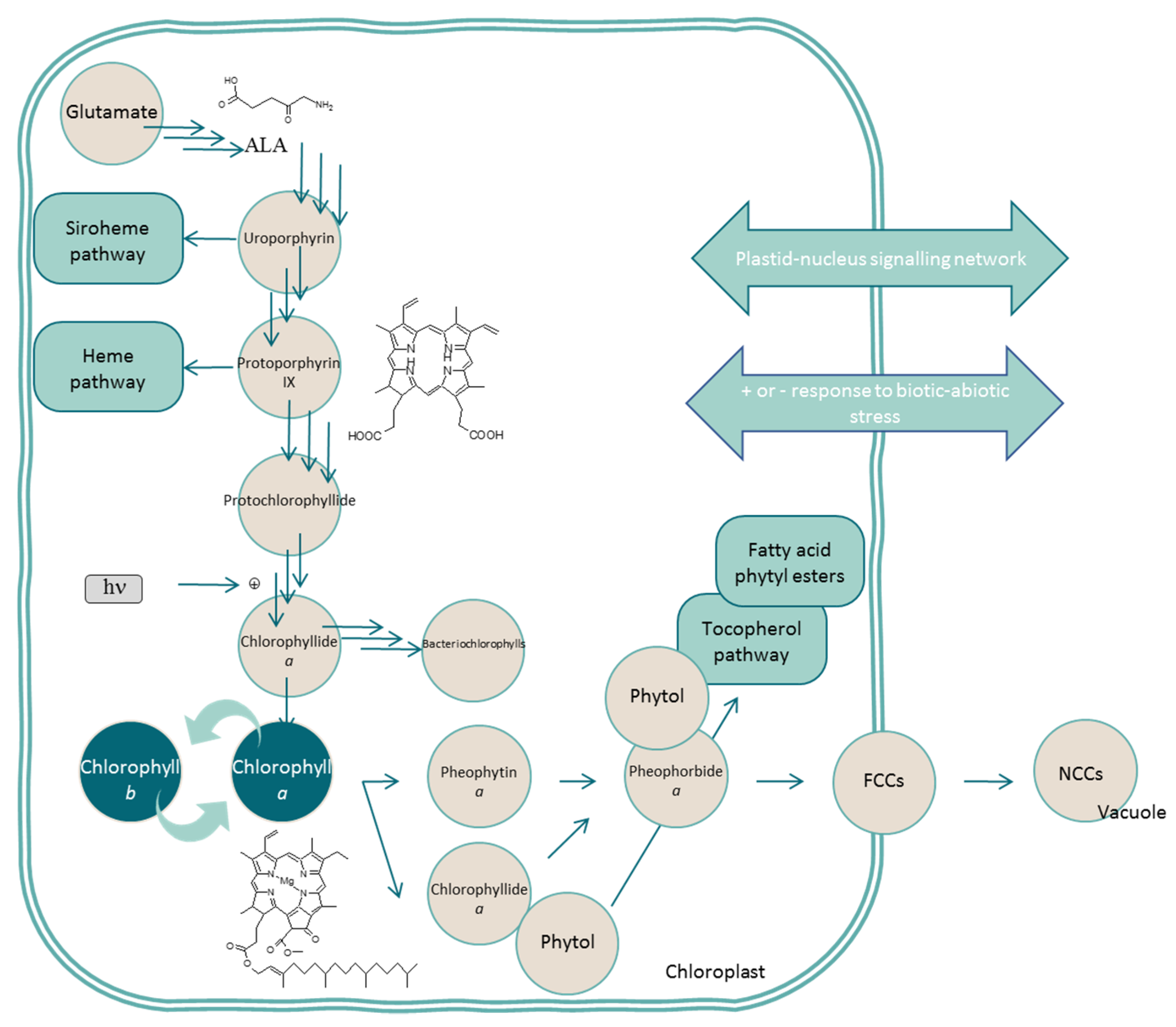
2.1. Chlorophylls
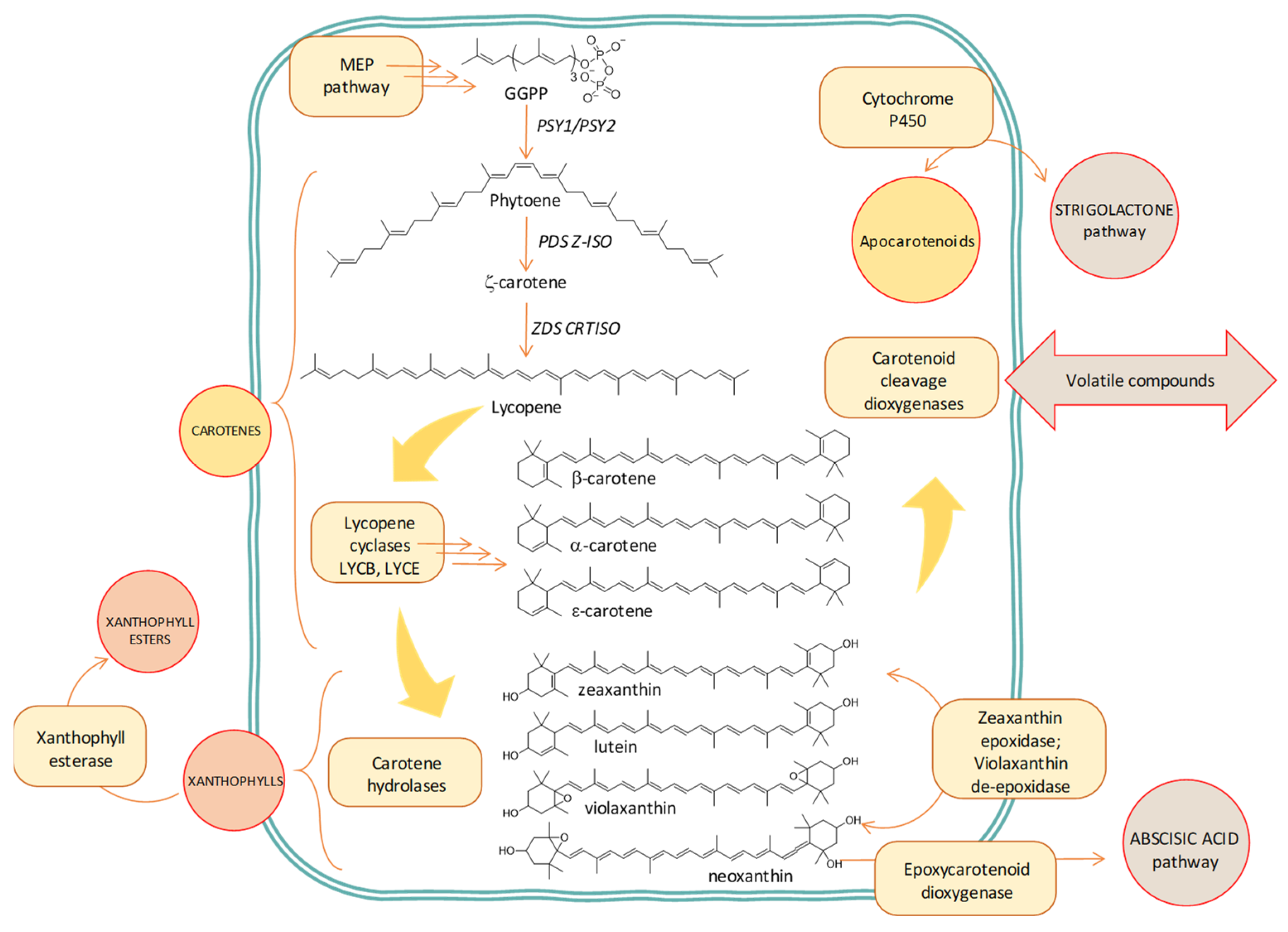
2.2. Carotenoids
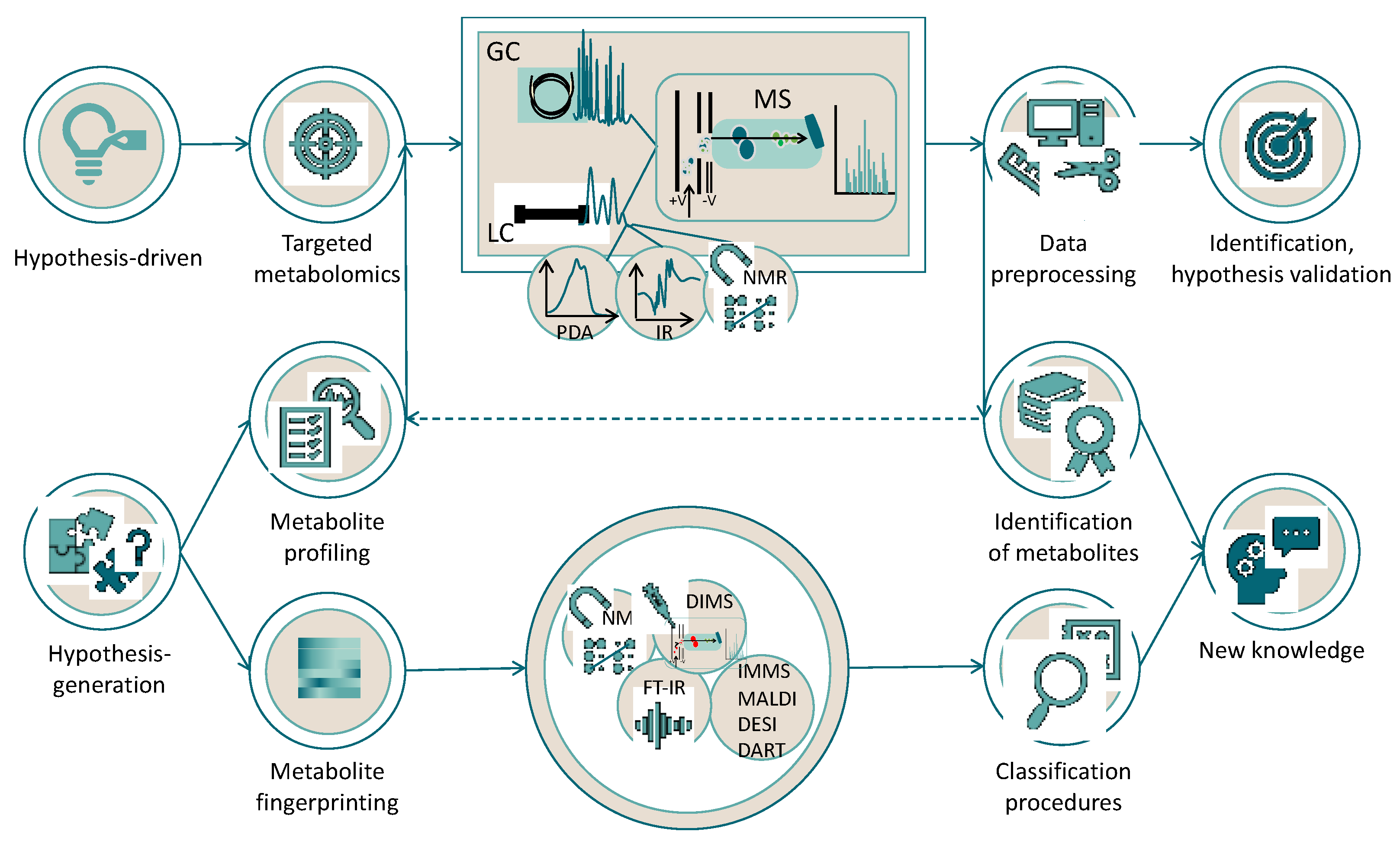
3. The Praxis of Metabolomics: Essential Steps and Challenges for the Experimental Design
3.1. Material Preparation and Extraction Protocols
3.2. Technologies: Instrumental Techniques for the Acquisition of Spectroscopic and Spectrometric Data
3.3. Application of Different Approaches to Pigment Metabolomics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Extraction Solvent | Instrumental Techniques | Strategy for Metabolomic Study | Ref. |
|---|---|---|---|---|
| A. thaliana | MeOH:H2O (8:2) | LC/ESI-TOF | Metabolite profiling based on isotope labeling-assisted elemental composition | [86] |
| A. thaliana | MeOH:MTBE:H2O (1:3:1) and subsequent separation with MeOH:H2O (1:3) | Multiplatform approach (UPLC-FT-MS and MS/MS, GC-MS, nUPLC-QTOF-MS, and MS/MS) | Metabolite profiling based on isotope labeling-assisted elemental composition | [87] |
| Olive oil, canned green vegetables | N,N-dimethylformamide | LC/APCI-ESI/hr-QTOF-MS | Metabolite profiling based on isotopic pattern | [88] |
| Microalgae | EtOH:hexane (2:1) and H2O:hexane 1:2 | UPLC-UV-TWIM-MS | Untargeted metabolomics | [82] |
| Lemon (Citrus lemon L.) | Acetone | LC/ESI/hr-QTOF-MS | Targeted metabolomics | [89] |
| A. thaliana | Ethanol | UPLC/TOF-MS | Targeted metabolomics | [92] |
| Wheat (Triticum aestivum) | Methanol:acetonitrile:water (4:4:2) | Multiplatform approach (GC-MS, GC-QTOF-MS, LC-MS, and LC-QTOF-MS) | Targeted and untargeted metabolomics | [93] |
| Tamarillo fruits (Solanum betaceum) | CO2:MeOH (95:5 or 90:10) | SFE-SFC-MS | Untargeted metabolomics | [96] |
| Tomato (Solanum lycopersicum L.) | MeOH followed by hexane:acetone (1:1) | LC-APCI-QTOF-MS | Metabolite profiling | [97] |
| A. thaliana | Chloroform:MeOH:H2O (2:6:2) and derivatization with methoxyamine hydrochloride and N-methyl-N-(trimethylsilyl) trifluoroacetamide | GC-TOF/MS | Metabolite profiling | [98] |
| S. lycopersicum L. | MeOH or MeOH:H2O (75:25) | LC-QTOF-MS and LC-PDA-FD | Metabolite profiling | [99] |
| Zea mays | MeOH and dH2O with ribitol; derivatization with methoxyamine, N,Obis(trimethylsilyl)trifluoroacetamide, and trimethylchlorosilane | GC-TOF-MS and spectrophotometry | Metabolite profiling | [100] |
| S. lycopersicum L. | MeOH and dH2O with ribitol; derivatization with methoxyamine, N,Obis(trimethylsilyl)trifluoroacetamide, and trimethylchlorosilane | GC-TOF-MS and LC-PDA | Metabolite profiling | [101] |
| Cucumis melo L. | Hexane:acetone:ethanol (50:25:25) | LC-PDA | Metabolite profiling | [102] |
| Daucus carota, Brassica oleracea, S. lycopersicum L. | MeOH:chloroform:Tris-buffer (1.25:1:1.25, 50 mM, pH 7.5) | LC-PDA, LC-PDA-QTOF-MS, GC-MS, and 1H-NMR | Targeted and untargeted metabolomics | [71] |
| Cuminum cyminum L. | N,N-dimethylformamide; trichloroacetic acid; chloroform:MeOH:phosphate buffer (1:2:0.9, pH 7.5) | Multiplatform approach (spectrophotometry, LC-PDA, LC-MS, and GC-MS) | Metabolite profiling | [103] |
| Potato (Solanum tuberosum) | MeOH:H2O (87.5:12.5) | LC-ESI-QTOF-MS | Metabolite profiling | [104] |
4. Metabolome-Based Studies of Chlorophylls and Carotenoids
4.1. Application in Abiotic Factors Studies
4.2. Application in Biotic Factor Studies
4.3. Application in Physiologic and Molecular Biology Studies
4.4. Application in Human Health (Health Status, Cancer, Hypertension, and Digestive Efficiency) Studies
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hall, R.; Beale, M.; Fiehn, O.; Hardy, N.; Sumner, L.; Bino, R. Plant metabolomics: The missing link in functional genomics strategies. Plant Cell 2002, 14, 1437–1440. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K. Modern plant metabolomics: Advanced natural product gene discoveries, improved technologies, and future prospects. Nat. Prod. Rep. 2015, 32, 212–229. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Fiehn, O. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comp. Funct. Genom. 2001, 2, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Brzezowski, P.; Richter, A.S.; Grimm, B. Regulation and function of tetrapyrrole biosynthesis in plants and algae. Biochim. Biophys. Acta 2015, 1847, 968–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuai, B.; Chen, J.; Hörtensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J. Exp. Bot. 2018, 69, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Grimm, B. Metabolism, Structure and Function of Plant Tetrapyrroles: Control Mechanisms of Chlorophyll Biosynthesis and Analysis of Chlorophyll-Binding Proteins; Academic Press: London, UK, 2019. [Google Scholar]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Biophys. Acta 2011, 1807, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vom Dorp, K.; Hölz, G.; Plohmann, C.; Eisenhut, M.; Abraham, M.; Weber, A.P.M.; Hanson, A.D.; Dörmann, P. Remobilization of phytol from chlorophyll degradation is essential for tocopherol synthesis and growth of Arabidopsis. Plant Cell 2015, 27, 2846–2859. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Gálvez, A.; Roca, M. Phyllobilins: A new group of bioactive compounds. In Studies of Natural Products Chemistry; Atta-ur-Rahman, F.R.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 159–191. [Google Scholar]
- Christ, B.; Hauenstein, M.; Hörtensteiner, S. A liquid chromatography-mass spectrometry platform for the analysis of phyllobilins, the major degradation products of chlorophyll in Arabidopsis thaliana. Plant J. 2016, 88, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Busch, A.W.; Montgomery, B.L. Interdependence of tetrapyrrole metabolism, the generation of oxidative stress and the mitigative oxidative stress response. Redox Biol. 2015, 4, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef] [Green Version]
- Wurtzel, E.T. Changing form and function through carotenoids and synthetic biology. Plant Physiol. 2019, 179, 830–843. [Google Scholar] [CrossRef] [Green Version]
- Al-Babili, S.; Bouwmeester, H.J. Strigolactones, a novel carotenoid-derived plant hormone. Annu. Rev. Plant Biol. 2015, 66, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Felemban, A.; Braguy, J.; Zurbriggen, M.D.; Al-Babili, S. Apocarotenoids involved in plant development and stress response. Front. Plant Sci. 2019, 10, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, S. Recent advances in the synthesis of carotenoid-derived flavours and fragrances. Molecules 2015, 20, 12817–12840. [Google Scholar] [CrossRef] [Green Version]
- Águila Ruiz-Sola, M.; Rodríguez-Concepción, M. Carotenoid biosynthesis in Arabidopsis: A colorful pathway. Arabidopsis Book 2012, 10, e0158. [Google Scholar] [CrossRef] [Green Version]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Viera, I.; Pérez-Gálvez, A.; Roca, M. Bioaccessibility of marine carotenoids. Mar. Drugs 2018, 16, 397. [Google Scholar] [CrossRef] [Green Version]
- Shumskaya, M.; Wurtzel, E.T. The carotenoid biosynthetic pathway: Thinking in all dimensions. Plant Sci. 2013, 208, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorente, B. Regulation of carotenoid biosynthesis in photosynthetic organs. In Carotenoids in Nature. Subcellular Biochemistry; Stange, C., Ed.; Springer: Cham, Switzerland; New York, NY, USA, 2016; Volume 79, pp. 141–160. [Google Scholar]
- Lado, J.; Zacarías, L.; Rodrigo, M.J. Regulation of carotenoid biosynthesis during fruit development. In Carotenoids in Nature. Subcellular Biochemistry; Stange, C., Ed.; Springer: Cham, Switzerland; New York, NY, USA, 2016; Volume 79, pp. 161–198. [Google Scholar]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Handbook; Birkhäuser Verlag: Basel, Switzerland, 2004. [Google Scholar]
- Rivera, S.M.; Canela-Garayoa, R. Analytical tools for the analysis of carotenoids in diverse materials. J. Chromatogr. A 2012, 1224, 1–10. [Google Scholar] [CrossRef]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry. Mass Spectrom Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Gálvez, A.; Roca, M. Recent developments in the analysis of carotenoids by mass spectrometry. In Progress in Carotenoid Research; Zepka, L., Ed.; IntechOpen: London, UK, 2018; pp. 17–44. [Google Scholar]
- Fernandes, A.S.; Petry, F.C.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. HPLC-PDA-MS/MS as a strategy to characterize and quantify natural pigments from microalgae. Curr. Res. Food Sci. 2020, 8, 100–112. [Google Scholar] [CrossRef]
- Fiehn, O.; Sumner, L.W.; Rhee, S.; Ward, J.; Dickerson, J.; Lange, B.M.; Lane, G.; Roessner, U.; Last, R.; Nikolau, B. Minimum reporting standards for plant biology context information in metabolomics studies. Metabolomics 2007, 3, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Morgan, J.A. Analysis of metabolic flux using dynamic labelling and metabolic modelling. Plant Cell. Environ. 2013, 36, 1738–1750. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Update on natural food pigments—A mini-review on carotenoids, anthocyanins, and betalains. Food Res. Int. 2019, 124, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Silva Miranda, P.H.; Dos Santos, A.C.; De Freitas, B.C.B.; De Souza Martins, G.A.; De Barros Vilas Boas, E.V.; Damiani, C. A scientific approach to extraction methods and stability of pigments from Amazonian fruits. Trends Food Sci. Technol. 2021, 113, 335–345. [Google Scholar] [CrossRef]
- Macías-Sánchez, M.D.; Mantell, C.; Rodríguez, M.; Martínez de la Ossa, E.; Lubián, L.M.; Montero, O. Comparison of supercritical fluid and ultrasound-assisted extraction of carotenoids and chlorophyll a from Dunaliella salina. Talanta 2009, 77, 948–952. [Google Scholar] [CrossRef]
- Pereira, C.G.; Meireles, M.A.A. Supercritical Fluid Extraction of Bioactive Compounds: Fundamentals, Applications and Economic Perspectives. Food Bioprocess. Technol. 2010, 3, 340–372. [Google Scholar] [CrossRef]
- Pasquet, V.; Chérouvrier, J.; Farhat, F.; Thiéry, V.; Piot, J.; Bérard, J.; Kaas, R.; Serive, B.; Patrice, T.; Cadoret, J.; et al. Study on the microalgal pigments extraction process: Performance of microwave assisted extraction. Process. Biochem. 2011, 46, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, B.; Christen, P. Recent extraction techniques for natural products: Microwave-assisted extraction and pressurised solvent extraction. Phytochem. Anal. 2002, 13, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Srivastav, S.; Sharanagat, V.S. Ultrasound assisted extraction (UAE) of bioactive compounds from fruit and vegetable processing by-products: A review. Ultrason Sonochem. 2021, 70, 105325. [Google Scholar] [CrossRef]
- Martínez, J.M.; Schottroff, F.; Haas, K.; Fauster, T.; Sajfrtová, M.; Álvarez, I.; Raso, J.; Jaeger, H. Evaluation of pulsed electric fields technology for the improvement of subsequent carotenoid extraction from dried Rhodotorula glutinis yeast. Food Chem. 2020, 323, 126824. [Google Scholar] [CrossRef]
- Leonhardt, L.; Käferböck, A.; Smetana, S.; de Vos, R.; Toepfl, S.; Parniakov, O. Bio-refinery of Chlorella sorokiniana with pulsed electric field pre-treatment. Bioresour. Technol. 2020, 301, 122743. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; De Souza Mesquita, L.M.; Vaz, B.M.C.; Dias, A.C.R.V.; Torres-Acosta, M.A.; Quéguineur, B.; Coutinho, J.A.P.; Ventura, S.P.M. Extraction and fractionation of pigments from Saccharina latissima (Linnaeus, 2006) using an ionic liquid + oil + water System. ACS Sustain. Chem. Eng. 2021, 9, 6599–6612. [Google Scholar] [CrossRef]
- Krishnan, P.; Kruger, N.J.; Ratcliffe, R.G. Metabolite fingerprinting and profiling in plants using NMR. J. Exp. Bot. 2005, 56, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Furihata, K.; Koda, M.; Hu, F.; Kato, R.; Miyakawa, T.; Tanokura, M. 13C NMR-based metabolomics for the classification of green coffee beans according to variety and origin. J. Agric. Food Chem. 2012, 60, 10118–10125. [Google Scholar] [CrossRef]
- Ward, J.L.; Baker, J.M.; Miller, S.J.; Deborde, C.; Maucourt, M.; Biais, B.; Rolin, D.; Moing, A.; Moco, S.; Vervoort, J.; et al. An inter-laboratory comparison demonstrates that [H]-NMR metabolite fingerprinting is a robust technique for collaborative plant metabolomic data collection. Metabolomics 2010, 6, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Florentino-Ramos, E.; Villa-Ruano, N.; Hidalgo-Martínez, D.; Ramírez-Meraz, M.; Méndez-Aguilar, R.; Velásquez-Valle, R.; Zepeda-Vallejo, L.G.; Pérez-Hernández, N.; Becerra-Martínez, E. 1H NMR-based fingerprinting of eleven Mexican Capsicum annuum cultivars. Food Res. Int. 2019, 121, 12–19. [Google Scholar] [CrossRef]
- Djukovic, D.; Liu, S.; Henry, I.; Tobias, B.; Raftery, D. Signal enhancement in HPLC/microcoil NMR using automated column trapping. Anal. Chem. 2006, 78, 7154–7160. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.W.; Kim, C.Y.; Song, D.G.; Pan, C.H.; Cha, K.H.; Lee, D.U.; Um, B.H. Rapid identification of furanocoumarins in Angelica dahurica using the online LC-MMR-MS and their nitric oxide inhibitory activity in RAW 264.7 cells. Phytochem. Anal. 2010, 21, 322–327. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry-based metabolite profiling in plants. Nat. Protoc. 2006, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite profiling: From diagnostics to systems biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef]
- Vorst, O.; de Vos, C.H.R.; Lommen, A.; Staps, R.V.; Visser, R.G.F.; Bino, R.J.; Hall, R.D. A non-directed approach to the differential analysis of multiple LC-MS-derived metabolic profiles. Metabolomics 2005, 1, 169–180. [Google Scholar] [CrossRef]
- Nagashima, Y.; He, K.; Singh, J.; Metrani, R.; Crosby, K.M.; Jifon, J.; Jayaprakasha, G.K.; Patil, B.; Qian, X.; Koiwa, H. Transition of aromatic volatile and transcriptome profiles during melon fruit ripening. Plant Sci. 2021, 304, 110809. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.J.; Hyeon, H.; Park, N.I.; Yi, T.G.; Lim, S.H.; Park, S.Y.; Ha, S.H.; Kim, J.K. A high-throughput platform for interpretation of metabolite profile data from pepper (Capsicum) fruits of 13 phenotypes associated with different fruit maturity states. Food Chem. 2020, 331, 127286. [Google Scholar] [CrossRef] [PubMed]
- Rambla, J.L.; Granell, A. Determination of plant volatile apocarotenoids. Methods Mol. Biol. 2020, 2083, 165–175. [Google Scholar]
- Wu, H.; Huang, W.; Chen, Z.; Chen, Z.; Shi, J.; Kong, Q.; Sun, S.; Jiang, X.; Chen, D.; Yan, S. GC-MS-based metabolomic study reveals dynamic changes of chemical compositions during black tea processing. Food Res. Int. 2019, 120, 330–338. [Google Scholar] [CrossRef]
- Park, Y.J.; Baek, S.-A.; Choi, Y.; Kim, J.K.; Park, S.-U. Metabolic profiling of nine Mentha species and prediction of their antioxidant properties using chemometrics. Molecules 2019, 24, 258. [Google Scholar] [CrossRef] [Green Version]
- Rydberg, J.; Cooke, C.A.; Tolu, J.; Wolfe, A.P.; Vinebrooke, R.D. An assessment of chlorophyll preservation in lake sediments using multiple analytical techniques applied to the annually laminated lake sediments of Nylandssjön. J. Paleolimnol. 2020, 64, 379–388. [Google Scholar] [CrossRef]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; de Groot, J.; van Beek, T.A.; Vervoort, J.; de Vos, C.H. A liquid chromatography-mass spectrometry-based metabolome database for tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glauser, G.; Veyrat, N.; Rochat, B.; Wolfender, J.L.; Turlings, T.C. Ultra-high pressure liquid chromatography-mass spectrometry for plant metabolomics: A systematic comparison of high-resolution quadrupole-time-of-flight and single stage Orbitrap mass spectrometers. J. Chromatogr. A 2013, 1292, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Reig, M.; Jaumot, J.; Baglai, A.; Vivó-Truyols, G.; Schoenmakers, P.J.; Tauler, R. Untargeted comprehensive two-dimensional liquid chromatography coupled with high-resolution mass spectrometry analysis of rice metabolome using multivariate curve resolution. Anal. Chem. 2017, 89, 7675–7683. [Google Scholar] [CrossRef]
- Oliver, J.; Palou, A. Chromatographic determination of carotenoids in foods. J. Chromatogr. A 2000, 881, 543–555. [Google Scholar] [CrossRef]
- Delpino-Rius, A.; Cosovanu, D.; Eras, J.; Vilaró, F.; Balcells, M.; Canela-Garayoa, R. A fast and reliable ultrahigh-performance liquid chromatography method to assess the fate of chlorophylls in teas and processed vegetable foodstuff. J. Chromatogr A 2018, 1568, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zapata, M.; Rodriguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C-8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Rowley, K.G.; Balazs, N.D. Carotenoids: Separation methods applicable to biological samples. J. Chromatogr. B 2002, 781, 393–418. [Google Scholar] [CrossRef]
- Unlu, N.Z.; Bohn, T.; Francis, D.; Clinton, S.K.; Schwartz, S.J. Carotenoid absorption in humans consuming tomato sauces obtained from tangerine or high-beta-carotene varieties of tomatoes. J. Agric. Food Chem. 2007, 55, 1597–1603. [Google Scholar] [CrossRef]
- Lee, B.L.; Ong, C.N. Comprehensive high-performance liquid chromatographic method for the measurements of lipophilic antioxidants in human plasma. J. Chromatogr. A 2009, 1216, 3131–3137. [Google Scholar] [CrossRef]
- Li, S.; Nugroho, A.; Rocheford, T.; White, W.S. Vitamin A equivalence of the ß-carotene in ß-carotene-biofortified maize porridge consumed by women. Am. J. Clin. Nutr. 2010, 92, 1105–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puspitasari-Nienaber, N.L.; Ferruzzi, M.G.; Schwartz, S.J. Simultaneous detection of tocopherols, carotenoids, and chlorophylls in vegetable oils by direct injection C30 RP-HPLC with coulometric electrochemical array detection. J. Am. Oil Chem. Soc. 2002, 79, 633–640. [Google Scholar] [CrossRef]
- Bustamam, M.S.A.; Pantami, H.A.; Azizan, A.; Shaari, K.; Min, C.C.; Abas, F.; Nagao, N.; Maulidiani, M.; Banerjee, S.; Sulaiman, F.; et al. Complementary analytical platforms of NMR spectroscopy and LCMS analysis in the metabolite profiling of Isochrysis galbana. Mar. Drugs 2021, 19, 139. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, P.; de Vos, R.C.H.; Jonker, H.H.; Mumm, R.; Hall, R.D.; Bialek, L.; Leenman, R.; Strassburg, K.; Vreeken, R.; Hankemeier, T.; et al. Comprehensive metabolomics to evaluate the impact of industrial processing on the phytochemical composition of vegetable purees. Food Chem. 2015, 168, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Viera, I.; Roca, M.; Pérez-Gálvez, A. Mass spectrometry of non-allomerized chlorophylls a and b derivatives from plants. Curr. Org. Chem. 2018, 22, 842–876. [Google Scholar] [CrossRef] [Green Version]
- Lei, Z.; Sumner, B.W.; Bhatia, A.; Sarma, S.J.; Sumner, L.W. UHPLC-MS analyses of plant flavonoids. Curr. Protoc. Plant Biol. 2019, 4, e20085. [Google Scholar] [CrossRef] [Green Version]
- Breithaupt, D.E.; Wirt, U.; Bamedi, A. Differentiation between lutein monoester regioisomers and detection of lutein diesters from marigold flowers (Tagetes erecta L.) and several fruits by liquid chromatography-mass spectrometry. J. Agric. Food Chem. 2002, 50, 66–70. [Google Scholar] [CrossRef]
- Giuffrida, D.; Pintea, A.; Dugo, P.; Torre, G.; Pop, R.M.; Mondello, L. Determination of carotenoids and their esters in fruits of sea buckthorn (Hippophae rhamnoides L.) by HPLC-DAD-APCI-MS. Phytochem. Anal. 2012, 23, 267–273. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Mariutti, L.R.B.; Mercadante, A.Z. Two-step cleanup procedure for the identification of carotenoid esters by liquid chromatography-atmospheric pressure chemical ionization-tandem mass spectrometry. J. Chromatogr. A 2016, 1457, 116–124. [Google Scholar] [CrossRef]
- Chen, K.; Ríos, J.J.; Pérez-Gálvez, A.; Roca, M. Development of an accurate and high-throughput methodology for structural comprehension of chlorophylls derivatives. (I) Phytylated derivatives. J. Chromatogr. A 2015, 1406, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.S.; Nascimento, T.C.; Pinheiro, P.N.; De Rosso, V.V.; De Menezes, C.R.; Jacob-Lopes, E.; Zepka, L.Q. Insights on the intestinal absorption of chlorophyll series from microalgae. Food Res. Int. 2021, 140, 110031. [Google Scholar] [CrossRef] [PubMed]
- Murador, D.C.; De Souza Mesquita, L.M.; Neves, B.V.; Braga, A.R.C.; Martins, P.L.G.; Zepka, L.Q.; De Rosso, V.V. Bioaccessibility and cellular uptake by Caco-2 cells of carotenoids and chlorophylls from orange peels: A comparison between conventional and ionic liquid mediated extractions. Food Chem. 2021, 339, 127818. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Ríos, J.J.; Roca, M.; Pérez-Gálvez, A. Development of an accurate and high-throughput methodology for structural comprehension of chlorophylls derivatives. (II) Dephytylated derivatives. J. Chromatogr. A 2015, 1412, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Roca, M.; Ríos, J.J.; Pérez-Gálvez, A. Mass spectrometry: The indispensable tool for plant metabolomics of colourless chlorophyll catabolites. Phytochem. Rev. 2018, 17, 453–468. [Google Scholar] [CrossRef] [Green Version]
- Pacini, T.; Fu, W.; Gudmundsson, S.; Chiaravalle, A.E.; Brynjolfson, S.; Palsson, B.O.; Astarita, G.; Paglia, G. Multidimensional analytical approach based on UHPLC-UV-ion mobility-MS for the screening of natural pigments. Anal. Chem. 2015, 87, 2593–2599. [Google Scholar] [CrossRef]
- Fraser, P.D.; Enfisse, E.M.A.; Goodfellow, M.; Eguchi, T.; Bramley, P.M. Metabolite profiling of plant carotenoids using the matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Plant J. 2007, 49, 552–564. [Google Scholar] [CrossRef]
- Calvano, C.D.; Ventura, G.; Cataldi, T.R.; Palmisano, F. Improvement of chlorophyll identification in foodstuffs by MALDI ToF/ToF mass spectrometry using 1,5-diaminonaphthalene electron transfer secondary reaction matrix. Anal. Bioanal. Chem. 2015, 407, 6369–6379. [Google Scholar] [CrossRef]
- Maroneze, M.M.; Caballero-Guerrero, B.; Zepka, L.Q.; Jacob-Lopes, E.; Pérez-Gálvez, A.; Roca, M. Accomplished high-resolution metabolomic and molecular studies identify new carotenoid biosynthetic reactions in Cyanobacteria. J. Agric. Food Chem. 2020, 68, 6212–6220. [Google Scholar] [CrossRef]
- Hegeman, A.D.; Schulte, C.F.; Cui, Q.; Lewis, I.A.; Huttlin, E.L.; Eghbalnia, H.; Harms, A.C.; Ulrich, E.L.; Markley, J.L.; Sussman, M.R. Stable isotope assisted assignment of elemental compositions for metabolomics. Anal. Chem. 2007, 79, 6912–6921. [Google Scholar] [CrossRef]
- Giavalisco, P.; Li, Y.; Matthes, A.; Eckhardt, A.; Hubberten, H.-M.; Hesse, H.; Segu, S.; Hummel, J.; Köhl, K.; Willmitzer, L. Elemental formula annotation of polar and lipophilic metabolites using 13 C, 15 N and 34 S isotope labelling, in combination with high-resolution mass spectrometry: Isotope labelling for unbiased plant metabolomics. Plant J. 2011, 68, 364–376. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Development of an accurate and direct method for the green food colorants detection. Food Res. Int. 2020, 136, 109484. [Google Scholar] [CrossRef]
- Ríos, J.J.; Roca, M.; Pérez-Gálvez, A. Systematic HPLC/ESI-high resolution-qTOF-MS methodology for metabolomic studies in nonfluorescent chlorophyll catabolites pathway. J. Anal. Methods Chem. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Wehrens, R.; Carvalho, E.; Masuero, D.; de Juan, A.; Martens, S. High-throughput carotenoid profiling using multivariate curve resolution. Anal. Bioanal. Chem. 2013, 405, 5075–5086. [Google Scholar] [CrossRef]
- Wehrens, R.; Bloemberg, T.G.; Eilers, P.H.C. Fast parametric time warping of peak lists. Bioinformatics 2015, 31, 3063–3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Tohge, T.; Balazadeh, S.; Erban, A.; Giavalisco, P.; Kopka, J.; Mueller-Roeber, B.; Fernie, A.R.; Hoefgen, R. Comprehensive metabolomics studies of plant developmental senescence. In Plant Senescence; Guo, Y., Ed.; Springer: New York, NY, USA, 2018; Volume 1744, pp. 339–358. [Google Scholar]
- Herzog, M.; Fukao, T.; Winkel, A.; Konnerup, D.; Lamichhane, S.; Alpuerto, J.B.; Hasler-Sheetal, H.; Pedersen, O. Physiology, gene expression, and metabolome of two wheat cultivars with contrasting submergence tolerance: Submergence tolerance in two wheat cultivars. Plant Cell Environ. 2018, 41, 1632–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pino Del Carpio, D.; Basnet, R.K.; De Vos, R.C.H.; Maliepaard, C.; Paulo, M.J.; Bonnema, G. Comparative methods for association studies: A case study on metabolite variation in a Brassica rapa core collection. PLoS ONE 2011, 6, e19624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamba, T.; Lee, J.W.; Matsubara, A.; Fukusaki, E. Metabolic profiling of lipids by supercritical fluid chromatography/mass spectrometry. J. Chromatogr. A 2012, 1250, 212–219. [Google Scholar] [CrossRef]
- Giuffrida, D.; Zoccali, M.; Arigò, A.; Cacciola, F.; Roa, C.O.; Dugo, P.; Mondello, L. Comparison of different analytical techniques for the analysis of carotenoids in tamarillo (Solanum betaceum Cav.). Arch. Biochem. Biophys. 2018, 646, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Cichon, M.J.; Riedl, K.M.; Schwartz, S.J. A metabolomic evaluation of the phytochemical composition of tomato juices being used in human clinical trials. Food Chem. 2017, 228, 270–278. [Google Scholar] [CrossRef]
- Fukushima, A.; Kusano, M.; Nakamichi, N.; Kobayashi, M.; Hayashi, N.; Sakakibara, H.; Mizuno, T.; Saito, K. Impact of clock-associated Arabidopsis pseudo-response regulators in metabolic coordination. Proc. Natl. Acad. Sci. USA 2009, 106, 7251–7256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moco, S.; Capanoglu, E.; Tikunov, Y.; Bino, R.J.; Boyacioglu, D.; Hall, R.D.; Vervoort, J.; De Vos, R.C.H. Tissue specialization at the metabolite level is perceived during the development of tomato fruit. J. Exp. Bot. 2007, 58, 4131–4146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witt, S.; Galicia, L.; Lisec, J.; Cairns, J.; Tiessen, A.; Araus, J.L.; Palacios-Rojas, N.; Fernie, A.R. Metabolic and phenotypic responses of greenhouse-grown maize hybrids to experimentally controlled drought stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bemer, M.; Karlova, R.; Ballester, A.R.; Tikunov, Y.M.; Bovy, A.G.; Wolters-Arts, M.; de Barros Rossetto, P.; Angenent, G.C.; de Maagd, R.A. The tomato FRUITFULL homologs TDR4/FUL1 and MBP7/FUL2 regulate ethylene-independent aspects of fruit ripening. Plant Cell 2012, 24, 4437–4451. [Google Scholar] [CrossRef] [Green Version]
- Harel-Beja, R.; Tzuri, G.; Portnoy, V.; Lotan-Pompan, M.; Lev, S.; Cohen, S.; Dai, N.; Yeselson, L.; Meir, A.; Libhaber, S.E.; et al. A genetic map of melon highly enriched with fruit quality QTLs and EST markers, including sugar and carotenoid metabolism genes. Theor. Appl. Genet. 2010, 121, 511–533. [Google Scholar] [CrossRef]
- Pandey, S.; Patel, M.K.; Mishra, A.; Jha, B. Physio-Biochemical composition and untargeted metabolomics of cumin (Cuminum cyminum L.) make it promising functional food and help in mitigating salinity stress. PLoS ONE 2015, 10, e014446. [Google Scholar] [CrossRef] [Green Version]
- Acharjee, A.; Kloosterman, B.; de Vos, R.C.H.; Werij, J.S.; Bachem, C.W.B.; Visser, R.G.F.; Maliepaard, C. Data integration and network reconstruction with omics data using Random Forest regression in potato. Anal. Chim. Acta 2011, 705, 56–63. [Google Scholar] [CrossRef]
- Le Lay, P.; Isaure, M.P.; Sarry, J.E.; Kuhn, L.; Fayard, B.; Le Bail, J.L.; Bastien, O.; Garin, J.; Roby, C.; Bourguignon, J. Metabolomic, proteomic and biophysical analyses of Arabidopsis thaliana cells exposed to a caesium stress. Influence of potassium supply. Biochimie 2006, 88, 1533–1547. [Google Scholar] [CrossRef]
- León, J.; Costa, A.; Castillo, M.C. Nitric oxide triggers a transient metabolic reprogramming in Arabidopsis. Sci. Rep. 2016, 6, 37945. [Google Scholar] [CrossRef]
- Mwamba, T.M.; Islam, F.; Ali, B.; Lwalaba, J.L.W.; Gill, R.A.; Zhang, F.; Farooq, M.A.; Ali, S.; Ulhassan, Z.; Huang, Q.; et al. Comparative metabolomic responses of low- and high-cadmium accumulating genotypes reveal the cadmium adaptive mechanism in Brassica napus. Chemosphere 2020, 250, 126308. [Google Scholar] [CrossRef]
- Hédiji, H.; Djebali, W.; Cabasson, C.; Maucourt, M.; Baldet, P.; Bertrand, A.; Boulila Zoghlami, L.; Deborde, C.; Moing, A.; Brouquisse, R.; et al. Effects of long-term cadmium exposure on growth and metabolomic profile of tomato plants. Ecotox. Environ. Safety 2010, 73, 1965–1974. [Google Scholar] [CrossRef]
- Hu, X.; Gao, Y.; Fang, Z. Integrating metabolic analysis with biological endpoints provides insight into nanotoxicological mechanisms of graphene oxide: From effect onset to cessation. Carbon 2016, 109, 65–73. [Google Scholar] [CrossRef]
- Allen, A.E.; LaRoche, J.; Maheswari, U.; Lommer, M.; Schauer, N.; Lopez, P.J.; Finazzi, G.; Fernie, A.R.; Bowler, C. Whole-cell response of the pennate diatom Phaeodactylum tricornutum to iron starvation. Proc. Natl. Acad. Sci. USA 2008, 105, 10438–10443. [Google Scholar] [CrossRef] [Green Version]
- Matich, E.K.; Ghafari, M.; Camgoz, E.; Caliskan, E.; Pfeifer, B.A.; Haznedaroglu, B.Z.; Atilla-Gokcumen, G.E. Time-series lipidomic analysis of the oleaginous green microalga species Ettlia oleoabundans under nutrient stress. Biotechnol. Biofuels 2018, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipanah, L.; Rohloff, J.; Winge, P.; Bones, A.M.; Brembu, T. Whole-cell response to nitrogen deprivation in the diatom Phaeodactylum tricornutum. EXBOTJ 2015, 66, 6281–6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.Y.; Park, J.-J.; Barupal, D.K.; Fiehn, O. System sesponse of metabolic networks in Chlamydomonas reinhardtii to total available ammonium. Mol. Cell. Proteom. 2012, 11, 973–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, H.; Meng, N.; Fu, J.; Chen, X.; Xu, X.; Feng, Q.; Jiang, H.; Dai, J.; Yuan, X.; Lu, Y.; et al. Genome-wide transcriptome and antioxidant analyses on gamma-irradiated phases of Deinococcus radiodurans R1. PLoS ONE 2014, 9, e85649. [Google Scholar] [CrossRef] [PubMed]
- Hansler, A.; Chen, Q.; Ma, Y.; Gross, S.S. Untargeted metabolite profiling reveals that nitric oxide bioynthesis is an endogenous modulator of carotenoid biosynthesis in Deinococcus radiodurans and is required for extreme ionizing radiation resistance. Arch. Biochem. Biophys. 2016, 589, 38–52. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Huang, X.; Sun, W.; Too, H.Z.; Laserna, A.K.C.; Li, S.F.Y. A global metabolomic insight into the oxidative stress and membrane damage of copper oxide nanoparticles and microparticles on microalga Chlorella vulgaris. Environ. Pollut. 2020, 258, 113647. [Google Scholar] [CrossRef]
- Reddy Pullagurala, V.L.; Adisa, I.O.; Rawat, S.; Kalagara, S.; Hernandez-Viezcas, J.A.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. ZnO nanoparticles increase photosynthetic pigments and decrease lipid peroxidation in soil grown cilantro (Coriandrum sativum). Plant Physiol. Biochem. 2018, 132, 120–127. [Google Scholar] [CrossRef]
- Maldini, M.; Natella, F.; Baima, S.; Morelli, G.; Scaccini, C.; Langridge, J.; Astarita, G. Untargeted metabolomics reveals predominant alterations in lipid metabolism following light exposure in broccoli sprouts. Int. J. Mol. Sci. 2015, 16, 13678–13691. [Google Scholar] [CrossRef] [Green Version]
- Will, S.E.; Henke, P.; Boedeker, C. Day and night: Metabolic profiles and evolutionary relationships of six axenic non-marine cyanobacteria. Genome Biol. Evol. 2019, 11, 270–294. [Google Scholar] [CrossRef]
- Gong, G.; Liu, L.; Zhang, X. Multi-omics metabolism analysis on irradiation-induced oxidative stress to Rhodotorula glutinis. Appl. Microbiol. Biotechnol. 2019, 103, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, C.A.; Airs, R.L.; Farnham, G.; Greig, C. Synthesis, regulation and degradation of carotenoids under low level UV-B radiation in the filamentous cyanobacterium Chlorogloeopsis fritschii PCC 6912. Front. Microbiol. 2020, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntagkas, N.; de Vos, R.C.H.; Woltering, E.J.; Nicole, C.C.S.; Labrie, C.; Marcelis, L.F.M. Modulation of the tomato fruit metabolome by LED light. Metabolites 2020, 10, 266. [Google Scholar] [CrossRef]
- Yan, Z.; Zuo, J.; Zhou, F.; Shi, J.; Xu, D.; Hu, W.; Jiang, A.; Liu, Y.; Wang, Q. Integrated analysis of transcriptomic and metabolomic data reveals the mechanism by which LED light irradiation extends the postharvest quality of Pak-choi (Brassica campestris L. ssp. chinensis (L.) Makino var. communis Tsen et Lee). Biomolecules 2020, 10, 252. [Google Scholar]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 2016, 16, 67. [Google Scholar] [CrossRef] [Green Version]
- Uarrota, V.G.; Segatto, C.; Voytena, A.P.L.; Coelho, C.M.M.; Souza, C.A. Metabolic fingerprinting of water-stressed soybean cultivars by gas chromatography, near-infrared and UV-visible spectroscopy combined with chemometrics. J. Agron. Crop Sci. 2019, 205, 141–156. [Google Scholar] [CrossRef]
- Lucini, L.; Miras-Moreno, B.; Busconi, M.; Marocco, A.; Gatti, M.; Poni, S. Molecular basis of rootstock-related tolerance to water deficit in Vitis vinifera L. cv. Sangiovese: A physiological and metabolomic combined approach. Plant Sci. 2020, 299, 110600. [Google Scholar] [PubMed]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. A combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef]
- Li, Z.; Cheng, B.; Yong, B.; Liu, T.; Peng, Y.; Zhang, X.; Ma, X.; Huang, L.; Liu, W.; Nie, G. Metabolomics and physiological analyses reveal β-sitosterol as an important plant growth regulator inducing tolerance to water stress in white clover. Planta 2019, 250, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Marín, B.; Nadal, M.; Gago, J.; Fernie, A.R.; López-Pozo, M.; Artetxe, U.; García-Plazaola, J.I.; Verhoeven, A. Born to revive: Molecular and physiological mechanisms of double tolerance in a paleotropical and resurrection plant. New Phytol. 2020, 226, 741–759. [Google Scholar] [CrossRef]
- Rouphael, Y.; Raimondi, G.; Lucini, L.; Carillo, P.; Kyriacou, M.C.; Colla, G.; Cirillo, V.; Pannico, A.; El-Nakhel, C.; De Pascale, S. Physiological and metabolic responses triggered by omeprazole improve tomato plant tolerance to NaCl stress. Front. Plant Sci. 2018, 9, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, D.; Zhou, R.; Wang, X.; Dossa, K.; Wang, L.; Zhang, Y.; Yu, J.; Gong, H.; Zhang, X.; et al. Transcriptome and metabolome analyses of two contrasting sesame genotypes reveal the crucial biological pathways involved in rapid adaptive response to salt stress. BMC Plant Biol. 2019, 19, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, X.; Zhu, Y.; Hu, Y.; Zhang, R.; Cheng, L.; Zhu, Z.; Zhao, T.; Zhang, X.; Wang, Y. Integrated physiologic, proteomic, and metabolomic analyses of Malus halliana adaptation to saline-alkali stress. Hortic. Res. 2019, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.L.; Pirastru, L.; Popovic, R.; Sleno, L. Carotenogenesis up-regulation in Scenedesmus sp. using a targeted metabolomics approach by liquid chromatography−high-resolution mass spectrometry. J. Agric. Food Chem. 2011, 59, 3004–3013. [Google Scholar] [CrossRef]
- Van Meulebroek, L.; Vanden Bussche, J.; Steppe, K.; Vanhaecke, L. High-resolution Orbitrap mass spectrometry for the analysis of carotenoids in tomato fruit: Validation and comparative evaluation towards UV-VIS and tandem mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 2613–2626. [Google Scholar] [CrossRef]
- Mandelli, F.; Couger, M.B.; Paixão, D.A.A.; Machado, C.B.; Carnielli, C.M.; Aricetti, J.A.; Polikarpov, I.; Prade, R.; Caldana, C.; Paes Leme, A.F.; et al. Thermal adaptation strategies of the extremophile bacterium Thermus filiformis based on multi-omics analysis. Extremophiles 2017, 21, 775–788. [Google Scholar] [CrossRef]
- Almeida, J.; Perez-Fons, L.; Fraser, P.D. A transcriptomic, metabolomic and cellular approach to the physiological adaptation of tomato fruit to high temperature. Plant Cell Environ. 2020, 2020, 1–19. [Google Scholar] [CrossRef]
- Fogelman, E.; Oren-Shamir, M.; Hirschberg, J.; Mandolino, G.; Parisi, B.; Ovadia, R.; Tanami, Z.; Faigenboim, A.; Ginzberg, I. Nutritional value of potato (Solanum tuberosum) in hot climates: Anthocyanins, carotenoids, and steroidal glycoalkaloids. Planta 2019, 249, 1143–1155. [Google Scholar] [CrossRef]
- Chou, L.; Kenig, F.; Murray, A.E.; Fritsen, C.H.; Doran, P.T. Effects of legacy metabolites from previous ecosystems on the environmental metabolomics of the brine of Lake Vida, East Antarctica. Org. Geochem. 2018, 122, 161–170. [Google Scholar] [CrossRef]
- Wang, L.; Huang, X.; Lim, D.J.; Laserna, A.K.C.; Li, S.F.Y. Uptake and toxic effects of triphenyl phosphate on freshwater microalgae Chlorella vulgaris and Scenedesmus obliquus: Insights from untargeted metabolomics. Sci. Total Environ. 2019, 650, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Stopka, S.A.; Mansour, T.R.; Shrestha, B.; Maréchal, É.; Falconet, D.; Vertes, A. Turnover rates in microorganisms by laser ablation electrospray ionization mass spectrometry and pulse-chase analysis. Anal. Chim. Acta 2016, 902, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Brown, P.; Ponchet, M. (Eds.) Proceedings of the 1st World Congress on the Use of Biostimulants in Agriculture; Acta Horticulturae Number 1009; ISHS: Leuven, Belgium, 2013; pp. 1–251. [Google Scholar]
- Barrajón-Catalán, E.; Álvarez-Martínez, F.J.; Borrás, F.; Pérez, D.; Herrero, N.; Ruiz, J.J.; Micol, V. Metabolomic analysis of the effects of a commercial complex biostimulant on pepper crops. Food Chem. 2020, 310, 125818. [Google Scholar] [CrossRef]
- Sofo, A.; Fausto, C.; Mininni, A.N.; Dichio, B.; Lucini, L. Soil management type differentially modulates the metabolomic profile of olive xylem sap. Plant Physiol. Biochem. 2019, 139, 707–714. [Google Scholar] [CrossRef]
- Errard, A.; Ulrichs, C.; Kühne, S.; Mewis, I.; Drungowski, M.; Schreiner, M.; Baldermann, S. Single- versus multiple-pest infestation affects differently the biochemistry of tomato (Solanum lycopersicum ‘Ailsa Craig’). J. Agric. Food Chem. 2015, 63, 10103–10111. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Chamoun, R.; Jabaji, S. Metabolic responses of willow (Salix purpurea L.) leaves to mycorrhization as revealed by mass spectrometry and 1H NMR spectroscopy metabolite profiling. Front. Plant Sci. 2015, 6, 344. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Zeng, Y.; Edlund, A.; Melnik, A.V.; Sanchez, L.M.; Mohimani, H.; Gurevich, A.; Miao, V.; Schiffler, S.; Lim, Y.W.; et al. Spatial molecular architecture of the microbial community of a Peltigera lichen. mSystems 2016, 1, e00139-16. [Google Scholar] [CrossRef] [Green Version]
- Paix, B.; Carriot, N.; Barry-Martinet, R. A multi-omics analysis suggests links between the differentiated surface metabolome and epiphytic microbiota along the thallus of a mediterranean seaweed holobiont. Front. Microbiol. 2020, 11, 494. [Google Scholar] [CrossRef]
- Sørensen, M.E.S.; Wood, A.J.; Minter, E.J.A.; Lowe, C.D.; Cameron, D.D.; Brockhurst, M.A. Comparison of independent evolutionary origins reveals both convergence and divergence in the metabolic mechanisms of symbiosis. Curr. Biol. 2020, 30, 328–334.e4. [Google Scholar] [CrossRef]
- Cheng, Q.; Jia, W.; Hu, C.; Shi, G.; Yang, D.; Cai, M.; Zhan, T.; Tang, Y.; Zhou, Y.; Sun, X.; et al. Enhancement and improvement of selenium in soil to the resistance of rape stem against Sclerotinia sclerotiorum and the inhibition of dissolved organic matter derived from rape straw on mycelium. Environ. Pollut. 2020, 265, 114827. [Google Scholar] [CrossRef]
- Tan, F.Q.; Zhang, M.; Xie, K.D.; Fan, Y.J.; Song, X.; Wang, R.; Wu, X.M.; Zhang, H.Y.; Guo, W.W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin (Citrus reticulata Blanco). Plant Sci. 2019, 289, 110276. [Google Scholar] [CrossRef]
- Maulidiani, M.; Mediani, A.; Abas, F.; Park, Y.S.; Park, Y.-K.; Kim, Y.M.; Gorinstein, S. 1H NMR and antioxidant profiles of polar and non-polar extracts of persimmon (Diospyros kaki L.)—Metabolomics study based on cultivars and origins. Talanta 2018, 184, 277–286. [Google Scholar] [CrossRef]
- Masetti, O.; Ciampa, A.; Nisini, L.; Sequi, P.; Dell’Abate, M.T. A multifactorial approach in characterizing geographical origin of Sicilian cherry tomatoes using 1 H-NMR profiling. Food Res. Int. 2017, 100, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Llano, S.M.; Muñoz-Jiménez, A.M.; Jiménez-Cartagena, C.; Londoño-Londoño, J.; Medina, S. Untargeted metabolomics reveals specific withanolides and fatty acyl glycoside as tentative metabolites to differentiate organic and conventional Physalis peruviana fruits. Food Chem. 2018, 244, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, J.; Zhao, Y.; Lu, X.; Zhou, Z.; Zhao, C.; Xu, G. Comprehensive investigation of tobacco leaves during natural early senescence via multi-platform metabolomics analyses. Sci. Rep. 2016, 6, 37976. [Google Scholar] [CrossRef] [PubMed]
- Heavisides, E.; Rouger, C.; Reichel, A.; Ulrich, C.; Wenzel-Storjohann, A.; Sebens, S.; Tasdemir, D. Seasonal variations in the metabolome and bioactivity profile of Fucus vesiculosus extracted by an optimised, pressurised liquid extraction protocol. Mar. Drugs 2018, 16, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masetti, O.; Ciampa, A.; Nisini, L.; Valentini, M.; Sequi, P.; Dell’Abate, M.T. Cherry tomatoes metabolic profile determined by 1H-High Resolution-NMR spectroscopy as influenced by growing season. Food Chem. 2014, 162, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Juin, C.; Bonnet, A.; Nicolau, E.; Bérard, J.-B.; Devillers, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. UPLC-MSE Profiling of phytoplankton metabolites: Application to the identification of pigments and structural analysis of metabolites in Porphyridium purpureum. Mar. Drugs 2015, 13, 2541–2558. [Google Scholar] [CrossRef] [Green Version]
- Marcellin-Gros, R.; Piganeau, G.; Stien, D. Metabolomic insights into marine phytoplankton diversity. Mar. Drugs 2020, 18, 78. [Google Scholar] [CrossRef] [Green Version]
- Drapal, M.; Carvalho, E.; Ovalle Rivera, T.M.; Becerra Lopez-Lavalle, L.A.; Fraser, P.D. Capturing biochemical diversity in Cassava (Manihot esculenta Crantz) through the application of metabolite profiling. J. Agric. Food Chem. 2019, 673, 986–993. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Rocchetti, G.; Miras Moreno, M.B.; Lucini, L.; Incrocci, L.; Pardossi, A.; Guidi, L. Hydroponically grown Sanguisorba minor Scop.: Effects of cut and storage on fresh-cut produce. Antioxidants 2019, 8, 631. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Luo, Z.; Ban, Z.; Lu, H.; Li, D.; Yang, D.; Aghdam, M.S.; Li, L. The effect of the layer-by-layer (LBL) edible coating on strawberry quality and metabolites during storage. Postharvest Biol. Technol. 2019, 147, 29–38. [Google Scholar] [CrossRef]
- Santin, M.; Lucini, L.; Castagna, A.; Chiodelli, G.; Hauser, M.-T.; Ranieri, A. Post-harvest UV-B radiation modulates metabolite profile in peach fruit. Postharvest Biol. Technol. 2018, 139, 127–134. [Google Scholar] [CrossRef]
- Zhou, F.; Zuo, J.; Gao, L. An untargeted metabolomic approach reveals significant postharvest alterations in vitamin metabolism in response to LED irradiation in pak-choi (Brassica campestris L. ssp. chinensis (L.) Makino var. communis Tsen et Lee. Metabolomics 2019, 15, 155. [Google Scholar] [CrossRef] [PubMed]
- Uarrota, V.G.; Moresco, R.; Coelho, B.; Nunes, E.d.C.; Peruch, L.A.M.; Neubert, E.d.O.; Rocha, M.; Maraschin, M. Metabolomics combined with chemometric tools (PCA, HCA, PLS-DA and SVM) for screening cassava (Manihot esculenta Crantz) roots during postharvest physiological deterioration. Food Chem. 2014, 161, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Luo, J.; Wang, S. The diversity of nutritional metabolites: Origin, dissection, and application in crop breeding. Front. Plant Sci. 2019, 10, 01028. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Sato, M.; Okamoto, M. Metabolome-based discrimination of chrysanthemum cultivars for the efficient generation of flower color variations in mutation breeding. Metabolomics 2019, 15, 118. [Google Scholar] [CrossRef]
- Hao, Z.; Liu, S.; Hu, L.; Shi, J.; Chen, J. Transcriptome analysis and metabolic profiling reveal the key role of carotenoids in the petal coloration of Liriodendron tulipifera. Hortic. Res. 2020, 7, 70. [Google Scholar] [CrossRef]
- Heng, Z.; Sheng, O.; Huang, W.; Zhang, S.; Fernie, A.R.; Motorykin, I.; Kong, Q.; Yi, G.; Yan, S. Integrated proteomic and metabolomic analysis suggests high rates of glycolysis are likely required to support high carotenoid accumulation in banana pulp. Food Chem. 2019, 297, 125016. [Google Scholar] [CrossRef]
- Enfissi, E.M.A.; Barneche, F.; Ahmed, I.; Lichtlé, C.; Gerrish, C.; McQuinn, R.P.; Giovannoni, J.J.; Lopez-Juez, E.; Bowler, C.; Bramley, P.M.; et al. Integrative transcript and metabolite analysis of nutritionally enhanced DE-ETIOLATED1 downregulated tomato fruit. Plant Cell. 2010, 22, 1190–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Gong, Z.; Li, N.; Zhao, Y.; Zhang, H.; Yang, X.; Liu, Y.; Rao, Z.; Yu, X. A negative regulator of carotenogenesis in Blakeslea trispora. Appl. Environ. Microbiol. 2020, 86, e02462-19. [Google Scholar] [CrossRef] [PubMed]
- Farré, G.; Maiam Rivera, S.; Alves, R.; Vilaprinyo, E.; Sorribas, A.; Canela, R.; Naqvi, S.; Sandmann, G.; Capell, T.; Zhu, C.; et al. Targeted transcriptomic and metabolic profiling reveals temporal bottlenecks in the maize carotenoid pathway that may be addressed by multigene engineering. Plant J. 2013, 75, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Alós, E.; Roca, M.; Iglesias, D.J.; Mínguez-Mosquera, M.I.; Damasceno, C.M.B.; Thannhauser, T.W.; Rose, J.K.C.; Talón, M.; Cercós, M. An evaluation of the basis and consequences of a stay-green mutation in the navel negra citrus mutant using transcriptomic and proteomic profiling and metabolite analysis. Plant Physiol. 2008, 147, 1300–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Fons, L.; Wells, T.; Corol, D.I.; Ward, J.L.; Gerrish, C.; Beale, M.H.; Seymour, G.B.; Bramley, P.M.; Fraser, P.D. A genome-wide metabolomic resource for tomato fruit from Solanum pennellii. Sci. Rep. 2015, 4, 3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.L.; Chen, L.; Shi, J.; Trzcinski, A.; Chen, W.-N. Metabolomic profiling of Rhodosporidium toruloides grown on glycerol for carotenoid production during different growth phases. J. Agric. Food Chem. 2014, 62, 10203–10209. [Google Scholar] [CrossRef]
- Farré, G.; Perez-Fons, L.; Decourcelle, M.; Breitenbach, J.; Hem, S.; Zhu, C.; Capell, T.; Christou, P.; Fraser, P.D.; Sandmann, G. Metabolic engineering of astaxanthin biosynthesis in maize endosperm and characterization of a prototype high oil hybrid. Transgenic Res. 2016, 25, 477–489. [Google Scholar] [CrossRef]
- Saiman, M.Z.; Miettinen, K.; Mustafa, N.R.; Choi, Y.H.; Verpoorte, R.; Schulte, A.E. Metabolic alteration of Catharanthus roseus cell suspension cultures overexpressing geraniol synthase in the plastids or cytosol. Plant Cell Tiss. Organ. Cult. 2018, 134, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Hasunuma, T.; Takaki, A.; Matsuda, M.; Kato, Y.; Vavricka, C.J.; Kondo, A. Single-stage astaxanthin production enhances the nonmevalonate pathway and photosynthetic central metabolism in Synechococcus sp. PCC 7002. Synth. Biol. 2019, 8, 2701–2709. [Google Scholar]
- Comas, J.; Benfeitas, R.; Vilaprinyo, E.; Sorribas, A.; Solsona, F.; Farré, G.; Berman, J.; Zorrilla, U.; Capell, T.; Sandmann, G.; et al. Identification of line-specific strategies for improving carotenoid production in synthetic maize through data-driven mathematical modeling. Plant J. 2016, 87, 455–471. [Google Scholar] [CrossRef] [Green Version]
- Maroneze, M.M.; Zepka, L.Q.; Jacob-Lopes, E.; Pérez-Gálvez, A.; Roca, M. Chlorophyll oxidative metabolism during the phototrophic and heterotrophic growth of Scenesdesmus obliquus. Antioxidants 2019, 8, 600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, P.M.; Batista, D.L.J.; Ribeiro, L.A.F.; Boffo, E.F.; de Cerqueira, M.D.; Martins, D.; de Castro, R.D.; de Souza-Neta, L.C.; Pinto, E.; Zambotti-Villela, L.; et al. Identification of antioxidant and antimicrobial compounds from the oilseed crop Ricinus communis using a multiplatform metabolite profiling approach. Ind. Crop. Prod. 2018, 124, 834–844. [Google Scholar] [CrossRef]
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; Hall, R.; de Vos, R. Changes in antioxidant and metabolite profiles during production of tomato paste. J. Agric. Food Chem. 2008, 56, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Beleggia, R.; Platani, C.; Papa, R.; Di Chio, A.; Barros, E.; Mashaba, C.; Wirth, J.; Fammartino, A.; Sautter, C.; Conner, S.; et al. Metabolomics and food processing: From semolina to pasta. J. Agric. Food Chem. 2011, 59, 9366–9377. [Google Scholar] [CrossRef]
- Van den Broek, T.J.; Kremer, B.H.; Rezende, M.M.; Hoevenaars, F.; Weber, P.; Hoeller, U.; van Ommen, B.; Wopereis, S. The impact of micronutrient status on health: Correlation network analysis to understand the role of micronutrients in metabolic-inflammatory processes regulating homeostasis and phenotypic flexibility. Genes Nutr. 2017, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Moser, S.; Erhart, T.; Neuhauser, S.; Kräutler, B. Phyllobilins from senescence-associated chlorophyll breakdown in the leaves of Basil (Ocimum basilicum) show increased abundance upon herbivore attack. J. Agric. Food Chem. 2020, 68, 7132–7142. [Google Scholar] [CrossRef]
- McQuinn, R.P.; Giovannoni, J.J.; Pogson, B.J. More than meets the eye: From carotenoid biosynthesis, to new insights into apocarotenoid signaling. Curr. Opin. Plant Biol. 2015, 27, 172–179. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Kohlen, W.; Charnikhova, T.; Mulder, P.; Undas, A.K.; Sergeant, M.J.; Verstappen, F.; Bugg, T.D.H.; Thompson, A.J.; Ruyter-Spira, C.; et al. Does abscisic acid affect strigolactone biosynthesis? New Phytol. 2010, 187, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Huttanus, H.M.; Vu, T.; Guruli, G.; Tracey, A.; Carswell, W.; Said, N.; Du, P.; Parkinson, B.G.; Orlando, G.; Robertson, J.L.; et al. Raman chemometric urinalysis (Rametrix) as a screen for bladder cancer. PLoS ONE 2020, 15, e0237070. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Parat, M.-O.; Shaw, P.N.; Hewavitharana, A.K.; Hodson, M.P. Traditional aboriginal preparation alters the chemical profile of Carica papaya leaves and impacts on cytotoxicity towards human squamous cell carcinoma. PLoS ONE 2016, 11, e0147956. [Google Scholar] [CrossRef] [Green Version]
- Arathi, B.P.; Sowmya, P.R.-R.; Kuriakose, G.C.; Vijay, K.; Baskaran, V.; Jayabaskaran, C.; Lakshminarayana, R. Enhanced cytotoxic and apoptosis inducing activity of lycopene oxidation products in different cancer cell lines. Food Chem. Toxicol. 2016, 97, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Rebholz, C.M.; Lichtenstein, A.H.; Zheng, Z.; Appel, L.J.; Coresh, J. Serum untargeted metabolomic profile of the dietary approaches to stop hypertension (DASH) dietary pattern. Am. J. Clin. Nutr. 2018, 108, 243–255. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roca, M.; Pérez-Gálvez, A. Metabolomics of Chlorophylls and Carotenoids: Analytical Methods and Metabolome-Based Studies. Antioxidants 2021, 10, 1622. https://doi.org/10.3390/antiox10101622
Roca M, Pérez-Gálvez A. Metabolomics of Chlorophylls and Carotenoids: Analytical Methods and Metabolome-Based Studies. Antioxidants. 2021; 10(10):1622. https://doi.org/10.3390/antiox10101622
Chicago/Turabian StyleRoca, María, and Antonio Pérez-Gálvez. 2021. "Metabolomics of Chlorophylls and Carotenoids: Analytical Methods and Metabolome-Based Studies" Antioxidants 10, no. 10: 1622. https://doi.org/10.3390/antiox10101622