
Potential of Persimmon Dietary Fiber Obtained from Byproducts as Antioxidant, Prebiotic and Modulating Agent of the Intestinal Epithelial Barrier Function

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material
2.3. Solvent Assisted Extraction (SAE)
2.4. Physicochemical Analysis
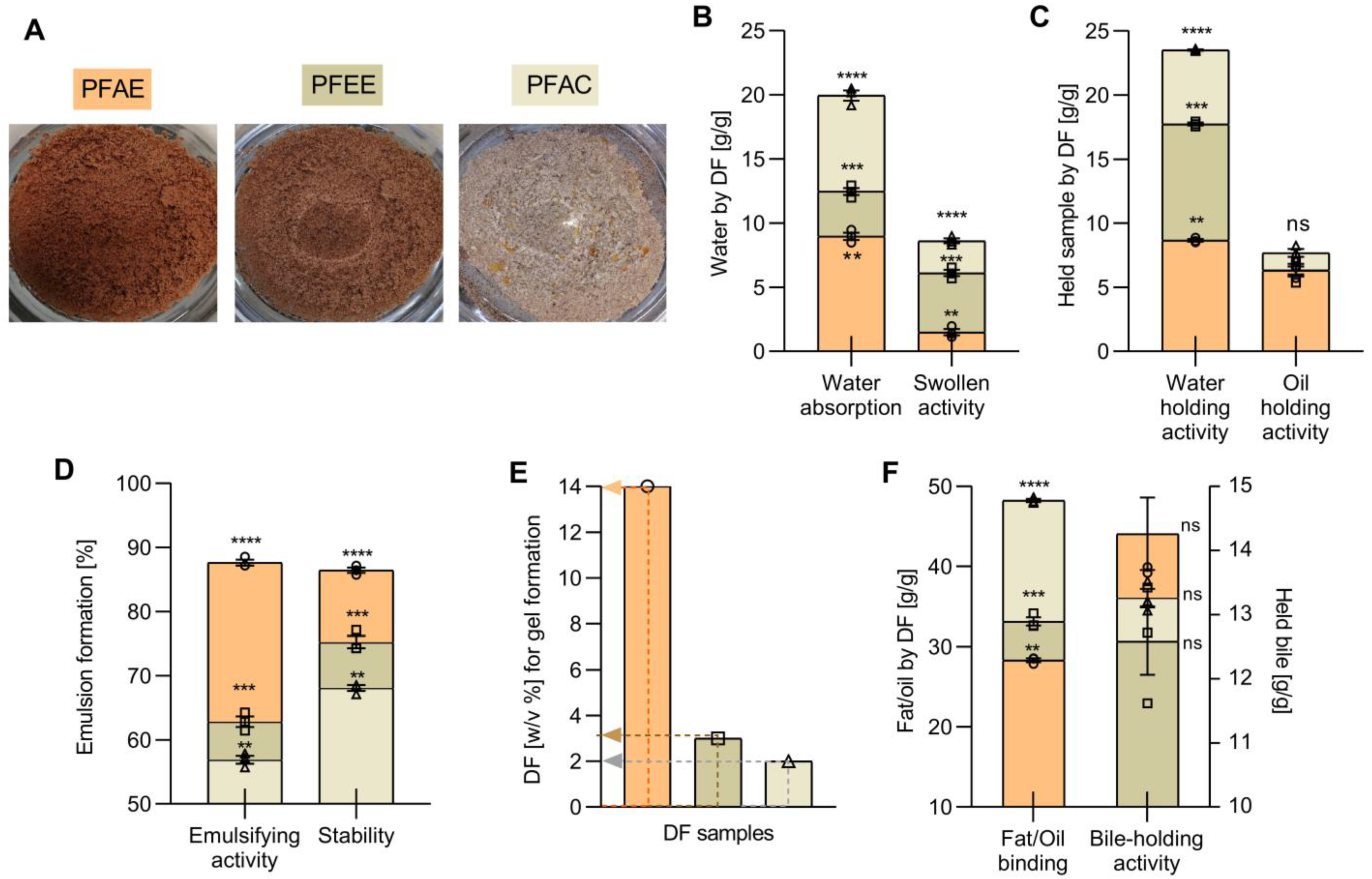
2.5. Techno-Functional Properties
2.6. Physio-Functional Properties
2.7. In Vitro Gastrointestinal Digestion
2.8. Probiotic Fermentation Process (PFP)
2.9. Cell Culture
2.9.1. Cell Viability
2.9.2. Trans Epithelial Electrical Resistance (TEER) Response of Caco-2 Cell Monolayer
2.10. Determination of Biocompounds
2.10.1. Sample Preparation
2.10.2. The Folin-Ciocalteu Reagent Assay
2.10.3. Total Carotenoid Content (TCC)
2.10.4. Total Flavonoid Content (TFC)
2.11. Recovery and Bioaccessibility Index of Free and Bonded Compounds
2.12. Antioxidant Activity
2.12.1. The 2,2′-Azino-Bis(3-Ethylbenzothiazoline-6-Sulfonic Acid) (ABTS•+) Radical Cation-Based Decolorization Assay
2.12.2. The 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical-Based Assay
2.13. High Performance Liquid Chromatography Analysis (HPLC-DAD)
2.14. HPLC Coupled to Electro-Spray Ion Trap Mass Spectrometry (HPLC-DAD-ESI-IT-MSn)
2.15. Statistical Analysis
3. Results
3.1. Physicochemical Analysis
3.2. Techno-Functional Properties
3.3. Physio-Functional Properties
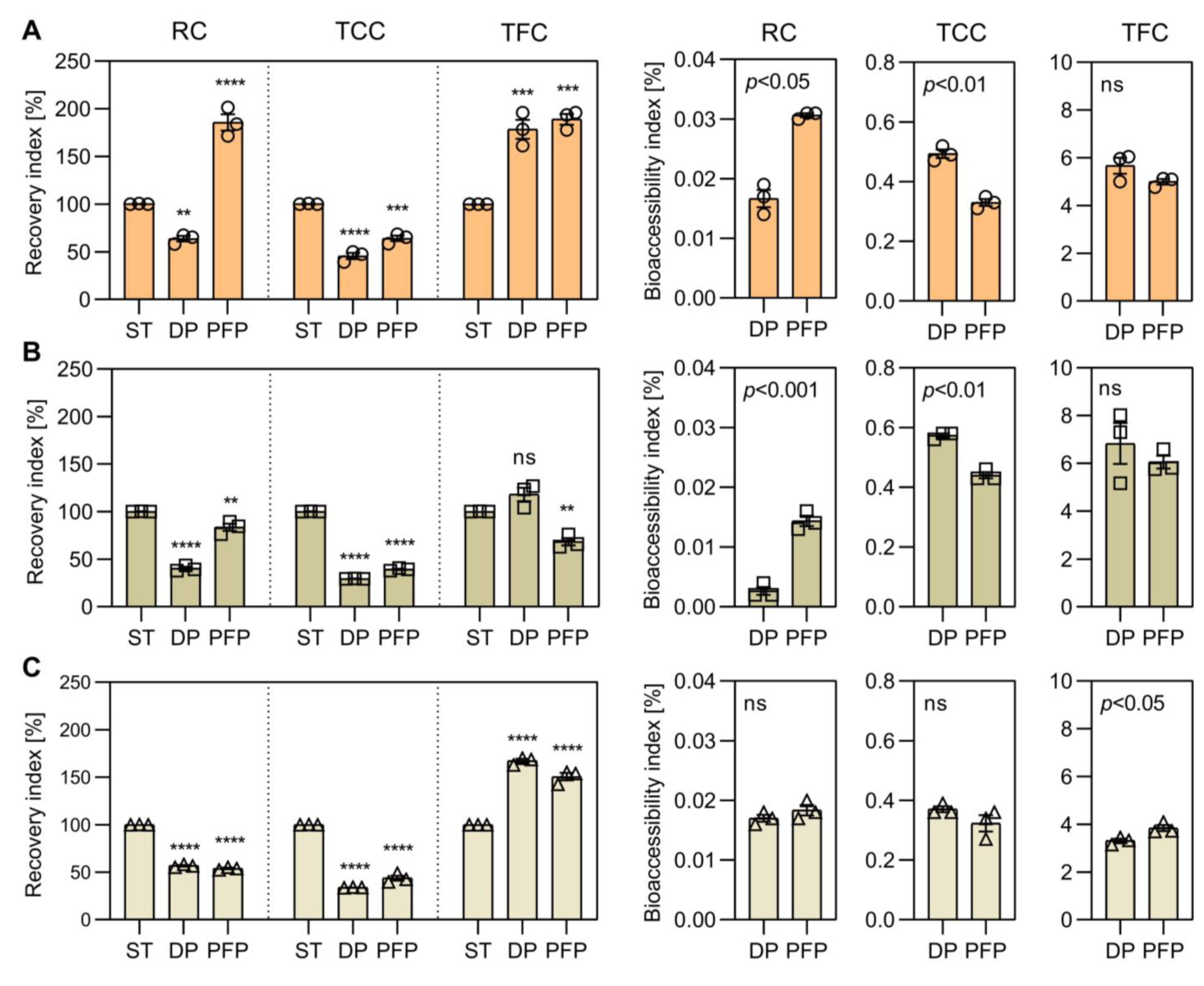
3.4. Recovery and Bioaccessibility of Free and Bonded Biocompounds
3.5. Antioxidant Activity
3.6. Phenolic Profile
3.7. Polyphenolic Quantification
3.8. SCFA Profile
3.9. Cytotoxicity Assays
3.10. TEER Response of Cell Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kearney, J. Food consumption trends and drivers. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2793–2807. [Google Scholar] [CrossRef]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomas-Barberan, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Foods Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Matheus, J.R.V.; Andrade, C.J.D.; Miyahira, R.F.; Fai, A.E.C. Persimmon (Diospyros kaki L.): Chemical properties, bioactive compounds and potential use in the development of new products—A review. Food Rev. Int. 2020, 1–18. [Google Scholar] [CrossRef]
- Matsumura, Y.; Ito, T.; Yano, H.; Kita, E.; Mikasa, K.; Okada, M.; Furutani, A.; Murono, Y.; Shibata, M.; Nishii, Y.; et al. Antioxidant potential in non-extractable fractions of dried persimmon (Diospyros kaki Thunb.). Food Chem. 2016, 202, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Cano, M.P.; Gómez-Maqueo, A.; Fernández-López, R.; Welti-Chanes, J.; García-Cayuela, T. Impact of high hydrostatic pressure and thermal treatment on the stability and bioaccessibility of carotenoid and carotenoid esters in astringent persimmon (Diospyros kaki Thunb, var. Rojo Brillante). Food Res. Int. 2019, 123, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Gea-Botella, S.; Agulló, L.; Martí, N.; Martínez-Madrid, M.C.; Lizama, V.; Martín-Bermudo, F.; Berná, G.; Saura, D.; Valero, M. Carotenoids from persimmon juice processing. Food Res. Int. 2021, 141, 109882. [Google Scholar] [CrossRef]
- Jiménez-Sánchez, C.; Lozano-Sánchez, J.; Marti, N.; Saura, D.; Valero, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Characterization of polyphenols, sugars, and other polar compounds in persimmon juices produced under different technologies and their assessment in terms of compositional variations. Food Chem. 2015, 182, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.N.; Shin, M.R.; Shin, S.H.; Lee, A.R.; Lee, J.Y.; Seo, B.I.; Kim, M.Y.; Kim, T.H.; Noh, J.S.; Rhee, M.H.; et al. Study of Antiobesity Effect through Inhibition of Pancreatic Lipase Activity of Diospyros kaki Fruit and Citrus unshiu Peel. BioMed Res. Int. 2016, 2016, 1723042. [Google Scholar] [CrossRef] [Green Version]
- Son, J.E.; Hwang, M.K.; Lee, E.; Seo, S.G.; Kim, J.E.; Jung, S.K.; Kim, J.R.; Ahn, G.H.; Lee, K.W.; Lee, H.J. Persimmon peel extract attenuates PDGF-BB-induced human aortic smooth muscle cell migration and invasion through inhibition of c-Src activity. Food Chem. 2013, 141, 3309–3316. [Google Scholar] [CrossRef]
- Ueda, K.; Kawabata, R.; Irie, T.; Nakai, Y.; Tohya, Y.; Sakaguchi, T. Inactivation of pathogenic viruses by plant-derived tannins: Strong effects of extracts from persimmon (Diospyros kaki) on a broad range of viruses. PLoS ONE 2013, 8, e55343. [Google Scholar] [CrossRef]
- Orejuela-Escobar, L.M.; Landázuri, A.C.; Goodell, B. Second generation biorefining in Ecuador: Circular bioeconomy, zero waste technology, environment and sustainable development: The nexus. J. Bioresour. Bioprod. 2021, 6, 83–107. [Google Scholar] [CrossRef]
- Soliman, G.A. Dietary Fiber, Atherosclerosis, and Cardiovascular Disease. Nutrients 2019, 11, 1155. [Google Scholar] [CrossRef] [Green Version]
- Blad, C.C.; Tang, C.; Offermanns, S. G protein-coupled receptors for energy metabolites as new therapeutic targets. Nat. Rev. Drug Discov. 2012, 11, 603–619. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Ahmed, M.; Eun, J.B. Effect of blanchig and drying temperatures on the physicochemical characteristics, dietary fiber and antioxidant-related parameters of dried persimmons peel powder. Int. J. Food Sci. Nutr. 2010, 61, 11. [Google Scholar] [CrossRef]
- AOAC Association of Official Analytical Chemist. Official Methods of Analysis; Association of Official Analytical Chemist: Washington, DC, USA, 2012. [Google Scholar]
- Raghavendra, S.N.; Rastogi, N.K.; Raghavarao, K.S.M.S.; Tharanathan, R.N. Dietary fiber from coconut residue: Effects of different treatments and particle size on the hydration properties. Eur. Food Res. Technol. 2004, 218, 563–567. [Google Scholar] [CrossRef]
- Robertson, J.A.; de Monredon, F.D.; Dysseler, P.; Guillon, F.; Amado, R.; Thibault, J.F. Hydration properties of dietary fibre and resistant starch: A European collaborative study. LWT-Food Sci. Technol. 2000, 33, 72–79. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Dietary fibre and physicochemical properties of several edible seaweeds from the northwestern Spanish coast. Food Res. Int. 2010, 43, 2289–2294. [Google Scholar] [CrossRef]
- Yasumatsu, K.; Sawada, K.; Moritaka, S.; Misaki, M.; Toda, J.; Wada, T.; Ishii, K. Whipping and emulsifying properties of soybean products. Agric. Biol. Chem. 1972, 36, 719–727. [Google Scholar] [CrossRef]
- Chau, C.F.; Cheung, P.C.K. Functional properties of flours prepared from three Chinese indigenous legume seeds. Food Chem. 1998, 61, 429–433. [Google Scholar] [CrossRef]
- Eastwood, M.A.; Kirkpatrick, J.R.; Mitchell, W.D.; Bone, A.; Hamilton, T. Effects of dietary supplements of wheat bran and cellulose on faeces and bowel function. Britrish Med. J. 1973, 4, 392–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food–An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Zamora, A.; Pastoriza, S.; Rufián-Henares, J. Revalorization of coffee by-products. Prebiotic, antimicrobial and antioxidant properties. LWT-Food Sci. Technol. 2015, 61, 12–18. [Google Scholar] [CrossRef]
- Kwon, H.J.; Kim, M.Y.; Seo, K.H.; Lee, H.G.; Kim, H. In-vitro prebiotic activity of grape seed fluor highly rich in flavonoid and dietary fiber. J. Food Nutr. Res. 2018, 6, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Praveen, M.A.; Parvathy, K.K.; Jayabalan, R.; Balasubramanian, P. Dietary fiber from Indian edible seaweeds and its in-vitro prebiotic effect on the gut microbiota. Food Hydrocoll. 2019, 96, 343–353. [Google Scholar] [CrossRef]
- Zarinah, Z.; Anis, A.A.; Napisah, H.; Shazila, S. Prebiotic activity score of breadfruit resistant starch (Artocarpus altilis), breadfruit flour, and inulin during in-vitro fermentation by pure cultures (Lactobacillus plantarum and Bifidobacterium bifidum). J. Agrobiotechnology 2018, 9, 122–131. [Google Scholar]
- Saotome, K.; Morita, H.; Umeda, M. Cytotoxicity test with simplified crystal violet staining method using microtitre plates and its application to injection drugs. Toxicol. Vitr. 1989, 3, 317–321. [Google Scholar] [CrossRef]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of butyrate on intestinal barrier function in a Caco-2 cell monolayer model of intestinal barrier. Pediatric Res. 2007, 61, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Kim, C.Y.; Kaur, A.; Lamothe, L.; Shaikh, M.; Keshavarzian, A.; Hamaker, B.R. Dietary fibre-based SCFA mixtures promote both protection and repair of intestinal epithelial barrier function in a Caco-2 cell model. Food Funct. 2017, 8, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Hubatsch, I.; Ragnarsson, E.G.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Górnaś, P.; Dwiecki, K.; Siger, A.; Tomaszewska-Gras, J.; Michalak, M.; Polewski, K. Contribution of phenolic acids isolated from green and roasted boiled-type coffee brews to total coffee antioxidant capacity. Eur. Food Res. Technol. 2015, 242, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Cuvi, M.J.; Moreno, C.; Zaro, M.J.; Vicente, A.R.; Concellón, A. Improvement of the antioxidant properties and postharvest life of three exotic Andean fruits by UV-C treatment. J. Food Qual. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Luximon-Ramma, A.; Bahorun, T.; Soobrattee, M.A.; Aruoma, O.I. Antioxidant activities of phenolic, proanthocyanidin, and flavonoid components in extracts of Cassia fistula. J. Agric. Food Chem. 2002, 50, 5042–5047. [Google Scholar] [CrossRef]
- Ortega, N.; Macià, A.; Romero, M.P.; Reguant, J.; Motilva, M.J. Matrix composition effect on the digestibility of carob flour phenols by an in-vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Calero, R.R.; Lagoa-Costa, B.; Fernandez-Feal, M.M.D.C.; Kennes, C.; Veiga, M.C. Volatile fatty acids production from cheese whey: Influence of pH, solid retention time and organic loading rate. J. Chem. Technol. Biotechnol. 2018, 93, 1742–1747. [Google Scholar] [CrossRef]
- Martínez-Las Heras, R.; Landines, E.F.; Heredia, A.; Castelló, M.L.; Andrés, A. Influence of drying process and particle size of persimmon fibre on its physicochemical, antioxidant, hydration and emulsifying properties. J. Food Sci. Technol. 2017, 54, 2902–2912. [Google Scholar] [CrossRef] [PubMed]
- Hamauzu, Y.; Suwannachot, J. Non-extractable polyphenols and in vitro bile acid-binding capacity of dried persimmon (Diospyros kaki) fruit. Food Chem. 2019, 293, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Maroun, G.R.; Rajha, H.N.; El Darra, N.; El Kantar, S.; Chacar, S.; Debs, E.; Louka, N. Emerging technologies for the extraction of polyphenols from natural sources. In Polyphenols: Properties, Recovery, and Applications; Galanakis, C.M., Ed.; Woodhead Publishing: Vienna, Austria, 2018; pp. 265–293. [Google Scholar]
- Lucas-González, R.; Viuda-Martos, M.; Pérez-Álvarez, J.; Fernández-López, J. Evaluation of Particle Size Influence on Proximate Composition, Physicochemical, Techno-Functional and Physio-Functional Properties of Flours Obtained from Persimmon (Diospyros kaki Trumb.) Coproducts. Plant Foods Hum. Nutr. 2017, 72, 67–73. [Google Scholar] [CrossRef]
- Park, Y.-S.; Jung, S.-T.; Kang, S.-G.; Delgado-Licon, E.; Leticia Martinez Ayala, A.; Tapia, M.S.; Martín-Belloso, O.; Trakhtenberg, S.; Gorinstein, S. Drying of persimmons (Diospyros kaki L.) and the following changes in the studied bioactive compounds and the total radical scavenging activities. LWT Food Sci. Technol. 2006, 39, 748–755. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, Y.; Huang, J.; Zhang, H.; Lin, Q.; Han, L.; Liu, J.; Wang, J.; Liu, H. Insoluble dietary fiber from soy hulls regulates the gut microbiota in vitro and increases the abundance of bifidobacteriales and lactobacillales. J. Food Sci. Technol. 2020, 57, 152–162. [Google Scholar] [CrossRef]
- Tessmer, M.A.; Besada, C.; Hernando, I.; Appezzato-da-Gloria, B.; Aquiles, A.; Salvador, A. Microstructural changes while persimmon fruits mature and ripen. Comparison between astringent and non-astringent cultivars. Postharvest Biol. Technol. 2016, 120, 52–60. [Google Scholar] [CrossRef]
- Salvador, A.; Arnal, L.; Besada, C.; Larrea, V.; Quiles, A.; Pérez-Munuera, I. Physiological and structural changes during ripening and deastringency treatment of persimmon fruit cv. ‘Rojo Brillante’. Postharvest Biol. Technol. 2007, 46, 181–188. [Google Scholar] [CrossRef]
- Rodríguez-Garayar, M.; Martín-Cabrejas, M.A.; Esteban, R.M. High hydrostatic pressure in astringent and non-astringent persimmons to obtain fiber-enriched ingredients with improved functionality. Food Bioprocess Technol. 2017, 10, 854–865. [Google Scholar] [CrossRef]
- Akubor, P.I.; Ukwuru, M.U. Functional properties and biscuit making potential of soybean and cassava flour blends. Plant Foods Hum. Nutr. 2003, 58, 1–12. [Google Scholar] [CrossRef]
- Houten, S.M.; Watanabe, M.; Auwerx, J. Endocrine functions of bile acids. EMBO J. 2006, 25, 1419–1425. [Google Scholar] [CrossRef]
- Niu, Y.; Xie, Z.; Zhang, H.; Sheng, Y.; Yu, L.L. Effects of structural modifications on physicochemical and bile acid-binding properties of psyllium. J. Agric. Food Chem. 2013, 61, 596–601. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mamet, T.; Ge, Z.Z.; Zhang, Y.; Li, C.M. Interactions between highly galloylated persimmon tannins and pectins. Int. J. Biol. Macromol. 2018, 106, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Barba, F.J.; Mariutti, L.R.; Bragagnolo, N.; Mercadante, A.Z.; Barbosa-Canovas, G.V.; Orlien, V. Bioaccessibility of bioactive compounds from fruits and vegetables after thermal and nonthermal processing. Trends Food Sci. Technol. 2017, 67, 195–206. [Google Scholar] [CrossRef]
- Padayachee, A.; Netzel, G.; Netzel, M.; Day, L.; Mikkelsen, D.; Gidley, M.J. Lack of release of bound anthocyanins and phenolic acids from carrot plant cell walls and model composites during simulated gastric and small intestinal digestion. Food Funct. 2013, 4, 906–916. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary Fiber Confers Protection against Flu by Shaping Ly6c. Immunity 2018, 48, 992–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada-Ortigoza, V.; Garcia-Amezquita, L.E.; Kazem, A.E.; Campanella, O.H.; Cano, M.P.; Hamaker, B.R.; Serna-Saldívar, S.O.; Welti-Chanes, J. In vitro fecal fermentation of high pressure-treated fruit peels used as dietary fiber sources. Molecules 2019, 24, 697. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Chandel, D.; Shukla, G. Antigenotoxicity and cytotoxic potentials of metabiotics extracted from isolated probiotic, Lactobacillus rhamnosus MD 14 on Caco-2 and HT-29 human colon cancer cells. Nutr. Cancer 2020, 72, 110–119. [Google Scholar] [CrossRef]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothari, C.; Patel, A.T.; Purama, R.K.; Dave, J.M.; Vyas, B.R. Potential of probiotics, prebiotics and synbiotics for management of colorectal cancer. Gut Microbes 2013, 4, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Arun, K.B.; Madhavan, A.; Reshmitha, T.R.; Sithara, T.; Nisha, P. Short chain fatty acids enriched fermentation metabolites of soluble dietary fibre from Musa paradisiaca drives HT29 colon cancer cells to apoptosis. PLoS ONE 2019, 14, e0216604. [Google Scholar] [CrossRef]
- Wang, P.; Henning, S.M.; Heber, D. Limitations of MTT and MTS-based assays for measurement of antiproliferative activity of green tea polyphenols. PLoS ONE 2010, 5, e10202. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.Y.; Osaka, T.; Moriyama, E.; Date, Y.; Kikuchi, J.; Tsuneda, S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol. Rep. 2015, 3, e12327. [Google Scholar] [CrossRef] [Green Version]
- Castro-Herrera, V.M.; Rasmussen, C.; Wellejus, A.; Miles, E.A.; Calder, P.C. In Vitro Effects of Live and Heat-Inactivated Bifidobacterium animalis subsp. lactis, BB-12 and Lactobacillus rhamnosus GG on Caco-2 Cells. Nutrients 2020, 12, 1719. [Google Scholar] [CrossRef]
- Ohata, A.; Usami, M.; Miyoshi, M. Short-chain fatty acids alter tight junction permeability in intestinal monolayer cells via lipoxygenase activation. Nutrition 2005, 21, 838–847. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | PFAE | PFEE | PFAC |
|---|---|---|---|
| Protein | 0.0006 ± 0.0001 a | 0.0014 ± 0.0000 a, b | 0.0021 ± 0.0001 b |
| Ashes | 0.08 ± 0.00 a | 0.16 ± 0.00 b | 0.04 ± 0.00 a |
| pH | 5.50 ± 0.02 a | 6.72 ± 0.01 b | 6.83 ± 0.02 c |
| TSS * | 1.17 ± 0.29 a | 20.33 ± 0.29 b | 21.17 ± 0.29 b |
| IDF | 0.62 ± 0.13 a | 0.68 ± 0.05 a | 0.64 ± 0.08 a |
| SDF | 0.19 ± 0.02 a | 0.14 ± 0.03 a | 0.30 ± 0.16 b |
| TDF | 0.82 ± 0.11 a | 0.81 ± 0.08 a | 0.94 ± 0.08 b |
| ABTS•+ | DPPH | ||
|---|---|---|---|
| Samples | Treatments | mg Trolox/g Sample | mg Trolox/g Sample |
| Extracted fiber | PFAE | 2.81 ± 0.34 a | 2.12 ± 0.13 a |
| PFEE | 2.71 ± 0.09 b | 2.04 ± 0.09 b | |
| PFAC | 2.71 ± 0.05 b | 2.04 ± 0.04 b | |
| Digested fiber | PFAE | 1.21 ± 0.11 f | 0.91 ± 0.11 f |
| PFEE | 1.43 ± 0.30 e | 1.08 ± 0.09 e | |
| PFAC | 1.05 ± 0.09 g | 0.79 ± 0.19 g | |
| Supernatant digested fiber | PFAE | 1.68 ± 0.11 d | 1.26 ± 0.11 d |
| PFEE | 1.68 ± 0.09 c, d | 1.26 ± 0.09 c, d | |
| PFAC | 1.68 ± 0.05 c, d | 1.26 ± 0.05 c, d | |
| Fermented fiber | PFAE | 0.73 ± 0.12 h | 0.55 ± 0.25 h |
| PFEE | 0.67 ± 0.32 i | 0.51 ± 0.12 i | |
| PFAC | 0.61 ± 0.13 j | 0.46 ± 0.15 j | |
| Supernatant fermented fiber | PFAE | 1.68 ± 0.10 c, d | 1.26 ± 0.10 c, d |
| PFEE | 1.68 ± 0.13 c, d | 1.26 ± 0.13 c, d | |
| PFAC | 1.68 ± 0.22 c | 1.26 ± 0.22 c |
| No. | RT (Min) | HPLC-DAD UV−vis Spectrum λmax (nm) | [M−H]− m/z | HPLC-DAD-ESI-IT-MSn Experiments m/z | Compound Identity | Molecular Formula | Extracts Present |
|---|---|---|---|---|---|---|---|
| 1 | 7.7 | 272/280 | 126.1 (100) 170.5 (57) | MS2 [170.5] 126.1 (100) 168.4 (31) 124.1 (15.4) | Gallic acid * | C7H6O5 | U1, U2, U3, DP1, DP2, DP3, FP1, FP2, FP3 |
| 2 | 16.0 | 305/272 | 164.5 (100) | MS2 [164.5] 120.1 (100) | p-Coumaric acid * | C9H8O3 | U1, U2, U3, DP1, DP2, DP3, DS3, |
| 3 | 16.1 | 260/272 | 303.7 (100) | MS2 [303.7] 301.6 (100) 259.2 (60) 186.5 (16.9) | Ellagic acid * | C14H6O8 | U1, U2, U3, DP1 DP2, DP3, DS1, DS2, DS3 |
| 4 | 16.6 | 320/272 | 209.8 (100) 225.0 (45) | MS2 [209.8] 165.4 (100) 150.3 (60) 164.4 (22) | 3,5-Dimethoxy-4-hydroxycinnamic acid * | C11H12O5 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3 |
| 5 | 18.7 | 305/280 | 301.6 (100) 241.1 (94) 138.2 (76) | MS2 [301.6] 138.2 (100) 139.2 (10) 94.0 (4) | Salicylic acid * | C7H6O3 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3 |
| 6 | 20.8 | 270/332 | 514.3 (100) | MS2 [514.3] 170.9 (100) 342.1 (31) | 3,5-Dicaffeoylquinic acid ** | C25H24O12 | DP1, DP2, DP3 |
| 7 | 20.9 | 270/332 | 1021.3 (100) 948.1 (42) 194.7 (42) | MS2 [948.6] 888.5 (100) 909.7 (47) 930.1 (42) | Spinacetin 3-O- (2″-p-coumaroylglucosyl) (1->6)-[apiosyl (1->2)]-glucoside ** | C43H48O24 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 8 | 21.6 | 270/332 | 498.3 (100) | MS2 [498.6] 350.6 (100) 412.9 (94) | 6″-O-Malonyldaidzin ** | C24H22O12 | DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 9 | 22.4 | 270/332 | 331.2 (100) | MS2 [331.3] 228.9 (100) 293 (96) 210.8 (90) | Galloyl-hexoside I ** | C13H16O10 | DP1, DP2, DP3, FP1, FP2, FP3 |
| 10 | 22.6 | 270/332 | 334.4 (100) | MS2 [334.4] 316.1 (100) 172.6 (22) 332.2 (16) 287.7 (2) | Galloyl-hexoside II ** | C13H16O10 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 11 | 23.2 | 270/332 | 448.3 (100) | MS2 [448.3] 384.1 (100) 402.2 (54) | Cyanidin 3-O-galactoside ** | C21H21O11 | DP1, DP2, DP3, DS1, DS2, FP1, FP2, FP3 |
| 12 | 23.5 | 270/332 | 289.8 (100) | MS2 [289.8] 271.6 (100) 142.2 (22) 130.2 (7) | Epicatechin * | C15H14O6 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 13 | 24.2 | 280/320 | 446.3 (100) | MS2 [446.1] 249.2 (100) | Kaempferol-7-glucoside ** | C21H19O11 | DP1, DP2, DP3, DS1, DS2, DS3, FP1 |
| 14 | 25.2 | 280/320 | 783.5 (100) 391.3 (27.7) | MS2 [391.3] 343.1 (100) 170.9 (56) | Quercetin glucoside I ** | C33H40O21 | DP1, DP2, DP3, FP1, FP2, FP3 |
| 15 | 25.9 | 272/280 | 389.5 (100) | MS2 [389.6] 339 (100) 342.2 (90) 297.2 (43) | Resveratrol glucoside I ** | C20H22O8 | DP1, DP2, FP1, FP1, FP2, FP3 |
| 16 | 26.1 | 320/360 | 316.2 (100) | MS2 [316.5] 172.6 (100) 297.9 (75) 128.3 (7) | Methoxyluteolin ** | C16H12O7 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 17 | 26.4 | 272/280 | 389.2 (100) | MS2 [389.2] 369.1 (100) 352.9 (56) 296.1 (38) 343.1 (21) | Resveratrol glucoside II ** | C20H22O8 | DP1, DP2, DP3, DS1, DS2, U3, FP1 |
| 18 | 27.2 | 280/320 | 783.6 (100) 391.3 (30) | MS2 [391.3] 218.9 (100) 357.1 (92) | Quercetin glucoside II ** | C33H40O21 | DP1, DP2, DP3 |
| 19 | 27.6 | 280/320 | 754.2 (100) 718 (95) | MS2 [754.2] 718.4 (100) MS3 [718.4] 661.4 (100) | Kaempferol 3-O-glucosyl-rhamnosyl-galactoside ** | C21H20O11 | U1, U2, U3, DP1, DP2, DP3, DS1, DS2, DS3, FP1, FP2, FP3 |
| 20 | 28.5 | 280/320 | 267.6 (100) | MS2 [267.6] 98.0 (100) 297.9 (75) 128.3 (7) | 7-Hydroxy-4′-methoxyisoflavone ** | C16H12O4 | U1, U2, U3, FP1, FP2 |
| 21 | 29.0 | 280/320 | 312.1 (100) | MS2 [312.1] 98.0 (100) 310.0 (18) 124.1 (17) | 5,4′-Dihydroxy-6,7-dimethoxyflavone ** | C17H14O6 | U1, U2, U3, DP1, DS1, DS2, DS3, FP1, FP2, FP3 |
| 22 | 29.5 | 320/360 | 355.2 (100) | MS2 [355.2] 265 (100) 291 (56) 234.8 (29) | Ferulic acid glucoside ** | C16H20O9 | DP1, DP2, DP3 |
| 23 | 30.2 | 320/360 | 297.2 (100) | MS2 [297.4] 277.1 (100) 234.5 (30) | p-Coumaroyl tartaric acid ** | C13H12O8 | DP1, DP2, FP1 |
| Phenolic Compounds (mg/g or mL Sample) | ||||||
|---|---|---|---|---|---|---|
| Samples | Treatment | Gallic Acid | Sinapic Acid | p-Coumaric Acid | Salicylic Acid | Ellagic Acid |
| Extracted fiber | PFAE | 58.63 ± 0.46 c | 0.69 ± 0.04 b | 0.57 ± 0.01 d | 2.19 ± 0.18 b | 1.26 ± 0.01 c |
| PFEE | 91.31 ± 2.45 b | 0.69 ± 0.04 b | 0.14 ± 0.01 d | 2.18 ± 0.15 b | 1.89 ± 0.19 b | |
| PFAC | 114.72 ± 2.60 a | 0.77 ± 0.08 a | 3.73 ± 0.22 b | 2.24 ± 0.09 b | 2.05 ± 0.08 b | |
| Digested fiber | PFAE | 80.42 ± 4.24 b | 0.46 ± 0.00 d | 1.28 ± 0.08 c | 3.57 ± 0.18 a | 1.03 ± 0.05 d |
| PFEE | 64.94 ± 3.32 c | 0.34 ± 0.03 f | 0.08 ± 0.00 e | 1.16 ± 0.09 e | 1.90 ± 0.11 b, c | |
| PFAC | 104.20 ± 9.38 a | 0.67 ± 0.03 b | 3.57 ± 0.18 b | 1.35 ± 0.11 e | 4.22 ± 0.13 a | |
| Supernatant digested fiber | PFAE | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| PFEE | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| PFAC | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Fermented fiber | PFAE | 33.06 ± 2.64 d | 0.51 ± 0.01 d | <0.001 | 1.87 ± 0.15 c | 1.40 ± 0.06 c |
| PFEE | 50.04 ± 0.97 c | 0.47 ± 0.04 d | <0.001 | 1.71 ± 0.14 d | 2.01 ± 0.04 b | |
| PFAC | 57.01 ± 3.04 c | 0.57 ± 0.01 c | 4.56 ± 0.09 a | 2.20 ± 0.11 b | 4.59 ± 0.37 a | |
| Supernatant fermented fiber | PFAE | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| PFEE | 0.006 ± 0.00 e | <0.001 | <0.001 | <0.001 | <0.001 | |
| PFAC | 0.004 ± 0.00 e | <0.001 | <0.001 | <0.001 | <0.001 | |
| SCFA (mg/L) | Treatment | Undigested Fraction | Digested Fraction |
|---|---|---|---|
| Acetic acid | PFAE | 0.65 ± 0.01 a, b | 0.67 ± 0.01 a, b |
| PFEE | 0.65 ± 0.00 a | 0.78 ± 0.01 c | |
| PFAC | 0.68 ± 0.01 b | 0.76 ± 0.01 c | |
| Propionic acid | PFAE | 0.09 ± 0,00 a | 0.23 ± 0.01 b |
| PFEE | n.d. | 0.21 ± 0.01 b | |
| PFAC | 0.20 ± 0.02 b | 0.20 ± 0.01 b | |
| Butyric acid | PFAE | 0.95 ± 0.01 a | 1.29 ± 0.02 d |
| PFEE | 1.03 ± 0.00 b | 1.10 ± 0.00 d | |
| PFAC | 1.06 ± 0.00 c | 1.01 ± 0.00 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Bermeo, J.; Moreno-Chamba, B.; Martínez-Madrid, M.C.; Saura, D.; Valero, M.; Martí, N. Potential of Persimmon Dietary Fiber Obtained from Byproducts as Antioxidant, Prebiotic and Modulating Agent of the Intestinal Epithelial Barrier Function. Antioxidants 2021, 10, 1668. https://doi.org/10.3390/antiox10111668
Salazar-Bermeo J, Moreno-Chamba B, Martínez-Madrid MC, Saura D, Valero M, Martí N. Potential of Persimmon Dietary Fiber Obtained from Byproducts as Antioxidant, Prebiotic and Modulating Agent of the Intestinal Epithelial Barrier Function. Antioxidants. 2021; 10(11):1668. https://doi.org/10.3390/antiox10111668
Chicago/Turabian StyleSalazar-Bermeo, Julio, Bryan Moreno-Chamba, María Concepción Martínez-Madrid, Domingo Saura, Manuel Valero, and Nuria Martí. 2021. "Potential of Persimmon Dietary Fiber Obtained from Byproducts as Antioxidant, Prebiotic and Modulating Agent of the Intestinal Epithelial Barrier Function" Antioxidants 10, no. 11: 1668. https://doi.org/10.3390/antiox10111668