
Neck Muscle Vibration Alters Cerebellar Processing Associated with Motor Skill Acquisition of a Proprioceptive-Based Task

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. SEP Stimulation Protocol and Recording Parameters
2.3. Neck Muscle Vibration
2.4. Force Matching Tracking Task
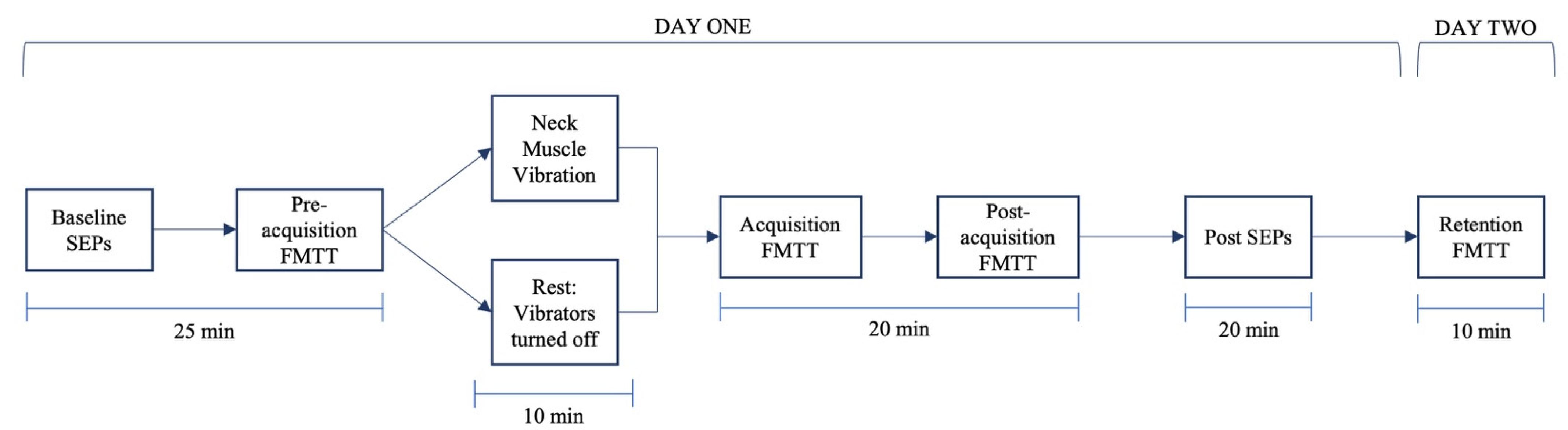
2.5. Experimental Procedure
2.6. Data Processing
2.7. Statistical Analysis
3. Results
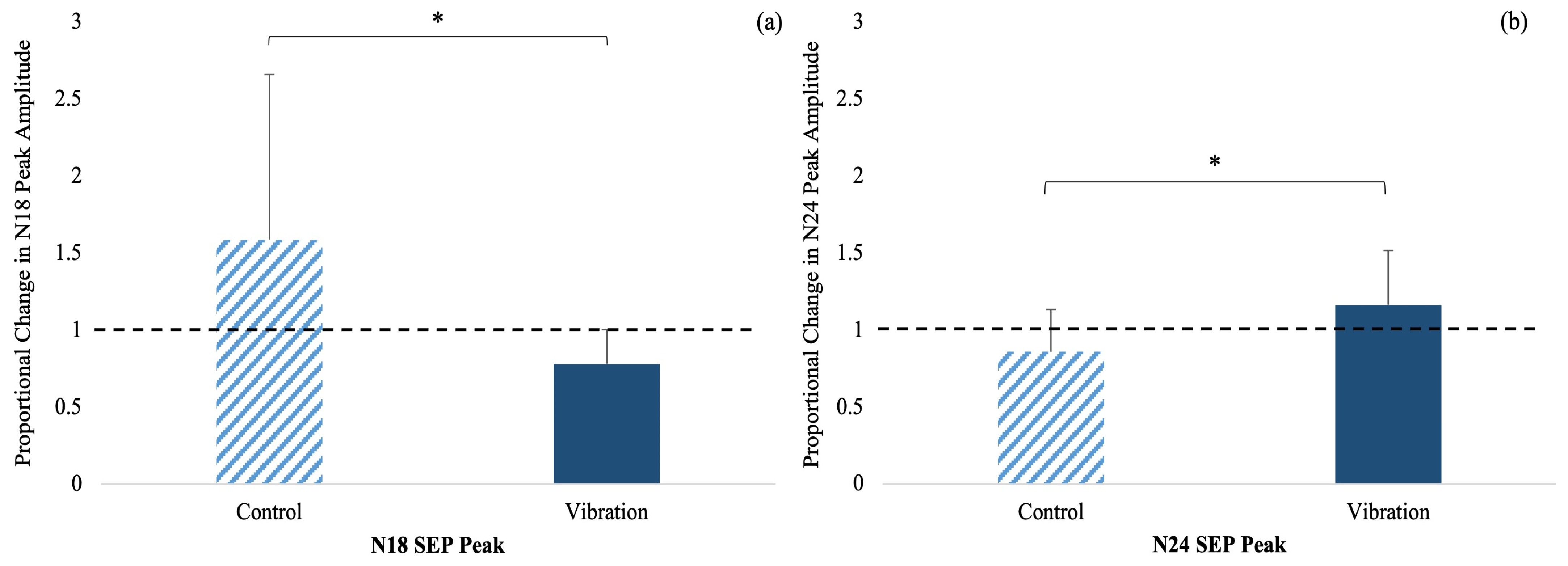
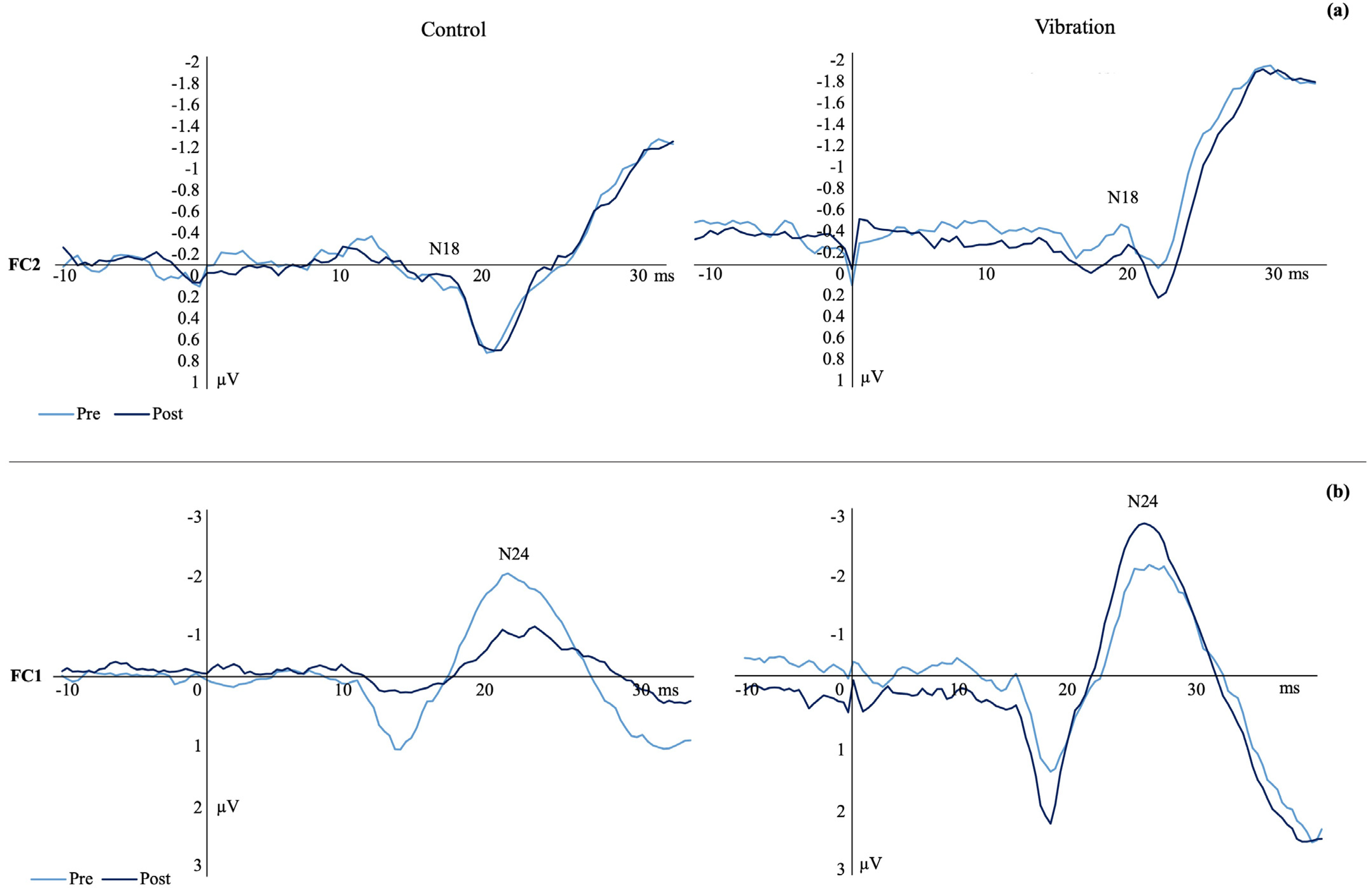
3.1. SEP Peak Amplitudes
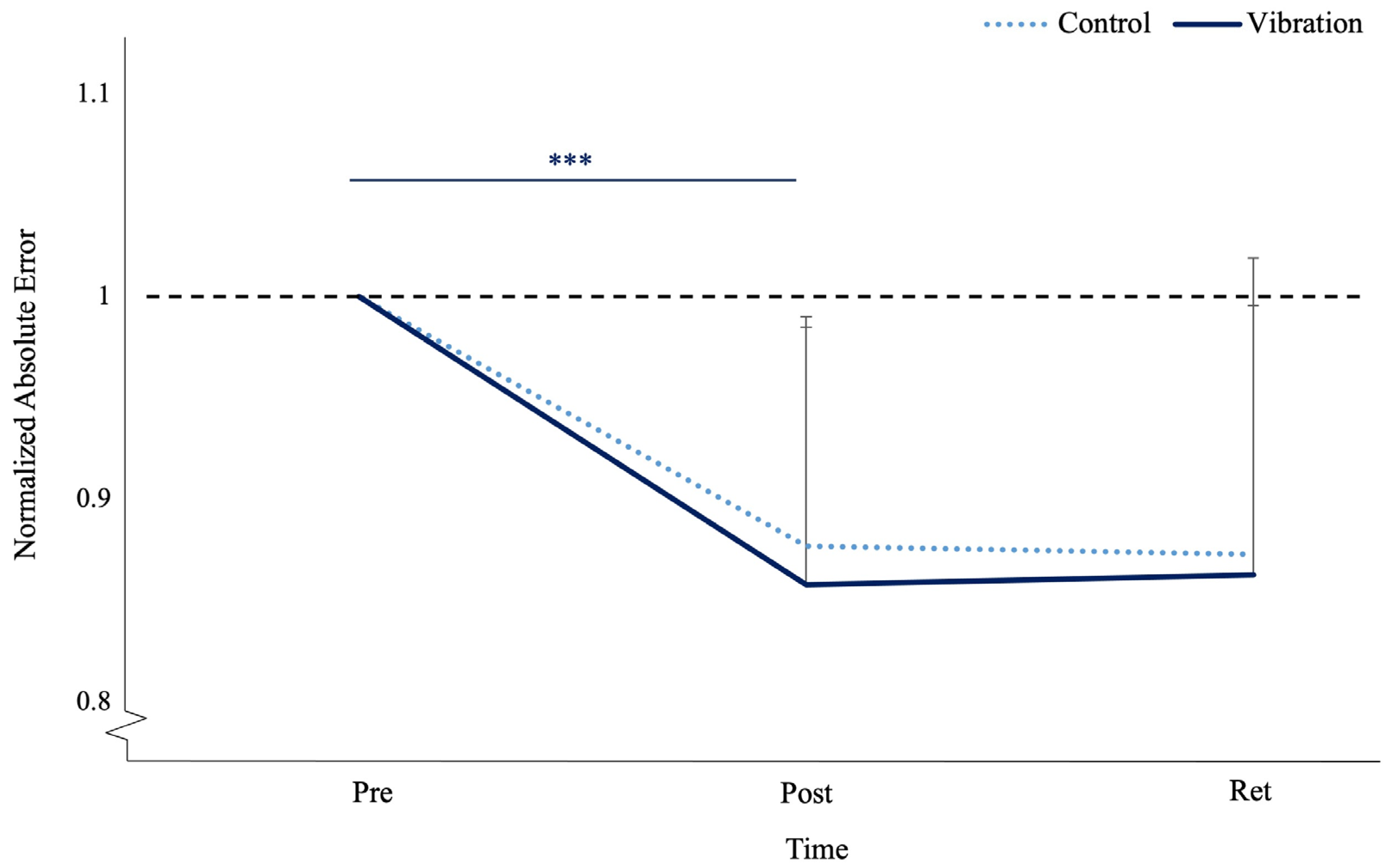
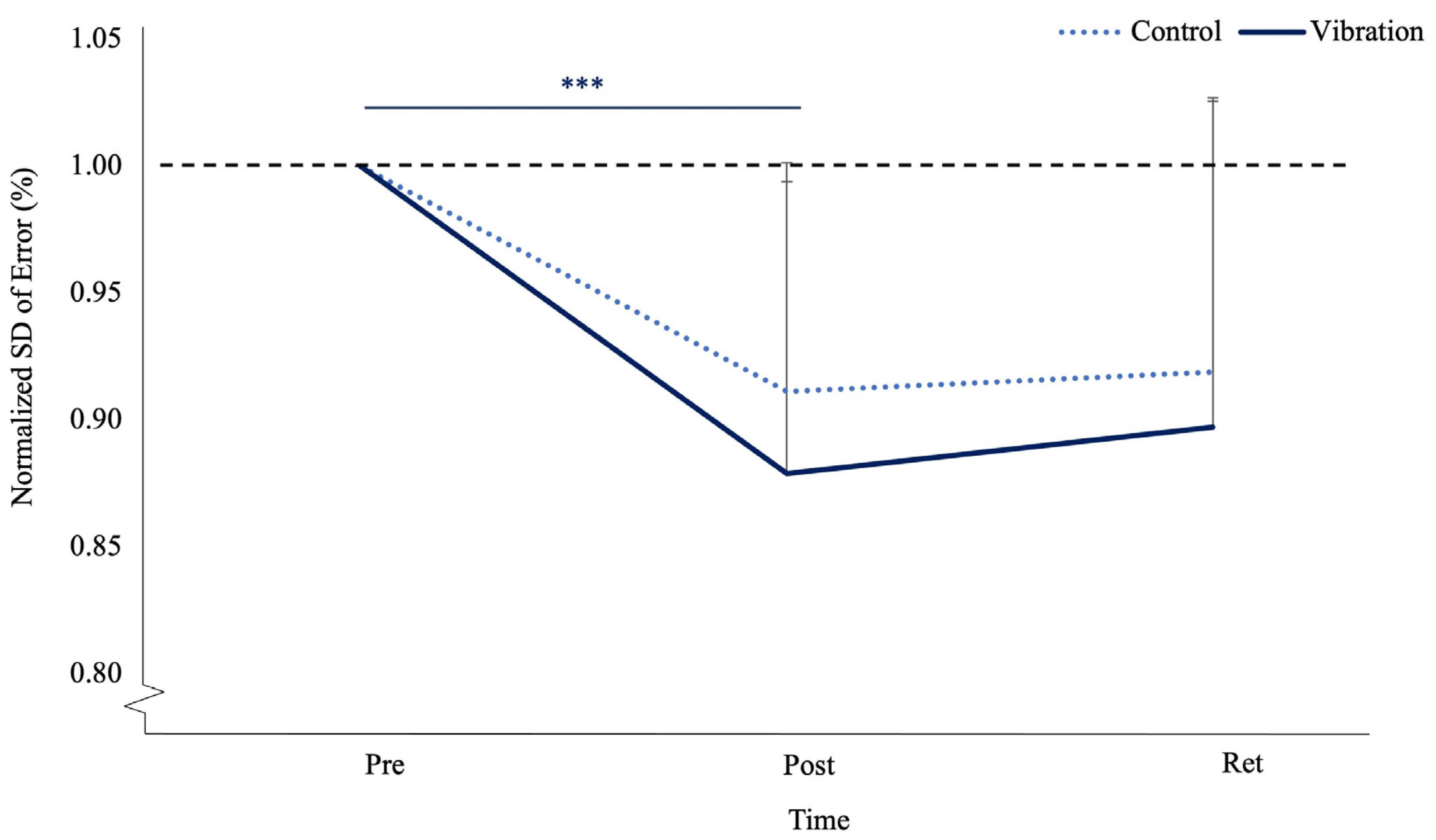
3.2. Motor Performance Accuracy
4. Discussion
4.1. SEPs
4.2. Motor Behavior
4.3. Real-World Application
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krajnak, K. Health effects associated with occupational exposure to hand-arm or whole body vibration. J. Toxicol. Environ. Health Part B 2018, 21, 320–334. [Google Scholar] [CrossRef]
- Holmes, N.P.; Spence, C. The body schema and multisensory representation (s) of peripersonal space. Cogn. Process. 2004, 5, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Blakemore, S.-J.; Frith, C.D.; Wolpert, D.M. The cerebellum is involved in predicting the sensory consequences of action. Neuroreport 2001, 12, 1879–1884. [Google Scholar] [CrossRef] [PubMed]
- Head, H.; Rivers, W.H.R.; Sherren, J.C.; Holmes, G.C.; Thompson, T.C.; Riddoch, G.C. Studies in Neurology. In Two Volumes, Vol. 1; Leopold Classic Library: South Yarra, VIC, Australia, 1920. [Google Scholar]
- Haggard, P.; Wolpert, D.M. Disorders of body scheme. In Higher-Order Motor Disorders; Freund, H.J., Jeannerod, M., Hallett, M., Leiguarda, R., Eds.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Nixon, P.D.; Passingham, R.E. Predicting sensory events. Exp. Brain Res. 2001, 138, 251–257. [Google Scholar] [CrossRef]
- Abbruzzese, G.; Berardelli, A. Sensorimotor integration in movement disorders. Mov. Disord. 2003, 18, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Albus, J.S. A theory of cerebellar function. Math. Biosci. 1971, 10, 25–61. [Google Scholar] [CrossRef]
- Gilbert, P.; Thach, W. Purkinje cell activity during motor learning. Brain Res. 1977, 128, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Passmore, S.R.; Murphy, B.; Lee, T.D. The origin, and application of somatosensory evoked potentials as a neurophysiological technique to investigate neuroplasticity. J. Can. Chiropr. Assoc. 2014, 58, 170. [Google Scholar]
- Waberski, T.D.; Buchner, H.; Perkuhn, M.; Gobbelé, R.; Wagner, M.; Kücker, W.; Silny, J. N30 and the effect of explorative finger movements: A model of the contribution of the motor cortex to early somatosensory potentials. Clin. Neurophysiol. 1999, 110, 1589–1600. [Google Scholar] [CrossRef]
- Britton, J.W.; Frey, L.C.; Hopp, J.L.; Korb, P.; Koubeissi, M.Z.; Lievens, W.E.; Pestana-Knight, E.M.; St Louis, E. Electroencephalography (EEG): An Introductory Text and Atlas of Normal and Abnormal Findings in Adults, Children, and Infants; American Epilepsy Society: Orlando, FL, USA, 2016. [Google Scholar]
- Andrew, D.; Yielder, P.; Haavik, H.; Murphy, B. The effects of subclinical neck pain on sensorimotor integration following a complex motor pursuit task. Exp. Brain Res. 2018, 236, 1–11. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Yielder, P.; Berkers, V.; Ambalavanar, U.; Holmes, M.; Murphy, B. Neck muscle fatigue impacts plasticity and sensorimotor integration in cerebellum and motor cortex in response to novel motor skill acquisition. J. Neurophysiol. 2020, 124, 844–855. [Google Scholar] [CrossRef]
- Guerraz, M.; Caudron, S.; Thomassin, N.; Blouin, J. Influence of head orientation on visually and memory-guided arm movements. Acta Psychol. 2011, 136, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Huysmans, M.A.; Hoozemans, M.J.; van der Beek, A.J.; de Looze, M.P.; van Dieen, J.H. Position sense acuity of the upper extremity and tracking performance in subjects with non-specific neck and upper extremity pain and healthy controls. J. Rehabil. Med. 2010, 42, 876–883. [Google Scholar] [CrossRef]
- Eklund, G. Position sense and state of contraction; the effects of vibration. J. Neurol. Neurosurg. Psychiatry 1972, 35, 606–611. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Holmes, M.W.; Murphy, B. Neck muscle fatigue alters upper limb proprioception. Exp. Brain Res. 2015, 233, 1663–1675. [Google Scholar] [CrossRef]
- Stanton, T.R.; Leake, H.B.; Chalmers, K.J.; Moseley, G.L. Evidence of impaired proprioception in chronic, idiopathic neck pain: Systematic review and meta-analysis. Phys. Ther. 2016, 96, 876–887. [Google Scholar] [CrossRef]
- Paulus, I.; Brumagne, S. Altered interpretation of neck proprioceptive signals in persons with subclinical recurrent neck pain. J. Rehabil. Med. 2008, 40, 426–432. [Google Scholar] [CrossRef]
- Lackner, J.R. Some proprioceptive influences on the perceptual representation of body shape and orientation. Brain 1988, 111, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.; Cordo, P.; Skoss, R.; Durrant, S.; Hodges, P. Illusory changes in head position induced by neck muscle vibration can alter the perception of elbow position. Behav. Neurosci. 2006, 120, 1211. [Google Scholar] [CrossRef] [PubMed]
- Daligadu, J.; Haavik, H.; Yielder, P.C.; Baarbe, J.; Murphy, B. Alterations in cortical and cerebellar motor processing in subclinical neck pain patients following spinal manipulation. J. Manip. Physiol. Ther. 2013, 36, 527–537. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Yielder, P.; Holmes, M.W.; Murphy, B. Neck muscle fatigue affects performance of an eye-hand tracking task. J. Electromyogr. Kinesiol. 2019, 47, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ambalavanar, U.; Delfa, N.L.; McCracken, H.; Zabihhosseinian, M.; Yielder, P.; Murphy, B. Differential changes in somatosensory evoked potentials and motor performance: Pursuit movement task versus force matching tracking task. J. Neurophysiol. 2022, 128, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Yielder, P.; Murphy, B. Do pursuit movement tasks lead to differential changes in early somatosensory evoked potentials related to motor learning compared with typing tasks? J. Neurophysiol. 2015, 113, 1156–1164. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Yielder, P.; Wise, R.; Holmes, M.; Murphy, B. Effect of Neck Muscle Fatigue on Hand Muscle Motor Performance and Early Somatosensory Evoked Potentials. Brain Sci. 2021, 11, 1481. [Google Scholar] [CrossRef]
- McCracken, H.; Murphy, B.; Ambalavanar, U.; Zabihhosseinian, M.; Yielder, P. Sensorimotor Integration and Motor Learning During a Novel Visuomotor Tracing Task in Young Adults with Attention-Deficit/Hyperactivity Disorder. J. Neurophysiol. 2023, 129, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.; Chandy, M.; Babu, K. Quantitative study of muscle spindles in suboccipital muscles of human foetuses. Neurol. India 2001, 49, 355. [Google Scholar]
- Cooper, S.; Daniel, P. Muscle spindles in man; their morphology in the lumbricals and the deep muscles of the neck. Brain 1963, 86, 563–586. [Google Scholar] [CrossRef]
- Peng, B.; Yang, L.; Li, Y.; Liu, T.; Liu, Y. Cervical Proprioception Impairment in Neck Pain-Pathophysiology, Clinical Evaluation, and Management: A Narrative Review. Pain Ther. 2021, 10, 143–164. [Google Scholar] [CrossRef]
- Richmond, F.; Abrahams, V. What are the proprioceptors of the neck? Prog. Brain Res. 1979, 50, 245–254. [Google Scholar]
- Ganguly, J.; Kulshreshtha, D.; Almotiri, M.; Jog, M. Muscle tone physiology and abnormalities. Toxins 2021, 13, 282. [Google Scholar] [CrossRef]
- Zabihhosseinian, M.; Holmes, M.W.; Howarth, S.; Ferguson, B.; Murphy, B. Neck muscle fatigue differentially alters scapular and humeral kinematics during humeral elevation in subclinical neck pain participants versus healthy controls. J. Electromyogr. Kinesiol. 2017, 33, 73–82. [Google Scholar] [CrossRef]
- Falla, D.; Farina, D. Neuromuscular adaptation in experimental and clinical neck pain. J. Electromyogr. Kinesiol. 2008, 18, 255–261. [Google Scholar] [CrossRef]
- Alcaraz-Clariana, S.; García-Luque, L.; Garrido-Castro, J.L.; Fernández-de-Las-Peñas, C.; Carmona-Pérez, C.; Rodrigues-de-Souza, D.P.; Alburquerque-Sendín, F. Paravertebral muscle mechanical properties and spinal range of motion in patients with acute neck or low back pain: A case-control study. Diagnostics 2021, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Schieppati, M.; Nardone, A.; Schmid, M. Neck muscle fatigue affects postural control in man. Neuroscience 2003, 121, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Hellsing, G. Distortion of mandibular kinesthesia induced by vibration of human jaw muscles. Eur. J. Oral Sci. 1978, 86, 486–494. [Google Scholar] [CrossRef]
- Tabbert, H.; Ambalavanar, U.; Murphy, B. Neck Muscle Vibration Alters Upper Limb Proprioception as Demonstrated by Changes in Accuracy and Precision during an Elbow Repositioning Task. Brain Sci. 2022, 12, 1532. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.M.; McCloskey, D.I.; Matthews, P.B. Proprioceptive illusions induced by muscle vibration: Contribution by muscle spindles to perception? Science 1972, 175, 1382–1384. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Engberg, I.; Matthews, P. The relative sensitivity to vibration of muscle receptors of the cat. J. Physiol. 1967, 192, 773–800. [Google Scholar] [CrossRef] [PubMed]
- Cordo, P.; Bevan, L.; Gurfinkel, V.; Carlton, L.; Carlton, M.; Kerr, G. Proprioceptive coordination of discrete movement sequences: Mechanism and generality. Can. J. Physiol. Pharmacol. 1995, 73, 305–315. [Google Scholar] [CrossRef]
- Pettorossi, V.E.; Panichi, R.; Botti, F.M.; Biscarini, A.; Filippi, G.M.; Schieppati, M. Long-lasting effects of neck muscle vibration and contraction on self-motion perception of vestibular origin. Clin. Neurophysiol. 2015, 126, 1886–1900. [Google Scholar]
- Mehrkanoon, S.; Boonstra, T.W.; Breakspear, M.; Hinder, M.; Summers, J.J. Upregulation of cortico-cerebellar functional connectivity after motor learning. Neuroimage 2016, 128, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Dal Maso, F.; Desormeau, B.; Boudrias, M.-H.; Roig, M. Acute cardiovascular exercise promotes functional changes in cortico-motor networks during the early stages of motor memory consolidation. Neuroimage 2018, 174, 380–392. [Google Scholar] [CrossRef]
- Pearce, A.J.; Kidgell, D.J. Comparison of corticomotor excitability during visuomotor dynamic and static tasks. J. Sci. Med. Sport 2010, 13, 167–171. [Google Scholar] [CrossRef]
- Brown, G.G.; Caligiuri, M.; Meloy, M.; Eberson, S.C.; Kindermann, S.S.; Frank, L.R.; Eyler Zorrilla, L.T.; Lohr, J.B. Functional brain asymmetries during visuomotor tracking. J. Clin. Exp. Neuropsychol. 2004, 26, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Vernon, H. The Neck Disability Index: State-of-the-art, 1991–2008. J. Manip. Physiol. Ther. 2008, 31, 491–502. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.S.; Murphy, B.A.; Glazebrook, C.M.; Burkitt, J.J.; Karellas, A.M.; Yielder, P.C. Audiovisual multisensory integration and evoked potentials in young adults with and without attention-deficit/hyperactivity disorder. Front. Hum. Neurosci. 2019, 13, 95. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.S.; Murphy, B.A.; Burkitt, J.J.; Glazebrook, C.M.; Yielder, P.C. Audiovisual Multisensory Processing in Young Adults with Attention-Deficit/Hyperactivity Disorder. Multisens. Res. 2020, 33, 599–623. [Google Scholar] [CrossRef] [PubMed]
- McCracken, H.S.; Murphy, B.A.; Ambalavanar, U.; Glazebrook, C.M.; Yielder, P.C. Source Localization of Audiovisual Multisensory Neural Generators in Young Adults with Attention-Deficit/Hyperactivity Disorder. Brain Sciences 2022, 12, 809. [Google Scholar] [CrossRef] [PubMed]
- Nuwer, M.R.; Aminoff, M.; Desmedt, J.; Eisen, A.A.; Goodin, D.; Matsuoka, S.; Mauguière, F.; Shibasaki, H.; Sutherling, W.; Vibert, J.-F. IFCN recommended standards for short latency somatosensory evoked potentials. Report of an IFCN committee. Electroencephalogr. Clin. Neurophysiol. 1994, 91, 6–11. [Google Scholar] [CrossRef]
- Haavik, H.; Murphy, B. An approach using fast rate stimulation to investigate changes in cerebellar processing. Clin. Chiropr. 2011, 14, 174. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics; Sparky House Publishing: Baltimore, MD, USA, 2009; Volume 2. [Google Scholar]
- Pallant, J. SPSS Survival Manual: A Step by Step Guide to Data Analysis Using IBM SPSS; McGraw-Hill Education: London, UK, 2020. [Google Scholar]
- Miles, J.; Shevlin, M. Applying Regression and Correlation: A Guide for Students and Researchers; Sage: Thousand Oaks, CA, USA, 2001. [Google Scholar]
- Tabbert, H.; Murphy, B. The effects of neck muscle vibration on short latency somatosensory evoked potentials. In Proceedings of the Southern Ontario Motor Behaviour Symposium (SOMBS) and Centre for Motor Control (CMC), Toronto, ON, Canada, 5 May 2023. [Google Scholar]
- Sonoo, M. Anatomic origin and clinical application of the widespread N18 potential in median nerve somatosensory evoked potentials. J. Clin. Neurophysiol. 2000, 17, 258–268. [Google Scholar] [CrossRef]
- Sonoo, M.; Genba, K.; Zai, W.; Iwata, M.; Mannen, T.; Kanazawa, I. Origin of the widespread N18 in median nerve SEP. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials Sect. 1992, 84, 418–425. [Google Scholar] [CrossRef]
- Miall, R.C.; Wolpert, D.M. Forward models for physiological motor control. Neural Netw. 1996, 9, 1265–1279. [Google Scholar] [CrossRef]
- Ambalavanar, U. Development and Validation of Tools to Examine Neural Correlates and Constructs Associated with Sensorimotor Integration; Ontario Tech University: Oshawa, ON, Canada, 2021. [Google Scholar]
- Felten, D.L.; O’Banion, M.K.; Maida, M.E. Netter’s Atlas of Neuroscience; Elsevier Health Sciences: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Noël, P.; Ozaki, I.; Desmedt, J.E. Origin of N18 and P14 far-fields of median nerve somatosensory evoked potentials studied in patients with a brain-stem lesion. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Allison, T.; McCarthy, G.; Wood, C.C. The relationship between human long-latency somatosensory evoked potentials recorded from the cortical surface and from the scalp. Electroencephalogr. Clin. Neurophysiol. Evoked Potentials Sect. 1992, 84, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Mauguiere, F.; Allison, T.; Babiloni, C.; Buchner, H.; Eisen, A.; Goodin, D.; Jones, S.; Kakigi, R.; Matsuoka, S.; Nuwer, M. Somatosensory evoked potentials. The International Federation of Clinical Neurophysiology. Electroencephalogr. Clin. Neurophysiol. 1999, 52, 79–90. [Google Scholar]
- Restuccia, D.; Valeriani, M.; Barba, C.; Le Pera, D.; Capecci, M.; Filippini, V.; Molinari, M. Functional changes of the primary somatosensory cortex in patients with unilateral cerebellar lesions. Brain 2001, 124, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Cruccu, G.; Aminoff, M.; Curio, G.; Guerit, J.; Kakigi, R.; Mauguiere, F.; Rossini, P.; Treede, R.-D.; Garcia-Larrea, L. Recommendations for the clinical use of somatosensory-evoked potentials. Clin. Neurophysiol. 2008, 119, 1705–1719. [Google Scholar] [CrossRef]
- Doyon, J.; Song, A.W.; Karni, A.; Lalonde, F.; Adams, M.M.; Ungerleider, L.G. Experience-dependent changes in cerebellar contributions to motor sequence learning. Proc. Natl. Acad. Sci. USA 2002, 99, 1017–1022. [Google Scholar] [CrossRef]
- Doyon, J.; Owen, A.M.; Petrides, M.; Sziklas, V.; Evans, A.C. Functional anatomy of visuomotor skill learning in human subjects examined with positron emission tomography. Eur. J. Neurosci. 1996, 8, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Doyon, J.; Ungerleider, L.G. Functional anatomy of motor skill learning. In Neuropsychology of Memory; The Guilford Press: New York, NY, USA, 2002; pp. 225–238. [Google Scholar]
- Doyon, J.; Penhune, V.; Ungerleider, L.G. Distinct contribution of the cortico-striatal and cortico-cerebellar systems to motor skill learning. Neuropsychologia 2003, 41, 252–262. [Google Scholar] [CrossRef]
- Macerollo, A.; Brown, M.J.; Kilner, J.M.; Chen, R. Neurophysiological changes measured using somatosensory evoked potentials. Trends Neurosci. 2018, 41, 294–310. [Google Scholar] [CrossRef]
- Fitts, P.M.; Posner, M.I. Human Performance; American Psychological Association: Washingtion, DC, USA, 1967. [Google Scholar]
- Schmidt, R.A.; Lee, T. Motor Learning and Control: A Behavioral Emphasis Champaign; Human Kinetics: Champaign, IL, USA, 1988. [Google Scholar]
- Shadmehr, R.; Holcomb, H.H. Neural correlates of motor memory consolidation. Science 1997, 277, 821–825. [Google Scholar] [CrossRef]
- Borragán, G.; Urbain, C.; Schmitz, R.; Mary, A.; Peigneux, P. Sleep and memory consolidation: Motor performance and proactive interference effects in sequence learning. Brain Cogn. 2015, 95, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Guadagnoli, M.A.; Lee, T.D. Challenge point: A framework for conceptualizing the effects of various practice conditions in motor learning. J. Mot. Behav. 2004, 36, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Akizuki, K.; Ohashi, Y. Measurement of functional task difficulty during motor learning: What level of difficulty corresponds to the optimal challenge point? Hum. Mov. Sci. 2015, 43, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.; Hunter, I.; Hopkins, T.; Feland, B. The short-term effect of whole body vibration training on sprint start performance in collegiate athletes. Int. J. Exerc. Sci. 2009, 2, 264. [Google Scholar] [PubMed]
- Fernandez-Rio, J.; Terrados, N.; Fernandez-Garcia, B.; Suman, O.E. Effects of vibration training on force production in female basketball players. J. Strength Cond. Res. 2010, 24, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Feltroni, L.; Monteleone, S.; Petrucci, L.; Carlisi, E.; Mazzacane, B.; Schieppati, M.; Dalla Toffola, E. Potentiation of muscle strength by focal vibratory stimulation on quadriceps femoris. G Ital. Med. Lav. Ergon. 2018, 40, 90–96. [Google Scholar]
- Filippi, G.; Fattorini, L.; Summa, A.; Zagaglia, A.; Rodio, A. Effects of focal vibration on power and work in multiple wingate tests. Biol. Sport 2020, 37, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Baarbé, J.K.; Yielder, P.; Haavik, H.; Holmes, M.W.; Murphy, B.A. Subclinical recurrent neck pain and its treatment impacts motor training-induced plasticity of the cerebellum and motor cortex. PLoS ONE 2018, 13, e0193413. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | |||
|---|---|---|---|
| Control | Vibration | ||
| Proportional Change in SEP Peak Amplitudes | |||
| N9 Peak Amplitude | 0.97 ± 0.10 | 1.03 ± 0.10 | |
| CI (95%) | Lower Bound | 0.91 | 0.97 |
| Upper Bound | 1.03 | 1.10 | |
| N11 Peak Amplitude | 1.18 ± 0.79 | 0.96 ± 0.33 | |
| CI (95%) | Lower Bound | 0.66 | 0.71 |
| Upper Bound | 1.61 | 1.12 | |
| N13 Peak Amplitude | 1.03 ± 0.31 | 1.06 ± 0.45 | |
| CI (95%) | Lower Bound | 0.83 | 0.72 |
| Upper Bound | 1.19 | 1.29 | |
| N18 Peak Amplitude | 1.59 ± 1.07 | 0.78 ± 0.22 | |
| CI (95%) | Lower Bound | 0.94 | 0.63 |
| Upper Bound | 2.24 | 1.94 | |
| N20 Peak Amplitude | 1.02 ± 0.29 | 1.21 ± 0.41 | |
| CI (95%) | Lower Bound | 0.84 | 0.95 |
| Upper Bound | 1.19 | 1.52 | |
| N24 Peak Amplitude | 0.86 ± 0.28 | 1.16 ± 0.35 | |
| CI (95%) | Lower Bound | 0.69 | 0.91 |
| Upper Bound | 1.03 | 1.40 | |
| P25 Peak Amplitude | 1.09 ± 0.24 | 1.19 ± 0.39 | |
| CI (95%) | Lower Bound | 0.95 | 0.93 |
| Upper Bound | 1.23 | 1.48 | |
| N30 Peak Amplitude | 1.07 ± 0.14 | 1.11 ± 0.28 | |
| CI (95%) | Lower Bound | 0.99 | 0.92 |
| Upper Bound | 1.16 | 1.31 | |
| N60 Peak Amplitude | 0.90 ± 0.35 | 1.00 ± 0.23 | |
| CI (95%) | Lower Bound | 0.69 | 0.83 |
| Upper Bound | 1.11 | 1.15 | |
| Time | |||
| Pre-Acquisition | Post-Acquisition | Retention | |
| Absolute Motor Performance Error (%) | |||
| Control Group | 0.679 ± 0.150 | 0.595 ± 0.115 | 0.592 ± 0.099 |
| 95% CI: Lower Bound | 0.62 | 0.55 | 0.55 |
| 95% CI: Upper Bound | 0.74 | 0.64 | 0.63 |
| Vibration Group | 0.673 ± 0.170 | 0.577 ± 0.097 | 0.581 ± 0.095 |
| 95% CI: Lower Bound | 0.59 | 0.54 | 0.55 |
| 95% CI: Upper Bound | 0.75 | 0.61 | 0.61 |
| Absolute SD of Motor Performance Error (%) | |||
| Control Group | 1.23 ± 0.117 | 1.11 ± 0.087 | 1.12 ± 0.101 |
| 95% CI: Lower Bound | 1.15 | 1.06 | 1.06 |
| 95% CI: Upper Bound | 1.30 | 1.16 | 1.18 |
| Vibration Group | 1.22 ± 0.191 | 1.05 ± 0.090 | 1.07 ± 0.078 |
| 95% CI: Lower Bound | 1.10 | 0.99 | 1.02 |
| 95% CI: Upper Bound | 1.34 | 1.11 | 1.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabbert, H.; Ambalavanar, U.; Murphy, B. Neck Muscle Vibration Alters Cerebellar Processing Associated with Motor Skill Acquisition of a Proprioceptive-Based Task. Brain Sci. 2023, 13, 1412. https://doi.org/10.3390/brainsci13101412
Tabbert H, Ambalavanar U, Murphy B. Neck Muscle Vibration Alters Cerebellar Processing Associated with Motor Skill Acquisition of a Proprioceptive-Based Task. Brain Sciences. 2023; 13(10):1412. https://doi.org/10.3390/brainsci13101412
Chicago/Turabian StyleTabbert, Hailey, Ushani Ambalavanar, and Bernadette Murphy. 2023. "Neck Muscle Vibration Alters Cerebellar Processing Associated with Motor Skill Acquisition of a Proprioceptive-Based Task" Brain Sciences 13, no. 10: 1412. https://doi.org/10.3390/brainsci13101412