
Using Neck Muscle Afferentation to Control an Ongoing Limb Movement? Individual Differences in the Influence of Brief Neck Vibration

Abstract
:1. Introduction
1.1. Neck Muscle Vibration Affects Pointing
1.2. Online Control of Voluntary Actions
2. Methodology
2.1. Participants
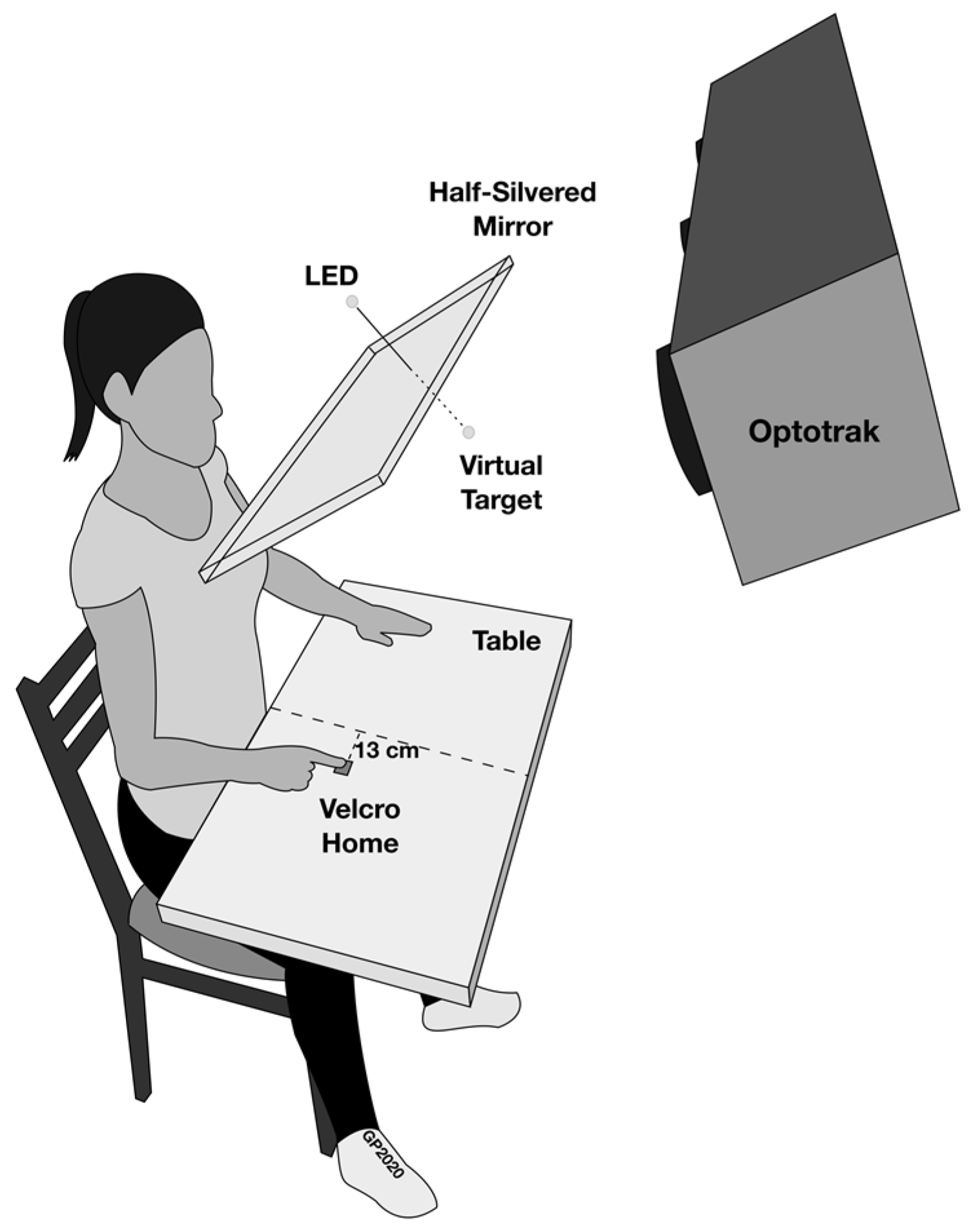
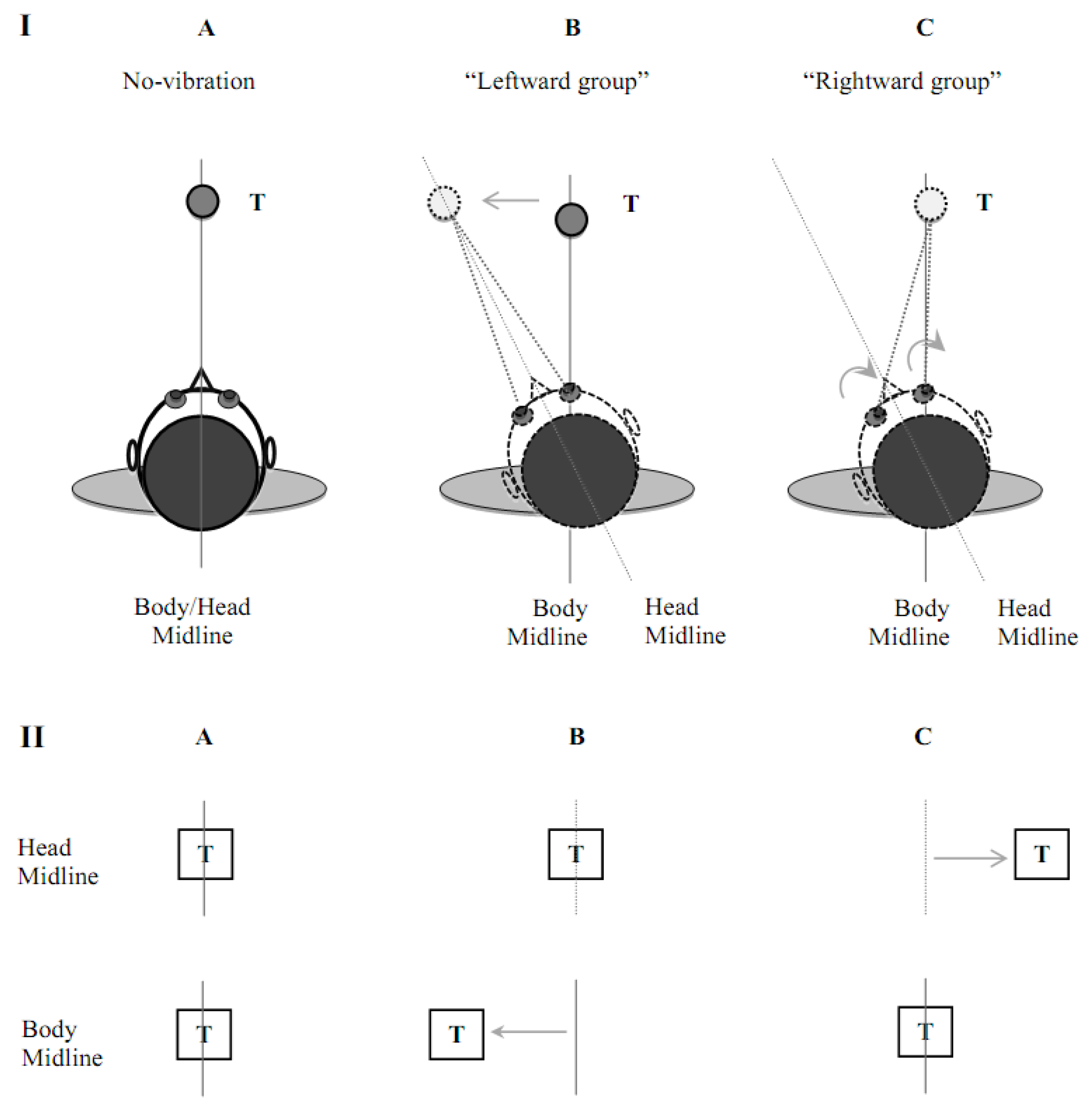
2.2. Apparatus
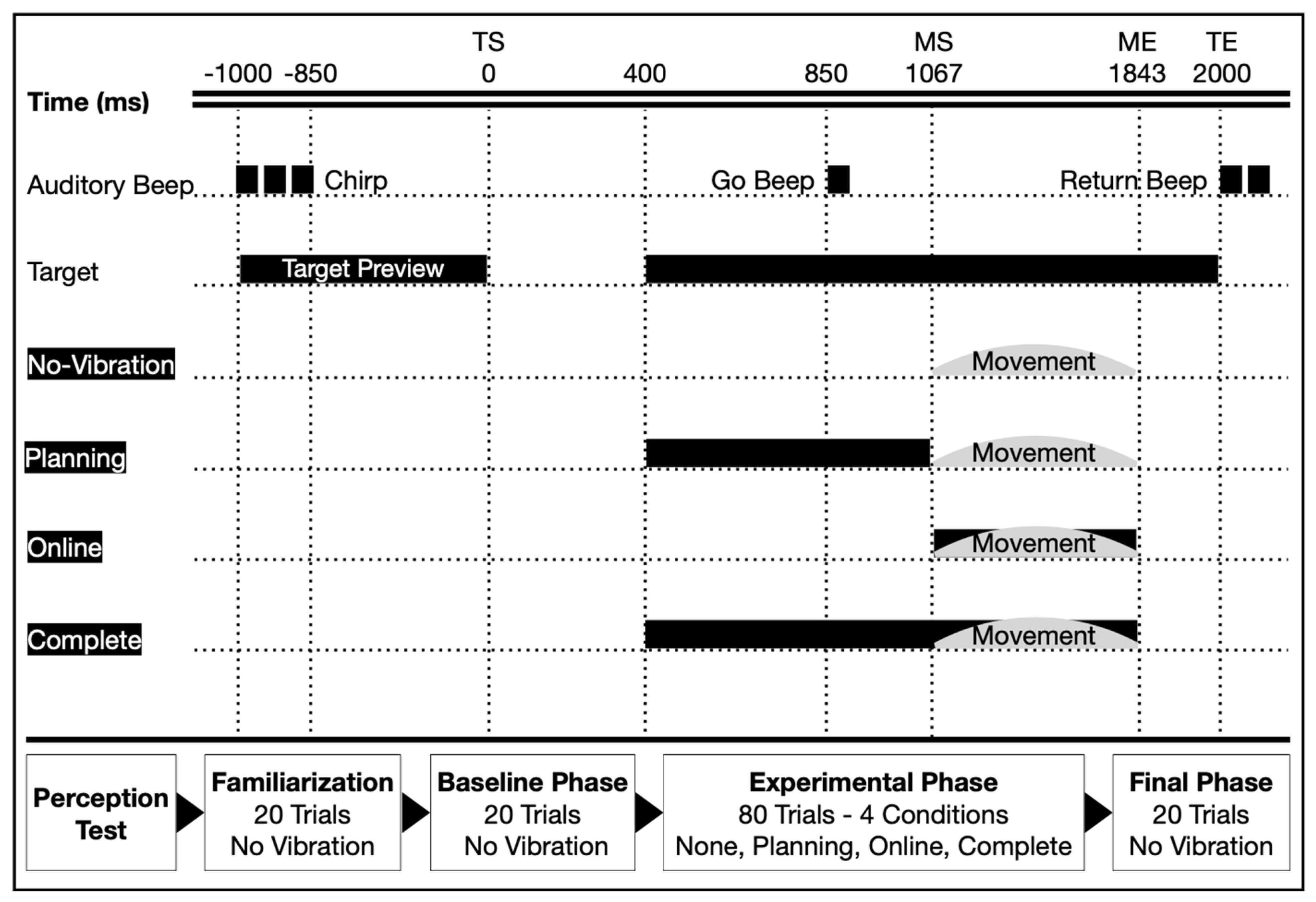
2.3. Experimental Procedure
2.4. Data Reduction and Analyses
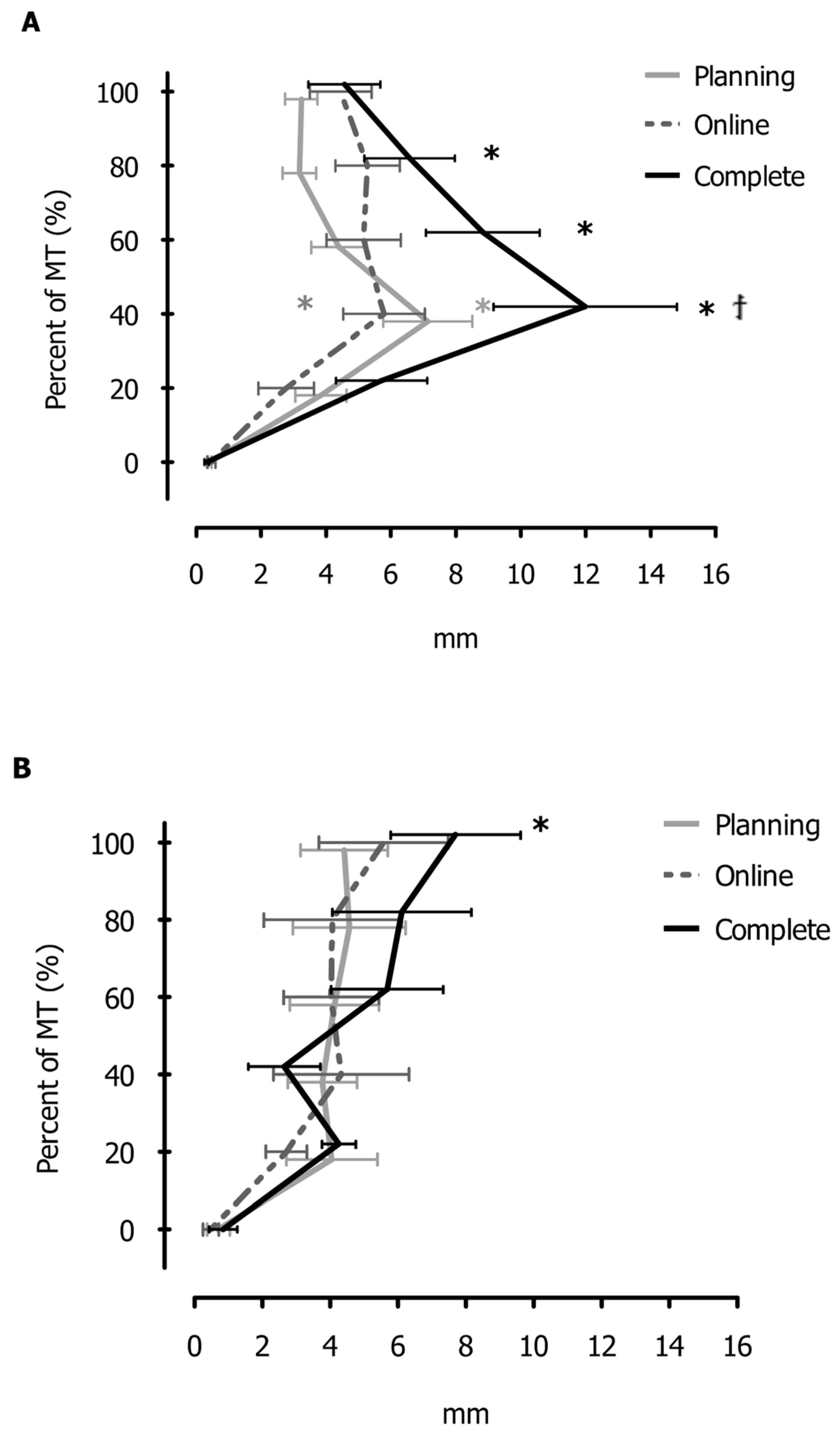
3. Results
4. Discussion
4.1. Neck Vibration and the Online Control of Reaching
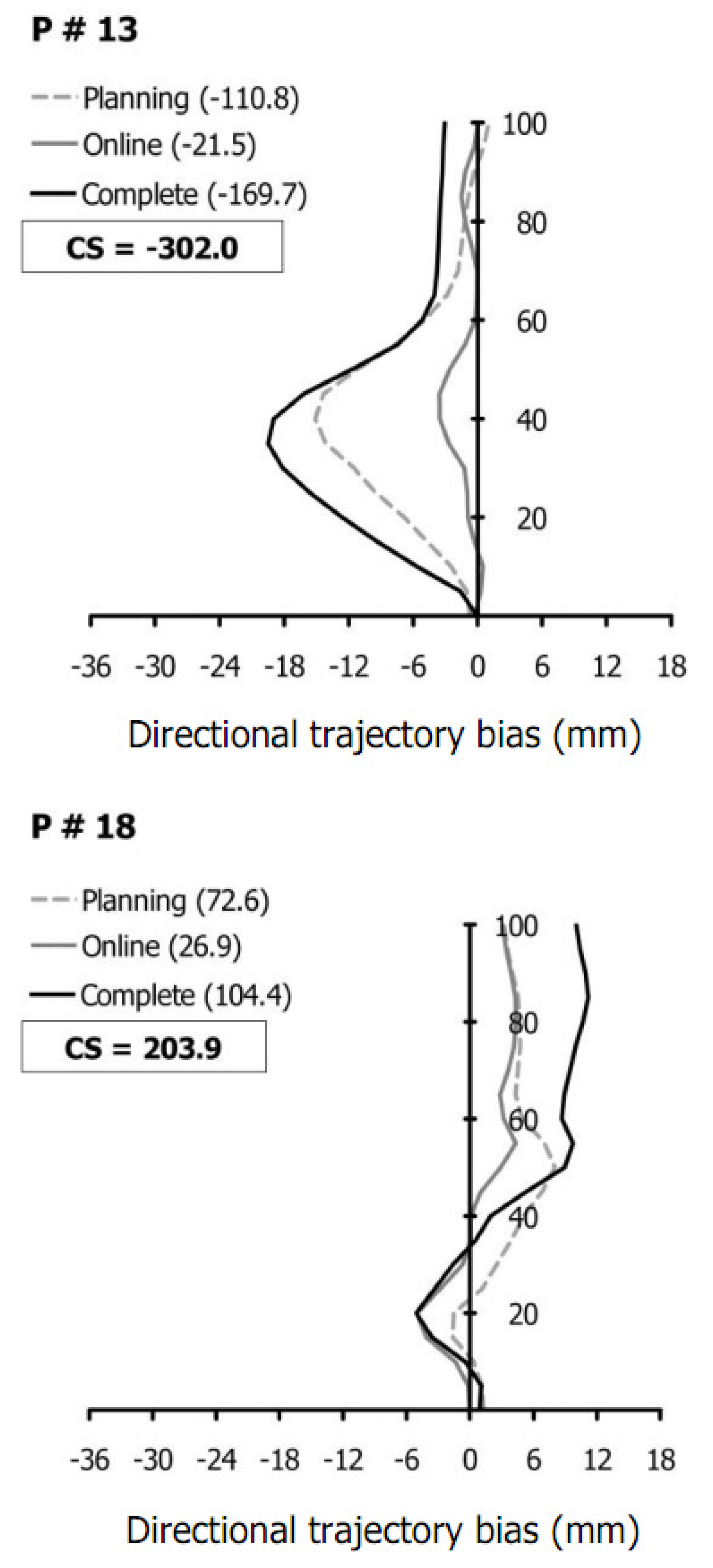
4.2. Using Limb Trajectories to Segregate Participants
4.3. Individual Differences in Prioritizing Sensory Cues
4.4. Different Patterns of Trajectory Biases and Sensory Prioritization
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrahams, V.C.; Richmond, F.J. Specialization of sensorimotor organization in the neck muscle system. Prog. Brain Res. 1988, 76, 125–135. [Google Scholar]
- Cohen, L.A. Role of eye and neck proprioceptive mechanisms in body orientation and motor coordination. J. Neurophysiol. 1961, 24, 1–11. [Google Scholar] [CrossRef]
- Corna, S.; Ito, Y.; von Brevern, M.; Bronstein, A.M.; Gresty, M.A. Reflex (unloading) and (defensive capitulation) responses in human neck muscle. J. Physiol. 1996, 496, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Corna, S.; von Brevern, M.; Bronstein, A.; Gresty, M. The functional effectiveness of neck muscle reflexes for head-righting in response to sudden fall. Exp. Brain Res. 1997, 117, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Bakker, D.A.; Richmond, F.J.R. Muscle Spindle Complexes in Muscles Around Upper Cervical Vertebrae in the Cat. J. Neurophysiol. 1982, 48, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.; Chandy, M.J.; Babu, K.S. Quantitative study of muscle spindles in suboccipital muscles of human foetuses. Neurol. India 2001, 49, 355–359. [Google Scholar]
- Bard, C.; Fleury, M.; Paillard, J. Different patterns in aiming accuracy for head-movers and non-head-movers. J. Hum. Mov. Stud. 1990, 18, 37–48. [Google Scholar]
- Flanders, M.; Tillery, S.I.H.; Soechting, J.F. Early stages in a sensorimotor transformation. Behav. Brain Sci. 1992, 15, 309–320. [Google Scholar] [CrossRef]
- Guerraz, M.; Navarro, J.; Ferrero, F.; Cremieux, J.; Blouin, J. Perceived versus actual head-on-trunk orientation during arm movement control. Exp. Brain Res. 2006, 172, 221–229. [Google Scholar] [CrossRef]
- Biguer, B.; Donaldson, I.M.; Hein, A.; Jeannerod, M. Neck muscle vibration modifies the representation of visual motion and direction in man. Brain 1988, 111, 1405–1424. [Google Scholar] [CrossRef]
- Mars, F.; Honoré, J.; Richard, C.; Coquery, J.M. Effects of all illusory orientation of the head on straight-ahead pointing movements. Cah. De Psychol. Cogn.-Curr. Psychol. Cogn. 1998, 17, 749–762. [Google Scholar]
- Guerraz, M.; Luyat, M.; Poquin, D.; Ohlmann, T. The role of neck afferents in subjective orientation in the visual and tactile sensory modalities. Acta Otolaryngol. 2000, 120, 735–738. [Google Scholar]
- Strupp, M.; Arbusow, V.; Dieterich, M.; Sautier, W.; Brandt, T. Perceptual and oculomotor effects of neck muscle vibration in vestibular neuritis. Ipsilateral somatosensory substitution of vestibular function. Brain 1998, 121, 677–685. [Google Scholar] [CrossRef]
- Blouin, J.; Okada, T.; Wolsley, C.; Bronstein, A. Encoding target-trunk relative position: Cervical versus vestibular contribution. Exp. Brain Res. 1998, 122, 101–107. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, S.; Seizova-Cajic, T. Neck muscle vibration in full cues affects pointing. J. Vis. 2007, 7, 1–8. [Google Scholar] [CrossRef]
- Guinet, M.; Michel, C. Prism adaptation and neck muscle vibration in healthy individuals: Are two methods better than one? Neuroscience 2003, 254, 443–451. [Google Scholar] [CrossRef]
- Han, Y.; Lennerstrand, G. Changes of visual localization induced by eye and neck muscle vibration in normal and strabismic subjects. Graefes Arch. Clin. Exp. Ophthalmol. 1999, 237, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Roll, R.; Velay, J.L.; Roll, J.P. Eye and neck proprioceptive messages contribute to the spatial coding of retinal input in visually oriented activities. Exp. Brain Res. 1991, 85, 423–431. [Google Scholar] [CrossRef]
- Goodwin, G.M.; McCloskey, D.I.; Matthews, P.B. The contribution of muscle afferents to kinesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain 1972, 95, 705–748. [Google Scholar] [CrossRef]
- Ernst, M.; Bülthoff, H. Merging the Senses into a Robust Percept. Trends Cogn. Sci. 2004, 8, 162–169. [Google Scholar] [CrossRef]
- Henriques, D.Y.; Klier, E.M.; Smith, M.A.; Lowy, D.; Crawford, J.D. Gaze-centered remapping of remembered visual space in an open-loop pointing task. J. Neurosci. 1998, 18, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Elliott, D.; Hansen, S.; Grierson, L.E.; Lyons, J.; Bennett, S.J.; Hayes, S.J. Goal-directed aiming: Two components but multiple processes. Psychol. Bull. 2010, 136, 1023–1044. [Google Scholar] [CrossRef] [PubMed]
- Elliott, D.; Helsen, W.F.; Chua, R. A century later: Woodworth’s (1899) two-component model of goal-directed aiming. Psychol. Bull. 2001, 127, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Keele, S.W. Movement control in skilled motor performance. Psychol. Bull. 1968, 70, 387–403. [Google Scholar] [CrossRef]
- Proteau, L.; Roujoula, A.; Messier, J. Evidence for continuous processing of visual information in a manual video-aiming task. J. Mot. Behav. 2009, 41, 219–231. [Google Scholar] [CrossRef]
- Saunders, J.A.; Knill, D.C. Humans use continuous visual feedback from the hand to control fast reaching movements. Exp. Brain Res. 2003, 152, 341–352. [Google Scholar] [CrossRef]
- Manson, G.A.; Blouin, J.; Kumawat, A.; Crainic, V.A.; Tremblay, L. Rapid online corrections for upper limb reaches to perturbed somatosensory targets: Evidence for non-visual sensorimotor transformation processes. Exp. Brain Res. 2019, 237, 839–853. [Google Scholar] [CrossRef]
- Bresciani, J.P.; Blouin, J.; Popov, K.; Bourdin, C.; Sarlegna, F.; Vercher, J.L.; Gauthier, G.M. Galvanic vestibular stimulation in humans produces online arm movement deviations when reaching towards memorized visual targets. Neurosci. Lett. 2002, 318, 34–38. [Google Scholar] [CrossRef]
- Mars, F.; Archambault, P.S.; Feldman, A.G. Vestibular contribution to combined arm and trunk motion. Exp. Brain Res. 2003, 150, 515–519. [Google Scholar] [CrossRef]
- Moreau-Debord, I.; Martin, C.Z.; Landry, M.; Green, A.M. Evidence for a reference frame transformation of vestibular signal contributions to voluntary reaching. J. Neurophysiol. 2014, 111, 1903–1919. [Google Scholar] [CrossRef]
- Smith, C.P.; Reynolds, R.F. Vestibular feedback maintains reaching accuracy during body movement. J. Physiol. 2017, 595, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Day, B.L.; Cauquil, A.S.; Bartolomei, L.; Pastor, M.A.; Lyon, I.N. Human body-segment tilts induced by galvanic stimulation: A vestibularly driven balance protection mechanism. J. Physiol. 1997, 500, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Salmoni, A.W.; Schmidt, R.A.; Walter, C.B. Knowledge of results and motor learning: A review and critical reappraisal. Psychol. Bull. 1984, 95, 355. [Google Scholar] [CrossRef]
- Knox, J.; Cordo, P.; Skoss, R.; Durrant, S.; Hodges, P. Illusory changes in head position induced by neck muscle vibration can alter the perception of elbow position. Behav. Neurosci. 2006, 120, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, M.; Andersson, G.; Gomez, S.; Johansson, R.; Martensson, A.; Karlberg, M.; Fransson, P.A. Cervical muscle afferents play a dominant role over vestibular afferents during bilateral vibration of neck muscles. J. Vestib. Res. 2006, 16, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; McCloskey, D.I. Illusions of head and visual target displacement induced by vibration of neck muscles. Brain 1991, 114, 755–759. [Google Scholar] [CrossRef]
- Fitts, P.M. The information capacity of the human motor system in controlling the amplitude of movment. J. Exp. Psychol. 1954, 47, 281–391. [Google Scholar] [CrossRef]
- Karnath, H.O.; Reich, E.; Rorden, C.; Fetter, M.; Driver, J. The perception of body orientation after neck-proprioceptive stimulation: Effects of time and of visual cueing. Exp. Brain Res. 2002, 143, 350–358. [Google Scholar] [CrossRef]
- Welch, R.B. Perceptual Modification: Adapting to Altered Sensory Environments; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Kornilova, L.N.; Sagalovitch, S.V.; Temnikova, V.V.; Yakushev, A.G. Static and dynamic vestibulo-cervico-ocular responses after prolonged exposure to microgravity. J. Vestib. Res. 2007, 17, 217–226. [Google Scholar] [CrossRef]
- Reschke, M.F.; Bloomberg, J.J.; Harm, D.L.; Paloski, W.H.; Layne, C.; McDonald, V. Posture, locomotion, spatial orientation, and motion sickness as a function of space flight. Brain Res. Rev. 1998, 28, 102–117. [Google Scholar] [CrossRef]
- Reinking, R.; Goldstein, G.; Houston, B.K. Cognitive style, proprioceptive skills, task set, stress, and the rod-and-frame test of field orientation. J. Personal. Soc. Psychol. 1974, 30, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Seizova-Cajic, T.; Sachtler, W.L.; Curthoys, I.S. Eye movements cannot explain vibration-induced visual motion and motion aftereffect. Exp. Brain Res. 2006, 173, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, L.; Elliott, D.; Starkes, J.L. Gender differences in the perception of self-orientation: Software or hardware? Perception 2004, 33, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Grierson, L.; Elliott, D. Goal-directed aiming and the relative contribution of two online control processes. Am. J. Psychol. 2009, 122, 309–324. [Google Scholar] [CrossRef]
- Chua, R.; Elliott, D. Visual regulation of manual aiming. Hum. Mov. Sci. 1993, 12, 365–401. [Google Scholar] [CrossRef]
- Elliott, D.; Carson, R.G.; Goodman, D.; Chua, R. Discrete vs continuous visual control of manual aiming. Hum. Mov. Sci. 1991, 10, 393–418. [Google Scholar] [CrossRef]
- Alekhina, M. The Role of Neck Muscles Afferentation in Planning and Online Control of Goal-Directed Movement. Master’s Thesis, University of Toronto, Toronto, ON, Canada, 2011. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant # /Condition | Planning | Online | Complete | CS | Group |
|---|---|---|---|---|---|
| P11 | −86.6 | −162.9 | −377.5 | −627.0 | Leftward |
| P16 | −125.3 | −69.6 | −188.7 | −383.6 | Leftward |
| P7 | −98.1 | −39.4 | −223.0 | −360.5 | Leftward |
| P1 | −114.6 | −41.3 | −151.2 | −307.1 | Leftward |
| P13 | −110.8 | −21.5 | −169.7 | −302.0 | Leftward |
| P10 | −5.4 | −125.6 | −96.2 | −227.2 | Leftward |
| P5 | −16.3 | −43.5 | −103.1 | −162.9 | Leftward |
| P14 | −69.1 | −30.5 | −61.4 | −161.0 | Leftward |
| P3 | −28.4 | −42.2 | −75.1 | −145.6 | Leftward |
| P9 | −115.7 | 89.8 | −115.2 | −141.1 | Leftward |
| P8 | −35.8 | −19.8 | −59.8 | −115.4 | Leftward |
| P2 | −62.7 | −14.1 | 8.3 | −68.5 | Leftward |
| P19 | 6.0 | −9.8 | 17.9 | 14.0 | Rightward |
| P4 | 26.9 | −1.4 | 47.6 | 73.1 | Rightward |
| P17 | 110.8 | −5.1 | 9.5 | 115.3 | Rightward |
| P15 | 124.2 | 1.8 | 68.1 | 194.0 | Rightward |
| P18 | 72.6 | 26.9 | 104.4 | 203.9 | Rightward |
| P6 | 51.6 | 138.7 | 70.2 | 260.4 | Rightward |
| P12 | 78.7 | 178.5 | 199.5 | 456.7 | Rightward |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekhina, M.; Perkic, G.; Manson, G.A.; Blouin, J.; Tremblay, L. Using Neck Muscle Afferentation to Control an Ongoing Limb Movement? Individual Differences in the Influence of Brief Neck Vibration. Brain Sci. 2023, 13, 1407. https://doi.org/10.3390/brainsci13101407
Alekhina M, Perkic G, Manson GA, Blouin J, Tremblay L. Using Neck Muscle Afferentation to Control an Ongoing Limb Movement? Individual Differences in the Influence of Brief Neck Vibration. Brain Sciences. 2023; 13(10):1407. https://doi.org/10.3390/brainsci13101407
Chicago/Turabian StyleAlekhina, Maria, Goran Perkic, Gerome Aleandro Manson, Jean Blouin, and Luc Tremblay. 2023. "Using Neck Muscle Afferentation to Control an Ongoing Limb Movement? Individual Differences in the Influence of Brief Neck Vibration" Brain Sciences 13, no. 10: 1407. https://doi.org/10.3390/brainsci13101407