
Imaging the Limbic System in Parkinson’s Disease—A Review of Limbic Pathology and Clinical Symptoms

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. What Is the Limbic System?
1.2. Imaging Changes
1.3. Aim of Review
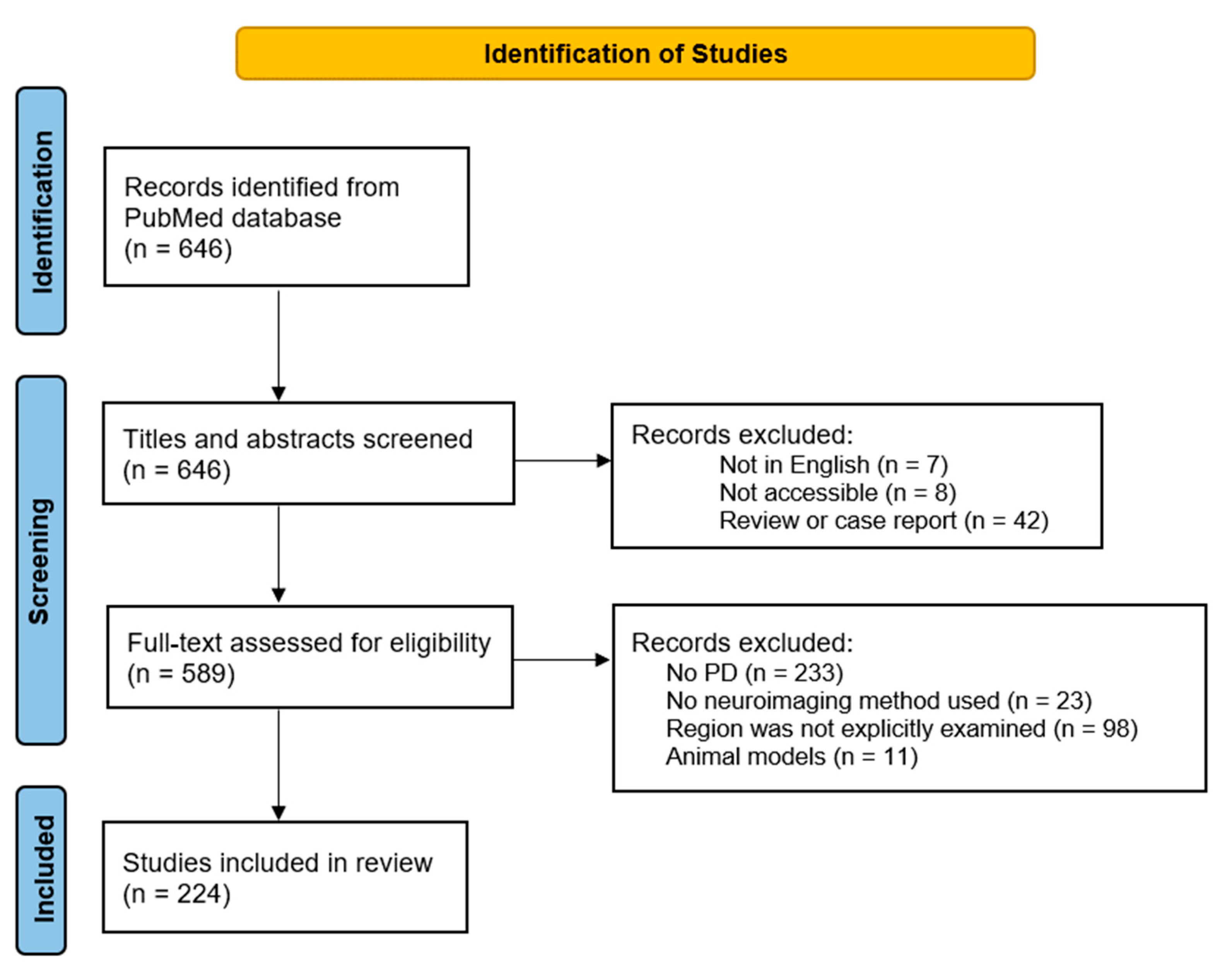
2. Methods
2.1. Literature Search and Selection Strategy
2.1.1. Inclusion Criteria
- English language;
- Parkinson’s disease;
- Use of neuroimaging methods (i.e., PET, MRI, SPECT);
- Human study.
2.1.2. Exclusion Criteria
- The limbic ROI was not explicitly examined in the study;
- Review paper or case report;
- Not accessible and not freely available.
2.2. Limbic Parkinson’s Disease Wordcloud
3. Results
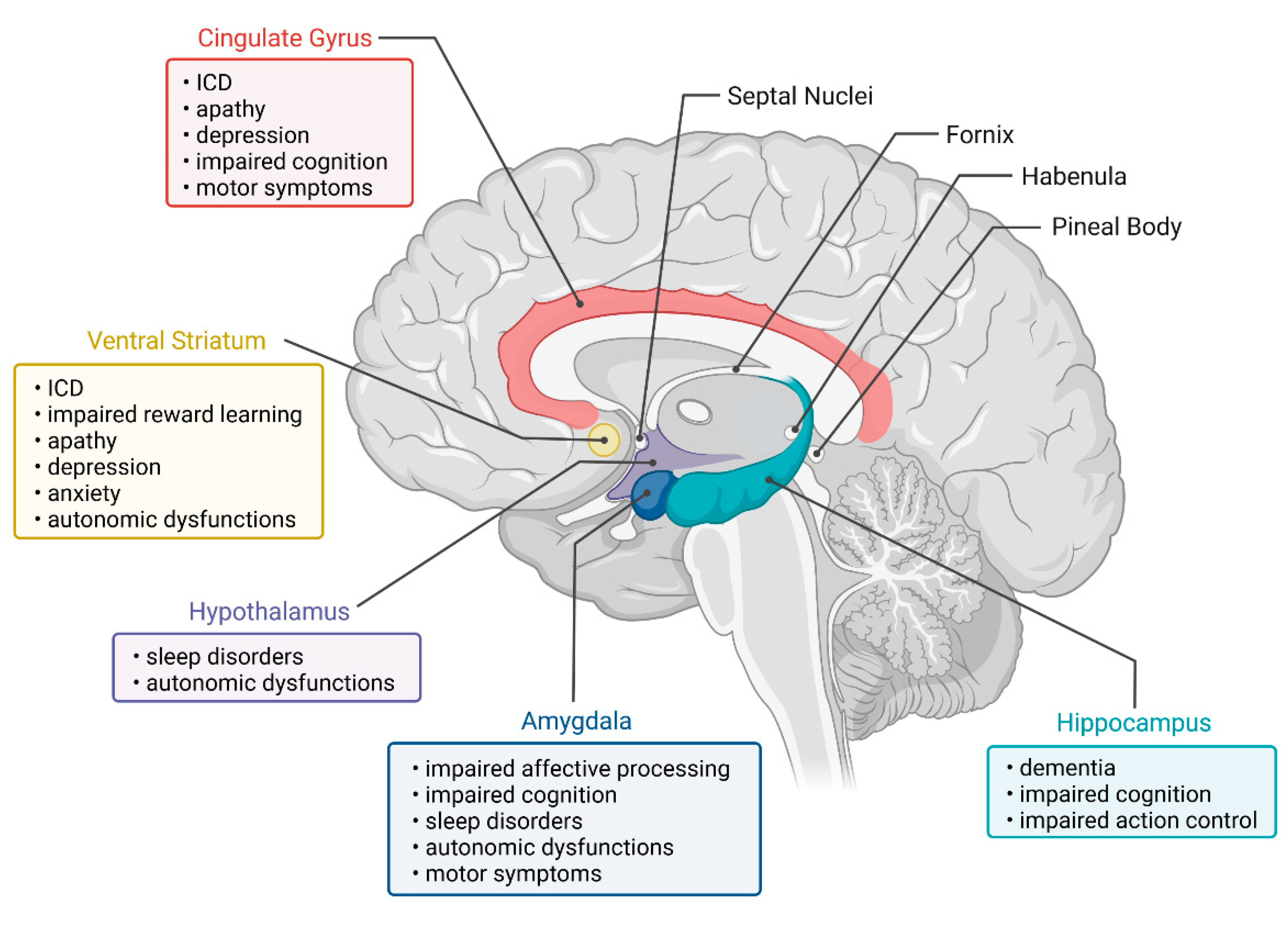
4. Regions of the Limbic System
4.1. Amygdala
4.1.1. Anatomy and Function
4.1.2. Pathology
4.1.3. Neuroimaging Evidence in PD
Neuropsychiatric Symptoms
Cognitive Symptoms
Motor and Other Symptoms
Conclusions
4.2. Hippocampus
4.2.1. Anatomy and Function
4.2.2. Pathology
4.2.3. Neuroimaging Evidence in PD
Cognitive Symptoms
Neuropsychiatric Symptoms
Motor and Other Symptoms
Conclusions
4.3. Hypothalamus
4.3.1. Anatomy and Function
4.3.2. Pathology
4.3.3. Neuroimaging Evidence in PD
Autonomous Symptoms and Sleep Disturbance
Conclusions
4.4. Cingulate Gyrus
4.4.1. Anatomy and Function
4.4.2. Pathology
4.4.3. Neuroimaging Evidence in PD
Neuropsychiatric Symptoms
Cognitive Symptoms
Motor and Other Symptoms
Conclusions
4.5. Substantia Innominata and Septal Nuclei
4.5.1. Anatomy and Function
4.5.2. Pathology
4.5.3. Neuroimaging Evidence in PD
Neuropsychiatric and Motor Symptoms
Cognitive Symptoms
4.6. Fornix and Mammillary Bodies
4.6.1. Anatomy and Function
4.6.2. Pathology
4.6.3. Neuroimaging Evidence in PD
4.7. Habenula and Pineal Body
4.7.1. Anatomy and Function
4.7.2. Pathology
4.7.3. Neuroimaging Evidence in PD
Neuropsychiatric Symptoms
4.8. Ventral Striatum
4.8.1. Anatomy and Function
4.8.2. Pathology
4.8.3. Neuroimaging Evidence in PD
Neuropsychiatric Symptoms
Cognitive and Other Symptoms
Conclusions
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dauer, W.; Przedborski, S. Parkinson’s Disease: Mechanisms and Models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- McGregor, M.M.; Nelson, A.B. Circuit Mechanisms of Parkinson’s Disease. Neuron 2019, 101, 1042–1056. [Google Scholar] [CrossRef] [PubMed]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson Disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, T.; Delong, M.R.; Guridi, J.; Obeso, J.A. Milestones in Research on the Pathophysiology of Parkinson’s Disease. Mov. Disord. 2011, 26, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.; Fox, S.H.; Lang, A.E. Overview of the Extranigral Aspects of Parkinson Disease. Arch. Neurol. 2009, 66, 167–172. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Pathoanatomy of Parkinson’s Disease. J. Neurol. 2000, 247, II3–II10. [Google Scholar] [CrossRef]
- Nelson, P.T.; Abner, E.L.; Patel, E.; Anderson, S.; Wilcock, D.M.; Kryscio, R.J.; van Eldik, L.J.; Jicha, G.A.; Gal, Z.; Nelson, R.S.; et al. The Amygdala as a Locus of Pathologic Misfolding in Neurodegenerative Diseases. J. Neuropathol. Exp. Neurol. 2018, 77, 2–20. [Google Scholar] [CrossRef]
- Braak, H.; Rüb, U.; Braak, E. Neuroanatomy of Parkinson Disease. Changes in the Neuronal Cytoskeleton of a Few Disease-Susceptible Types of Neurons Lead to Progressive Destruction of Circumscribed Areas in the Limbic and Motor Systems. Nervenarzt 2000, 71, 459–469. [Google Scholar] [CrossRef]
- Hawkes, C.H.; del Tredici, K.; Braak, H. A Timeline for Parkinson’s Disease. Parkinsonism Relat. Disord. 2010, 16, 79–84. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E.; Yilmazer, D.; de Vos, R.A.I.; Jansen, E.N.H.; Bohl, J.; Jellinger, K. Amygdala Pathology in Parkinson’s Disease. Acta Neuropathol. 1994, 88, 493–500. [Google Scholar] [CrossRef]
- Rolls, E.T. Limbic Systems for Emotion and for Memory, but No Single Limbic System. Cortex 2015, 62, 119–157. [Google Scholar] [CrossRef]
- Willis, T. Cerebri Anatome: Cui Accessit. Nervorum Descriptio et Usus; apud Gerbrandum Schagen; Forgotten Books: London, UK, 1973; Volume 1. [Google Scholar]
- Broca, P. Le Grand Lobe Limbique et La Scissure Limbique Dans La Série Des Mammifères. Rev. D’anthropologie 1878, 2, 385–598. [Google Scholar]
- Roxo, M.R.; Franceschini, P.R.; Zubaran, C.; Kleber, F.D.; Sander, J.W. The Limbic System Conception and Its Historical Evolution. Science 2011, 11, 2427–2440. [Google Scholar] [CrossRef]
- Kötter, R.; Meyer, N. The Limbic System: A Review of Its Empirical Foundation. Behav. Brain Res. 1992, 52, 105–127. [Google Scholar] [CrossRef]
- Rajmohan, V.; Mohandas, E. The Limbic System. Indian J. Psychiatry 2007, 49, 132. [Google Scholar] [CrossRef]
- Mark, L.P.; Daniels, D.L.; Naidich, T.P.; Hendrix, L.E. Limbic Connections. AJNR Am. J. Neuroradiol. 1995, 16, 1303. [Google Scholar]
- Mogenson, G.J.; Jones, D.L.; Yim, C.Y. From Motivation to Action: Functional Interface between the Limbic System and the Motor System. Prog. Neurobiol. 1980, 14, 69–97. [Google Scholar] [CrossRef]
- Sokolowski, K.; Corbin, J.G. Wired for Behaviors: From Development to Function of Innate Limbic System Circuitry. Front. Mol. Neurosci. 2012, 5, 55. [Google Scholar] [CrossRef]
- Giguère, N.; Nanni, S.B.; Trudeau, L.E. On Cell Loss and Selective Vulnerability of Neuronal Populations in Parkinson’s Disease. Front. Neurol. 2018, 9, 455. [Google Scholar] [CrossRef]
- Pavese, N.; Brooks, D.J. Imaging Neurodegeneration in Parkinson’s Disease. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2009, 1792, 722–729. [Google Scholar] [CrossRef]
- Moustafa, A.A.; Chakravarthy, S.; Phillips, J.R.; Gupta, A.; Keri, S.; Polner, B.; Frank, M.J.; Jahanshahi, M. Motor Symptoms in Parkinson’s Disease: A Unified Framework. Neurosci. Biobehav. Rev. 2016, 68, 727–740. [Google Scholar] [CrossRef]
- Kessler, R.M. Buitelaar Imaging Methods for Evaluating Brain Function in Man. Neurobiol. Aging 2003, 24, S21–S35. [Google Scholar] [CrossRef]
- LeDoux, J. The Amygdala. Curr. Biol. 2007, 17. [Google Scholar] [CrossRef] [PubMed]
- Swanson, L.W.; Petrovich, G.D. What Is the Amygdala? Trends Neurosci. 1998, 21, 323–331. [Google Scholar] [CrossRef]
- Janak, P.H.; Tye, K.M. From Circuits to Behaviour in the Amygdala. Nature 2015, 517, 284. [Google Scholar] [CrossRef]
- Balleine, B.W.; Killcross, S. Parallel Incentive Processing: An Integrated View of Amygdala Function. Trends Neurosci. 2006, 29, 272–279. [Google Scholar] [CrossRef]
- Braak, H.; del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Vasconcellos, L.F.; Pereira, J.S.; Adachi, M.; Greca, D.; Cruz, M.; Malak, A.L.; Charchat-Fichman, H. Volumetric Brain Analysis as a Predictor of a Worse Cognitive Outcome in Parkinson’s Disease. J. Psychiatr. Res. 2018, 102, 254–260. [Google Scholar] [CrossRef]
- Tinaz, S.; Kamel, S.; Aravala, S.S.; Sezgin, M.; Elfil, M.; Sinha, R. Distinct Neural Circuits Are Associated with Subclinical Neuropsychiatric Symptoms in Parkinson’s Disease. J. Neurol. Sci. 2021, 423, 117365. [Google Scholar] [CrossRef]
- Thaler, A.; Kliper, E.; Maidan, I.; Herman, T.; Rosenberg-Katz, K.; Bregman, N.; Gurevich, T.; Shiner, T.; Hausdorff, J.M.; Orr-Urtreger, A.; et al. Cerebral Imaging Markers of GBA and LRRK2 Related Parkinson’s Disease and Their First-Degree Unaffected Relatives. Brain. Topogr. 2018, 31, 1029–1036. [Google Scholar] [CrossRef]
- Banwinkler, M.; Dzialas, V.; Hoenig, M.C.; van Eimeren, T. Gray Matter Volume Loss in Proposed Brain-First and Body-First Parkinson’s Disease Subtypes. Mov. Disord. 2022. [Google Scholar] [CrossRef] [PubMed]
- Laansma, M.A.; Bright, J.K.; Al-Bachari, S.; Anderson, T.J.; Ard, T.; Assogna, F.; Baquero, K.A.; Berendse, H.W.; Blair, J.; Cendes, F.; et al. International Multicenter Analysis of Brain Structure Across Clinical Stages of Parkinson’s Disease. Mov. Disord. 2021, 36, 2583–2594. [Google Scholar] [CrossRef]
- Betrouni, N.; Lopes, R.; Defebvre, L.; Leentjens, A.F.G.; Dujardin, K. Texture Features of Magnetic Resonance Images: A Marker of Slight Cognitive Deficits in Parkinson’s Disease. Mov. Disord. 2020, 35, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Leocadi, M.; Canu, E.; Donzuso, G.; Stojkovic, T.; Basaia, S.; Kresojević, N.; Stankovic, I.; Sarasso, E.; Piramide, N.; Tomic, A.; et al. Longitudinal Clinical, Cognitive, and Neuroanatomical Changes over 5 Years in GBA-Positive Parkinson’s Disease Patients. J. Neurol. 2022, 269, 1485–1500. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Niccolini, F.; Pellicano, C.; Politis, M. Cortical Thinning across Parkinson’s Disease Stages and Clinical Correlates. J. Neurol. Sci. 2019, 398, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Lee, P.L.; Lu, C.H.; Lin, C.P.; Chou, K.H. Linking Stage-Specific Plasma Biomarkers to Gray Matter Atrophy in Parkinson Disease. AJNR Am. J. Neuroradiol. 2021, 42, 1444–1451. [Google Scholar] [CrossRef]
- He, H.; Liang, L.; Tang, T.; Luo, J.; Wang, Y.; Cui, H. Progressive Brain Changes in Parkinson’s Disease: A Meta-Analysis of Structural Magnetic Resonance Imaging Studies. Brain Res. 2020, 1740. [Google Scholar] [CrossRef]
- Bell, P.T.; Gilat, M.; Shine, J.M.; McMahon, K.L.; Lewis, S.J.G.; Copland, D.A. Neural Correlates of Emotional Valence Processing in Parkinson’s Disease: Dysfunction in the Subcortex. Brain Imaging Behav. 2019, 13, 189–199. [Google Scholar] [CrossRef]
- Criaud, M.; Kim, J.H.; Zurowski, M.; Lobaugh, N.; Chavez, S.; Houle, S.; Strafella, A.P. Anxiety in Parkinson’s Disease: Abnormal Resting Activity and Connectivity. Brain Res. 2021, 1753. [Google Scholar] [CrossRef]
- Knolle, F.; Garofalo, S.; Viviani, R.; Justicia, A.; Ermakova, A.O.; Blank, H.; Williams, G.B.; Arrondo, G.; Ramachandra, P.; Tudor-Sfetea, C.; et al. Altered Subcortical Emotional Salience Processing Differentiates Parkinson’s Patients with and without Psychotic Symptoms. Neuroimage Clin. 2020, 27, 102277. [Google Scholar] [CrossRef]
- Oosterwijk, C.S.; Vriend, C.; Berendse, H.W.; van der Werf, Y.D.; van den Heuvel, O.A. Anxiety in Parkinson’s Disease Is Associated with Reduced Structural Covariance of the Striatum. J. Affect Disord. 2018, 240, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qiu, Y.; Luo, Y.; Xu, P.; Li, Z.; Zhu, W.; Wu, Q.; Tao, W.; Guan, Q.; Chen, F. The Relationship of Anxious and Depressive Symptoms in Parkinson’s Disease with Voxel-Based Neuroanatomical and Functional Connectivity Measures. J. Affect Disord. 2019, 245, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Carey, G.; Lopes, R.; Viard, R.; Betrouni, N.; Kuchcinski, G.; Devignes, Q.; Defebvre, L.; Leentjens, A.F.G.; Dujardin, K. Anxiety in Parkinson’s Disease Is Associated with Changes in the Brain Fear Circuit. Parkinsonism Relat. Disord. 2020, 80, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Boccalini, C.; Carli, G.; Pilotto, A.; Padovani, A.; Perani, D. Gender Differences in Dopaminergic System Dysfunction in de Novo Parkinson’s Disease Clinical Subtypes. Neurobiol. Dis. 2022, 167, 105668. [Google Scholar] [CrossRef]
- Lin, H.; Cai, X.; Zhang, D.; Liu, J.; Na, P.; Li, W. Functional Connectivity Markers of Depression in Advanced Parkinson’s Disease. Neuroimage Clin. 2020, 25, 102130. [Google Scholar] [CrossRef]
- Fu, J.F.; Klyuzhin, I.; Liu, S.; Shahinfard, E.; Vafai, N.; McKenzie, J.; Neilson, N.; Mabrouk, R.; Sacheli, M.A.; Wile, D.; et al. Investigation of Serotonergic Parkinson’s Disease-Related Covariance Pattern Using [11C]-DASB/PET. Neuroimage Clin. 2018, 19, 652–660. [Google Scholar] [CrossRef]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Koros, C.; Yousaf, T.; Picillo, M.; Polychronis, S.; Simitsi, A.; Giordano, B.; Chappell, Z.; et al. Serotonergic Pathology and Disease Burden in the Premotor and Motor Phase of A53T α-Synuclein Parkinsonism: A Cross-Sectional Study. Lancet Neurol. 2019, 18, 748–759. [Google Scholar] [CrossRef]
- Nicastro, N.; Garibotto, V.; Burkhard, P.R. Extrastriatal 123 I-FP-CIT SPECT Impairment in Parkinson’s Disease—The PPMI Cohort. BMC Neurol. 2020, 20, 192. [Google Scholar] [CrossRef]
- Kang, Y.; Henchcliffe, C.; Verma, A.; Vallabhajosula, S.; He, B.; Kothari, P.J.; Pryor, K.O.; Mozley, P.D. 18F-FPEB PET/CT Shows MGluR5 Upregulation in Parkinson’s Disease. J. Neuroimaging 2019, 29, 97–103. [Google Scholar] [CrossRef]
- Marín-Lahoz, J.; Sampedro, F.; Horta-Barba, A.; Martínez-Horta, S.; Aracil-Bolaños, I.; Camacho, V.; Bejr-kasem, H.; Pascual-Sedano, B.; Pérez-Pérez, J.; Gironell, A.; et al. Preservation of Brain Metabolism in Recently Diagnosed Parkinson’s Impulse Control Disorders. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 2165–2174. [Google Scholar] [CrossRef]
- Stark, A.J.; Smith, C.T.; Petersen, K.J.; Trujillo, P.; van Wouwe, N.C.; Donahue, M.J.; Kessler, R.M.; Deutch, A.Y.; Zald, D.H.; Claassen, D.O. [18 F]Fallypride Characterization of Striatal and Extrastriatal D 2/3 Receptors in Parkinson’s Disease. Neuroimage Clin. 2018, 18, 433–442. [Google Scholar] [CrossRef] [PubMed]
- De Schipper, L.J.; Hafkemeijer, A.; van der Grond, J.; Marinus, J.; Henselmans, J.M.L.; van Hilten, J.J. Regional Structural Hippocampal Differences between Dementia with Lewy Bodies and Parkinson’s Disease. J. Parkinsons Dis. 2019, 9, 775–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.S.; Chen, H.L.; Lu, C.H.; Lee, C.Y.; Chou, K.H.; Chen, M.H.; Yu, C.C.; Lai, Y.R.; Chiang, P.L.; Lin, W.C. The Corticolimbic Structural Covariance Network as an Early Predictive Biosignature for Cognitive Impairment in Parkinson’s Disease. Sci. Rep. 2021, 11, 862. [Google Scholar] [CrossRef]
- Kim, J.; Zhang, K.; Cai, W.; YorkWilliams, S.; Ua Cruadhlaoich, M.A.I.; Llanes, S.; Menon, V.; Poston, K.L. Dopamine-Related Dissociation of Cortical and Subcortical Brain Activations in Cognitively Unimpaired Parkinson’s Disease Patients OFF and ON Medications. Neuropsychologia 2018, 119, 24–33. [Google Scholar] [CrossRef]
- Liu, A.; Lin, S.J.; Mi, T.; Chen, X.; Chan, P.; Wang, Z.J.; McKeown, M.J. Decreased Subregional Specificity of the Putamen in Parkinson’s Disease Revealed by Dynamic Connectivity-Derived Parcellation. Neuroimage Clin. 2018, 20, 1163–1175. [Google Scholar] [CrossRef]
- Wang, E.W.; Du, G.; Lewis, M.M.; Lee, E.Y.; de Jesus, S.; Kanekar, S.; Kong, L.; Huang, X. Multimodal MRI Evaluation of Parkinsonian Limbic Pathologies. Neurobiol. Aging 2019, 76, 194–200. [Google Scholar] [CrossRef]
- Hou, Y.; Wei, Q.; Ou, R.; Yang, J.; Song, W.; Gong, Q.; Shang, H. Impaired Topographic Organization in Cognitively Unimpaired Drug-Naïve Patients with Rigidity-Dominant Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 56, 52–57. [Google Scholar] [CrossRef]
- Wu, Q.; Gao, Y.; Liu, A.S.; Xie, L.Z.; Qian, L.; Yang, X.G. Large-Scale Cortical Volume Correlation Networks Reveal Disrupted Small World Patterns in Parkinson’s Disease. Neurosci. Lett. 2018, 662, 374–380. [Google Scholar] [CrossRef]
- Barrett, M.J.; Murphy, J.M.; Zhang, J.; Blair, J.C.; Flanigan, J.L.; Nawaz, H.; Dalrymple, W.A.; Sperling, S.A.; Patrie, J.; Druzgal, T.J. Olfaction, Cholinergic Basal Forebrain Degeneration, and Cognition in Early Parkinson Disease. Parkinsonism Relat. Disord. 2021, 90, 27–32. [Google Scholar] [CrossRef]
- Kim, M.; Yoo, S.; Kim, D.; Cho, J.W.; Kim, J.S.; Ahn, J.H.; Mun, J.K.; Choi, I.; Lee, S.K.; Youn, J. Extra-Basal Ganglia Iron Content and Non-Motor Symptoms in Drug-Naïve, Early Parkinson’s Disease. Neurol. Sci. 2021, 42, 5297–5304. [Google Scholar] [CrossRef]
- Guo, T.; Wu, J.; Zhou, C.; Guan, X.; Gao, T.; Bai, X.; Song, Z.; Xuan, M.; Gu, Q.; Huang, P.; et al. Aberrant Fiber Coherence of Amygdala-Accumbens-Pallidum Pathway Is Associated with Disorganized Nigrostriatal-Nigropallidal Pathway in Parkinson’s Disease. J. Magn. Reson. Imaging 2020, 52, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.; van Wouwe, N.; Stark, A.; Lin, Y.C.; Kang, H.; Trujillo-Diaz, P.; Kessler, R.; Zald, D.; Donahue, M.J.; Claassen, D.O. Ventral Striatal Network Connectivity Reflects Reward Learning and Behavior in Patients with Parkinson’s Disease. Hum. Brain Mapp. 2018, 39, 509–521. [Google Scholar] [CrossRef] [Green Version]
- Tuovinen, N.; Seppi, K.; de Pasquale, F.; Müller, C.; Nocker, M.; Schocke, M.; Gizewski, E.R.; Kremser, C.; Wenning, G.K.; Poewe, W.; et al. The Reorganization of Functional Architecture in the Early-Stages of Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 50, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, C.; Abbasi, N.; Zeighami, Y.; Yau, Y.; Dadar, M.; Rahayel, S.; Dagher, A. Sex Effects on Brain Structure in de Novo Parkinson’s Disease: A Multimodal Neuroimaging Study. Brain 2020, 143, 3052–3066. [Google Scholar] [CrossRef]
- Gilat, M.; Ehgoetz Martens, K.A.; Miranda-Domínguez, O.; Arpan, I.; Shine, J.M.; Mancini, M.; Fair, D.A.; Lewis, S.J.G.; Horak, F.B. Dysfunctional Limbic Circuitry Underlying Freezing of Gait in Parkinson’s Disease. Neuroscience 2018, 374, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, N.; Watanabe, H.; Kawabata, K.; Bagarinao, E.; Hara, K.; Tsuboi, T.; Tanaka, Y.; Ohdake, R.; Imai, K.; Masuda, M.; et al. Severe Hyposmia and Aberrant Functional Connectivity in Cognitively Normal Parkinson’s Disease. PLoS ONE 2018, 13, e0190072. [Google Scholar] [CrossRef] [PubMed]
- Mann, L.G.; Hay, K.R.; Song, A.K.; Errington, S.P.; Trujillo, P.; Zald, D.H.; Yan, Y.; Kang, H.; Logan, G.D.; Claassen, D.O. D 2-Like Receptor Expression in the Hippocampus and Amygdala Informs Performance on the Stop-Signal Task in Parkinson’s Disease. J. Neurosci. 2021, 41, 10023–10030. [Google Scholar] [CrossRef]
- Knierim, J.J. The Hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef]
- Anand, K.; Dhikav, V. Hippocampus in Health and Disease: An Overview. Ann. Indian Acad. Neurol. 2012, 15, 239. [Google Scholar] [CrossRef]
- Scoville, W.B.; Milner, B. Loss of Recent Memory after Bilateral Hippocampal Lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, 11. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Otto, T.; Cohen, N.J. The Hippocampus—What Does It Do? Behav. Neural. Biol. 1992, 57, 2–36. [Google Scholar] [CrossRef]
- Manns, J.R.; Eichenbaum, H. Evolution of Declarative Memory. Hippocampus 2006, 16, 795–808. [Google Scholar] [CrossRef]
- Howard, M.W.; Eichenbaum, H. Time and Space in the Hippocampus. Brain Res. 2015, 1621, 345–354. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Dostrovsky, J. The Hippocampus as a Spatial Map. Preliminary Evidence from Unit Activity in the Freely-Moving Rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef]
- Hall, H.; Reyes, S.; Landeck, N.; Bye, C.; Leanza, G.; Double, K.; Thompson, L.; Halliday, G.; Kirik, D. Hippocampal Lewy Pathology and Cholinergic Dysfunction Are Associated with Dementia in Parkinson’s Disease. Brain 2014, 137, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.K.L.; Chau, T.W.; Lim, E.J.; Ahmed, I.; Chang, R.C.C.; Kalaitzakis, M.E.; Graeber, M.B.; Gentleman, S.M.; Pearce, R.K.B. Hippocampal CA2 Lewy Pathology Is Associated with Cholinergic Degeneration in Parkinson’s Disease with Cognitive Decline. Acta Neuropathol. Commun. 2019, 7, 61. [Google Scholar] [CrossRef]
- Calabresi, P.; Castrioto, A.; di Filippo, M.; Picconi, B. New Experimental and Clinical Links between the Hippocampus and the Dopaminergic System in Parkinson’s Disease. Lancet Neurol. 2013, 12, 811–821. [Google Scholar] [CrossRef]
- Hely, M.A.; Reid, W.G.J.; Adena, M.A.; Halliday, G.M.; Morris, J.G.L. The Sydney Multicenter Study of Parkinson’s Disease: The Inevitability of Dementia at 20 Years. Mov. Disord. 2008, 23, 837–844. [Google Scholar] [CrossRef]
- Novellino, F.; Vasta, R.; Sarica, A.; Chiriaco, C.; Salsone, M.; Morelli, M.; Arabia, G.; Saccà, V.; Nicoletti, G.; Quattrone, A. Relationship between Hippocampal Subfields and Category Cued Recall in AD and PDD: A Multimodal MRI Study. Neuroscience 2018, 371, 506–517. [Google Scholar] [CrossRef]
- Becker, S.; Granert, O.; Timmers, M.; Pilotto, A.; van Nueten, L.; Roeben, B.; Salvadore, G.; Galpern, W.R.; Streffer, J.; Scheffler, K.; et al. Association of Hippocampal Subfields, CSF Biomarkers, and Cognition in Patients with Parkinson Disease Without Dementia. Neurology 2021, 96, e904–e915. [Google Scholar] [CrossRef]
- Chung, S.J.; Park, Y.H.; Yun, H.J.; Kwon, H.; Yoo, H.S.; Sohn, Y.H.; Lee, J.M.; Lee, P.H. Clinical Relevance of Amnestic versus Non-Amnestic Mild Cognitive Impairment Subtyping in Parkinson’s Disease. Eur. J. Neurol. 2019, 26, 766–773. [Google Scholar] [CrossRef]
- La, C.; Linortner, P.; Bernstein, J.D.; Ua Cruadhlaoich, M.A.I.; Fenesy, M.; Deutsch, G.K.; Rutt, B.K.; Tian, L.; Wagner, A.D.; Zeineh, M.; et al. Hippocampal CA1 Subfield Predicts Episodic Memory Impairment in Parkinson’s Disease. Neuroimage Clin. 2019, 23, 101824. [Google Scholar] [CrossRef] [PubMed]
- Kunst, J.; Marecek, R.; Klobusiakova, P.; Balazova, Z.; Anderkova, L.; Nemcova-Elfmarkova, N.; Rektorova, I. Patterns of Grey Matter Atrophy at Different Stages of Parkinson’s and Alzheimer’s Diseases and Relation to Cognition. Brain Topogr. 2019, 32, 142–160. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Gao, Y.; Hu, N.; Wei, X.; Xiao, Y.; Wang, W.; Lui, S.; Gong, Q. Distinct Hippocampal Subfield Atrophy in Parkinson’s Disease Regarding Motor Subtypes. Parkinsonism Relat. Disord. 2021, 93, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Sarasso, E.; Piramide, N.; Stojkovic, T.; Stankovic, I.; Basaia, S.; Fontana, A.; Tomic, A.; Markovic, V.; Stefanova, E.; et al. Progressive Brain Atrophy and Clinical Evolution in Parkinson’s Disease. Neuroimage Clin. 2020, 28, 102374. [Google Scholar] [CrossRef]
- Hünerli, D.; Emek-Savaş, D.D.; Çavuşoğlu, B.; Dönmez Çolakoğlu, B.; Ada, E.; Yener, G.G. Mild Cognitive Impairment in Parkinson’s Disease Is Associated with Decreased P300 Amplitude and Reduced Putamen Volume. Clin. Neurophysiol. 2019, 130, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Trempler, I.; Binder, E.; El-Sourani, N.; Schiffler, P.; Tenberge, J.G.; Schiffer, A.M.; Fink, G.R.; Schubotz, R.I. Association of Grey Matter Changes with Stability and Flexibility of Prediction in Akinetic-Rigid Parkinson’s Disease. Brain Struct. Funct. 2018, 223, 2097–2111. [Google Scholar] [CrossRef]
- Wang, L.; Nie, K.; Zhao, X.; Feng, S.; Xie, S.; He, X.; Ma, G.; Wang, L.; Huang, Z.; Huang, B.; et al. Characteristics of Gray Matter Morphological Change in Parkinson’s Disease Patients with Semantic Abstract Reasoning Deficits. Neurosci. Lett. 2018, 673, 85–91. [Google Scholar] [CrossRef]
- Radziunas, A.; Deltuva, V.P.; Tamasauskas, A.; Gleizniene, R.; Pranckeviciene, A.; Petrikonis, K.; Bunevicius, A. Brain MRI Morphometric Analysis in Parkinson’s Disease Patients with Sleep Disturbances. BMC Neurol. 2018, 18, 88. [Google Scholar] [CrossRef]
- Tabi, Y.A.; Maio, M.R.; Attaallah, B.; Dickson, S.; Drew, D.; Idris, M.I.; Kienast, A.; Klar, V.; Nobis, L.; Plant, O.; et al. Vividness of Visual Imagery Questionnaire Scores and Their Relationship to Visual Short-Term Memory Performance. Cortex 2022, 146, 186–199. [Google Scholar] [CrossRef]
- Roh, H.; Kang, J.; Koh, S.B.; Kim, J.H. Hippocampal Volume Is Related to Olfactory Impairment in Parkinson’s Disease. J. Neuroimaging 2021, 31, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, L.; Yang, H.G.; Luo, X.G.; Fan, G.G. Do Multiple System Atrophy and Parkinson’s Disease Show Distinct Patterns of Volumetric Alterations across Hippocampal Subfields? An Exploratory Study. Eur. Radiol. 2019, 29, 4948–4956. [Google Scholar] [CrossRef] [PubMed]
- Mihaescu, A.S.; Masellis, M.; Graff-Guerrero, A.; Kim, J.; Criaud, M.; Cho, S.S.; Ghadery, C.; Valli, M.; Strafella, A.P. Brain Degeneration in Parkinson’s Disease Patients with Cognitive Decline: A Coordinate-Based Meta-Analysis. Brain Imaging Behav. 2019, 13, 1021–1034. [Google Scholar] [CrossRef]
- Berlot, R.; Pirtošek, Z.; Brezovar, S.; Koritnik, B.; Teipel, S.J.; Grothe, M.J.; Ray, N.J. Cholinergic Basal Forebrain and Hippocampal Structure Influence Visuospatial Memory in Parkinson’s Disease. Brain Imaging Behav. 2022, 16, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Tyacke, R.J.; Polychronis, S.; Myers, J.; Gunn, R.N.; Rabiner, E.A.; Nutt, D.; Politis, M. Imidazoline 2 Binding Sites Reflecting Astroglia Pathology in Parkinson’s Disease: An in Vivo11C-BU99008 PET Study. Brain 2019, 142, 3116–3128. [Google Scholar] [CrossRef] [PubMed]
- Schulz, J.; Pagano, G.; Fernández Bonfante, J.A.; Wilson, H.; Politis, M. Nucleus Basalis of Meynert Degeneration Precedes and Predicts Cognitive Impairment in Parkinson’s Disease. Brain 2018, 141, 1501–1516. [Google Scholar] [CrossRef]
- Filippi, M.; Canu, E.; Donzuso, G.; Stojkovic, T.; Basaia, S.; Stankovic, I.; Tomic, A.; Markovic, V.; Petrovic, I.; Stefanova, E.; et al. Tracking Cortical Changes Throughout Cognitive Decline in Parkinson’s Disease. Mov. Disord. 2020, 35, 1987–1998. [Google Scholar] [CrossRef]
- Low, A.; Foo, H.; Yong, T.T.; Tan, L.C.S.; Kandiah, N. Hippocampal Subfield Atrophy of CA1 and Subicular Structures Predict Progression to Dementia in Idiopathic Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2019, 90, 681–687. [Google Scholar] [CrossRef]
- Thomas, G.E.C.; Leyland, L.A.; Schrag, A.E.; Lees, A.J.; Acosta-Cabronero, J.; Weil, R.S. Brain Iron Deposition Is Linked with Cognitive Severity in Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2020, 91, 418–425. [Google Scholar] [CrossRef]
- Li, D.T.H.; Hui, E.S.; Chan, Q.; Yao, N.; Chua, S.E.; McAlonan, G.M.; Pang, S.Y.Y.; Ho, S.L.; Mak, H.K.F. Quantitative Susceptibility Mapping as an Indicator of Subcortical and Limbic Iron Abnormality in Parkinson’s Disease with Dementia. Neuroimage Clin. 2018, 20, 365–373. [Google Scholar] [CrossRef]
- Sejnoha Minsterova, A.; Klobusiakova, P.; Pies, A.; Galaz, Z.; Mekyska, J.; Novakova, L.; Nemcova Elfmarkova, N.; Rektorova, I. Patterns of Diffusion Kurtosis Changes in Parkinson’s Disease Subtypes. Parkinsonism Relat. Disord. 2020, 81, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Grothe, M.J.; Labrador-Espinosa, M.A.; Jesús, S.; Macías-García, D.; Adarmes-Gómez, A.; Carrillo, F.; Camacho, E.I.; Franco-Rosado, P.; Lora, F.R.; Martín-Rodríguez, J.F.; et al. In Vivo Cholinergic Basal Forebrain Degeneration and Cognition in Parkinson’s Disease: Imaging Results from the COPPADIS Study. Parkinsonism Relat. Disord. 2021, 88, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, F.; Gallea, C.; Mongin, M.; Pyatigorskaya, N.; Valabregue, R.; Ewenczyk, C.; Sarazin, M.; Yahia-Cherif, L.; Vidailhet, M.; Lehéricy, S. Multimodal Magnetic Resonance Imaging Investigation of Basal Forebrain Damage and Cognitive Deficits in Parkinson’s Disease. Mov. Disord. 2019, 34, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Paez, A.G.; Gu, C.; Rajan, S.; Miao, X.; Cao, D.; Kamath, V.; Bakker, A.; Unschuld, P.G.; Pantelyat, A.Y.; Rosenthal, L.S.; et al. Differential Changes in Arteriolar Cerebral Blood Volume between Parkinson’s Disease Patients with Normal and Impaired Cognition and Mild Cognitive Impairment (Mci) Patients without Movement Disorder—An Exploratory Study. Tomography 2020, 6, 333–342. [Google Scholar] [CrossRef]
- Bezdicek, O.; Ballarini, T.; Buschke, H.; Růžicka, F.; Roth, J.; Albrecht, F.; Růžicka, E.; Mueller, K.; Schroeter, M.L.; Jech, R. Memory Impairment in Parkinson’s Disease: The Retrieval versus Associative Deficit Hypothesis Revisited and Reconciled. Neuropsychology 2019, 33, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.J.; Banan, G.; Amin, M.; Tanner, J.J.; Hizel, L.; Nguyen, P.; Brumback, B.; Rodriguez, K.; McFarland, N.; Bowers, D.; et al. Statistically Defined Parkinson’s Disease Executive and Memory Cognitive Phenotypes: Demographic, Behavioral, and Structural Neuroimaging Comparisons. J. Parkinsons Dis. 2021, 11, 283–297. [Google Scholar] [CrossRef]
- Zarifkar, P.; Kim, J.; La, C.; Zhang, K.; YorkWilliams, S.; Levine, T.F.; Tian, L.; Borghammer, P.; Poston, K.L. Cognitive Impairment in Parkinson’s Disease Is Associated with Default Mode Network Subsystem Connectivity and Cerebrospinal Fluid Aβ. Parkinsonism Relat. Disord. 2021, 83, 71–78. [Google Scholar] [CrossRef]
- Andersen, K.B.; Hansen, A.K.; Damholdt, M.F.; Horsager, J.; Skjærbæk, C.; Gottrup, H.; Klit, H.; Schacht, A.C.; Danielsen, E.H.; Brooks, D.J.; et al. Reduced Synaptic Density in Patients with Lewy Body Dementia: An [11C]UCB-J PET Imaging Study. Mov. Disord. 2021, 36, 2057–2065. [Google Scholar] [CrossRef]
- Vriend, C.; van Balkom, T.D.; van Druningen, C.; Klein, M.; van der Werf, Y.D.; Berendse, H.W.; van den Heuvel, O.A. Processing Speed Is Related to Striatal Dopamine Transporter Availability in Parkinson’s Disease. Neuroimage Clin. 2020, 26, 102257. [Google Scholar] [CrossRef]
- Kamps, S.; van den Heuvel, O.A.; van der Werf, Y.D.; Berendse, H.W.; Weintraub, D.; Vriend, C. Smaller Subcortical Volume in Parkinson Patients with Rapid Eye Movement Sleep Behavior Disorder. Brain Imaging Behav. 2019, 13, 1352–1360. [Google Scholar] [CrossRef]
- Meles, S.K.; Renken, R.J.; Janzen, A.; Vadasz, D.; Pagani, M.; Arnaldi, D.; Morbelli, S.; Nobili, F.; Mayer, G.; Leenders, K.L.; et al. The Metabolic Pattern of Idiopathic REM Sleep Behavior Disorder Reflects Early-Stage Parkinson Disease. J. Nucl. Med. 2018, 59, 1437–1444. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Pagano, G.; Niccolini, F.; Muhlert, N.; Mehta, M.A.; Searle, G.; Gunn, R.N.; Rabiner, E.A.; Foltynie, T.; Politis, M. The Role of Phosphodiesterase 4 in Excessive Daytime Sleepiness in Parkinson’s Disease. Parkinsonism Relat. Disord. 2020, 77, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.W.; Yun, E.; Bang, M.; Yoon, U.; Yoo, J.Y.; Lee, K.S.; Shin, N.Y.; Kim, J.S. Blood Pressure Lability Is Associated with Subcortical Atrophy in Early Parkinson’s Disease. J. Hypertens. 2020, 38, 2043–2049. [Google Scholar] [CrossRef]
- Rahayel, S.; Gaubert, M.; Postuma, R.B.; Montplaisir, J.; Carrier, J.; Monchi, O.; Remillard-Pelchat, D.; Bourgouin, P.A.; Panisset, M.; Chouinard, S.; et al. Brain Atrophy in Parkinson’s Disease with Polysomnography-Confirmed REM Sleep Behavior Disorder. Sleep 2019, 42, zsz062. [Google Scholar] [CrossRef] [PubMed]
- Tessa, C.; Toschi, N.; Orsolini, S.; Valenza, G.; Lucetti, C.; Barbieri, R.; Diciotti, S. Central Modulation of Parasympathetic Outflow Is Impaired in de Novo Parkinson’s Disease Patients. PLoS ONE 2019, 14, e0210324. [Google Scholar] [CrossRef]
- Goto, M.; Kamagata, K.; Hatano, T.; Hattori, N.; Abe, O.; Aoki, S.; Hori, M.; Gomi, T. Depressive Symptoms in Parkinson’s Disease Are Related to Decreased Left Hippocampal Volume: Correlation with the 15-Item Shortened Version of the Geriatric Depression Scale. Acta Radiol. 2018, 59, 341–345. [Google Scholar] [CrossRef]
- Lenka, A.; Ingalhalikar, M.; Shah, A.; Saini, J.; Arumugham, S.S.; Hegde, S.; George, L.; Reddy, V.; Reddy, Y.C.J.; Yadav, R.; et al. Hippocampal Subfield Atrophy in Patients with Parkinson’s Disease and Psychosis. J. Neural. Transm. 2018, 125, 1361–1372. [Google Scholar] [CrossRef]
- Vignando, M.; Ffytche, D.; Lewis, S.J.G.; Lee, P.H.; Chung, S.J.; Weil, R.S.; Hu, M.T.; Mackay, C.E.; Griffanti, L.; Pins, D.; et al. Mapping Brain Structural Differences and Neuroreceptor Correlates in Parkinson’s Disease Visual Hallucinations. Nat. Commun. 2022, 13, 519. [Google Scholar] [CrossRef]
- van der Vegt, J.P.M.; Hulme, O.J.; Madsen, K.H.; Buhmann, C.; Bloem, B.R.; Münchau, A.; Helmich, R.C.; Siebner, H.R. Dopamine Agonist Treatment Increases Sensitivity to Gamble Outcomes in the Hippocampus in de Novo Parkinson’s Disease. Neuroimage Clin. 2020, 28, 102362. [Google Scholar] [CrossRef]
- Li, N.; Suo, X.; Zhang, J.; Lei, D.; Wang, L.; Li, J.; Peng, J.; Duan, L.; Gong, Q.; Peng, R. Disrupted Functional Brain Network Topology in Parkinson’s Disease Patients with Freezing of Gait. Neurosci. Lett. 2021, 759, 135970. [Google Scholar] [CrossRef]
- Kang, S.Y.; Bang, M.; Hong, J.Y.; Oh, J.; Kim, J.S.; Han, Y.M.; Chang, S.K.; Lee, S.A.; Yoon, U.; Shin, N.Y. Neural and Dopaminergic Correlates of Fatigue in Parkinson’s Disease. J. Neural. Transm. 2020, 127, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Sarasso, E.; Agosta, F.; Piramide, N.; Canu, E.; Volontè, M.A.; Filippi, M. Brain Activity of the Emotional Circuit in Parkinson’s Disease Patients with Freezing of Gait. Neuroimage Clin. 2021, 30, 102649. [Google Scholar] [CrossRef]
- Hall, J.M.; Shine, J.M.; Ehgoetz Martens, K.A.; Gilat, M.; Broadhouse, K.M.; Szeto, J.Y.Y.; Walton, C.C.; Moustafa, A.A.; Lewis, S.J.G. Alterations in White Matter Network Topology Contribute to Freezing of Gait in Parkinson’s Disease. J. Neurol. 2018, 265, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Kim, B.H.; Chung, S.J.; Yoo, H.S.; Lee, Y.H.; Baik, K.; Ye, B.S.; Sohn, Y.H.; Lee, J.M.; Lee, P.H. Motor Cerebellar Connectivity and Future Development of Freezing of Gait in De Novo Parkinson’s Disease. Mov. Disord. 2020, 35, 2240–2249. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Yoon, E.J.; Kibreab, M.; Kathol, I.; Cheetham, J.; Hammer, T.; Sarna, J.; Ismail, Z.; Monchi, O. Mild Behavioral Impairment in Parkinson’s Disease Is Associated with Altered Corticostriatal Connectivity. Neuroimage Clin. 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, E.; Roeben, B.; Granert, O.; Hanert, A.; Liepelt-Scarfone, I.; Leks, E.; Otterbein, S.; Saraykin, P.; Busch, J.H.; Synofzik, M.; et al. Effects of Exergaming on Hippocampal Volume and Brain-Derived Neurotrophic Factor Levels in Parkinson’s Disease. Eur. J. Neurol. 2022, 29, 441–449. [Google Scholar] [CrossRef]
- Joling, M.; Vriend, C.; Raijmakers, P.G.H.M.; van der Zande, J.J.; Lemstra, A.W.; Berendse, H.W.; Booij, J.; van den Heuvel, O.A. Striatal DAT and Extrastriatal SERT Binding in Early-Stage Parkinson’s Disease and Dementia with Lewy Bodies, Compared with Healthy Controls: An 123I-FP-CIT SPECT Study. Neuroimage Clin. 2019, 22, 101755. [Google Scholar] [CrossRef]
- Bernstein, H.G.; Dobrowolny, H.; Bogerts, B.; Keilhoff, G.; Steiner, J. The Hypothalamus and Neuropsychiatric Disorders: Psychiatry Meets Microscopy. Cell Tissue Res. 2018, 375, 243–258. [Google Scholar] [CrossRef]
- Saper, C.B.; Lowell, B.B. The Hypothalamus. Curr. Biol. 2014, 24, R1111–R1116. [Google Scholar] [CrossRef]
- Swaab, D.F. The Human Hypothalamus. Basic and Clinical Aspects: Part I: Nuclei of the Human Hypothalamus. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Alstadhaug, K. Migraine and the Hypothalamus. Cephalalgia 2009, 29, 809–817. [Google Scholar] [CrossRef]
- Langston, J.W.; Forno, L.S. The Hypothalamus in Parkinson Disease. Ann. Neurol. 1978, 3, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Lewy, F.H. Die Lehre Vom Tonus der Bewegung, Zugleich Systema—Tische Untersuchungen zur Klinik, Physiologie, Pathologie und Pathogenese Paralysis Agitans; Springer: Berlin/Heidelberg, Germany, 1923. [Google Scholar]
- Gorges, M.; Kuntz, B.; del Tredici, K.; Schmidt, D.G.; Müller, H.P.; Ludolph, A.C.; Dupuis, L.; Kassubek, J. Morphological MRI Investigations of the Hypothalamus in 232 Individuals with Parkinson’s Disease. Mov. Disord. 2019, 34, 1566–1570. [Google Scholar] [CrossRef] [Green Version]
- Dayan, E.; Sklerov, M.; Browner, N. Disrupted Hypothalamic Functional Connectivity in Patients with PD and Autonomic Dysfunction. Neurology 2018, 90, e2051–e2058. [Google Scholar] [CrossRef] [PubMed]
- Nishi, A.; Kuroiwa, M.; Miller, D.B.; O’callaghan, J.P.; Bateup, H.S.; Shuto, T.; Sotogaku, N.; Fukuda, T.; Heintz, N.; Greengard, P.; et al. Distinct Roles of PDE4 and PDE10A in the Regulation of CAMP/PKA Signaling in the Striatum. Soc. Neurosci. 2008, 28, 10460–10471. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Giordano, B.; Turkheimer, F.E.; Chaudhuri, K.R.; Politis, M. Serotonergic Dysregulation Is Linked to Sleep Problems in Parkinson’s Disease. Neuroimage Clin. 2018, 18, 630–637. [Google Scholar] [CrossRef]
- Broca, P. Sur La Circonvolution Limbique et La Scissure Limbique. Bull. Mem. Soc. Anthr. Paris 1877, 12, 646–657. [Google Scholar] [CrossRef]
- Vogt, B.A.; Nimchinsky, E.A.; Vogt, L.J.; Hof, P.R. Human Cingulate Cortex: Surface Features, Flat Maps, and Cytoarchitecture. J. Comp. Neurol. 1995, 359, 490–506. [Google Scholar] [CrossRef]
- Vogt, B.A. Pain and Emotion Interactions in Subregions of the Cingulate Gyrus. Nat. Rev. Neurosci. 2005, 6, 533–544. [Google Scholar] [CrossRef]
- Rolls, E.T. The Cingulate Cortex and Limbic Systems for Emotion, Action, and Memory. Brain Struct. Funct. 2019, 224, 3001–3018. [Google Scholar] [CrossRef]
- Alexander, W.H.; Brown, J.W. The Role of the Anterior Cingulate Cortex in Prediction Error and Signaling Surprise. Top. Cogn. Sci. 2019, 11, 119–135. [Google Scholar] [CrossRef]
- Elliott, R.; Rubinsztein, J.S.; Sahakian, B.J.; Dolan, R.J. Selective Attention to Emotional Stimuli in a Verbal Go/No-Go Task: An FMRI Study. Neuroreport 2000, 11, 1739–1744. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.M.; Heilbronner, S.R.; Barack, D.L.; Hayden, B.Y.; Platt, M.L. Posterior Cingulate Cortex: Adapting Behavior to a Changing World. Trends Cogn. Sci. 2011, 15, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammes, J.; Theis, H.; Giehl, K.; Hoenig, M.C.; Greuel, A.; Tittgemeyer, M.; Timmermann, L.; Fink, G.R.; Drzezga, A.; Eggers, C.; et al. Dopamine Metabolism of the Nucleus Accumbens and Fronto-Striatal Connectivity Modulate Impulse Control. Brain 2019, 142, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Navalpotro-Gomez, I.; Kim, J.; Paz-Alonso, P.M.; Delgado-Alvarado, M.; Quiroga-Varela, A.; Jimenez-Urbieta, H.; Carreiras, M.; Strafella, A.P.; Rodriguez-Oroz, M.C. Disrupted Salience Network Dynamics in Parkinson’s Disease Patients with Impulse Control Disorders. Parkinsonism Relat. Disord. 2020, 70, 74–81. [Google Scholar] [CrossRef]
- Girard, R.; Obeso, I.; Thobois, S.; Park, S.A.; Vidal, T.; Favre, E.; Ulla, M.; Broussolle, E.; Krack, P.; Durif, F.; et al. Wait and You Shall See: Sexual Delay Discounting in Hypersexual Parkinson’s Disease. Brain 2019, 142, 146–162. [Google Scholar] [CrossRef]
- Li, Y.; Huang, P.; Guo, T.; Guan, X.; Gao, T.; Sheng, W.; Zhou, C.; Wu, J.; Song, Z.; Xuan, M.; et al. Brain Structural Correlates of Depressive Symptoms in Parkinson’s Disease Patients at Different Disease Stage. Psychiatry Res. Neuroimaging 2020, 296, 111029. [Google Scholar] [CrossRef]
- Wu, J.Y.; Zhang, Y.; Wu, W.B.; Hu, G.; Xu, Y. Impaired Long Contact White Matter Fibers Integrity Is Related to Depression in Parkinson’s Disease. CNS Neurosci. 2018, 24, 108–114. [Google Scholar] [CrossRef]
- Ghazi Sherbaf, F.; Same, K.; Aarabi, M.H. High Angular Resolution Diffusion Imaging Correlates of Depression in Parkinson’s Disease: A Connectometry Study. Acta Neurol. Belg. 2018, 118, 573–579. [Google Scholar] [CrossRef]
- Wang, H.; Chen, H.; Wu, J.; Tao, L.; Pang, Y.; Gu, M.; Lv, F.; Luo, T.; Cheng, O.; Sheng, K.; et al. Altered Resting-State Voxel-Level Whole-Brain Functional Connectivity in Depressed Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 50, 74–80. [Google Scholar] [CrossRef]
- Wei, L.; Hu, X.; Yuan, Y.; Liu, W.; Chen, H. Abnormal Ventral Tegmental Area-Anterior Cingulate Cortex Connectivity in Parkinson’s Disease with Depression. Behav. Brain Res. 2018, 347, 132–139. [Google Scholar] [CrossRef]
- Qiu, Y.H.; Huang, Z.H.; Gao, Y.Y.; Feng, S.J.; Huang, B.; Wang, W.Y.; Xu, Q.H.; Zhao, J.H.; Zhang, Y.H.; Wang, L.M.; et al. Alterations in Intrinsic Functional Networks in Parkinson’s Disease Patients with Depression: A Resting-State Functional Magnetic Resonance Imaging Study. CNS Neurosci. 2021, 27, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.H.; Hu, J.B.; Chen, J.; Wang, X.Y.; Wang, X.L.; Pan, P.L.; Liu, C.F. Abnormal Spontaneous Neural Activity in Parkinson’s Disease With “Pure” Apathy. Front. Neurosci. 2020, 14, 830. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, J.; Wu, W.; Liu, R.; Pang, L.; Guan, D.; Xu, Y. Reduction of White Matter Integrity Correlates with Apathy in Parkinson’s Disease. Int. J. Neurosci. 2017, 128, 25–31. [Google Scholar] [CrossRef]
- Zhou, Z.; Müller, M.L.T.M.; Kanel, P.; Chua, J.; Kotagal, V.; Kaufer, D.I.; Albin, R.L.; Frey, K.A.; Bohnen, N.I. Apathy Rating Scores and β-Amyloidopathy in Patients with Parkinson Disease at Risk for Cognitive Decline. Neurology 2020, 94, e376–e383. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Kajiyama, Y.; Revankar, G.S.; Hashimoto, R.; Watanabe, Y.; Kishima, H.; Ikeda, M.; Mihara, M.; Mochizuki, H.; Hattori, N. Neural Networks Associated with Quality of Life in Patients with Parkinson’s Disease. Parkinsonism Relat. Disord. 2021, 89, 6–12. [Google Scholar] [CrossRef]
- Jørgensen, L.M.; Henriksen, T.; Mardosiene, S.; Keller, S.H.; Stenbæk, D.S.; Hansen, H.D.; Jespersen, B.; Thomsen, C.; Weikop, P.; Svarer, C.; et al. Parkinson Patients Have a Presynaptic Serotonergic Deficit: A Dynamic Deep Brain Stimulation PET Study. J. Cereb. Blood Flow Metab. 2021, 41, 1954–1963. [Google Scholar] [CrossRef] [PubMed]
- Varrone, A.; Svenningsson, P.; Forsberg, A.; Varnäs, K.; Tiger, M.; Nakao, R.; Halldin, C.; Nilsson, L.G.; Farde, L. Positron Emission Tomography Imaging of 5-Hydroxytryptamine1B Receptors in Parkinson’s Disease. Neurobiol. Aging 2014, 35, 867–875. [Google Scholar] [CrossRef]
- Maillet, A.; Météreau, E.; Tremblay, L.; Favre, E.; Klinger, H.; Lhommée, E.; le Bars, D.; Castrioto, A.; Prange, S.; Sgambato, V.; et al. Serotonergic and Dopaminergic Lesions Underlying Parkinsonian Neuropsychiatric Signs. Mov. Disord. 2021, 36, 2888–2900. [Google Scholar] [CrossRef]
- Maillet, A.; Krack, P.; Lhommée, E.; Météreau, E.; Klinger, H.; Favre, E.; le Bars, D.; Schmitt, E.; Bichon, A.; Pelissier, P.; et al. The Prominent Role of Serotonergic Degeneration in Apathy, Anxiety and Depression in de Novo Parkinson’s Disease. Brain 2016, 139, 2486–2502. [Google Scholar] [CrossRef]
- Prange, S.; Metereau, E.; Maillet, A.; Klinger, H.; Schmitt, E.; Lhommée, E.; Bichon, A.; Lancelot, S.; Meoni, S.; Broussolle, E.; et al. Limbic Serotonergic Plasticity Contributes to the Compensation of Apathy in Early Parkinson’s Disease. Mov. Disord. 2022, 37, 1211–1221. [Google Scholar] [CrossRef]
- Owens-Walton, C.; Jakabek, D.; Power, B.D.; Walterfang, M.; Hall, S.; van Westen, D.; Looi, J.C.L.; Shaw, M.; Hansson, O. Structural and Functional Neuroimaging Changes Associated with Cognitive Impairment and Dementia in Parkinson’s Disease. Psychiatry Res. Neuroimaging 2021, 312, 111273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, B.; Chen, Q.; Wang, L.; Zhang, L.; Nie, K.; Huang, Q.; Huang, R. Altered Microstructural Properties of Superficial White Matter in Patients with Parkinson’s Disease. Brain Imaging Behav. 2021, 16, 476–491. [Google Scholar] [CrossRef]
- Guo, T.; Guan, X.; Zeng, Q.; Xuan, M.; Gu, Q.; Xu, X.; Zhang, M. Correlations between CSF Proteins and Spontaneous Neuronal Activity in Parkinson’s Disease. Neurosci. Lett. 2018, 673, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.W.; Lin, L.Z.; Yu, E.H.; Xin, J.W.; Lin, L.; Lin, H.L.; Ye, Q.Y.; Chen, X.C.; Pan, X.D. Abnormal Resting-State Functional Connectivity in Posterior Cingulate Cortex of Parkinson’s Disease with Mild Cognitive Impairment and Dementia. CNS Neurosci. 2018, 24, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Weil, R.S.; Winston, J.S.; Leyland, L.A.; Pappa, K.; Mahmood, R.B.; Morris, H.R.; Rees, G. Neural Correlates of Early Cognitive Dysfunction in Parkinson’s Disease. Ann. Clin. Transl. Neurol. 2019, 6, 902–912. [Google Scholar] [CrossRef]
- Wolters, A.F.; Moonen, A.J.H.; Lopes, R.; Leentjens, A.F.G.; Duits, A.A.; Defebvre, L.; Delmaire, C.; Hofman, P.A.; van Bussel, F.C.; Dujardin, K. Grey Matter Abnormalities Are Associated Only with Severe Cognitive Decline in Early Stages of Parkinson’s Disease. Cortex 2020, 123, 1–11. [Google Scholar] [CrossRef]
- Fathy, Y.Y.; Hepp, D.H.; de Jong, F.J.; Geurts, J.J.G.; Foncke, E.M.J.; Berendse, H.W.; van de Berg, W.D.J.; Schoonheim, M.M. Anterior Insular Network Disconnection and Cognitive Impairment in Parkinson’s Disease. Neuroimage Clin. 2020, 28, 102364. [Google Scholar] [CrossRef]
- Jonkman, L.E.; Fathy, Y.Y.; Berendse, H.W.; Schoonheim, M.M.; van de Berg, W.D.J. Structural Network Topology and Microstructural Alterations of the Anterior Insula Associate with Cognitive and Affective Impairment in Parkinson’s Disease. Sci. Rep. 2021, 11, 16021. [Google Scholar] [CrossRef]
- van Balkom, T.D.; van den Heuvel, O.A.; Berendse, H.W.; van der Werf, Y.D.; Vriend, C. Eight-Week Multi-Domain Cognitive Training Does Not Impact Large-Scale Resting-State Brain Networks in Parkinson’s Disease. Neuroimage Clin. 2022, 33, 102952. [Google Scholar] [CrossRef]
- Pan, C.; Ren, J.; Li, L.; Li, Y.; Xu, J.; Xue, C.; Hu, G.; Yu, M.; Chen, Y.; Zhang, L.; et al. Differential Functional Connectivity of Insular Subdivisions in de Novo Parkinson’s Disease with Mild Cognitive Impairment. Brain Imaging Behav. 2022, 16, 1–10. [Google Scholar] [CrossRef]
- van der Zee, S.; Müller, M.L.T.M.; Kanel, P.; van Laar, T.; Bohnen, N.I. Cholinergic Denervation Patterns Across Cognitive Domains in Parkinson’s Disease. Mov. Disord. 2021, 36, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, I.; Lange, C.; Frings, L.; Klutmann, S.; Meyer, P.T.; Buchert, R. Nigrostriatal Degeneration in the Cognitive Part of the Striatum in Parkinson Disease Is Associated with Frontomedial Hypometabolism. Clin. Nucl. Med. 2020, 45, 95–99. [Google Scholar] [CrossRef]
- Barzgari, A.; Sojkova, J.; Maritza Dowling, N.; Pozorski, V.; Okonkwo, O.C.; Starks, E.J.; Oh, J.; Thiesen, F.; Wey, A.; Nicholas, C.R.; et al. Arterial Spin Labeling Reveals Relationships between Resting Cerebral Perfusion and Motor Learning in Parkinson’s Disease. Brain Imaging Behav. 2019, 13, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shen, Y.; Peng, J.; Wang, A.; Wu, X.; Chen, X.; Liu, J.; Wei, M.; Zou, D.; Han, Y.; et al. Different Functional Connectivity Modes of the Right Fronto-Insular Cortex in Akinetic-Rigid and Tremor-Dominant Parkinson’s Disease. Neurol. Sci. 2021, 42, 2937–2946. [Google Scholar] [CrossRef] [PubMed]
- Kanno, S.; Shinohara, M.; Kanno, K.; Gomi, Y.; Uchiyama, M.; Nishio, Y.; Baba, T.; Hosokai, Y.; Takeda, A.; Fukuda, H.; et al. Neural Substrates Underlying Progressive Micrographia in Parkinson’s Disease. Brain. Behav. 2020, 10, e01669. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Hirano, S.; Yamamoto, T.; Asahina, M.; Uchiyama, T.; Yamanaka, Y.; Nakano, Y.; Ishikawa, A.; Kojima, K.; Abe, M.; et al. Decline in Drawing Ability and Cerebral Perfusion in Parkinson’s Disease Patients after Subthalamic Nucleus Deep Brain Stimulation Surgery. Parkinsonism Relat. Disord. 2020, 70, 60–66. [Google Scholar] [CrossRef]
- Klobusiakova, P.; Mekyska, J.; Brabenec, L.; Galaz, Z.; Zvoncak, V.; Mucha, J.; Rapcsak, S.Z.; Rektorova, I. Articulatory Network Reorganization in Parkinson’s Disease as Assessed by Multimodal MRI and Acoustic Measures. Parkinsonism Relat. Disord. 2021, 84, 122–128. [Google Scholar] [CrossRef]
- Oltra, J.; Campabadal, A.; Segura, B.; Uribe, C.; Marti, M.J.; Compta, Y.; Valldeoriola, F.; Bargallo, N.; Iranzo, A.; Junque, C. Disrupted Functional Connectivity in PD with Probable RBD and Its Cognitive Correlates. Sci. Rep. 2021, 11, 24351. [Google Scholar] [CrossRef]
- Dimarzio, M.; Rashid, T.; Hancu, I.; Fiveland, E.; Prusik, J.; Gillogly, M.; Madhavan, R.; Joel, S.; Durphy, J.; Molho, E.; et al. Functional MRI Signature of Chronic Pain Relief from Deep Brain Stimulation in Parkinson Disease Patients. Clin. Neurosurg. 2019, 85, E1043–E1049. [Google Scholar] [CrossRef]
- Dellapina, E.; Pellaprat, J.; Adel, D.; Llido, J.; Harroch, E.; Martini, J.B.; Kas, A.; Salabert, A.S.; Ory-Magne, F.; Payoux, P.; et al. Dopaminergic Denervation Using [ 123 I]-FPCIT and Pain in Parkinson’s Disease: A Correlation Study. J. Neural Transm. 2019, 126, 279–287. [Google Scholar] [CrossRef]
- Zhang, P.F.; Gao, F. Neuroinflammation in Parkinson’s Disease: A Meta-Analysis of PET Imaging Studies. J. Neurol. 2022, 269, 2304–2314. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Ikeda, T.; Ishida, C.; Komai, K.; Yamada, M. Delusions and Visual Hallucinations in a Patient with Parkinson’s Disease with Dementia Showing Pronounced Lewy Body Pathology in the Nucleus Basalis of Meynert. Neuropathology 2019, 39, 319–323. [Google Scholar] [CrossRef]
- Ray, N.J.; Lawson, R.A.; Martin, S.L.; Sigurdsson, H.P.; Wilson, J.; Galna, B.; Lord, S.; Alcock, L.; Duncan, G.W.; Khoo, T.K.; et al. Free-Water Imaging of the Cholinergic Basal Forebrain and Pedunculopontine Nucleus in Parkinson’s Disease. Brain 2022, awac127. [Google Scholar] [CrossRef]
- Pieperhoff, P.; Südmeyer, M.; Dinkelbach, L.; Hartmann, C.J.; Ferrea, S.; Moldovan, A.S.; Minnerop, M.; Diaz-Pier, S.; Schnitzler, A.; Amunts, K. Regional Changes of Brain Structure during Progression of Idiopathic Parkinson’s Disease—A Longitudinal Study Using Deformation Based Morphometry. Cortex 2022, 151, 188–210. [Google Scholar] [CrossRef]
- Rogozinski, S.; Klietz, M.; Respondek, G.; Oertel, W.H.; Grothe, M.J.; Pereira, J.B.; Höglinger, G.U. Reduction in Volume of Nucleus Basalis of Meynert Is Specific to Parkinson’s Disease and Progressive Supranuclear Palsy but Not to Multiple System Atrophy. Front Aging Neurosci. 2022, 14, 851788. [Google Scholar] [CrossRef] [PubMed]
- Rong, S.; Li, Y.; Li, B.; Nie, K.; Zhang, P.; Cai, T.; Mei, M.; Wang, L.; Zhang, Y. Meynert Nucleus-Related Cortical Thinning in Parkinson’s Disease with Mild Cognitive Impairment. Quant. Imaging Med. Surg. 2021, 11, 1551566–1554566. [Google Scholar] [CrossRef]
- Gang, M.; Baba, T.; Hosokai, Y.; Nishio, Y.; Kikuchi, A.; Hirayama, K.; Hasegawa, T.; Aoki, M.; Takeda, A.; Mori, E.; et al. Clinical and Cerebral Metabolic Changes in Parkinson’s Disease with Basal Forebrain Atrophy. Mov. Disord. 2020, 35, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, C.; Zhang, H.; Dou, W.; Li, W.; Sami, M.U.; Xu, K. Disrupted Resting-State Functional Connectivity of the Nucleus Basalis of Meynert in Parkinson’s Disease with Mild Cognitive Impairment. Neuroscience 2020, 442, 228–236. [Google Scholar] [CrossRef]
- Lee, Y.; Ham, J.H.; Cha, J.; Park, Y.-H.; Lee, J.J.; Sunwoo, M.K.; Hong, J.Y.; Sohn, Y.H.; Lee, J.-M.; Lee, P.H. The Cholinergic Contribution to the Resting-State Functional Network in Non-Demented Parkinson’s Disease. Sci. Rep. 2018, 8, 7683. [Google Scholar] [CrossRef]
- Boshkovski, T.; Cohen-Adad, J.; Misic, B.; Arnulf, I.; Corvol, J.-C.; Vidailhet, M.; Lehéricy, S.; Stikov, N.; Mancini, M. The Myelin-Weighted Connectome in Parkinson’s Disease. Mov. Disord. 2022, 37, 724–733. [Google Scholar] [CrossRef]
- Nazmuddin, M.; van Dalen, J.-W.; Borra, R.J.H.; Stormezand, G.N.; van der Horn, H.J.; van der Zee, S.; Boertien, J.; van Laar, T. Postural and Gait Symptoms in de Novo Parkinson’s Disease Patients Correlate with Cholinergic White Matter Pathology. Parkinsonism Relat. Disord. 2021, 93, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Hepp, D.H.; Foncke, E.M.J.; Berendse, H.W.; Wassenaar, T.M.; Olde Dubbelink, K.T.E.; Groenewegen, H.J.; van de Berg, W.; Schoonheim, M.M. Damaged Fiber Tracts of the Nucleus Basalis of Meynert in Parkinson’s Disease Patients with Visual Hallucinations. Sci. Rep. 2017, 7, 10112. [Google Scholar] [CrossRef] [Green Version]
- Sperling, S.A.; Druzgal, J.; Blair, J.C.; Flanigan, J.L.; Stohlman, S.L.; Barrett, M.J. Cholinergic Nucleus 4 Grey Matter Density Is Associated with Apathy in Parkinson’s Disease. Clin. Neuropsychol. 2022, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Yarnall, A.J.; Craig, C.E.; Galna, B.; Lord, S.; Morris, R.; Lawson, R.A.; Alcock, L.; Duncan, G.W.; Khoo, T.K.; et al. Cholinergic Basal Forebrain Volumes Predict Gait Decline in Parkinson’s Disease. Mov. Disord. 2021, 36, 611–621. [Google Scholar] [CrossRef]
- Barrett, M.J.; Sperling, S.A.; Blair, J.C.; Freeman, C.S.; Flanigan, J.L.; Smolkin, M.E.; Manning, C.A.; Druzgal, T.J. Lower Volume, More Impairment: Reduced Cholinergic Basal Forebrain Grey Matter Density Is Associated with Impaired Cognition in Parkinson Disease. J. Neurol. Neurosurg. Psychiatry 2019, 90, 1251–1256. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.J.; Bradburn, S.; Murgatroyd, C.; Toseeb, U.; Mir, P.; Kountouriotis, G.K.; Teipel, S.J.; Grothe, M.J. In Vivo Cholinergic Basal Forebrain Atrophy Predicts Cognitive Decline in de Novo Parkinson’s Disease. Brain 2018, 141, 165–176. [Google Scholar] [CrossRef]
- Pereira, J.B.; Hall, S.; Jalakas, M.; Grothe, M.J.; Strandberg, O.; Stomrud, E.; Westman, E.; van Westen, D.; Hansson, O. Longitudinal Degeneration of the Basal Forebrain Predicts Subsequent Dementia in Parkinson’s Disease. Neurobiol. Dis. 2020, 139, 104831. [Google Scholar] [CrossRef]
- Wilson, H.; de Natale, E.R.; Politis, M. Chapter 12—Nucleus Basalis of Meynert Degeneration Predicts Cognitive Impairment in Parkinson’s Disease. In Handbook of Clinical Neurology. The Human Hypothalamus: Anterior Region; Swaab, D.F., Kreier, F., Lucassen, P.J., Salehi, A., Buijs, R.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 179, pp. 189–205. [Google Scholar]
- Guttuso Jr, T.; Sirica, D.; Tosun, D.; Zivadinov, R.; Pasternak, O.; Weintraub, D.; Baglio, F.; Bergsland, N. Thalamic Dorsomedial Nucleus Free Water Correlates with Cognitive Decline in Parkinson’s Disease. Mov. Disord. 2022, 37, 490–501. [Google Scholar] [CrossRef]
- Kübler, D.; Wellmann, S.K.; Kaminski, J.; Skowronek, C.; Schneider, G.-H.; Neumann, W.-J.; Ritter, K.; Kühn, A. Nucleus Basalis of Meynert Predicts Cognition after Deep Brain Stimulation in Parkinson’s Disease. Parkinsonism Relat. Disord. 2022, 94, 89–95. [Google Scholar] [CrossRef]
- Sasikumar, S.; Cohn, M.; Harmsen, I.E.; Loh, A.; Cho, S.S.; Sáenz-Farret, M.; Maciel, R.; Soh, D.; Boutet, A.; Germann, J.; et al. Single-Trajectory Multiple-Target Deep Brain Stimulation for Parkinsonian Mobility and Cognition. Mov. Disord. 2022, 37, 635–640. [Google Scholar] [CrossRef]
- Gratwicke, J.; Zrinzo, L.; Kahan, J.; Peters, A.; Beigi, M.; Akram, H.; Hyam, J.; Oswal, A.; Day, B.; Mancini, L.; et al. Bilateral Deep Brain Stimulation of the Nucleus Basalis of Meynert for Parkinson Disease Dementia: A Randomized Clinical Trial. JAMA Neurol. 2018, 75, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Rea, R.C.; Berlot, R.; Martin, S.L.; Craig, C.E.; Holmes, P.S.; Wright, D.J.; Bon, J.; Pirtošek, Z.; Ray, N.J. Quantitative EEG and Cholinergic Basal Forebrain Atrophy in Parkinson’s Disease and Mild Cognitive Impairment. Neurobiol. Aging 2021, 106, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Oswal, A.; Gratwicke, J.; Akram, H.; Jahanshahi, M.; Zaborszky, L.; Brown, P.; Hariz, M.; Zrinzo, L.; Foltynie, T.; Litvak, V. Cortical Connectivity of the Nucleus Basalis of Meynert in Parkinson’s Disease and Lewy Body Dementias. Brain 2021, 144, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Dell’Acqua, F.; de Schotten, M. A Revised Limbic System Model for Memory, Emotion and Behaviour. Neurosci. Biobehav. Rev. 2013, 37, 1724–1737. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Belke, M.; Menzler, K.; Heverhagen, J.T.; Keil, B.; Stiasny-Kolster, K.; Rosenow, F.; Diederich, N.J.; Mayer, G.; Möller, J.C.; et al. Diffusion Tensor Imaging in Idiopathic REM Sleep Behavior Disorder Reveals Microstructural Changes in the Brainstem, Substantia Nigra, Olfactory Region, and Other Brain Regions. Sleep 2010, 33, 767–773. [Google Scholar] [CrossRef]
- Haghshomar, M.; Rahmani, F.; Hadi Aarabi, M.; Shahjouei, S.; Sobhani, S.; Rahmani, M. White Matter Changes Correlates of Peripheral Neuroinflammation in Patients with Parkinson’s Disease. Neuroscience 2019, 403, 70–78. [Google Scholar] [CrossRef]
- Pu, W.; Shen, X.; Huang, M.; Li, Z.; Zeng, X.; Wang, R.; Shen, G.; Yu, H. Assessment of White Matter Lesions in Parkinson’s Disease: Voxel-Based Analysis and Tract-Based Spatial Statistics Analysis of Parkinson’s Disease with Mild Cognitive Impairment. Curr. Neurovasc. Res. 2020, 17, 480–486. [Google Scholar] [CrossRef]
- Zheng, Z.; Shemmassian, S.; Wijekoon, C.; Kim, W.; Bookheimer, S.Y.; Pouratian, N. DTI Correlates of Distinct Cognitive Impairments in Parkinson’s Disease. Hum. Brain. Mapp. 2014, 35, 1325–1333. [Google Scholar] [CrossRef]
- Matsui, H.; Nishinaka, K.; Oda, M.; Niikawa, H.; Komatsu, K.; Kubori, T.; Udaka, F. Disruptions of the Fornix Fiber in Parkinsonian Patients with Excessive Daytime Sleepiness. Parkinsonism Relat. Disord. 2006, 12, 319–322. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, S.J.; Kim, H.S.; Choi, C.G.; Kim, N.; Han, S.; Jang, E.H.; Chung, S.J.; Lee, C.S. Alterations of Mean Diffusivity in Brain White Matter and Deep Gray Matter in Parkinson’s Disease. Neurosci. Lett. 2013, 550, 64–68. [Google Scholar] [CrossRef]
- Li, X.-R.; Ren, Y.-D.; Cao, B.; Huang, X.-L. Analysis of White Matter Characteristics with Tract-Based Spatial Statistics According to Diffusion Tensor Imaging in Early Parkinson’s Disease. Neurosci. Lett. 2018, 675, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, S.; Rahmani, F.; Aarabi, M.H.; Sadr, A.V. Exploring White Matter Microstructure and Olfaction Dysfunction in Early Parkinson Disease: Diffusion MRI Reveals New Insight. Brain Imaging Behav. 2019, 13, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Foo, H.; Mak, E.; Chander, R.J.; Ng, A.; Au, W.L.; Sitoh, Y.Y.; Tan, L.C.S.; Kandiah, N. Associations of Hippocampal Subfields in the Progression of Cognitive Decline Related to Parkinson’s Disease. Neuroimage Clin. 2017, 14, 37–42. [Google Scholar] [CrossRef]
- Dolatshahi, M.; Ashraf-Ganjouei, A.; Wu, I.W.; Zhang, Y.; Aarabi, M.H.; Tosun, D. White Matter Changes in Drug-Naïve Parkinson’s Disease Patients with Impulse Control & Probable REM Sleep Behavior Disorders. J. Neurol. Sci. 2021, 430, 120032. [Google Scholar] [CrossRef] [PubMed]
- Ghazi Sherbaf, F.; Rahmani, F.; Jooyandeh, S.M.; Aarabi, M.H. Microstructural Changes in Patients with Parkinson Disease and REM Sleep Behavior Disorder: Depressive Symptoms versus Non-Depressed. Acta Neurol. Belg. 2018, 118, 415–421. [Google Scholar] [CrossRef]
- He, N.; Sethi, S.K.; Zhang, C.; Li, Y.; Chen, Y.; Sun, B.; Yan, F.; Haacke, E.M. Visualizing the Lateral Habenula Using Susceptibility Weighted Imaging and Quantitative Susceptibility Mapping. Magn. Reson. Imaging 2020, 65, 55–61. [Google Scholar] [CrossRef]
- Kochanski, R.B.; Dawe, R.; Kocak, M.; Sani, S. Identification of Stria Medullaris Fibers in the Massa Intermedia Using Diffusion Tensor Imaging. World Neurosurg. 2018, 112, e497–e504. [Google Scholar] [CrossRef]
- Sartorius, A.; Kiening, K.L.; Kirsch, P.; von Gall, C.C.; Haberkorn, U.; Unterberg, A.W.; Henn, F.A.; Meyer-Lindenberg, A. Remission of Major Depression Under Deep Brain Stimulation of the Lateral Habenula in a Therapy-Refractory Patient. Biol. Psychiatry 2010, 67, e9–e11. [Google Scholar] [CrossRef]
- Markovic, V.; Agosta, F.; Canu, E.; Inuggi, A.; Petrovic, I.; Stankovic, I.; Imperiale, F.; Stojkovic, T.; Kostic, V.S.; Filippi, M. Role of Habenula and Amygdala Dysfunction in Parkinson Disease Patients with Punding. Neurology 2017, 88, 2207–2215. [Google Scholar] [CrossRef]
- Sourani, D.; Eitan, R.; Gordon, N.; Goelman, G. The Habenula Couples the Dopaminergic and the Serotonergic Systems: Application to Depression in Parkinson’s Disease. Eur. J. Neurosci. 2012, 36, 2822–2829. [Google Scholar] [CrossRef]
- Breen, D.P.; Vuono, R.; Nawarathna, U.; Fisher, K.; Shneerson, J.M.; Reddy, A.B.; Barker, R.A. Sleep and Circadian Rhythm Regulation in Early Parkinson Disease. JAMA Neurol. 2014, 71, 589–595. [Google Scholar] [CrossRef]
- Videnovic, A.; Noble, C.; Reid, K.J.; Peng, J.; Turek, F.W.; Marconi, A.; Rademaker, A.W.; Simuni, T.; Zadikoff, C.; Zee, P.C. Circadian Melatonin Rhythm and Excessive Daytime Sleepiness in Parkinson Disease. JAMA Neurol. 2014, 71, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.P.; Nombela, C.; Vuono, R.; Jones, P.S.; Fisher, K.; Burn, D.J.; Brooks, D.J.; Reddy, A.B.; Rowe, J.B.; Barker, R.A. Hypothalamic Volume Loss Is Associated with Reduced Melatonin Output in Parkinson’s Disease. Mov. Disord. 2016, 31, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y.; Whone, A.L.; Brooks, D.J. Extrastriatal Monoamine Neuron Function in Parkinson’s Disease: An 18F-Dopa PET Study. Neurobiol. Dis. 2008, 29, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Pavese, N.; Moore, R.Y.; Scherfler, C.; Khan, N.L.; Hotton, G.; Quinn, N.P.; Bhatia, K.P.; Wood, N.W.; Brooks, D.J.; Lees, A.J.; et al. In Vivo Assessment of Brain Monoamine Systems in Parkin Gene Carriers: A PET Study. Exp. Neurol. 2010, 222, 120–124. [Google Scholar] [CrossRef]
- Kägi, G.; Klein, C.; Wood, N.W.; Schneider, S.A.; Pramstaller, P.P.; Tadic, V.; Quinn, N.P.; van de Warrenburg, B.P.C.; Bhatia, K.P. Nonmotor Symptoms in Parkin Gene-Related Parkinsonism. Mov. Disord. 2010, 25, 1279–1284. [Google Scholar] [CrossRef] [PubMed]
- Ghaemi, M.; Rudolf, J.; Hilker, R.; Herholz, K.; Heiss, W.D. Increased Pineal Fdopa Uptake Is Related to Severity of Parkinson’s Disease—A PET Study. J. Pineal. Res. 2001, 30, 213–219. [Google Scholar] [CrossRef]
- Floresco, S.B. The Nucleus Accumbens: An Interface between Cognition, Emotion, and Action. Annu. Rev. Psychol. 2015, 66, 25–32. [Google Scholar] [CrossRef]
- Groenewegen, H.J.; Trimble, M. The Ventral Striatum as an Interface Between the Limbic and Motor Systems. CNS Spectr. 2007, 12, 887–892. [Google Scholar] [CrossRef]
- Salgado, S.; Kaplitt, M.G. The Nucleus Accumbens: A Comprehensive Review. Stereotact. Funct. Neurosurg. 2015, 93, 75–93. [Google Scholar] [CrossRef]
- Volkow, N.D.; Michaelides, M.; Baler, R. The Neuroscience of Drug Reward and Addiction. Physiol. Rev. 2019, 99, 2115–2140. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. Is Braak Staging Valid for All Types of Parkinson’s Disease? J. Neural. Transm. 2019, 126, 423–431. [Google Scholar] [CrossRef]
- Stark, A.J.; Smith, C.T.; Lin, Y.C.; Petersen, K.J.; Trujillo, P.; van Wouwe, N.C.; Kang, X.H.; Donahue, M.J.; Kessler, R.M.; Zald, D.H.; et al. Nigrostriatal and Mesolimbic D2/3 Receptor Expression in Parkinson’s Disease Patients with Compulsive Reward-Driven Behaviors. J. Neurosci. 2018, 38, 3230–3239. [Google Scholar] [CrossRef] [PubMed]
- Pagano, G.; Molloy, S.; Bain, P.G.; Rabiner, E.A.; Ray Chaudhuri, K.; Brooks, D.J.; Pavese, N. Impulse Control Disorders Are Associated with Lower Ventral Striatum Dopamine D3 Receptor Availability in Parkinson’s Disease: A [11C]-PHNO PET Study. Parkinsonism Relat. Disord. 2021, 90, 52–56. [Google Scholar] [CrossRef]
- Navalpotro-Gomez, I.; Dacosta-Aguayo, R.; Molinet-Dronda, F.; Martin-Bastida, A.; Botas-Peñin, A.; Jimenez-Urbieta, H.; Delgado-Alvarado, M.; Gago, B.; Quiroga-Varela, A.; Rodriguez-Oroz, M.C. Nigrostriatal Dopamine Transporter Availability, and Its Metabolic and Clinical Correlates in Parkinson’s Disease Patients with Impulse Control Disorders. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2065–2076. [Google Scholar] [CrossRef]
- Theis, H.; Probst, C.; Fernagut, P.O.; van Eimeren, T. Unlucky Punches: The Vulnerability-Stress Model for the Development of Impulse Control Disorders in Parkinson’s Disease. npj Parkinson’s Disease 2021, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Paz-Alonso, P.M.; Navalpotro-Gomez, I.; Boddy, P.; Dacosta-Aguayo, R.; Delgado-Alvarado, M.; Quiroga-Varela, A.; Jimenez-Urbieta, H.; Carreiras, M.; Rodriguez-Oroz, M.C. Functional Inhibitory Control Dynamics in Impulse Control Disorders in Parkinson’s Disease. Mov. Disord. 2020, 35, 316–325. [Google Scholar] [CrossRef]
- Theis, H.; Probst, C.; Campabadal, A.; Goerlich, K.S.; Granert, O.; Wolff, S.; Witt, K.; Deuschl, G.; Van Eimeren, T. Inhibitory Framing in Hypersexual Patients with Parkinson’s Disease. An FMRI Pilot Study. Exp. Brain Res. 2022, 240, 2097–2107. [Google Scholar] [CrossRef]
- Mosley, P.E.; Paliwal, S.; Robinson, K.; Coyne, T.; Silburn, P.; Tittgemeyer, M.; Stephan, K.E.; Breakspear, M.; Perry, A. The Structural Connectivity of Discrete Networks Underlies Impulsivity and Gambling in Parkinson’s Disease. Brain 2019, 142, 3917–3935. [Google Scholar] [CrossRef]
- Mosley, P.E.; Paliwal, S.; Robinson, K.; Coyne, T.; Silburn, P.; Tittgemeyer, M.; Stephan, K.E.; Perry, A.; Breakspear, M. The Structural Connectivity of Subthalamic Deep Brain Stimulation Correlates with Impulsivity in Parkinson’s Disease. Brain 2020, 143, 2235–2254. [Google Scholar] [CrossRef]
- du Plessis, S.; Bossert, M.; Vink, M.; van den Heuvel, L.; Bardien, S.; Emsley, R.; Buckle, C.; Seedat, S.; Carr, J. Reward Processing Dysfunction in Ventral Striatum and Orbitofrontal Cortex in Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 48, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Hiebert, N.M.; Owen, A.M.; Ganjavi, H.; Mendonça, D.; Jenkins, M.E.; Seergobin, K.N.; MacDonald, P.A. Dorsal Striatum Does Not Mediate Feedback-Based, Stimulus-Response Learning: An Event-Related FMRI Study in Patients with Parkinson’s Disease Tested on and off Dopaminergic Therapy. Neuroimage 2019, 185, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Marín-Lahoz, J.; Martínez-Horta, S.; Sampedro, F.; Pagonabarraga, J.; Horta-Barba, A.; Bejr-kasem, H.; Botí, M.; Fernández-Bobadilla, R.; Pascual-Sedano, B.; Pérez-Pérez, J.; et al. Measuring Impulsivity in Parkinson’s Disease: A Correlational and Structural Neuroimaging Study Using Different Tests. Eur. J. Neurol. 2020, 27, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Ye, B.S.; Jeon, S.; Yoon, S.; Kang, S.W.; Baik, K.W.; Lee, Y.; Chung, S.J.; Oh, J.S.; Moon, H.; Kim, J.S.; et al. Effects of Dopaminergic Depletion and Brain Atrophy on Neuropsychiatric Symptoms in de Novo Parkinson’s Disease. J. Neurol. Neurosurg. Psychiatry 2018, 89, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Sacheli, M.A.; Murray, D.K.; Vafai, N.; Cherkasova, M.V.; Dinelle, K.; Shahinfard, E.; Neilson, N.; McKenzie, J.; Schulzer, M.; Appel-Cresswell, S.; et al. Habitual Exercisers versus Sedentary Subjects with Parkinson’s Disease: Multimodal PET and FMRI Study. Mov. Disord. 2018, 33, 1945–1950. [Google Scholar] [CrossRef]
- Sacheli, M.A.; Neva, J.L.; Lakhani, B.; Murray, D.K.; Vafai, N.; Shahinfard, E.; English, C.; McCormick, S.; Dinelle, K.; Neilson, N.; et al. Exercise Increases Caudate Dopamine Release and Ventral Striatal Activation in Parkinson’s Disease. Mov. Disord. 2019, 34, 1891–1900. [Google Scholar] [CrossRef]
- Kawashima, S.; Ueki, Y.; Kato, T.; Ito, K.; Matsukawa, N. Reduced Striatal Dopamine Release during Motor Skill Acquisition in Parkinson’s Disease. PLoS ONE 2018, 13, e0196661. [Google Scholar] [CrossRef]
- Yoo, S.W.; Oh, Y.S.; Hwang, E.J.; Ryu, D.W.; Lee, K.S.; Lyoo, C.H.; Kim, J.S. “Depressed” Caudate and Ventral Striatum Dopamine Transporter Availability in de Novo Depressed Parkinson’s Disease. Neurobiol. Dis. 2019, 132, 104563. [Google Scholar] [CrossRef]
- Kinugawa, K.; Mano, T.; Yamatani, Y.; Miyasaka, T.; Kataoka, H.; Sugie, K. Pain-Related Abnormal Neuronal Synchronization of the Nucleus Accumbens in Parkinson’s Disease. Brain Sci. 2022, 12, 84. [Google Scholar] [CrossRef]
- Chung, S.J.; Yoo, H.S.; Oh, J.S.; Kim, J.S.; Ye, B.S.; Sohn, Y.H.; Lee, P.H. Effect of Striatal Dopamine Depletion on Cognition in de Novo Parkinson’s Disease. Parkinsonism Relat. Disord. 2018, 51, 43–48. [Google Scholar] [CrossRef]
- Oh, Y.S.; Yoo, S.W.; Lyoo, C.H.; Yoo, J.Y.; Yoon, H.; Ha, S.; Lee, K.S.; Kim, J.S. The Association of β-Amyloid with Cognition and Striatal Dopamine in Early, Non-Demented Parkinson’s Disease. J. Parkinsons Dis. 2021, 11, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Chung, S.J.; Lee, S.; Cha, J.; Sohn, Y.H.; Yun, M.; Lee, P.H. Dysautonomia Is Linked to Striatal Dopamine Deficits and Regional Cerebral Perfusion in Early Parkinson Disease. Clin. Nucl. Med. 2020, 45, e342–e348. [Google Scholar] [CrossRef]
- Valli, M.; Cho, S.S.; Uribe, C.; Masellis, M.; Chen, R.; Mihaescu, A.; Strafella, A.P. VMAT2 Availability in Parkinson’s Disease with Probable REM Sleep Behaviour Disorder. Mol. Brain 2021, 14, 165. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Eickhoff, S.B.; Hoffstaedter, F.; Genon, S.; Caspers, S.; Reetz, K.; Dogan, I.; Eickhoff, C.R.; Chen, J.; Caspers, J.; et al. Joint Multi-Modal Parcellation of the Human Striatum: Functions and Clinical Relevance. Neurosci. Bull. 2020, 36, 1123–1136. [Google Scholar] [CrossRef]
- Khan, A.R.; Hiebert, N.M.; Vo, A.; Wang, B.T.; Owen, A.M.; Seergobin, K.N.; MacDonald, P.A. Biomarkers of Parkinson’s Disease: Striatal Sub-Regional Structural Morphometry and Diffusion MRI. Neuroimage Clin. 2019, 21, 101597. [Google Scholar] [CrossRef]
- Rolls, E.T. Emotion and Decision-Making Explained: A Précis. Cortex 2014, 59, 185–193. [Google Scholar] [CrossRef]
- Weintraub, D.; David, A.S.; Evans, A.H.; Grant, J.E.; Stacy, M. Clinical Spectrum of Impulse Control Disorders in Parkinson’s Disease. Mov. Disord. 2015, 30, 121–127. [Google Scholar] [CrossRef]
- Mazzoni, P.; Hristova, A.; Krakauer, J.W. Why Don’t We Move Faster? Parkinson’s Disease, Movement Vigor, and Implicit Motivation. J. Neurosci. 2007, 27, 7105–7116. [Google Scholar] [CrossRef]
- Baraduc, P.; Thobois, S.; Gan, J.; Broussolle, E.; Desmurget, M. A Common Optimization Principle for Motor Execution in Healthy Subjects and Parkinsonian Patients. J. Neurosci. 2013, 33, 665–677. [Google Scholar] [CrossRef]
- Le Bouc, R.; Rigoux, L.; Schmidt, L.; Degos, B.; Welter, M.L.; Vidailhet, M.; Daunizeau, J.; Pessiglione, M. Computational Dissection of Dopamine Motor and Motivational Functions in Humans. J. Neurosci. 2016, 36, 6623–6633. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banwinkler, M.; Theis, H.; Prange, S.; van Eimeren, T. Imaging the Limbic System in Parkinson’s Disease—A Review of Limbic Pathology and Clinical Symptoms. Brain Sci. 2022, 12, 1248. https://doi.org/10.3390/brainsci12091248
Banwinkler M, Theis H, Prange S, van Eimeren T. Imaging the Limbic System in Parkinson’s Disease—A Review of Limbic Pathology and Clinical Symptoms. Brain Sciences. 2022; 12(9):1248. https://doi.org/10.3390/brainsci12091248
Chicago/Turabian StyleBanwinkler, Magdalena, Hendrik Theis, Stéphane Prange, and Thilo van Eimeren. 2022. "Imaging the Limbic System in Parkinson’s Disease—A Review of Limbic Pathology and Clinical Symptoms" Brain Sciences 12, no. 9: 1248. https://doi.org/10.3390/brainsci12091248