
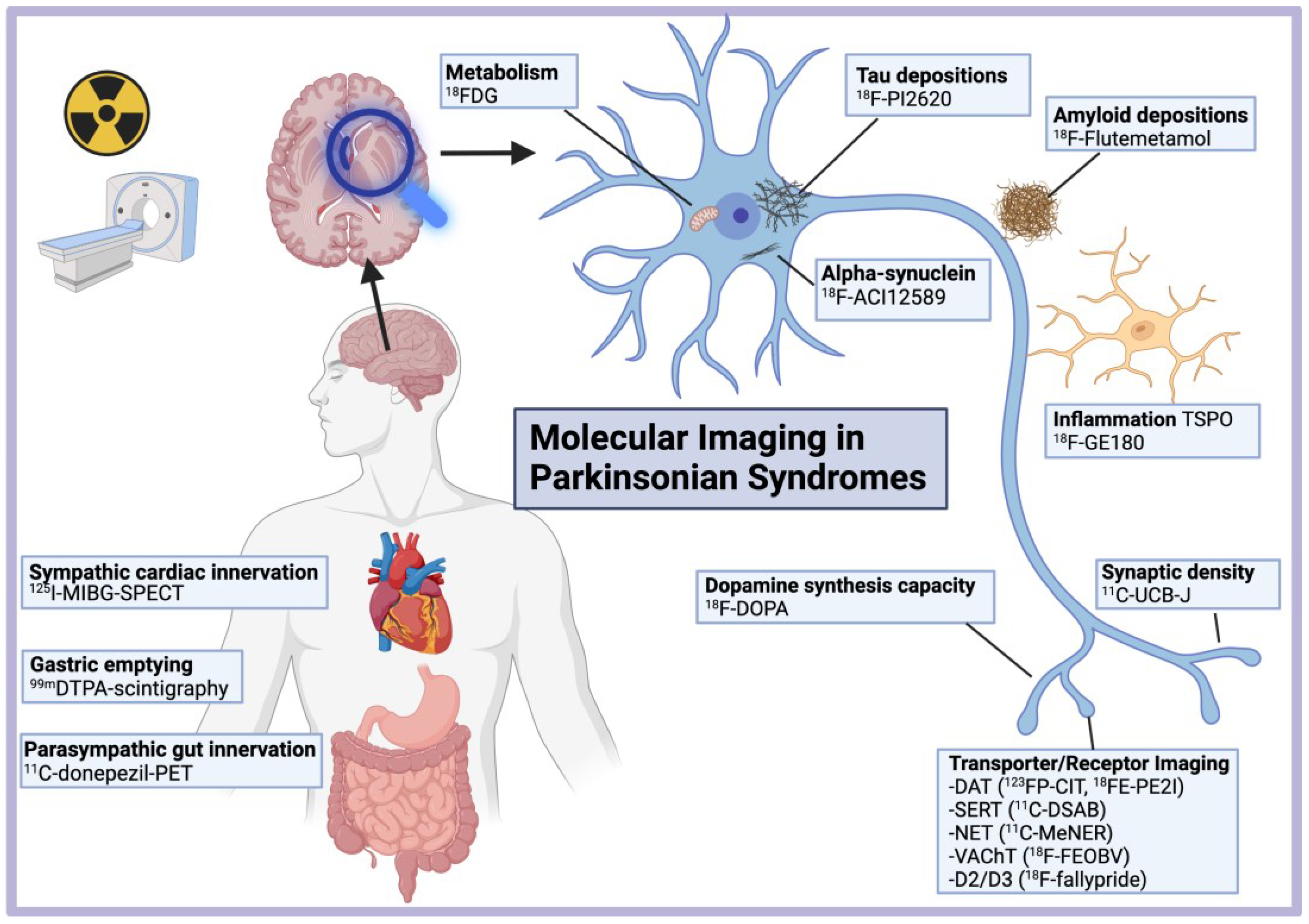
Molecular Imaging in Parkinsonian Disorders—What’s New and Hot?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Highlights
- Imaging of the dopamine transporter (DAT) (in particular using 11C- or 18F-PE2I) most reliably and precisely measures neurodegeneration of the dopaminergic system in parkinsonian disorders and its progression in longitudinal studies.
- Combining DAT imaging and iron- and neuromelanin-sensitive MR imaging in vivo unveiled the sequential progression of neurodegeneration along the sensorimotor, associative, and limbic territories in the striatum followed by the substantia nigra in Parkinson’s disease.
- DAT imaging is the most accurate marker for prodromal PD and predicts phenoconversion to clinical motor symptoms in patients with REM-sleep behavior disorders (RBD).
- Dopaminergic imaging can be used to investigate motor reserve in PD patients.
- Peripheral molecular imaging shows that autonomous denervation of the heart and gut in prodromal and de novo PD with RBD precedes striatal dopaminergic loss, thereby supporting peripheral starting points in those patients.
- Serotonergic denervation in the subcortical and cortical limbic regions is associated with apathy, depression, and anxiety in PD.
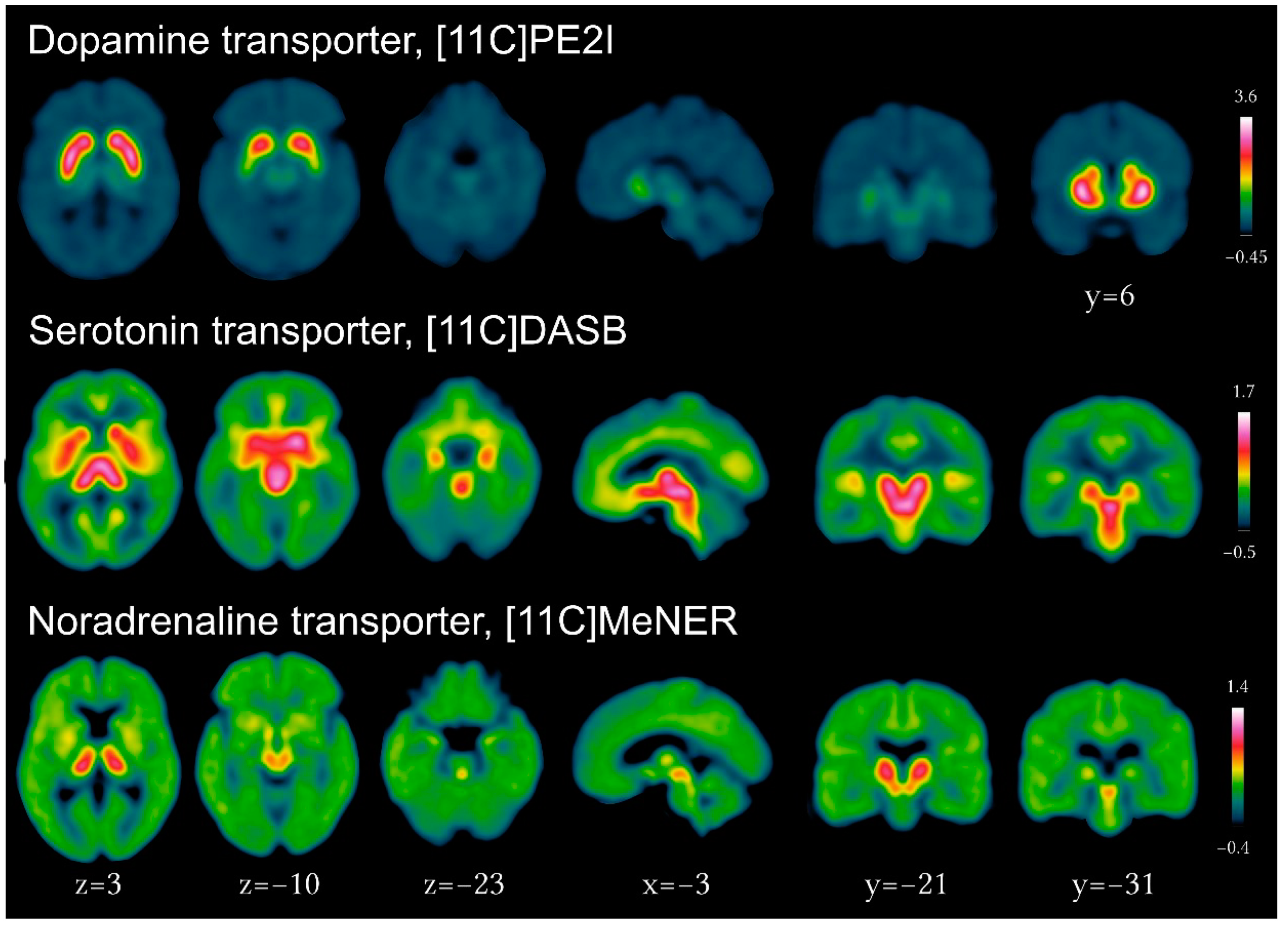
- Noradrenergic terminals loss plays a critical role in RBD and was shown to exceed locus coeruleus pathology in PD using 11C-MeNER PET.
- Cholinergic imaging is now complemented by the new PET tracer 18F-FEOBV, which binds to presynaptic transporters, particularly in subcortical structures.
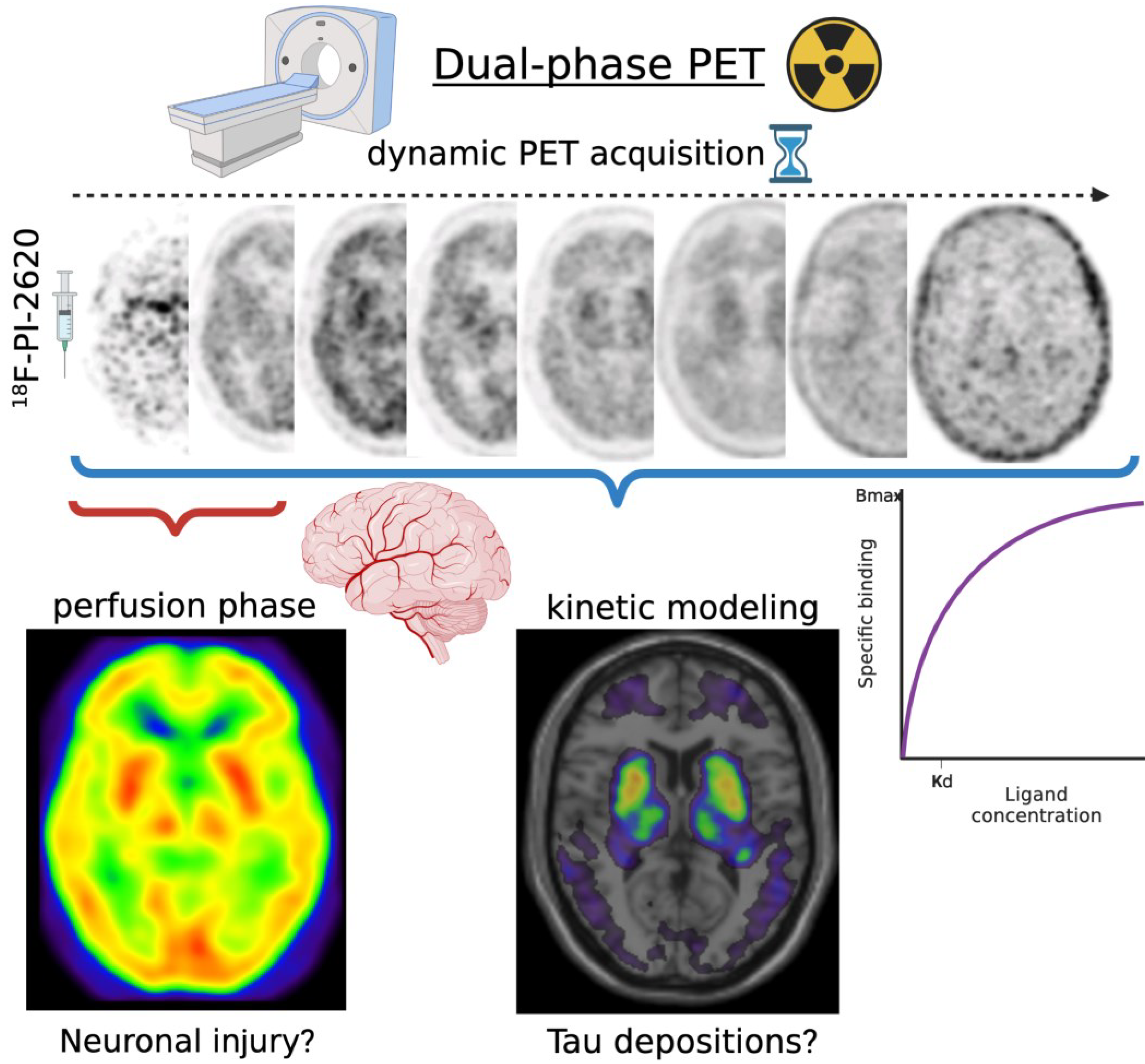
- Dual-phase dynamic PET imaging may enable to inform both metabolic and molecular disease-related patterns with a single injection, including tracers for DAT or tau.
- Using 11C-UCB-J, prominent loss of synaptic density in the substantia nigra and brainstem was demonstrated in PD, even in the early stage, extending to the cortex in DLB, as well as PSP and CBD.
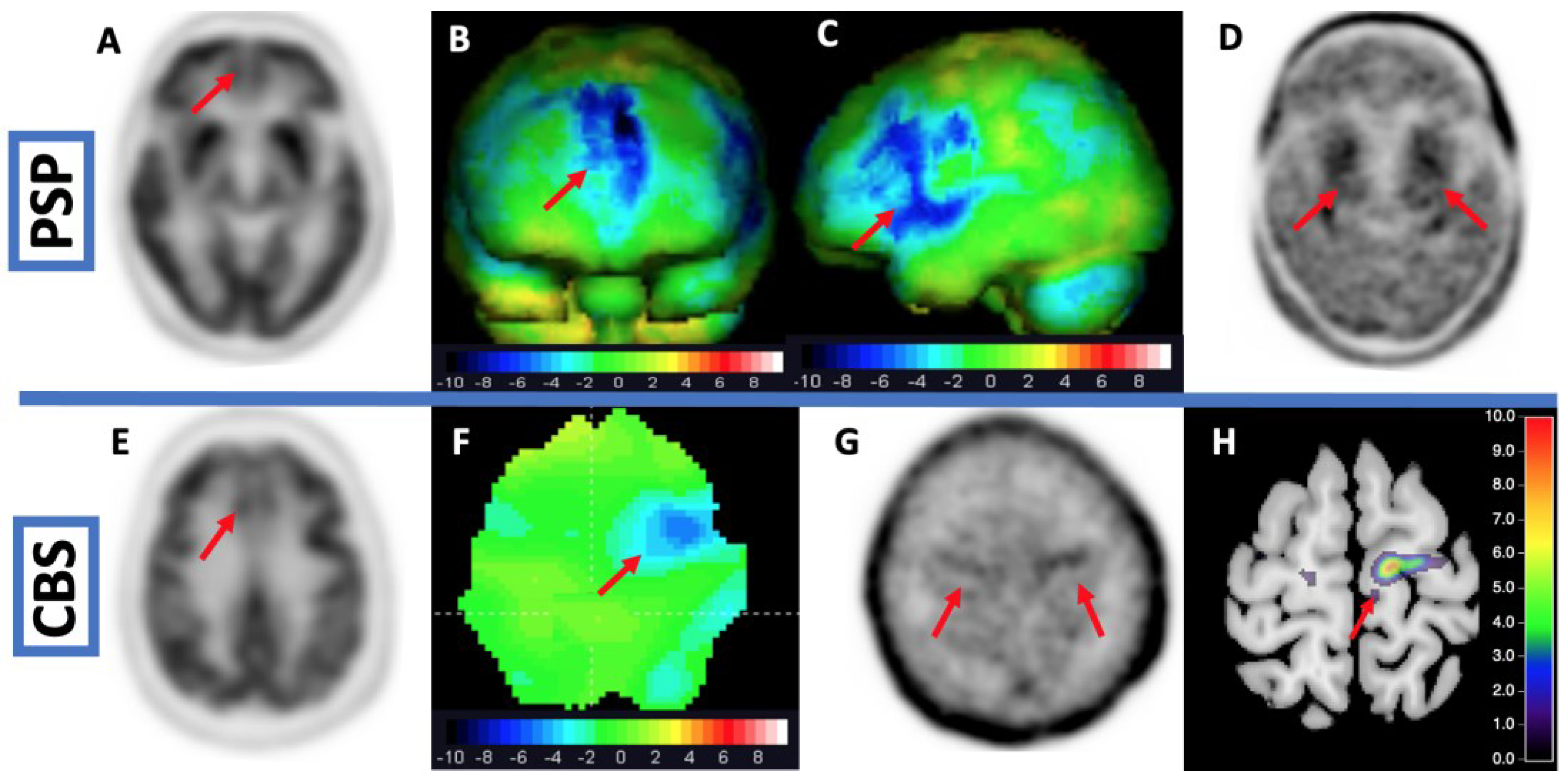
- Tau imaging using second-generation tracers with increased specificity greatly improved the diagnosis of tau aggregation in PSP and CBD.
- Despite recent advances and promising results in MSA patients using 18F-ACI12589 PET, no tracer is readily available to image α-synuclein aggregates in vivo in parkinsonian disorders.
- Computational modeling of tracer affinity in silico, coupled with cryogenic electron microscopy will improve efficiency and accelerate the development of novel tracers.
Abstract
1. Introduction
2. Materials and Methods
3. Neurotransmitter Imaging
3.1. Dopaminergic Imaging
3.1.1. Imaging the Dopaminergic System
3.1.2. Dopaminergic Imaging in PD
3.1.3. Dopaminergic Imaging in Genetic PD
3.1.4. Dopaminergic Imaging in Prodromal PD
3.1.5. Dopaminergic Imaging in Atypical Parkinsonian Disorders
3.1.6. Differential Diagnosis Using Dopaminergic Imaging and Use of Machine Learning
3.1.7. Implications of Advances in Dopaminergic Imaging
3.2. Imaging of Peripheral Neurotransmitter in Parkinsonian Disorders
3.3. Serotonergic Imaging in PD and Parkinsonian Disorders
3.4. Noradrenergic Imaging in PD and RBD
3.5. Cholinergic Imaging in PD
4. Imaging of Metabolism, Synaptic Density and Inflammation
4.1. Metabolic Imaging in Parkinsonian Disorders
4.2. Disease-Related Patterns Using Metabolic Imaging
4.3. Imaging of Synaptic Density
4.4. Imaging of Inflammation
5. Imaging of Protein Aggregates
5.1. Second Generation Tau Imaging
5.2. Imaging of α-Synuclein
6. New Perspectives for Tracer Development and Technical Advances
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peralta, C.; Strafella, A.P.; van Eimeren, T.; Ceravolo, R.; Seppi, K.; Kaasinen, V.; Arena, J.E.; Lehericy, S.; International Parkinson Movement Disorders Society-Neuroimaging Study Group. Pragmatic Approach on Neuroimaging Techniques for the Differential Diagnosis of Parkinsonisms. Mov. Disord. Clin. Pract. 2022, 9, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Strafella, A.P.; Bohnen, N.I.; Perlmutter, J.S.; Eidelberg, D.; Pavese, N.; Van Eimeren, T.; Piccini, P.; Politis, M.; Thobois, S.; Ceravolo, R.; et al. Molecular Imaging to Track Parkinson’s Disease and Atypical Parkinsonisms: New Imaging Frontiers. Mov. Disord. 2017, 32, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Morbelli, S.; Esposito, G.; Arbizu, J.; Barthel, H.; Boellaard, R.; Bohnen, N.I.; Brooks, D.J.; Darcourt, J.; Dickson, J.C.; Douglas, D.; et al. EANM Practice Guideline/SNMMI Procedure Standard for Dopaminergic Imaging in Parkinsonian Syndromes 1.0. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 1885–1912. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Zhang, C.; Liu, K.; Wagle Shukla, A.; Sun, B.; Guan, Y. Imaging of Dopamine Transporters in Parkinson Disease: A Meta-Analysis of 18F/123I-FP-CIT Studies. Ann. Clin. Transl. Neurol. 2020, 7, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Kramer, V.; Juri, C.; Riss, P.J.; Pruzzo, R.; Soza-Ried, C.; Flores, J.; Hurtado, A.; Rösch, F.; Chana-Cuevas, P.; Amaral, H. Pharmacokinetic Evaluation of [18F]PR04.MZ for PET/CT Imaging and Quantification of Dopamine Transporters in the Human Brain. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 1927–1937. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, C.; Tang, J.; Xu, Y.; Xie, M.; Chen, Z. An Efficient Automated Radiosynthesis and Bioactivity Confirmation of VMAT2 Tracer [18F]FP-(+)-DTBZ. Mol. Imaging Biol. 2020, 22, 265–273. [Google Scholar] [CrossRef]
- Delva, A.; Van Weehaeghe, D.; van Aalst, J.; Ceccarini, J.; Koole, M.; Baete, K.; Nuyts, J.; Vandenberghe, W.; Van Laere, K. Quantification and Discriminative Power of 18F-FE-PE2I PET in Patients with Parkinson’s Disease. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 1913–1926. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Takahata, K.; Kubota, M.; Takano, H.; Takeuchi, H.; Kimura, Y.; Sano, Y.; Kurose, S.; Ito, H.; Mimura, M.; et al. Differential Associations of Dopamine Synthesis Capacity with the Dopamine Transporter and D2 Receptor Availability as Assessed by PET in the Living Human Brain. NeuroImage 2021, 226, 117543. [Google Scholar] [CrossRef]
- Biondetti, E.; Gaurav, R.; Yahia-Cherif, L.; Mangone, G.; Pyatigorskaya, N.; Valabrègue, R.; Ewenczyk, C.; Hutchison, M.; François, C.; Arnulf, I.; et al. Spatiotemporal Changes in Substantia Nigra Neuromelanin Content in Parkinson’s Disease. Brain 2020, 143, 2757–2770. [Google Scholar] [CrossRef]
- Uchida, Y.; Kan, H.; Sakurai, K.; Inui, S.; Kobayashi, S.; Akagawa, Y.; Shibuya, K.; Ueki, Y.; Matsukawa, N. Magnetic Susceptibility Associates With Dopaminergic Deficits and Cognition in Parkinson’s Disease. Mov. Disord. 2020, 35, 1396–1405. [Google Scholar] [CrossRef]
- Martín-Bastida, A.; Lao-Kaim, N.P.; Roussakis, A.A.; Searle, G.E.; Xing, Y.; Gunn, R.N.; Schwarz, S.T.; Barker, R.A.; Auer, D.P.; Piccini, P. Relationship between Neuromelanin and Dopamine Terminals within the Parkinson’s Nigrostriatal System. Brain 2019, 142, 2023–2036. [Google Scholar] [CrossRef]
- Yang, J.; Archer, D.B.; Burciu, R.G.; Müller, M.L.T.M.; Roy, A.; Ofori, E.; Bohnen, N.I.; Albin, R.L.; Vaillancourt, D.E. Multimodal Dopaminergic and Free-Water Imaging in Parkinson’s Disease. Parkinsonism Relat. Disord. 2019, 62, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.F.; Klyuzhin, I.; McKenzie, J.; Neilson, N.; Shahinfard, E.; Dinelle, K.; McKeown, M.J.; Stoessl, A.J.; Sossi, V. Joint Pattern Analysis Applied to PET DAT and VMAT2 Imaging Reveals New Insights into Parkinson’s Disease Induced Presynaptic Alterations. NeuroImage Clin. 2019, 23, 101856. [Google Scholar] [CrossRef]
- Roussakis, A.-A.; Zeng, Z.; Lao-Kaim, N.P.; Martin-Bastida, A.; Piccini, P.; Barker, R.A.; Farrell, K.; Guzman, N.V.; He, X.; Lazic, S.E.; et al. Parkinson’s Disease Laterality: A 11C-PE2I PET Imaging Study. J. Neurol. 2021, 268, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Pak, K.; Kim, H.-K.; Nudelman, K.N.; Kim, J.H.; Kim, Y.H.; Kang, J.; Baek, M.S.; Lyoo, C.H. Genetic Factors Affecting Dopaminergic Deterioration during the Premotor Stage of Parkinson Disease. Npj Park. Dis. 2021, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.P.; Yang, B.; Rahmim, A.; Tang, J. Prediction of Outcome in Parkinson’s Disease Patients from DAT SPECT Images Using a Convolutional Neural Network. In Proceedings of the 2018 IEEE Nuclear Science Symposium and Medical Imaging Conference Proceedings (NSS/MIC), Sydney, NSW, Australia, 10–17 November 2018; pp. 1–4. [Google Scholar]
- Salmanpour, M.R.; Shamsaei, M.; Saberi, A.; Klyuzhin, I.S.; Tang, J.; Sossi, V.; Rahmim, A. Machine Learning Methods for Optimal Prediction of Motor Outcome in Parkinson’s Disease. Phys. Med. 2020, 69, 233–240. [Google Scholar] [CrossRef]
- Tang, J.; Yang, B.; Adams, M.P.; Shenkov, N.N.; Klyuzhin, I.S.; Fotouhi, S.; Davoodi-Bojd, E.; Lu, L.; Soltanian-Zadeh, H.; Sossi, V.; et al. Artificial Neural Network-Based Prediction of Outcome in Parkinson’s Disease Patients Using DaTscan SPECT Imaging Features. Mol. Imaging Biol. 2019, 21, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, T.; Pagano, G.; Niccolini, F.; Politis, M. Predicting Cognitive Decline with Non-Clinical Markers in Parkinson’s Disease (PRECODE-2). J. Neurol. 2019, 266, 1203–1210. [Google Scholar] [CrossRef]
- Ye, R.; Locascio, J.J.; Goodheart, A.E.; Quan, M.; Zhang, B.; Gomperts, S.N. Serum NFL Levels Predict Progression of Motor Impairment and Reduction in Putamen Dopamine Transporter Binding Ratios in de Novo Parkinson’s Disease: An 8-Year Longitudinal Study. Parkinsonism Relat. Disord. 2021, 85, 11–16. [Google Scholar] [CrossRef]
- Boonstra, J.T.; Michielse, S.; Temel, Y.; Hoogland, G.; Jahanshahi, A. Neuroimaging Detectable Differences between Parkinson’s Disease Motor Subtypes: A Systematic Review. Mov. Disord. Clin. Pract. 2021, 8, 175–192. [Google Scholar] [CrossRef]
- Chung, S.J.; Kim, H.-R.; Jung, J.H.; Lee, P.H.; Jeong, Y.; Sohn, Y.H. Identifying the Functional Brain Network of Motor Reserve in Early Parkinson’s Disease. Mov. Disord. 2020, 35, 577–586. [Google Scholar] [CrossRef]
- Sacheli, M.A.; Neva, J.L.; Lakhani, B.; Murray, D.K.; Vafai, N.; Shahinfard, E.; English, C.; McCormick, S.; Dinelle, K.; Neilson, N.; et al. Exercise Increases Caudate Dopamine Release and Ventral Striatal Activation in Parkinson’s Disease. Mov. Disord. 2019, 34, 1891–1900. [Google Scholar] [CrossRef]
- Sambin, S.; Lavisse, S.; Decaix, C.; Pyatigorskaya, N.; Mangone, G.; Valabrègue, R.; Arnulf, I.; Cormier, F.; Lesage, S.; Lehericy, S.; et al. Compensatory Mechanisms Nine Years Before Parkinson’s Disease Conversion in a LRRK2 R1441H Family. Mov. Disord. 2022, 37, 428–430. [Google Scholar] [CrossRef]
- Simuni, T.; Uribe, L.; Cho, H.R.; Caspell-Garcia, C.; Coffey, C.S.; Siderowf, A.; Trojanowski, J.Q.; Shaw, L.M.; Seibyl, J.; Singleton, A.; et al. Clinical and Dopamine Transporter Imaging Characteristics of Non-Manifest LRRK2 and GBA Mutation Carriers in the Parkinson’s Progression Markers Initiative (PPMI): A Cross-Sectional Study. Lancet Neurol. 2020, 19, 71–80. [Google Scholar] [CrossRef]
- Wile, D.J.; Agarwal, P.A.; Schulzer, M.; Mak, E.; Dinelle, K.; Shahinfard, E.; Vafai, N.; Hasegawa, K.; Zhang, J.; McKenzie, J.; et al. Serotonin and Dopamine Transporter PET Changes in the Premotor Phase of LRRK2 Parkinsonism: Cross-Sectional Studies. Lancet Neurol. 2017, 16, 351–359. [Google Scholar] [CrossRef]
- Droby, A.; Artzi, M.; Lerman, H.; Hutchison, R.M.; Bashat, D.B.; Omer, N.; Gurevich, T.; Orr-Urtreger, A.; Cohen, B.; Cedarbaum, J.M.; et al. Aberrant Dopamine Transporter and Functional Connectivity Patterns in LRRK2 and GBA Mutation Carriers. Npj Park. Dis. 2022, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Pont-Sunyer, C.; Tolosa, E.; Caspell-Garcia, C.; Coffey, C.; Alcalay, R.N.; Chan, P.; Duda, J.E.; Facheris, M.; Fernández-Santiago, R.; Marek, K.; et al. The Prodromal Phase of Leucine-Rich Repeat Kinase 2–Associated Parkinson Disease: Clinical and Imaging Studies. Mov. Disord. 2017, 32, 726–738. [Google Scholar] [CrossRef]
- Lopez, G.; Eisenberg, D.P.; Gregory, M.D.; Ianni, A.M.; Grogans, S.E.; Masdeu, J.C.; Kim, J.; Groden, C.; Sidransky, E.; Berman, K.F. Longitudinal Positron Emission Tomography of Dopamine Synthesis in Subjects with GBA1 Mutations. Ann. Neurol. 2020, 87, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, A.; Martínez-Rodríguez, I.; Sánchez-Juan, P.; Sierra, M.; González-Aramburu, I.; Rivera-Sánchez, M.; Andrés-Pacheco, J.; Gutierrez-González, Á.; García-Hernández, A.; Madera, J.; et al. Serial DaT-SPECT Imaging in Asymptomatic Carriers of LRRK2 G2019S Mutation: 8 Years’ Follow-Up. Eur. J. Neurol. 2021, 28, 4204–4208. [Google Scholar] [CrossRef] [PubMed]
- Sierra, M.; Martínez-Rodríguez, I.; Sánchez-Juan, P.; González-Aramburu, I.; Jiménez-Alonso, M.; Sánchez-Rodríguez, A.; Berciano, J.; Banzo, I.; Infante, J. Prospective Clinical and DaT-SPECT Imaging in Premotor LRRK2 G2019S-Associated Parkinson Disease. Neurology 2017, 89, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simuni, T.; Brumm, M.C.; Uribe, L.; Caspell-Garcia, C.; Coffey, C.S.; Siderowf, A.; Alcalay, R.N.; Trojanowski, J.Q.; Shaw, L.M.; Seibyl, J.; et al. Clinical and Dopamine Transporter Imaging Characteristics of Leucine Rich Repeat Kinase 2 (LRRK2) and Glucosylceramidase Beta (GBA) Parkinson’s Disease Participants in the Parkinson’s Progression Markers Initiative: A Cross-Sectional Study. Mov. Disord. 2020, 35, 833–844. [Google Scholar] [CrossRef]
- Greuel, A.; Trezzi, J.-P.; Glaab, E.; Ruppert, M.C.; Maier, F.; Jäger, C.; Hodak, Z.; Lohmann, K.; Ma, Y.; Eidelberg, D.; et al. GBA Variants in Parkinson’s Disease: Clinical, Metabolomic, and Multimodal Neuroimaging Phenotypes. Mov. Disord. 2020, 35, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Schindlbeck, K.A.; Vo, A.; Nguyen, N.; Tang, C.C.; Niethammer, M.; Dhawan, V.; Brandt, V.; Saunders-Pullman, R.; Bressman, S.B.; Eidelberg, D. LRRK2 and GBA Variants Exert Distinct Influences on Parkinson’s Disease-Specific Metabolic Networks. Cereb. Cortex 2020, 30, 2867–2878. [Google Scholar] [CrossRef] [PubMed]
- Pak, K.; Lee, M.J.; Kim, K.; Kim, I.J. No Effect of Parkinson’s Disease-Polygenic Load on Striatal Density of Dopaminergic Neuron in Healthy Subjects. Ann. Nucl. Med. 2021, 35, 1187–1192. [Google Scholar] [CrossRef]
- Shin, S.; Kim, K.; Lee, J.M.; Kim, E.J.; Kim, S.J.; Kim, I.J.; Pak, K.; Lee, M.J. Effect of Single-Nucleotide Polymorphisms on Decline of Dopamine Transporter Availability in Parkinson’s Disease. J. Clin. Neurol. Seoul Korea 2019, 15, 102–107. [Google Scholar] [CrossRef]
- Tan, M.M.X.; Lawton, M.A.; Jabbari, E.; Reynolds, R.H.; Iwaki, H.; Blauwendraat, C.; Kanavou, S.; Pollard, M.I.; Hubbard, L.; Malek, N.; et al. Genome-Wide Association Studies of Cognitive and Motor Progression in Parkinson’s Disease. Mov. Disord. 2021, 36, 424–433. [Google Scholar] [CrossRef]
- Valli, M.; Cho, S.S.; Masellis, M.; Chen, R.; Rusjan, P.; Kim, J.; Koshimori, Y.; Mihaescu, A.; Strafella, A.P. DRD2 Genotype-Based Variants Modulates D2 Receptor Distribution in Ventral Striatum. Mol. Neurobiol. 2019, 56, 6512–6520. [Google Scholar] [CrossRef]
- Heinzel, S.; Berg, D.; Gasser, T.; Chen, H.; Yao, C.; Postuma, R.B. Update of the MDS Research Criteria for Prodromal Parkinson’s Disease. Mov. Disord. 2019, 34, 1464–1470. [Google Scholar] [CrossRef]
- Arnaldi, D.; Chincarini, A.; Hu, M.T.; Sonka, K.; Boeve, B.; Miyamoto, T.; Puligheddu, M.; De Cock, V.C.; Terzaghi, M.; Plazzi, G.; et al. Dopaminergic Imaging and Clinical Predictors for Phenoconversion of REM Sleep Behaviour Disorder. Brain 2021, 144, 278–287. [Google Scholar] [CrossRef]
- Chahine, L.M.; Brumm, M.C.; Caspell-Garcia, C.; Oertel, W.; Mollenhauer, B.; Amara, A.; Fernandez-Arcos, A.; Tolosa, E.; Simonet, C.; Hogl, B.; et al. Dopamine Transporter Imaging Predicts Clinically-Defined α-Synucleinopathy in REM Sleep Behavior Disorder. Ann. Clin. Transl. Neurol. 2021, 8, 201–212. [Google Scholar] [CrossRef]
- Ganapathy, S.R.; Levová, K.; Kotačková, L.; Trnka, J.; Zogala, D.; Rusz, J.; Zima, T.; Devos, D.; Šonka, K.; Růžička, E.; et al. Increased Transferrin Sialylation Predicts Phenoconversion in Isolated REM Sleep Behavior Disorder. Mov. Disord. 2022, 37, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Miyamoto, M.; Numahata, K.; Onoue, H.; Akaiwa, Y.; Sairenchi, T. Reduced Dopamine Transporter Binding Predicts Early Transition to Lewy Body Disease in Japanese Patients with Idiopathic Rapid Eye Movement Sleep Behavior Disorder. J. Neurol. Sci. 2020, 414, 116821. [Google Scholar] [CrossRef] [PubMed]
- Siderowf, A.; Jennings, D.; Stern, M.; Seibyl, J.; Eberly, S.; Oakes, D.; Marek, K.; PARS Investigators. Clinical and Imaging Progression in the PARS Cohort: Long-Term Follow-Up. Mov. Disord. 2020, 35, 1550–1557. [Google Scholar] [CrossRef]
- Postuma, R.B.; Iranzo, A.; Hu, M.; Högl, B.; Boeve, B.F.; Manni, R.; Oertel, W.H.; Arnulf, I.; Ferini-Strambi, L.; Puligheddu, M.; et al. Risk and Predictors of Dementia and Parkinsonism in Idiopathic REM Sleep Behaviour Disorder: A Multicentre Study. Brain 2019, 142, 744–759. [Google Scholar] [CrossRef]
- Janzen, A.; Kogan, R.V.; Meles, S.K.; Sittig, E.; Renken, R.J.; Geibl, F.F.; Booij, J.; Stormezand, G.; Luster, M.; Mayer, G.; et al. Rapid Eye Movement Sleep Behavior Disorder: Abnormal Cardiac Image and Progressive Abnormal Metabolic Brain Pattern. Mov. Disord. 2022, 37, 624–629. [Google Scholar] [CrossRef]
- Kogan, R.V.; Janzen, A.; Meles, S.K.; Sittig, E.; Renken, R.J.; Gurvits, V.; Mayer, G.; Leenders, K.L.; Oertel, W.H.; REMPET Working Group. Four-Year Follow-up of [18F]Fluorodeoxyglucose Positron Emission Tomography–Based Parkinson’s Disease–Related Pattern Expression in 20 Patients with Isolated Rapid Eye Movement Sleep Behavior Disorder Shows Prodromal Progression. Mov. Disord. 2021, 36, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Barber, T.R.; Griffanti, L.; Bradley, K.M.; McGowan, D.R.; Lo, C.; Mackay, C.E.; Hu, M.T.; Klein, J.C. Nigrosome 1 Imaging in REM Sleep Behavior Disorder and Its Association with Dopaminergic Decline. Ann. Clin. Transl. Neurol. 2020, 7, 26–35. [Google Scholar] [CrossRef]
- Kazmi, H.; Walker, Z.; Booij, J.; Khan, F.; Shah, S.; Sudre, C.H.; Buckman, J.E.J.; Schrag, A.-E. Late Onset Depression: Dopaminergic Deficit and Clinical Features of Prodromal Parkinson’s Disease: A Cross-Sectional Study. J. Neurol. Neurosurg. Psychiatry 2021, 92, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Joling, M.; Vriend, C.; Raijmakers, P.G.H.M.; van der Zande, J.J.; Lemstra, A.W.; Berendse, H.W.; Booij, J.; van den Heuvel, O.A. Striatal DAT and Extrastriatal SERT Binding in Early-Stage Parkinson’s Disease and Dementia with Lewy Bodies, Compared with Healthy Controls: An 123I-FP-CIT SPECT Study. NeuroImage Clin. 2019, 22, 101755. [Google Scholar] [CrossRef]
- Kang, S.W.; Jeon, S.; Lee, Y.; Park, M.; Baik, K.; Jung, J.H.; Chung, S.J.; Yoo, H.S.; Jeong, S.H.; Yun, M.; et al. Implication of Metabolic and Dopamine Transporter PET in Dementia with Lewy Bodies. Sci. Rep. 2021, 11, 14394. [Google Scholar] [CrossRef]
- Sakakibara, S.; Hashimoto, R.; Katayama, T.; Kenjyo, M.; Yokokawa, Y.; Saito, Y.; Hirakawa, A.; Ito, M.; Nakamura, T.; Hara, K.; et al. Longitudinal Change of DAT SPECT in Parkinson’s Disease and Multiple System Atrophy. J. Park. Dis. 2020, 10, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Vergnet, S.; Hives, F.; Foubert-Samier, A.; Payoux, P.; Fernandez, P.; Meyer, M.; Dupouy, J.; Brefel-Courbon, C.; Ory-Magne, F.; Rascol, O.; et al. Dopamine Transporter Imaging for the Diagnosis of Multiple System Atrophy Cerebellar Type. Parkinsonism Relat. Disord. 2019, 63, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Kaasinen, V.; Vahlberg, T.; Stoessl, A.J.; Strafella, A.P.; Antonini, A. Dopamine Receptors in Parkinson’s Disease: A Meta-Analysis of Imaging Studies. Mov. Disord. 2021, 36, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Shigekiyo, T.; Arawaka, S. Laterality of Specific Binding Ratios on DAT-SPECT for Differential Diagnosis of Degenerative Parkinsonian Syndromes. Sci. Rep. 2020, 10, 15761. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishii, K.; Sousa, K.; Marumoto, K.; Kashibayashi, T.; Fujita, J.; Yokoyama, K. Distinctive Regional Asymmetry in Dopaminergic and Serotoninergic Dysfunction in Degenerative Parkinsonisms. J. Neurol. Sci. 2021, 423, 117363. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.S.; Lee, S.; Chung, S.J.; Lee, Y.H.; Lee, P.H.; Sohn, Y.H.; Lee, S.; Yun, M.; Ye, B.S. Dopaminergic Depletion, β-Amyloid Burden, and Cognition in Lewy Body Disease. Ann. Neurol. 2020, 87, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, J.R.; Brillman, S.; Chhabria, N.; Isaacson, S.H. Impact of DaTscan Imaging on Clinical Decision Making in Clinically Uncertain Parkinson’s Disease. J. Park. Dis. 2021, 11, 885–889. [Google Scholar] [CrossRef]
- Thobois, S.; Prange, S.; Scheiber, C.; Broussolle, E. What a Neurologist Should Know about PET and SPECT Functional Imaging for Parkinsonism: A Practical Perspective. Parkinsonism Relat. Disord. 2019, 59, 93–100. [Google Scholar] [CrossRef]
- Verger, A.; Grimaldi, S.; Ribeiro, M.-J.; Frismand, S.; Guedj, E. Single Photon Emission Computed Tomography/Positron Emission Tomography Molecular Imaging for Parkinsonism: A Fast-Developing Field. Ann. Neurol. 2021, 90, 711–719. [Google Scholar] [CrossRef]
- Schmitz-Steinkrüger, H.; Lange, C.; Apostolova, I.; Amthauer, H.; Lehnert, W.; Klutmann, S.; Buchert, R. Impact of the Size of the Normal Database on the Performance of the Specific Binding Ratio in Dopamine Transporter SPECT. EJNMMI Phys. 2020, 7, 34. [Google Scholar] [CrossRef]
- Schmitz-Steinkrüger, H.; Lange, C.; Apostolova, I.; Mathies, F.L.; Frings, L.; Klutmann, S.; Hellwig, S.; Meyer, P.T.; Buchert, R. Impact of Age and Sex Correction on the Diagnostic Performance of Dopamine Transporter SPECT. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 1445–1459. [Google Scholar] [CrossRef]
- Piccardo, A.; Cappuccio, R.; Bottoni, G.; Cecchin, D.; Mazzella, L.; Cirone, A.; Righi, S.; Ugolini, M.; Bianchi, P.; Bertolaccini, P.; et al. The Role of the Deep Convolutional Neural Network as an Aid to Interpreting Brain [18F]DOPA PET/CT in the Diagnosis of Parkinson’s Disease. Eur. Radiol. 2021, 31, 7003–7011. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, M.; Milletari, F.; Krüger, J.; Lange, C.; Schenk, M.; Apostolova, I.; Klutmann, S.; Ehrenburg, M.; Buchert, R. Automatic Classification of Dopamine Transporter SPECT: Deep Convolutional Neural Networks Can Be Trained to Be Robust with Respect to Variable Image Characteristics. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2800–2811. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, N.; Wegrzyk, J.; Preti, M.G.; Fleury, V.; Van de Ville, D.; Garibotto, V.; Burkhard, P.R. Classification of Degenerative Parkinsonism Subtypes by Support-Vector-Machine Analysis and Striatal 123I-FP-CIT Indices. J. Neurol. 2019, 266, 1771–1781. [Google Scholar] [CrossRef]
- Dotinga, M.; van Dijk, J.D.; Vendel, B.N.; Slump, C.H.; Portman, A.T.; van Dalen, J.A. Clinical Value of Machine Learning-Based Interpretation of I-123 FP-CIT Scans to Detect Parkinson’s Disease: A Two-Center Study. Ann. Nucl. Med. 2021, 35, 378–385. [Google Scholar] [CrossRef]
- Nazari, M.; Kluge, A.; Apostolova, I.; Klutmann, S.; Kimiaei, S.; Schroeder, M.; Buchert, R. Data-Driven Identification of Diagnostically Useful Extrastriatal Signal in Dopamine Transporter SPECT Using Explainable AI. Sci. Rep. 2021, 11, 22932. [Google Scholar] [CrossRef]
- Shiiba, T.; Arimura, Y.; Nagano, M.; Takahashi, T.; Takaki, A. Improvement of Classification Performance of Parkinson’s Disease Using Shape Features for Machine Learning on Dopamine Transporter Single Photon Emission Computed Tomography. PLoS ONE 2020, 15, e0228289. [Google Scholar] [CrossRef]
- Magesh, P.R.; Myloth, R.D.; Tom, R.J. An Explainable Machine Learning Model for Early Detection of Parkinson’s Disease Using LIME on DaTSCAN Imagery. Comput. Biol. Med. 2020, 126, 104041. [Google Scholar] [CrossRef]
- Manzanera, O.M.; Meles, S.K.; Leenders, K.L.; Renken, R.J.; Pagani, M.; Arnaldi, D.; Nobili, F.; Obeso, J.; Oroz, M.R.; Morbelli, S.; et al. Scaled Subprofile Modeling and Convolutional Neural Networks for the Identification of Parkinson’s Disease in 3D Nuclear Imaging Data. Int. J. Neural Syst. 2019, 29, 1950010. [Google Scholar] [CrossRef]
- Shen, T.; Jiang, J.; Lin, W.; Ge, J.; Wu, P.; Zhou, Y.; Zuo, C.; Wang, J.; Yan, Z.; Shi, K. Use of Overlapping Group LASSO Sparse Deep Belief Network to Discriminate Parkinson’s Disease and Normal Control. Front. Neurosci. 2019, 13, 396. [Google Scholar] [CrossRef] [Green Version]
- Arnaldi, D.; Mattioli, P.; Famà, F.; Girtler, N.; Brugnolo, A.; Pardini, M.; Donniaquio, A.; Massa, F.; Orso, B.; Raffa, S.; et al. Stratification Tools for Disease-Modifying Trials in Prodromal Synucleinopathy. Mov. Disord. 2022, 37, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, D.; Hill, D.; Cedarbaum, J.M.; Tome, M.; Vamvakas, S.; Romero, K.; Conrado, D.J.; Dexter, D.T.; Seibyl, J.; Jennings, D.; et al. The Qualification of an Enrichment Biomarker for Clinical Trials Targeting Early Stages of Parkinson’s Disease. J. Park. Dis. 2019, 9, 553–563. [Google Scholar] [CrossRef]
- Massa, J.; Chahine, L.M. Revision of Diagnosis in Early Parkinsonism with Abnormal Dopamine Transporter Imaging. J. Park. Dis. 2019, 9, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Arnaldi, D.; Famà, F.; Girtler, N.; Brugnolo, A.; Pardini, M.; Mattioli, P.; Meli, R.; Massa, F.; Orso, B.; Sormani, M.P.; et al. Rapid Eye Movement Sleep Behavior Disorder: A Proof-of-Concept Neuroprotection Study for Prodromal Synucleinopathies. Eur. J. Neurol. 2021, 28, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Horsager, J.; Andersen, K.B.; Knudsen, K.; Skjærbæk, C.; Fedorova, T.D.; Okkels, N.; Schaeffer, E.; Bonkat, S.K.; Geday, J.; Otto, M.; et al. Brain-First versus Body-First Parkinson’s Disease: A Multimodal Imaging Case-Control Study. Brain 2020, 143, 3077–3088. [Google Scholar] [CrossRef]
- Knudsen, K.; Fedorova, T.D.; Hansen, A.K.; Sommerauer, M.; Otto, M.; Svendsen, K.B.; Nahimi, A.; Stokholm, M.G.; Pavese, N.; Beier, C.P.; et al. In-Vivo Staging of Pathology in REM Sleep Behaviour Disorder: A Multimodality Imaging Case-Control Study. Lancet Neurol. 2018, 17, 618–628. [Google Scholar] [CrossRef]
- Berg, D.; Borghammer, P.; Fereshtehnejad, S.-M.; Heinzel, S.; Horsager, J.; Schaeffer, E.; Postuma, R.B. Prodromal Parkinson Disease Subtypes—Key to Understanding Heterogeneity. Nat. Rev. Neurol. 2021, 17, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.; Fedorova, T.D.; Horsager, J.; Andersen, K.B.; Skjærbæk, C.; Berg, D.; Schaeffer, E.; Brooks, D.J.; Pavese, N.; Van Den Berge, N.; et al. Asymmetric Dopaminergic Dysfunction in Brain-First versus Body-First Parkinson’s Disease Subtypes. J. Park. Dis. 2021, 11, 1677–1687. [Google Scholar] [CrossRef]
- Valli, M.; Cho, S.S.; Uribe, C.; Masellis, M.; Chen, R.; Mihaescu, A.; Strafella, A.P. VMAT2 Availability in Parkinson’s Disease with Probable REM Sleep Behaviour Disorder. Mol. Brain 2021, 14, 165. [Google Scholar] [CrossRef]
- Valli, M.; Cho, S.S.; Masellis, M.; Chen, R.; Koshimori, Y.; Diez-Cirarda, M.; Mihaescu, A.; Christopher, L.; Strafella, A.P. Extra-Striatal Dopamine in Parkinson’s Disease with Rapid Eye Movement Sleep Behavior Disorder. J. Neurosci. Res. 2021, 99, 1177–1187. [Google Scholar] [CrossRef]
- Pasquini, J.; Ceravolo, R.; Brooks, D.J.; Bonuccelli, U.; Pavese, N. Progressive Loss of Raphe Nuclei Serotonin Transporter in Early Parkinson’s Disease: A Longitudinal 123I-FP-CIT SPECT Study. Parkinsonism Relat. Disord. 2020, 77, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Koros, C.; Yousaf, T.; Picillo, M.; Polychronis, S.; Simitsi, A.; Giordano, B.; Chappell, Z.; et al. Serotonergic Pathology and Disease Burden in the Premotor and Motor Phase of A53T α-Synuclein Parkinsonism: A Cross-Sectional Study. Lancet Neurol. 2019, 18, 748–759. [Google Scholar] [CrossRef]
- Sampedro, F.; Marín-Lahoz, J.; Martínez-Horta, S.; Camacho, V.; Lopez-Mora, D.-A.; Pagonabarraga, J.; Kulisevsky, J. Extrastriatal SPECT-DAT Uptake Correlates with Clinical and Biological Features of de Novo Parkinson’s Disease. Neurobiol. Aging 2021, 97, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.F.; Matarazzo, M.; McKenzie, J.; Neilson, N.; Vafai, N.; Dinelle, K.; Felicio, A.C.; McKeown, M.J.; Stoessl, A.J.; Sossi, V. Serotonergic System Impacts Levodopa Response in Early Parkinson’s and Future Risk of Dyskinesia. Mov. Disord. 2021, 36, 389–397. [Google Scholar] [CrossRef]
- Maillet, A.; Krack, P.; Lhommée, E.; Météreau, E.; Klinger, H.; Favre, E.; Bars, D.L.; Schmitt, E.; Bichon, A.; Pelissier, P.; et al. The Prominent Role of Serotonergic Degeneration in Apathy, Anxiety and Depression in de Novo Parkinson’s Disease. Brain 2016, 139, 2486–2502. [Google Scholar] [CrossRef]
- Maillet, A.; Météreau, E.; Tremblay, L.; Favre, E.; Klinger, H.; Lhommée, E.; Le Bars, D.; Castrioto, A.; Prange, S.; Sgambato, V.; et al. Serotonergic and Dopaminergic Lesions Underlying Parkinsonian Neuropsychiatric Signs. Mov Disord 2021, 36, 2888–2900. [Google Scholar] [CrossRef]
- Prange, S.; Metereau, E.; Maillet, A.; Klinger, H.; Schmitt, E.; Lhommée, E.; Bichon, A.; Lancelot, S.; Meoni, S.; Broussolle, E.; et al. Limbic Serotonergic Plasticity Contributes to the Compensation of Apathy in Early Parkinson’s Disease. Mov. Disord. 2022, 37, 1211–1221. [Google Scholar] [CrossRef]
- Jørgensen, L.M.; Henriksen, T.; Mardosiene, S.; Keller, S.H.; Stenbæk, D.S.; Hansen, H.D.; Jespersen, B.; Thomsen, C.; Weikop, P.; Svarer, C.; et al. Parkinson Patients Have a Presynaptic Serotonergic Deficit: A Dynamic Deep Brain Stimulation PET Study. J. Cereb. Blood Flow Metab. 2021, 41, 1954–1963. [Google Scholar] [CrossRef]
- Pilotto, A.; Schiano di Cola, F.; Premi, E.; Grasso, R.; Turrone, R.; Gipponi, S.; Scalvini, A.; Cottini, E.; Paghera, B.; Garibotto, V.; et al. Extrastriatal Dopaminergic and Serotonergic Pathways in Parkinson’s Disease and in Dementia with Lewy Bodies: A 123I-FP-CIT SPECT Study. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 1642–1651. [Google Scholar] [CrossRef]
- van der Zande, J.J.; Joling, M.; Happach, I.G.; Vriend, C.; Scheltens, P.; Booij, J.; Lemstra, A.W. Serotonergic Deficits in Dementia with Lewy Bodies with Concomitant Alzheimer’s Disease Pathology: An 123I-FP-CIT SPECT Study. NeuroImage Clin. 2020, 25, 102062. [Google Scholar] [CrossRef]
- Joling, M.; Vriend, C.; van den Heuvel, O.A.; Raijmakers, P.G.H.M.; Jones, P.A.; Berendse, H.W.; Booij, J. Analysis of Extrastriatal 123I-FP-CIT Binding Contributes to the Differential Diagnosis of Parkinsonian Diseases. J. Nucl. Med. 2017, 58, 1117–1123. [Google Scholar] [CrossRef]
- Meyer, M.; Lamare, F.; Asselineau, J.; Foubert-Samier, A.; Mazère, J.; Zanotti-Fregonara, P.; Rizzo, G.; Delamarre, A.; Spampinato, U.; Rascol, O.; et al. Brain 5-HT1A Receptor Binding in Multiple System Atrophy: An [18F]-MPPF PET Study. Mov. Disord. 2021, 36, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kudo, T.; Lapa, C.; Buck, A.; Higuchi, T. Recent Advances in Radiotracers Targeting Norepinephrine Transporter: Structural Development and Radiolabeling Improvements. J. Neural Transm. Vienna Austria 1996 2020, 127, 851–873. [Google Scholar] [CrossRef]
- Laurencin, C.; Lancelot, S.; Gobert, F.; Redouté, J.; Mérida, I.; Iecker, T.; Liger, F.; Irace, Z.; Greusard, E.; Lamberet, L.; et al. Modeling [11C]Yohimbine PET Human Brain Kinetics with Test-Retest Reliability, Competition Sensitivity Studies and Search for a Suitable Reference Region. NeuroImage 2021, 240, 118328. [Google Scholar] [CrossRef]
- Doppler, C.E.J.; Kinnerup, M.B.; Brune, C.; Farrher, E.; Betts, M.; Fedorova, T.D.; Schaldemose, J.L.; Knudsen, K.; Ismail, R.; Seger, A.D.; et al. Regional Locus Coeruleus Degeneration Is Uncoupled from Noradrenergic Terminal Loss in Parkinson’s Disease. Brain 2021, 144, 2732–2744. [Google Scholar] [CrossRef]
- Doppler, C.E.J.; Smit, J.A.M.; Hommelsen, M.; Seger, A.; Horsager, J.; Kinnerup, M.B.; Hansen, A.K.; Fedorova, T.D.; Knudsen, K.; Otto, M.; et al. Microsleep Disturbances Are Associated with Noradrenergic Dysfunction in Parkinson’s Disease. Sleep 2021, 44, zsab040. [Google Scholar] [CrossRef]
- Kinnerup, M.B.; Sommerauer, M.; Damholdt, M.F.; Schaldemose, J.L.; Ismail, R.; Terkelsen, A.J.; Stær, K.; Hansen, A.; Fedorova, T.D.; Knudsen, K.; et al. Preserved Noradrenergic Function in Parkinson’s Disease Patients with Rest Tremor. Neurobiol. Dis. 2021, 152, 105295. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.B.; Hansen, A.K.; Sommerauer, M.; Fedorova, T.D.; Knudsen, K.; Vang, K.; Van Den Berge, N.; Kinnerup, M.; Nahimi, A.; Pavese, N.; et al. Altered Sensorimotor Cortex Noradrenergic Function in Idiopathic REM Sleep Behaviour Disorder—A PET Study. Parkinsonism Relat. Disord. 2020, 75, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Lenka, A.; Lamotte, G.; Goldstein, D.S. Cardiac 18F-Dopamine PET Distinguishes PD with Orthostatic Hypotension from Parkinsonian MSA. Mov. Disord. Clin. Pract. 2021, 8, 582–586. [Google Scholar] [CrossRef]
- Kuten, J.; Linevitz, A.; Lerman, H.; Freedman, N.; Kestenbaum, M.; Shiner, T.; Giladi, N.; Even-Sapir, E. [18F] FDOPA PET May Confirm the Clinical Diagnosis of Parkinson’s Disease by Imaging the Nigro-Striatal Pathway and the Sympathetic Cardiac Innervation: Proof-of-Concept Study. J. Integr. Neurosci. 2020, 19, 489–494. [Google Scholar] [CrossRef]
- van der Zee, S.; Vállez García, D.; Elsinga, P.H.; Willemsen, A.T.M.; Boersma, H.H.; Gerritsen, M.J.J.; Spikman, J.M.; van Laar, T. [18F]Fluoroethoxybenzovesamicol in Parkinson’s Disease Patients: Quantification of a Novel Cholinergic Positron Emission Tomography Tracer. Mov. Disord. 2019, 34, 924–926. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Kanel, P.; Zhou, Z.; Koeppe, R.A.; Frey, K.A.; Dauer, W.T.; Albin, R.L.; Müller, M.L.T.M. Cholinergic System Changes of Falls and Freezing of Gait in Parkinson’s Disease. Ann. Neurol. 2019, 85, 538–549. [Google Scholar] [CrossRef]
- van der Zee, S.; Müller, M.L.T.M.; Kanel, P.; van Laar, T.; Bohnen, N.I. Cholinergic Denervation Patterns Across Cognitive Domains in Parkinson’s Disease. Mov. Disord. 2021, 36, 642–650. [Google Scholar] [CrossRef]
- Pasquini, J.; Brooks, D.J.; Pavese, N. The Cholinergic Brain in Parkinson’s Disease. Mov. Disord. Clin. Pract. 2021, 8, 1012–1026. [Google Scholar] [CrossRef]
- Bohnen, N.I.; Kanel, P.; Müller, M.L.T.M. Molecular Imaging of the Cholinergic System in Parkinson’s Disease. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 141, pp. 211–250. ISBN 978-0-12-815418-2. [Google Scholar]
- van der Zee, S.; Kanel, P.; Gerritsen, M.J.J.; Boertien, J.M.; Slomp, A.C.; Müller, M.L.T.M.; Bohnen, N.I.; Spikman, J.M.; van Laar, T. Altered Cholinergic Innervation in De Novo Parkinson’s Disease with and Without Cognitive Impairment. Mov. Disord. 2022, 37, 713–723. [Google Scholar] [CrossRef]
- Bedard, M.-A.; Aghourian, M.; Legault-Denis, C.; Postuma, R.B.; Soucy, J.-P.; Gagnon, J.-F.; Pelletier, A.; Montplaisir, J. Brain Cholinergic Alterations in Idiopathic REM Sleep Behaviour Disorder: A PET Imaging Study with 18F-FEOBV. Sleep Med. 2019, 58, 35–41. [Google Scholar] [CrossRef]
- Gersel Stokholm, M.; Iranzo, A.; Østergaard, K.; Serradell, M.; Otto, M.; Bacher Svendsen, K.; Garrido, A.; Vilas, D.; Fedorova, T.D.; Santamaria, J.; et al. Cholinergic Denervation in Patients with Idiopathic Rapid Eye Movement Sleep Behaviour Disorder. Eur. J. Neurol. 2020, 27, 644–652. [Google Scholar] [CrossRef]
- Kanel, P.; Müller, M.L.T.M.; van der Zee, S.; Sanchez-Catasus, C.A.; Koeppe, R.A.; Frey, K.A.; Bohnen, N.I. Topography of Cholinergic Changes in Dementia With Lewy Bodies and Key Neural Network Hubs. J. Neuropsychiatry Clin. Neurosci. 2020, 32, 370–375. [Google Scholar] [CrossRef]
- Ji, L.; Fang, Y.; Tang, J.; Liu, C.; Huang, C.; Hu, Q.; Li, Q.; Chen, Z. Synthesis and Biological Evaluation of 18F-Labelled Dopamine D3 Receptor Selective Ligands. Bioorg. Med. Chem. Lett. 2022, 62, 128630. [Google Scholar] [CrossRef] [PubMed]
- Lehnert, W.; Riss, P.J.; Hurtado de Mendoza, A.; Lopez, S.; Fernandez, G.; Ilheu, M.; Amaral, H.; Kramer, V. Whole-Body Biodistribution and Radiation Dosimetry of [18F]PR04.MZ: A New PET Radiotracer for Clinical Management of Patients with Movement Disorders. EJNMMI Res. 2022, 12, 1. [Google Scholar] [CrossRef]
- Pain, C.D.; O’Keefe, G.J.; Ackermann, U.; Dore, V.; Villemagne, V.L.; Rowe, C.C. Human Biodistribution and Internal Dosimetry of 4-[ 18F]Fluorobenzyl-Dexetimide: A PET Radiopharmaceutical for Imaging Muscarinic Acetylcholine Receptors in the Brain and Heart. EJNMMI Res. 2020, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Rowe, C.C.; Krishnadas, N.; Ackermann, U.; Doré, V.; Goh, R.Y.W.; Guzman, R.; Chong, L.; Bozinovski, S.; Mulligan, R.; Kanaan, R.; et al. PET Imaging of Brain Muscarinic Receptors with 18F-Fluorobenzyl-Dexetimide: A First in Human Study. Psychiatry Res. Neuroimaging 2021, 316, 111354. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.H.; Toussaint, M.; Teodoro, R.; Dukić-Stefanović, S.; Kranz, M.; Deuther-Conrad, W.; Moldovan, R.-P.; Brust, P. Synthesis and Biological Evaluation of a Novel 18F-Labeled Radiotracer for PET Imaging of the Adenosine A2A Receptor. Int. J. Mol. Sci. 2021, 22, 1182. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Q.; Zhang, Y.; Fang, J.; Jiang, D.; Mou, Z.; Liu, H.; Su, R.; Wang, C.; He, F.; et al. 18F-Labelled Pyrrolopyrimidines Reveal Brain Leucine-Rich Repeat Kinase 2 Expression Implicated in Parkinson’s Disease. Eur. J. Med. Chem. 2021, 214, 113245. [Google Scholar] [CrossRef] [PubMed]
- Malik, N.; Kornelsen, R.; McCormick, S.; Colpo, N.; Merkens, H.; Bendre, S.; Benard, F.; Sossi, V.; Schirrmacher, R.; Schaffer, P. Development and Biological Evaluation of[18F]FMN3PA & [18F]FMN3PU for Leucine-Rich Repeat Kinase 2 (LRRK2) in Vivo PET Imaging. Eur. J. Med. Chem. 2021, 211, 113005. [Google Scholar] [CrossRef]
- Thobois, S.; Brefel-Courbon, C.; Le Bars, D.; Sgambato-Faure, V. Molecular Imaging of Opioid System in Idiopathic Parkinson’s Disease. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 141, pp. 275–303. ISBN 978-0-12-815418-2. [Google Scholar]
- Green, D.G.J.; Kim, J.; Kish, S.J.; Tyndale, R.F.; Hill, M.N.; Strafella, A.P.; Tong, J.; McCluskey, T.; Westwood, D.J.; Houle, S.; et al. Fatty Acid Amide Hydrolase Binding Is Inversely Correlated with Amygdalar Functional Connectivity: A Combined Positron Emission Tomography and Magnetic Resonance Imaging Study in Healthy Individuals. J. Psychiatry Neurosci. 2021, 46, E238–E246. [Google Scholar] [CrossRef] [PubMed]
- Gjerum, L.; Frederiksen, K.S.; Henriksen, O.M.; Law, I.; Anderberg, L.; Andersen, B.B.; Bjerregaard, E.; Hejl, A.-M.; Høgh, P.; Hasselbalch, S.G. A Visual Rating Scale for Cingulate Island Sign on 18F-FDG-PET to Differentiate Dementia with Lewy Bodies and Alzheimer’s Disease. J. Neurol. Sci. 2020, 410, 116645. [Google Scholar] [CrossRef]
- Iizuka, T.; Fukasawa, M.; Kameyama, M. Deep-Learning-Based Imaging-Classification Identified Cingulate Island Sign in Dementia with Lewy Bodies. Sci. Rep. 2019, 9, 8944. [Google Scholar] [CrossRef] [PubMed]
- Morbelli, S.; Chincarini, A.; Brendel, M.; Rominger, A.; Bruffaerts, R.; Vandenberghe, R.; Kramberger, M.G.; Trost, M.; Garibotto, V.; Nicastro, N.; et al. Metabolic Patterns across Core Features in Dementia with Lewy Bodies. Ann. Neurol. 2019, 85, 715–725. [Google Scholar] [CrossRef]
- Park, K.W.; Ko, J.H.; Choi, N.; Jo, S.; Park, Y.J.; Lee, E.-J.; Kim, S.J.; Chung, S.J.; Lee, C.S. Cortical Hypometabolism Associated with Cognitive Impairment of Multiple System Atrophy. Parkinsonism Relat. Disord. 2020, 81, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Chen, Q.-S.; Zuo, C.-T.; Liu, F.-T.; Wang, J. The Frontal and Cerebellar Metabolism Related to Cognitive Dysfunction in Multiple System Atrophy. Front. Aging Neurosci. 2022, 14, 788166. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, S.; Boucekine, M.; Witjas, T.; Fluchere, F.; Azulay, J.-P.; Guedj, E.; Eusebio, A. Early Atypical Signs and Insula Hypometabolism Predict Survival in Multiple System Atrophy. J. Neurol. Neurosurg. Psychiatry 2021, 92, 881–889. [Google Scholar] [CrossRef]
- Grimaldi, S.; Boucekine, M.; Witjas, T.; Fluchère, F.; Renaud, M.; Azulay, J.-P.; Guedj, E.; Eusebio, A. Multiple System Atrophy: Phenotypic Spectrum Approach Coupled with Brain 18-FDG PET. Parkinsonism Relat. Disord. 2019, 67, 3–9. [Google Scholar] [CrossRef]
- Orso, B.; Arnaldi, D.; Girtler, N.; Brugnolo, A.; Doglione, E.; Mattioli, P.; Biassoni, E.; Fancellu, R.; Massa, F.; Bauckneht, M.; et al. Dopaminergic and Serotonergic Degeneration and Cortical [18F]Fluorodeoxyglucose Positron Emission Tomography in De Novo Parkinson’s Disease. Mov. Disord. 2021, 36, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, M.C.; Greuel, A.; Tahmasian, M.; Schwartz, F.; Stürmer, S.; Maier, F.; Hammes, J.; Tittgemeyer, M.; Timmermann, L.; van Eimeren, T.; et al. Network Degeneration in Parkinson’s Disease: Multimodal Imaging of Nigro-Striato-Cortical Dysfunction. Brain 2020, 143, 944–959. [Google Scholar] [CrossRef]
- Ruppert, M.C.; Greuel, A.; Freigang, J.; Tahmasian, M.; Maier, F.; Hammes, J.; van Eimeren, T.; Timmermann, L.; Tittgemeyer, M.; Drzezga, A.; et al. The Default Mode Network and Cognition in Parkinson’s Disease: A Multimodal Resting-State Network Approach. Hum. Brain Mapp. 2021, 42, 2623–2641. [Google Scholar] [CrossRef]
- Huber, M.; Beyer, L.; Prix, C.; Schönecker, S.; Palleis, C.; Rauchmann, B.-S.; Morbelli, S.; Chincarini, A.; Bruffaerts, R.; Vandenberghe, R.; et al. Metabolic Correlates of Dopaminergic Loss in Dementia with Lewy Bodies. Mov. Disord. 2020, 35, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Emsen, B.; Villafane, G.; David, J.-P.; Evangelista, E.; Chalaye, J.; Lerman, L.; Authier, F.-J.; Gracies, J.-M.; Itti, E. Clinical Impact of Dual-Tracer FDOPA and FDG PET/CT for the Evaluation of Patients with Parkinsonian Syndromes. Medicine 2020, 99, e23060. [Google Scholar] [CrossRef] [PubMed]
- Spetsieris, P.G.; Ko, J.H.; Tang, C.C.; Nazem, A.; Sako, W.; Peng, S.; Ma, Y.; Dhawan, V.; Eidelberg, D. Metabolic Resting-State Brain Networks in Health and Disease. Proc. Natl. Acad. Sci. USA 2015, 112, 2563–2568. [Google Scholar] [CrossRef] [PubMed]
- Schindlbeck, K.A.; Eidelberg, D. Network Imaging Biomarkers: Insights and Clinical Applications in Parkinson’s Disease. Lancet Neurol. 2018, 17, 629–640. [Google Scholar] [CrossRef]
- Spetsieris, P.G.; Eidelberg, D. Spectral Guided Sparse Inverse Covariance Estimation of Metabolic Networks in Parkinson’s Disease. NeuroImage 2021, 226, 117568. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.C.; Holtbernd, F.; Ma, Y.; Spetsieris, P.; Oh, A.; Fink, G.R.; Timmermann, L.; Eggers, C.; Eidelberg, D. Hemispheric Network Expression in Parkinson’s Disease: Relationship to Dopaminergic Asymmetries. J. Park. Dis. 2020, 10, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Schindlbeck, K.A.; Vo, A.; Mattis, P.J.; Villringer, K.; Marzinzik, F.; Fiebach, J.B.; Eidelberg, D. Cognition-Related Functional Topographies in Parkinson’s Disease: Localized Loss of the Ventral Default Mode Network. Cereb. Cortex 2021, 31, 5139–5150. [Google Scholar] [CrossRef] [PubMed]
- Martí-Andrés, G.; van Bommel, L.; Meles, S.K.; Riverol, M.; Valentí, R.; Kogan, R.V.; Renken, R.J.; Gurvits, V.; van Laar, T.; Pagani, M.; et al. Multicenter Validation of Metabolic Abnormalities Related to PSP According to the MDS-PSP Criteria. Mov. Disord. 2020, 35, 2009–2018. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, C.; Ge, J.; Lu, J.; Li, L.; Yu, H.; Wu, J.; Wang, J.; Wu, P.; Zuo, C. The Impact of Probable Rapid Eye Movement Sleep Behavior Disorder on Parkinson’s Disease: A Dual-Tracer PET Imaging Study. Parkinsonism Relat. Disord. 2022, 95, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Lee, J.-Y.; Nam, H.; Kim, H.-J.; Jeon, B.; Jeong, J.M.; Kim, Y.K. A New Metabolic Network Correlated with Olfactory and Executive Dysfunctions in Idiopathic Rapid Eye Movement Sleep Behavior Disorder. J. Clin. Neurol. 2019, 15, 175–183. [Google Scholar] [CrossRef]
- Kim, R.; Lee, J.-Y.; Kim, Y.K.; Kim, H.; Yoon, E.J.; Shin, J.H.; Yoo, D.; Nam, H.; Jeon, B. Longitudinal Changes in Isolated Rapid Eye Movement Sleep Behavior Disorder-Related Metabolic Pattern Expression. Mov. Disord. 2021, 36, 1889–1898. [Google Scholar] [CrossRef]
- Chun, K.; Kong, E.; Cho, I. Comparison of Perfusion 18F-FP-CIT PET and 99mTc-ECD SPECT in Parkinsonian Disorders. Medicine (Baltimore) 2021, 100, e27019. [Google Scholar] [CrossRef]
- Peng, S.; Tang, C.; Schindlbeck, K.; Rydzinski, Y.; Dhawan, V.; Spetsieris, P.G.; Ma, Y.; Eidelberg, D. Dynamic 18 F-FPCIT PET: Quantification of Parkinson Disease Metabolic Networks and Nigrostriatal Dopaminergic Dysfunction in a Single Imaging Session. J. Nucl. Med. 2021, 62, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Carson, R.E.; Naganawa, M.; Toyonaga, T.; Koohsari, S.; Yang, Y.; Chen, M.-K.; Matuskey, D.; Finnema, S.J. Imaging of Synaptic Density in Neurodegenerative Disorders. J. Nucl. Med. 2022, 63, 60S–67S. [Google Scholar] [CrossRef] [PubMed]
- Matuskey, D.; Tinaz, S.; Wilcox, K.C.; Naganawa, M.; Toyonaga, T.; Dias, M.; Henry, S.; Pittman, B.; Ropchan, J.; Nabulsi, N.; et al. Synaptic Changes in Parkinson Disease Assessed with in Vivo Imaging. Ann. Neurol. 2020, 87, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Delva, A.; Van Weehaeghe, D.; Koole, M.; Van Laere, K.; Vandenberghe, W. Loss of Presynaptic Terminal Integrity in the Substantia Nigra in Early Parkinson’s Disease. Mov. Disord. 2020, 35, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Pagano, G.; de Natale, E.R.; Mansur, A.; Caminiti, S.P.; Polychronis, S.; Middleton, L.T.; Price, G.; Schmidt, K.F.; Gunn, R.N.; et al. Mitochondrial Complex 1, Sigma 1, and Synaptic Vesicle 2A in Early Drug-Naive Parkinson’s Disease. Mov. Disord. 2020, 35, 1416–1427. [Google Scholar] [CrossRef]
- Andersen, K.B.; Hansen, A.K.; Damholdt, M.F.; Horsager, J.; Skjærbæk, C.; Gottrup, H.; Klit, H.; Schacht, A.C.; Danielsen, E.H.; Brooks, D.J.; et al. Reduced Synaptic Density in Patients with Lewy Body Dementia: An [11C]UCB-J PET Imaging Study. Mov. Disord. 2021, 36, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.; Jones, P.S.; Savulich, G.; Wiggins, J.K.; Hong, Y.T.; Fryer, T.D.; Manavaki, R.; Sephton, S.M.; Boros, I.; Malpetti, M.; et al. Synaptic Loss in Primary Tauopathies Revealed by [11C]UCB-J Positron Emission Tomography. Mov. Disord. 2020, 35, 1834–1842. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-F.; Gao, F. Neuroinflammation in Parkinson’s Disease: A Meta-Analysis of PET Imaging Studies. J. Neurol. 2021. [Google Scholar] [CrossRef]
- Gersel Stokholm, M.; Garrido, A.; Tolosa, E.; Serradell, M.; Iranzo, A.; Østergaard, K.; Borghammer, P.; Møller, A.; Parbo, P.; Stær, K.; et al. Imaging Dopamine Function and Microglia in Asymptomatic LRRK2 Mutation Carriers. J. Neurol. 2020, 267, 2296–2300. [Google Scholar] [CrossRef]
- Mullin, S.; Stokholm, M.G.; Hughes, D.; Mehta, A.; Parbo, P.; Hinz, R.; Pavese, N.; Brooks, D.J.; Schapira, A.H.V. Brain Microglial Activation Increased in Glucocerebrosidase (GBA) Mutation Carriers without Parkinson’s Disease. Mov. Disord. 2021, 36, 774–779. [Google Scholar] [CrossRef]
- Malpetti, M.; Passamonti, L.; Jones, P.S.; Street, D.; Rittman, T.; Fryer, T.D.; Hong, Y.T.; Rodriguez, P.V.; Bevan-Jones, W.R.; Aigbirhio, F.I.; et al. Neuroinflammation Predicts Disease Progression in Progressive Supranuclear Palsy. J. Neurol. Neurosurg. Psychiatry 2021, 92, 769–775. [Google Scholar] [CrossRef]
- Best, L.; Ghadery, C.; Pavese, N.; Tai, Y.F.; Strafella, A.P. New and Old TSPO PET Radioligands for Imaging Brain Microglial Activation in Neurodegenerative Disease. Curr. Neurol. Neurosci. Rep. 2019, 19, 24. [Google Scholar] [CrossRef]
- Wimberley, C.; Lavisse, S.; Hillmer, A.; Hinz, R.; Turkheimer, F.; Zanotti-Fregonara, P. Kinetic Modeling and Parameter Estimation of TSPO PET Imaging in the Human Brain. Eur. J. Nucl. Med. Mol. Imaging 2021, 49, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.R.; Yeo, A.J.; Gunn, R.N.; Song, K.; Wadsworth, G.; Lewis, A.; Rhodes, C.; Pulford, D.J.; Bennacef, I.; Parker, C.A.; et al. An 18-KDa Translocator Protein (TSPO) Polymorphism Explains Differences in Binding Affinity of the PET Radioligand PBR28. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2012, 32, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Varnäs, K.; Cselényi, Z.; Jucaite, A.; Halldin, C.; Svenningsson, P.; Farde, L.; Varrone, A. PET Imaging of [11C]PBR28 in Parkinson’s Disease Patients Does Not Indicate Increased Binding to TSPO despite Reduced Dopamine Transporter Binding. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Ghadery, C.; Koshimori, Y.; Coakeley, S.; Harris, M.; Rusjan, P.; Kim, J.; Houle, S.; Strafella, A.P. Microglial Activation in Parkinson’s Disease Using [18F]-FEPPA. J. Neuroinflamm. 2017, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Koshimori, Y.; Ko, J.-H.; Mizrahi, R.; Rusjan, P.; Mabrouk, R.; Jacobs, M.F.; Christopher, L.; Hamani, C.; Lang, A.E.; Wilson, A.A.; et al. Imaging Striatal Microglial Activation in Patients with Parkinson’s Disease. PLoS ONE 2015, 10, e0138721. [Google Scholar] [CrossRef] [PubMed]
- Lavisse, S.; Goutal, S.; Wimberley, C.; Tonietto, M.; Bottlaender, M.; Gervais, P.; Kuhnast, B.; Peyronneau, M.-A.; Barret, O.; Lagarde, J.; et al. Increased Microglial Activation in Patients with Parkinson Disease Using [18F]-DPA714 TSPO PET Imaging. Parkinsonism Relat. Disord. 2021, 82, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Jucaite, A.; Cselényi, Z.; Kreisl, W.C.; Rabiner, E.A.; Varrone, A.; Carson, R.E.; Rinne, J.O.; Savage, A.; Schou, M.; Johnström, P.; et al. Glia Imaging Differentiates Multiple System Atrophy from Parkinson’s Disease: A Positron Emission Tomography Study with [11C]PBR28 and Machine Learning Analysis. Mov. Disord. 2022, 37, 119–129. [Google Scholar] [CrossRef]
- Laurell, G.L.; Plavén-Sigray, P.; Jucaite, A.; Varrone, A.; Cosgrove, K.P.; Svarer, C.; Knudsen, G.M.; Consortium, K.S.P.; Ogden, R.T.; Zanderigo, F.; et al. Nondisplaceable Binding Is a Potential Confounding Factor in 11C-PBR28 Translocator Protein PET Studies. J. Nucl. Med. 2021, 62, 412–417. [Google Scholar] [CrossRef]
- Lee, J.-H.; Simeon, F.G.; Liow, J.-S.; Morse, C.L.; Gladding, R.L.; Montero Santamaria, J.A.; Henter, I.D.; Zoghbi, S.S.; Pike, V.W.; Innis, R.B. In Vivo Evaluation of Six Analogs of 11C-ER176 as Candidate 18F-Labeled Radioligands for Translocator Protein 18 KDa (TSPO). J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2022, 63, 1252–1258. [Google Scholar] [CrossRef]
- Mahler, C.; Schumacher, A.-M.; Unterrainer, M.; Kaiser, L.; Höllbacher, T.; Lindner, S.; Havla, J.; Ertl-Wagner, B.; Patzig, M.; Seelos, K.; et al. TSPO PET Imaging of Natalizumab-Associated Progressive Multifocal Leukoencephalopathy. Brain 2021, 144, 2683–2695. [Google Scholar] [CrossRef]
- Palleis, C.; Sauerbeck, J.; Beyer, L.; Harris, S.; Schmitt, J.; Morenas-Rodriguez, E.; Finze, A.; Nitschmann, A.; Ruch-Rubinstein, F.; Eckenweber, F.; et al. In Vivo Assessment of Neuroinflammation in 4-Repeat Tauopathies. Mov. Disord. 2021, 36, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Dervenoulas, G.; Pagano, G.; Tyacke, R.J.; Polychronis, S.; Myers, J.; Gunn, R.N.; Rabiner, E.A.; Nutt, D.; Politis, M. Imidazoline 2 Binding Sites Reflecting Astroglia Pathology in Parkinson’s Disease: An in Vivo11C-BU99008 PET Study. Brain 2019, 142, 3116–3128. [Google Scholar] [CrossRef] [PubMed]
- Ezura, M.; Kikuchi, A.; Ishiki, A.; Okamura, N.; Hasegawa, T.; Harada, R.; Watanuki, S.; Funaki, Y.; Hiraoka, K.; Baba, T.; et al. Longitudinal Changes in 18F-THK5351 Positron Emission Tomography in Corticobasal Syndrome. Eur. J. Neurol. 2019, 26, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Harada, R.; Furumoto, S.; Kudo, Y.; Yanai, K.; Villemagne, V.L.; Okamura, N. Imaging of Reactive Astrogliosis by Positron Emission Tomography. Front. Neurosci. 2022, 16, 807435. [Google Scholar] [CrossRef]
- Van Weehaeghe, D.; Koole, M.; Schmidt, M.E.; Deman, S.; Jacobs, A.H.; Souche, E.; Serdons, K.; Sunaert, S.; Bormans, G.; Vandenberghe, W.; et al. [11C]JNJ54173717, a Novel P2X7 Receptor Radioligand as Marker for Neuroinflammation: Human Biodistribution, Dosimetry, Brain Kinetic Modelling and Quantification of Brain P2X7 Receptors in Patients with Parkinson’s Disease and Healthy Volunteers. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2051–2064. [Google Scholar] [CrossRef]
- Lucot, K.L.; Stevens, M.Y.; Bonham, T.A.; Azevedo, E.C.; Chaney, A.M.; Webber, E.D.; Jain, P.; Klockow, J.L.; Jackson, I.M.; Carlson, M.L.; et al. Tracking Innate Immune Activation in a Mouse Model of Parkinson’s Disease Using Trem1 and Tspo Pet Tracers. J. Nucl. Med. 2022, 63. [Google Scholar] [CrossRef]
- Horti, A.G.; Naik, R.; Foss, C.A.; Minn, I.; Misheneva, V.; Du, Y.; Wang, Y.; Mathews, W.B.; Wu, Y.; Hall, A.; et al. PET Imaging of Microglia by Targeting Macrophage Colony-Stimulating Factor 1 Receptor (CSF1R). Proc. Natl. Acad. Sci. USA 2019, 116, 1686–1691. [Google Scholar] [CrossRef]
- Hammes, J.; Drzezga, A.; van Eimeren, T. The Role of Tau Imaging in Parkinsonian Disorders. Curr. Neurol. Neurosci. Rep. 2018, 18, 86. [Google Scholar] [CrossRef]
- Ng, K.P.; Pascoal, T.A.; Mathotaarachchi, S.; Therriault, J.; Kang, M.S.; Shin, M.; Guiot, M.-C.; Guo, Q.; Harada, R.; Comley, R.A.; et al. Monoamine Oxidase B Inhibitor, Selegiline, Reduces 18F-THK5351 Uptake in the Human Brain. Alzheimers Res. Ther. 2017, 9, 25. [Google Scholar] [CrossRef]
- Bischof, G.N.; Dodich, A.; Boccardi, M.; van Eimeren, T.; Festari, C.; Barthel, H.; Hansson, O.; Nordberg, A.; Ossenkoppele, R.; Sabri, O.; et al. Clinical Validity of Second-Generation Tau PET Tracers as Biomarkers for Alzheimer’s Disease in the Context of a Structured 5-Phase Development Framework. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2110–2120. [Google Scholar] [CrossRef]
- Brendel, M.; Barthel, H.; van Eimeren, T.; Marek, K.; Beyer, L.; Song, M.; Palleis, C.; Gehmeyr, M.; Fietzek, U.; Respondek, G.; et al. Assessment of 18F-PI-2620 as a Biomarker in Progressive Supranuclear Palsy. JAMA Neurol. 2020, 77, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Willroider, M.; Roeber, S.; Horn, A.K.E.; Arzberger, T.; Scheifele, M.; Respondek, G.; Sabri, O.; Barthel, H.; Patt, M.; Mishchenko, O.; et al. Superiority of Formalin-Fixed Paraffin-Embedded Brain Tissue for in Vitro Assessment of Progressive Supranuclear Palsy Tau Pathology With [18F]PI-2620. Front. Neurol. 2021, 12, 684523. [Google Scholar] [CrossRef] [PubMed]
- Kroth, H.; Oden, F.; Molette, J.; Schieferstein, H.; Capotosti, F.; Mueller, A.; Berndt, M.; Schmitt-Willich, H.; Darmency, V.; Gabellieri, E.; et al. Discovery and Preclinical Characterization of [18F]PI-2620, a next-Generation Tau PET Tracer for the Assessment of Tau Pathology in Alzheimer’s Disease and Other Tauopathies. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, T.; Takahata, K.; Seki, M.; Tabuchi, H.; Momota, Y.; Shiraiwa, M.; Suzuki, N.; Morimoto, A.; Nakahara, T.; Iwabuchi, Y.; et al. Evaluation of [18F]PI-2620, a Second-Generation Selective Tau Tracer, for Assessing Four-Repeat Tauopathies. Brain Commun. 2021, 3, fcab190. [Google Scholar] [CrossRef]
- Palleis, C.; Brendel, M.; Finze, A.; Weidinger, E.; Bötzel, K.; Danek, A.; Beyer, L.; Nitschmann, A.; Kern, M.; Biechele, G.; et al. Cortical [18F]PI-2620 Binding Differentiates Corticobasal Syndrome Subtypes. Mov. Disord. 2021, 36, 2104–2115. [Google Scholar] [CrossRef]
- Song, M.; Beyer, L.; Kaiser, L.; Barthel, H.; van Eimeren, T.; Marek, K.; Nitschmann, A.; Scheifele, M.; Palleis, C.; Respondek, G.; et al. Binding Characteristics of [18F]PI-2620 Distinguish the Clinically Predicted Tau Isoform in Different Tauopathies by PET. J. Cereb. Blood Flow Metab. 2021, 41, 2957–2972. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, J.; Nordberg, A.; Ågren, H. Dissecting the Binding Profile of PET Tracers to Corticobasal Degeneration Tau Fibrils. ACS Chem. Neurosci. 2021, 12, 3487–3496. [Google Scholar] [CrossRef]
- Beyer, L.; Nitschmann, A.; Barthel, H.; van Eimeren, T.; Unterrainer, M.; Sauerbeck, J.; Marek, K.; Song, M.; Palleis, C.; Respondek, G.; et al. Early-Phase [18F]PI-2620 Tau-PET Imaging as a Surrogate Marker of Neuronal Injury. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 2911–2922. [Google Scholar] [CrossRef]
- Hammes, J.; Leuwer, I.; Bischof, G.N.; Drzezga, A.; van Eimeren, T. Multimodal Correlation of Dynamic [18F]-AV-1451 Perfusion PET and Neuronal Hypometabolism in [18F]-FDG PET. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 2249–2256. [Google Scholar] [CrossRef]
- Song, M.; Scheifele, M.; Barthel, H.; van Eimeren, T.; Beyer, L.; Marek, K.; Eckenweber, F.; Palleis, C.; Kaiser, L.; Finze, A.; et al. Feasibility of Short Imaging Protocols for [18F]PI-2620 Tau-PET in Progressive Supranuclear Palsy. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 3872–3885. [Google Scholar] [CrossRef]
- Tagai, K.; Ono, M.; Kubota, M.; Kitamura, S.; Takahata, K.; Seki, C.; Takado, Y.; Shinotoh, H.; Sano, Y.; Yamamoto, Y.; et al. High-Contrast In Vivo Imaging of Tau Pathologies in Alzheimer’s and Non-Alzheimer’s Disease Tauopathies. Neuron 2021, 109, 42–58.e8. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, F.-T.; Li, M.; Lu, J.-Y.; Sun, Y.-M.; Liang, X.; Bao, W.; Chen, Q.-S.; Li, X.-Y.; Zhou, X.-Y.; et al. Clinical Utility of 18F-APN-1607 Tau PET Imaging in Patients with Progressive Supranuclear Palsy. Mov. Disord. 2021, 36, 2314–2323. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Choi, J.Y.; Lee, S.H.; Ryu, Y.H.; Lee, M.S.; Lyoo, C.H. 18F-AV-1451 Binds to Putamen in Multiple System Atrophy. Mov. Disord. 2017, 32, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Kuebler, L.; Buss, S.; Leonov, A.; Ryazanov, S.; Schmidt, F.; Maurer, A.; Weckbecker, D.; Landau, A.M.; Lillethorup, T.P.; Bleher, D.; et al. [11C]MODAG-001-towards a PET Tracer Targeting α-Synuclein Aggregates. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 1759–1772. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Azpiazu, P.; Svedberg, M.; Higuchi, M.; Ono, M.; Jia, Z.; Sunnemark, D.; Elmore, C.S.; Schou, M.; Varrone, A. Identification and in Vitro Characterization of C05-01, a PBB3 Derivative with Improved Affinity for Alpha-Synuclein. Brain Res. 2020, 1749, 147131. [Google Scholar] [CrossRef]
- Yousefi, B.H.; Shi, K.; Arzberger, T.; Wester, H.J.; Schwaiger, M.; Yakushev, I.; Weber, W. Translational Study of a Novel Alpha-Synuclein PET Tracer Designed for First-in-Human Investigating. In Proceedings of the Nuklearmedizin—NuclearMedicine; Georg Thieme Verlag KG, New York, NY, USA, 27 March 2019; Volume 58, p. L25. [Google Scholar]
- Chen, Y.-F.; Bian, J.; Zhang, P.; Bu, L.-L.; Shen, Y.; Yu, W.-B.; Lu, X.-H.; Lin, X.; Ye, D.-Y.; Wang, J.; et al. Design, Synthesis and Identification of N, N-Dibenzylcinnamamide (DBC) Derivatives as Novel Ligands for α-Synuclein Fibrils by SPR Evaluation System. Bioorg. Med. Chem. 2020, 28, 115358. [Google Scholar] [CrossRef]
- Kuang, G.; Murugan, N.A.; Ågren, H. Mechanistic Insight into the Binding Profile of DCVJ and α-Synuclein Fibril Revealed by Multiscale Simulations. ACS Chem. Neurosci. 2019, 10, 610–617. [Google Scholar] [CrossRef]
- Korat, Š.; Bidesi, N.S.R.; Bonanno, F.; Di Nanni, A.; Hoàng, A.N.N.; Herfert, K.; Maurer, A.; Battisti, U.M.; Bowden, G.D.; Thonon, D.; et al. Alpha-Synuclein PET Tracer Development—An Overview about Current Efforts. Pharmaceuticals 2021, 14, 847. [Google Scholar] [CrossRef] [PubMed]
- Alzghool, O.M.; van Dongen, G.; van de Giessen, E.; Schoonmade, L.; Beaino, W. α-Synuclein Radiotracer Development and In Vivo Imaging: Recent Advancements and New Perspectives. Mov. Disord. 2022, 37, 936–948. [Google Scholar] [CrossRef]
- McCluskey, S.P.; Plisson, C.; Rabiner, E.A.; Howes, O. Advances in CNS PET: The State-of-the-Art for New Imaging Targets for Pathophysiology and Drug Development. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 451–489. [Google Scholar] [CrossRef] [Green Version]
- Murugan, N.A.; Nordberg, A.; Ågren, H. Different Positron Emission Tomography Tau Tracers Bind to Multiple Binding Sites on the Tau Fibril: Insight from Computational Modeling. ACS Chem. Neurosci. 2018, 9, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Stéen, E.J.L.; Vugts, D.J.; Windhorst, A.D. The Application of in Silico Methods for Prediction of Blood-Brain Barrier Permeability of Small Molecule PET Tracers. Front. Nucl. Med. 2022, 2, 853475. [Google Scholar] [CrossRef]
- Ataeinia, B.; Heidari, P. Artificial Intelligence and the Future of Diagnostic and Therapeutic Radiopharmaceutical Development: In Silico Smart Molecular Design. PET Clin. 2021, 16, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, W.; Yang, Y.; Murzin, A.G.; Falcon, B.; Kotecha, A.; van Beers, M.; Tarutani, A.; Kametani, F.; Garringer, H.J.; et al. Structure-Based Classification of Tauopathies. Nature 2021, 598, 359–363. [Google Scholar] [CrossRef]
- Sehlin, D.; Syvänen, S.; Ballanger, B.; Barthel, H.; Bischof, G.N.; Boche, D.; Boecker, H.; Bohn, K.P.; Borghammer, P.; Cross, D.; et al. Engineered Antibodies: New Possibilities for Brain PET? Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2848–2858. [Google Scholar] [CrossRef]
- Roshanbin, S.; Xiong, M.; Hultqvist, G.; Söderberg, L.; Zachrisson, O.; Meier, S.; Ekmark-Lewén, S.; Bergström, J.; Ingelsson, M.; Sehlin, D.; et al. In Vivo Imaging of Alpha-Synuclein with Antibody-Based PET. Neuropharmacology 2022, 208, 108985. [Google Scholar] [CrossRef] [PubMed]
- Bani Sadr, A.; Testart, N.; Tylski, P.; Scheiber, C. Reduced Scan Time in 123I-FP-CIT SPECT Imaging Using a Large-Field Cadmium-Zinc-Telluride Camera. Clin. Nucl. Med. 2019, 44, 568–569. [Google Scholar] [CrossRef]
- Bordonne, M.; Chawki, M.B.; Marie, P.-Y.; Zaragori, T.; Roch, V.; Grignon, R.; Imbert, L.; Verger, A. High-Quality Brain Perfusion SPECT Images May Be Achieved with a High-Speed Recording Using 360° CZT Camera. EJNMMI Phys. 2020, 7, 65. [Google Scholar] [CrossRef]
- Tecklenburg, K.; Forgács, A.; Apostolova, I.; Lehnert, W.; Klutmann, S.; Csirik, J.; Garutti, E.; Buchert, R. Performance Evaluation of a Novel Multi-Pinhole Collimator for Dopamine Transporter SPECT. Phys. Med. Biol. 2020, 65, 165015. [Google Scholar] [CrossRef]
- Villain, N.; Béra, G.; Habert, M.-O.; Kas, A.; Aubert, J.; Jaubert, O.; Valabregue, R.; Fernandez-Vidal, S.; Corvol, J.-C.; Mangone, G.; et al. Dopamine Denervation in the Functional Territories of the Striatum: A New MR and Atlas-Based 123I-FP-CIT SPECT Quantification Method. J. Neural Transm. 2021, 128, 1841–1852. [Google Scholar] [CrossRef]
- Mahlknecht, P.; Marini, K.; Werkmann, M.; Poewe, W.; Seppi, K. Prodromal Parkinson’s Disease: Hype or Hope for Disease-Modification Trials? Transl. Neurodegener. 2022, 11, 11. [Google Scholar] [CrossRef]
- Bischof, G.N.; Ewers, M.; Franzmeier, N.; Grothe, M.J.; Hoenig, M.; Kocagoncu, E.; Neitzel, J.; Rowe, J.B.; Strafella, A.; Drzezga, A.; et al. Connectomics and Molecular Imaging in Neurodegeneration. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 2819–2830. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Strafella, A.P. Chapter 8—Metabolic Imaging and Plasticity. In Handbook of Clinical Neurology; Quartarone, A., Ghilardi, M.F., Boller, F., Eds.; Neuroplasticity; Elsevier: Amsterdam, The Netherlands, 2022; Volume 184, pp. 121–132. [Google Scholar]
- Liu, S.-Y.; Wile, D.J.; Fu, J.F.; Valerio, J.; Shahinfard, E.; McCormick, S.; Mabrouk, R.; Vafai, N.; McKenzie, J.; Neilson, N.; et al. The Effect of LRRK2 Mutations on the Cholinergic System in Manifest and Premanifest Stages of Parkinson’s Disease: A Cross-Sectional PET Study. Lancet Neurol. 2018, 17, 309–316. [Google Scholar] [CrossRef]
- Tondo, G.; Esposito, M.; Dervenoulas, G.; Wilson, H.; Politis, M.; Pagano, G. Hybrid PET-MRI Applications in Movement Disorders. Int. Rev. Neurobiol. 2019, 144, 211–257. [Google Scholar] [CrossRef] [PubMed]
- Newman-Tancredi, A.; Depoortère, R.Y.; Kleven, M.S.; Kołaczkowski, M.; Zimmer, L. Translating Biased Agonists from Molecules to Medications: Serotonin 5-HT1A Receptor Functional Selectivity for CNS Disorders. Pharmacol. Ther. 2022, 229, 107937. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prange, S.; Theis, H.; Banwinkler, M.; van Eimeren, T. Molecular Imaging in Parkinsonian Disorders—What’s New and Hot? Brain Sci. 2022, 12, 1146. https://doi.org/10.3390/brainsci12091146
Prange S, Theis H, Banwinkler M, van Eimeren T. Molecular Imaging in Parkinsonian Disorders—What’s New and Hot? Brain Sciences. 2022; 12(9):1146. https://doi.org/10.3390/brainsci12091146
Chicago/Turabian StylePrange, Stéphane, Hendrik Theis, Magdalena Banwinkler, and Thilo van Eimeren. 2022. "Molecular Imaging in Parkinsonian Disorders—What’s New and Hot?" Brain Sciences 12, no. 9: 1146. https://doi.org/10.3390/brainsci12091146