
Unexpected Effect of IL-1β on the Function of GABAA Receptors in Pediatric Focal Cortical Dysplasia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Membrane Preparation
2.3. Xenopus Oocytes Electrophysiology
2.4. RNA-Seq Library Preparation and Sequencing
2.5. Bioinformatics Analysis of RNA-Seq Data
3. Results
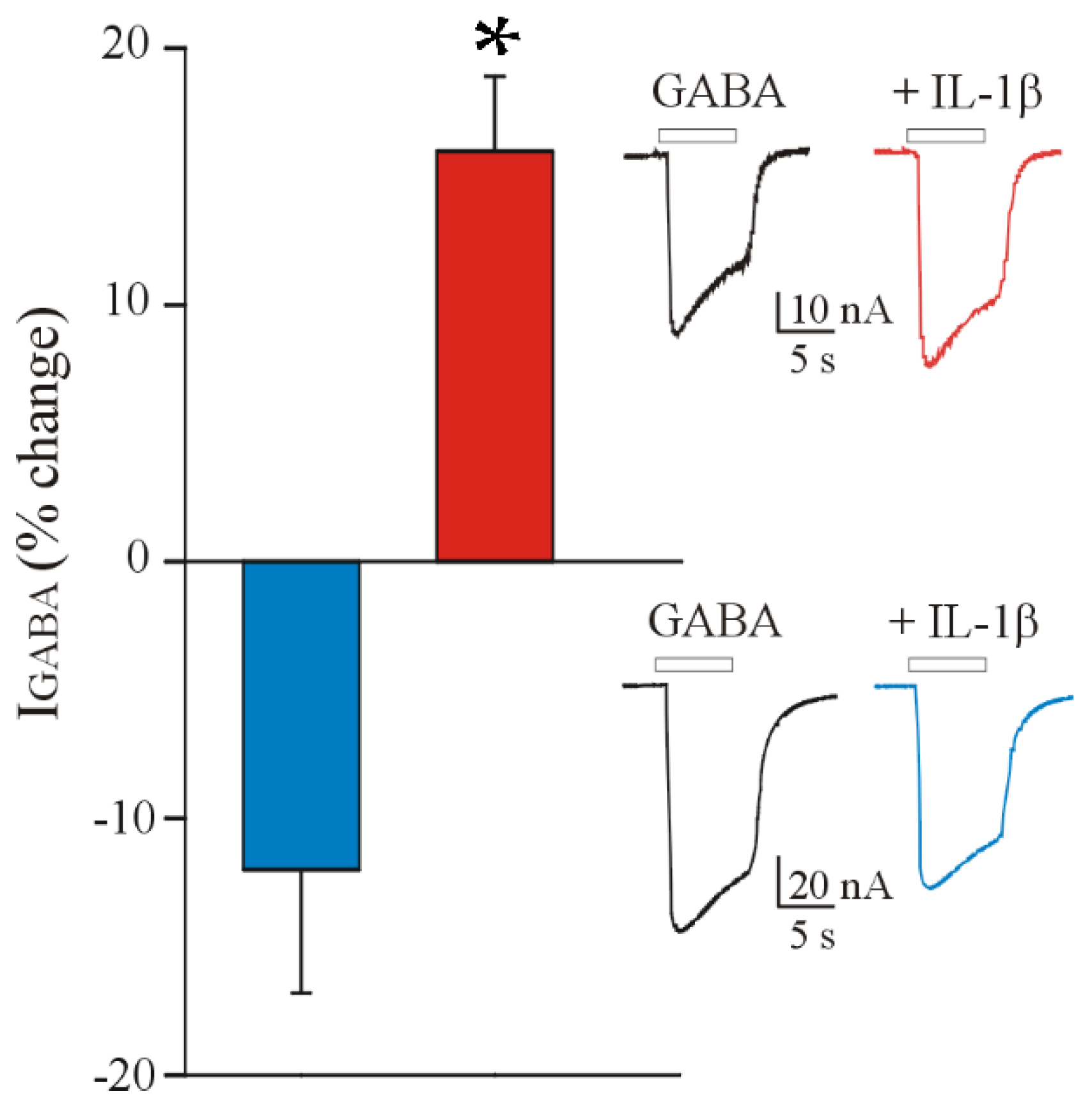
3.1. IL-1β Affects the GABA Current Amplitude in Oocytes Injected with FCD IIb Membranes
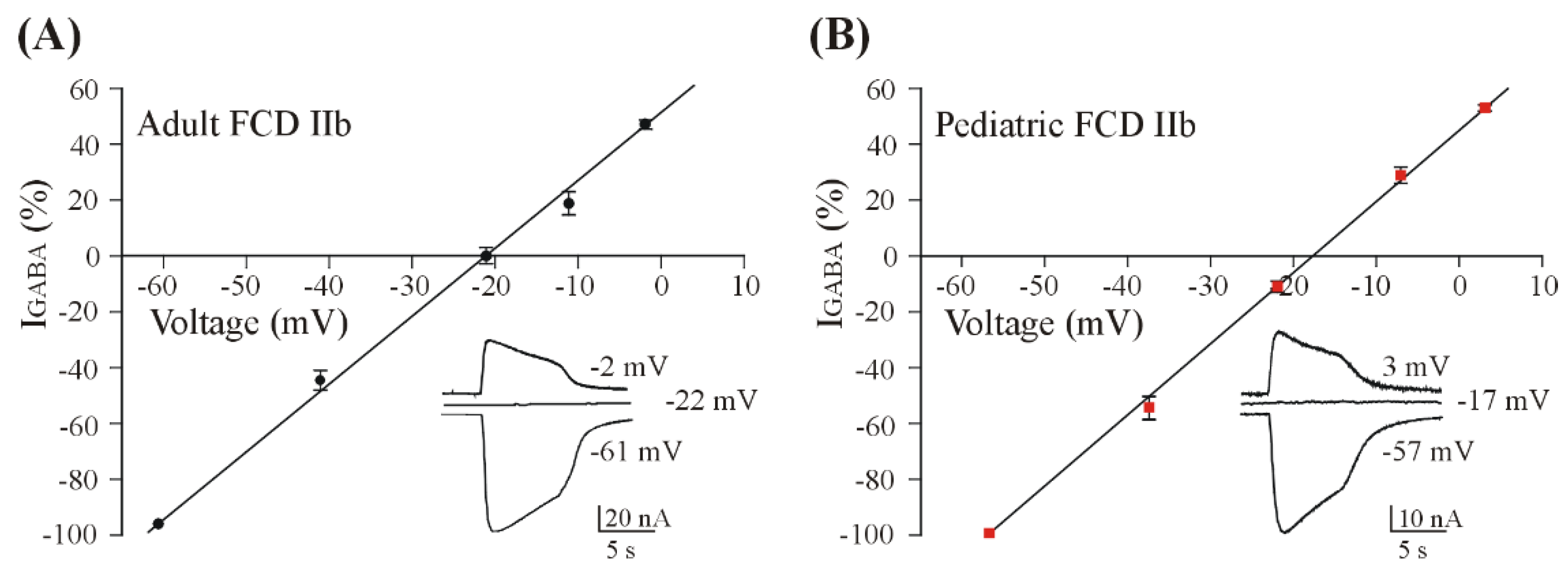
3.2. GABA Reversal Potential in Oocytes Injected with FCD IIb Membranes
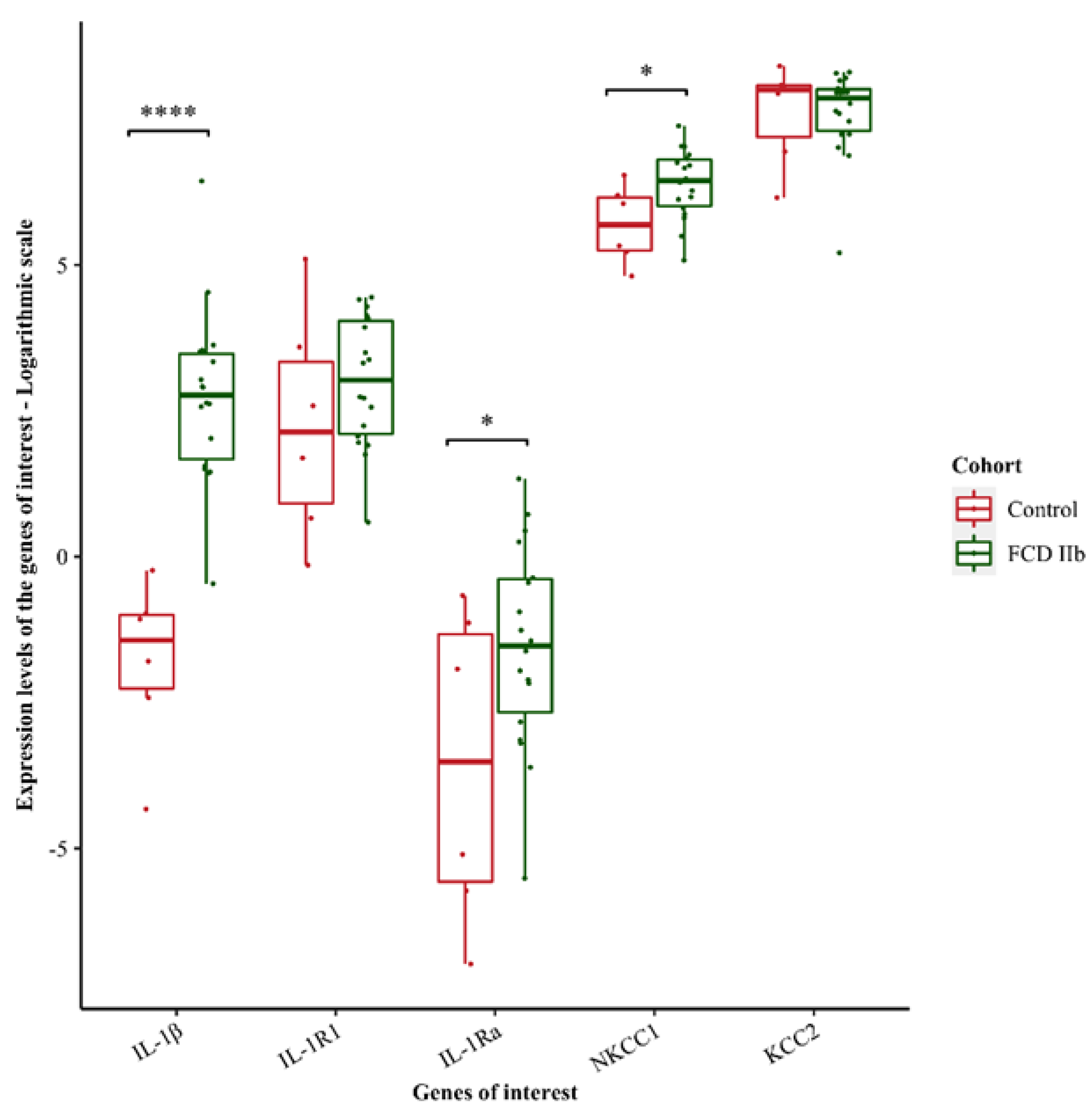
3.3. Transcriptomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iffland, P.H.; Crino, P.B. Focal Cortical Dysplasia: Gene Mutations, Cell Signaling, and Therapeutic Implications. Annu. Rev. Pathol. 2017, 12, 547–571. [Google Scholar] [CrossRef] [PubMed]
- Najm, I.; Lal, D.; Alonso Vanegas, M.; Cendes, F.; Lopes-Cendes, I.; Palmini, A.; Paglioli, E.; Sarnat, H.B.; Walsh, C.A.; Wiebe, S.; et al. The ILAE Consensus Classification of Focal Cortical Dysplasia: An Update Proposed by an Ad Hoc Task Force of the ILAE Diagnostic Methods Commission. Epilepsia 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Blumcke, I.; Spreafico, R.; Haaker, G.; Coras, R.; Kobow, K.; Bien, C.G.; Pfäfflin, M.; Elger, C.; Widman, G.; Schramm, J.; et al. Histopathological Findings in Brain Tissue Obtained during Epilepsy Surgery. N. Engl. J. Med. 2017, 377, 1648–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepeda, C.; André, V.M.; Levine, M.S.; Salamon, N.; Miyata, H.; Vinters, H.V.; Mathern, G.W. Epileptogenesis in Pediatric Cortical Dysplasia: The Dysmature Cerebral Developmental Hypothesis. Epilepsy Behav. 2006, 9, 219–235. [Google Scholar] [CrossRef]
- Blauwblomme, T.; Dossi, E.; Pellegrino, C.; Goubert, E.; Iglesias, B.G.; Sainte-Rose, C.; Rouach, N.; Nabbout, R.; Huberfeld, G. Gamma-Aminobutyric Acidergic Transmission Underlies Interictal Epileptogenicity in Pediatric Focal Cortical Dysplasia. Ann. Neurol. 2019, 85, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Cepeda, C.; Chen, J.Y.; Wu, J.Y.; Fisher, R.S.; Vinters, H.V.; Mathern, G.W.; Levine, M.S. Pacemaker GABA Synaptic Activity May Contribute to Network Synchronization in Pediatric Cortical Dysplasia. Neurobiol. Dis. 2014, 62, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Kaila, K.; Price, T.J.; Payne, J.A.; Puskarjov, M.; Voipio, J. Cation-Chloride Cotransporters in Neuronal Development, Plasticity and Disease. Nat. Rev. Neurosci. 2014, 15, 637–654. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA Excitatory/Inhibitory Shift in Brain Maturation and Neurological Disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef]
- Ben-Ari, Y. Excitatory Actions of Gaba during Development: The Nature of the Nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Cherubini, E.; Griguoli, M.; Safiulina, V.; Lagostena, L. The Depolarizing Action of GABA Controls Early Network Activity in the Developing Hippocampus. Mol. Neurobiol. 2011, 43, 97–106. [Google Scholar] [CrossRef]
- Talos, D.M.; Sun, H.; Kosaras, B.; Joseph, A.; Folkerth, R.D.; Poduri, A.; Madsen, J.R.; Black, P.M.; Jensen, F.E. Altered Inhibition in Tuberous Sclerosis and Type IIb Cortical Dysplasia. Ann. Neurol. 2012, 71, 539–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffolo, G.; Cifelli, P.; Miranda-Lourenço, C.; De Felice, E.; Limatola, C.; Sebastião, A.M.; Diógenes, M.J.; Aronica, E.; Palma, E. Rare Diseases of Neurodevelopment: Maintain the Mystery or Use a Dazzling Tool for Investigation? The Case of Rett Syndrome. Neuroscience 2019, 439, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Kim, J.; Zhou, L.; Wengert, E.; Zhang, L.; Wu, Z.; Carromeu, C.; Muotri, A.R.; Marchetto, M.C.N.; Gage, F.H.; et al. KCC2 Rescues Functional Deficits in Human Neurons Derived from Patients with Rett Syndrome. Proc. Natl. Acad. Sci. USA 2016, 113, 751–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffolo, G.; Cifelli, P.; Roseti, C.; Thom, M.; van Vliet, E.A.; Limatola, C.; Aronica, E.; Palma, E. A Novel GABAergic Dysfunction in Human Dravet Syndrome. Epilepsia 2018, 59, 2106–2117. [Google Scholar] [CrossRef] [Green Version]
- Cepeda, C.; André, V.M.; Wu, N.; Yamazaki, I.; Uzgil, B.; Vinters, H.V.; Levine, M.S.; Mathern, G.W. Immature Neurons and GABA Networks May Contribute to Epileptogenesis in Pediatric Cortical Dysplasia. Epilepsia 2007, 48 (Suppl. 5), 79–85. [Google Scholar] [CrossRef]
- Cepeda, C.; Levinson, S.; Nariai, H.; Yazon, V.-W.; Tran, C.; Barry, J.; Oikonomou, K.D.; Vinters, H.V.; Fallah, A.; Mathern, G.W.; et al. Pathological High Frequency Oscillations Associate with Increased GABA Synaptic Activity in Pediatric Epilepsy Surgery Patients. Neurobiol. Dis. 2020, 134, 104618. [Google Scholar] [CrossRef]
- Vezzani, A.; Viviani, B. Neuromodulatory Properties of Inflammatory Cytokines and Their Impact on Neuronal Excitability. Neuropharmacology 2015, 96 Pt A, 70–82. [Google Scholar] [CrossRef]
- Zimmer, T.S.; Broekaart, D.W.M.; Luinenburg, M.; Mijnsbergen, C.; Anink, J.J.; Sim, N.S.; Michailidou, I.; Jansen, F.E.; van Rijen, P.C.; Lee, J.H.; et al. Balloon Cells Promote Immune System Activation in Focal Cortical Dysplasia Type 2b. Neuropathol. Appl. Neurobiol. 2021, 47, 826–839. [Google Scholar] [CrossRef]
- Srivastava, A.; Dixit, A.B.; Paul, D.; Tripathi, M.; Sarkar, C.; Chandra, P.S.; Banerjee, J. Comparative Analysis of Cytokine/Chemokine Regulatory Networks in Patients with Hippocampal Sclerosis (HS) and Focal Cortical Dysplasia (FCD). Sci. Rep. 2017, 7, 15904. [Google Scholar] [CrossRef] [Green Version]
- Butler, T.; Li, Y.; Tsui, W.; Friedman, D.; Maoz, A.; Wang, X.; Harvey, P.; Tanzi, E.; Morim, S.; Kang, Y.; et al. Transient and Chronic Seizure-Induced Inflammation in Human Focal Epilepsy. Epilepsia 2016, 57, e191–e194. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory Pathways as Treatment Targets and Biomarkers in Epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Roseti, C.; van Vliet, E.A.; Cifelli, P.; Ruffolo, G.; Baayen, J.C.; Di Castro, M.A.; Bertollini, C.; Limatola, C.; Aronica, E.; Vezzani, A.; et al. GABAA Currents Are Decreased by IL-1β in Epileptogenic Tissue of Patients with Temporal Lobe Epilepsy: Implications for Ictogenesis. Neurobiol. Dis. 2015, 82, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Fuso, A.; Iyer, A.M.; van Scheppingen, J.; Maccarrone, M.; Scholl, T.; Hainfellner, J.A.; Feucht, M.; Jansen, F.E.; Spliet, W.G.; Krsek, P.; et al. Promoter-Specific Hypomethylation Correlates with IL-1β Overexpression in Tuberous Sclerosis Complex (TSC). J. Mol. Neurosci. 2016, 59, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Rossini, L.; Villani, F.; Granata, T.; Tassi, L.; Tringali, G.; Cardinale, F.; Aronica, E.; Spreafico, R.; Garbelli, R. FCD Type II and MTOR Pathway: Evidence for Different Mechanisms Involved in the Pathogenesis of Dysmorphic Neurons. Epilepsy Res. 2017, 129, 146–156. [Google Scholar] [CrossRef]
- Mills, J.D.; van Vliet, E.A.; Chen, B.J.; Janitz, M.; Anink, J.J.; Baayen, J.C.; Idema, S.; Devore, S.; Friedman, D.; Diehl, B.; et al. Coding and Non-Coding Transcriptome of Mesial Temporal Lobe Epilepsy: Critical Role of Small Non-Coding RNAs. Neurobiol. Dis. 2020, 134, 104612. [Google Scholar] [CrossRef]
- Palma, E.; Esposito, V.; Mileo, A.M.; Di Gennaro, G.; Quarato, P.; Giangaspero, F.; Scoppetta, C.; Onorati, P.; Trettel, F.; Miledi, R.; et al. Expression of Human Epileptic Temporal Lobe Neurotransmitter Receptors in Xenopus Oocytes: An Innovative Approach to Study Epilepsy. Proc. Natl. Acad. Sci. USA 2002, 99, 15078–15083. [Google Scholar] [CrossRef] [Green Version]
- Miledi, R.; Eusebi, F.; Martínez-Torres, A.; Palma, E.; Trettel, F. Expression of Functional Neurotransmitter Receptors in Xenopus Oocytes after Injection of Human Brain Membranes. Proc. Natl. Acad. Sci. USA 2002, 99, 13238–13242. [Google Scholar] [CrossRef] [Green Version]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate Paired Shotgun Read Merging via Overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The Reference Human Genome Annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon Provides Fast and Bias-Aware Quantification of Transcript Expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential Analyses for RNA-Seq: Transcript-Level Estimates Improve Gene-Level Inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Palma, E.; Amici, M.; Sobrero, F.; Spinelli, G.; Di Angelantonio, S.; Ragozzino, D.; Mascia, A.; Scoppetta, C.; Esposito, V.; Miledi, R.; et al. Anomalous Levels of Cl- Transporters in the Hippocampal Subiculum from Temporal Lobe Epilepsy Patients Make GABA Excitatory. Proc. Natl. Acad. Sci. USA 2006, 103, 8465–8468. [Google Scholar] [CrossRef] [Green Version]
- Palma, E.; Trettel, F.; Fucile, S.; Renzi, M.; Miledi, R.; Eusebi, F. Microtransplantation of Membranes from Cultured Cells to Xenopus Oocytes: A Method to Study Neurotransmitter Receptors Embedded in Native Lipids. Proc. Natl. Acad. Sci. USA 2003, 100, 2896–2900. [Google Scholar] [CrossRef] [Green Version]
- Blümcke, I.; Thom, M.; Aronica, E.; Armstrong, D.D.; Vinters, H.V.; Palmini, A.; Jacques, T.S.; Avanzini, G.; Barkovich, A.J.; Battaglia, G.; et al. The Clinico-Pathological Spectrum of Focal Cortical Dysplasias: A Consensus Classification Proposed by an Ad Hoc Task Force of the ILAE Diagnostic Methods Commission. Epilepsia 2011, 52, 158–174. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.; Wirrell, E.; Vezzani, A.; Nabbout, R.; Muscal, E.; Kaliakatsos, M.; Wickström, R.; Riviello, J.J.; Brunklaus, A.; Payne, E.; et al. Proposal to Optimize Evaluation and Treatment of Febrile Infection-Related Epilepsy Syndrome (FIRES): A Report from FIRES Workshop. Epilepsia Open 2021, 6, 62–72. [Google Scholar] [CrossRef]
- Pozzi, D.; Rasile, M.; Corradini, I.; Matteoli, M. Environmental Regulation of the Chloride Transporter KCC2: Switching Inflammation off to Switch the GABA On? Transl. Psychiatry 2020, 10, 349. [Google Scholar] [CrossRef]
- Semple, B.D.; O’Brien, T.J.; Gimlin, K.; Wright, D.K.; Kim, S.E.; Casillas-Espinosa, P.M.; Webster, K.M.; Petrou, S.; Noble-Haeusslein, L.J. Interleukin-1 Receptor in Seizure Susceptibility after Traumatic Injury to the Pediatric Brain. J. Neurosci. 2017, 37, 7864–7877. [Google Scholar] [CrossRef]
- Iori, V.; Frigerio, F.; Vezzani, A. Modulation of Neuronal Excitability by Immune Mediators in Epilepsy. Curr. Opin. Pharmacol. 2016, 26, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Ge, Q.; Yang, J.; Wang, M.; Zhou, Y.; Guo, J.-X.; Zhu, M.; Shi, J.; Hu, Y.; Shen, L.; et al. Pathological Networks Involving Dysmorphic Neurons in Type II Focal Cortical Dysplasia. Neurosci. Bull. 2022. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Ruffolo, G.; Iyer, A.; Cifelli, P.; Roseti, C.; Mühlebner, A.; van Scheppingen, J.; Scholl, T.; Hainfellner, J.A.; Feucht, M.; Krsek, P.; et al. Functional Aspects of Early Brain Development Are Preserved in Tuberous Sclerosis Complex (TSC) Epileptogenic Lesions. Neurobiol. Dis. 2016, 95, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Q.; Liu, M.; Wang, H.; Dong, Y.; Ji, T.; Liu, X.; Jiang, Y.; Cai, L.; Wu, Y. Upregulation of HMGB1-TLR4 Inflammatory Pathway in Focal Cortical Dysplasia Type II. J. Neuroinflammation 2018, 15, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Patient | Age at the Time of the Surgery (y) | Epilepsy Onset (y) | Gender | Diagnosis | ASMs |
|---|---|---|---|---|---|
| #1 | 2 | 0 | M | FCD IIb | OCZ, C, VPA, |
| #2 | 3 | 0 | F | FCD IIb | LEV, OCZ, C, VPA, CL |
| #3 | 5 | 3 | M | FCD IIb | C, CL |
| #4 | 18 | 2 | M | FCD IIb | C, VPA, LMT, LCM |
| #5 | 44 | 10 | F | FCD IIb | LEV, OCZ |
| #6 | 45 | 12 | M | FCD IIb | LEV, OCZ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfano, V.; Romagnolo, A.; Mills, J.D.; Cifelli, P.; Gaeta, A.; Morano, A.; Mühlebner, A.; Aronica, E.; Palma, E.; Ruffolo, G. Unexpected Effect of IL-1β on the Function of GABAA Receptors in Pediatric Focal Cortical Dysplasia. Brain Sci. 2022, 12, 807. https://doi.org/10.3390/brainsci12060807
Alfano V, Romagnolo A, Mills JD, Cifelli P, Gaeta A, Morano A, Mühlebner A, Aronica E, Palma E, Ruffolo G. Unexpected Effect of IL-1β on the Function of GABAA Receptors in Pediatric Focal Cortical Dysplasia. Brain Sciences. 2022; 12(6):807. https://doi.org/10.3390/brainsci12060807
Chicago/Turabian StyleAlfano, Veronica, Alessia Romagnolo, James D. Mills, Pierangelo Cifelli, Alessandro Gaeta, Alessandra Morano, Angelika Mühlebner, Eleonora Aronica, Eleonora Palma, and Gabriele Ruffolo. 2022. "Unexpected Effect of IL-1β on the Function of GABAA Receptors in Pediatric Focal Cortical Dysplasia" Brain Sciences 12, no. 6: 807. https://doi.org/10.3390/brainsci12060807