
Quantifying GABA in Addiction: A Review of Proton Magnetic Resonance Spectroscopy Studies

 ,
,
Abstract
:1. Introduction
1.1. GABA’s Role in Addiction
1.2. Proton Magnetic Resonance Spectroscopy (1H-MRS) in Clinical Research
2. Methods
3. GABAergic Alterations in Drug Addiction
3.1. Alcohol
3.2. Nicotine
3.3. Opioids
3.4. Cocaine
3.5. Methamphetamine
3.6. Cannabis
4. Technical Considerations of MRS in Addiction Studies
5. Discussion
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Volkow, N.D.; Michaelides, M.; Baler, R. The Neuroscience of Drug Reward and Addiction. Physiol. Rev. 2019, 99, 2115–2140. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurobiology of addiction: A neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Egerton, A. The potential of 1H-MRS in CNS drug development. Psychopharmacology 2021, 238, 1241–1254. [Google Scholar] [CrossRef]
- Chen, T.; Tan, H.; Lei, H.; Li, X.; Wu, Q.; Xu, X.; Ye, Y.; Zhong, N.; Du, J.; Jiang, H.; et al. Nature of glutamate alterations in substance dependence: A systematic review and meta-analysis of proton magnetic resonance spectroscopy studies. Psychiatry Res. Neuroimaging 2021, 315, 111329. [Google Scholar] [CrossRef]
- Yoon, S.; Kyoon Lyoo, I.; Renshaw, P.F. Application of magnetic resonance spectroscopic imaging to addiction research. In The Wiley-Blackwell Handbook of Addiction Psychopharmacology; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 707–750. [Google Scholar]
- Ende, G. Proton Magnetic Resonance Spectroscopy: Relevance of Glutamate and GABA to Neuropsychology. Neuropsychol. Rev. 2015, 25, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Kohut, S.J.; Kaufman, M.J. Magnetic resonance spectroscopy studies of substance use disorders: Current landscape and potential future directions. Pharm. Biochem. Behav. 2021, 200, 173090. [Google Scholar] [CrossRef]
- Moeller, S.J.; London, E.D.; Northoff, G. Neuroimaging markers of glutamatergic and GABAergic systems in drug addiction: Relationships to resting-state functional connectivity. Neurosci. Biobehav. Rev. 2016, 61, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Puts, N.A.; Edden, R.A. In vivo magnetic resonance spectroscopy of GABA: A methodological review. Prog. Nucl. Magn. Reson. Spectrosc. 2012, 60, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Peek, A.L.; Rebbeck, T.J.; Leaver, A.M.; Puts, N.A.; Foster, S.L.; Refshauge, K.M.; Oeltzschner, G. A Comprehensive Guide to Mega-Press for Gaba Measurement. medRxiv 2021. [Google Scholar] [CrossRef]
- Filip, M.; Frankowska, M.; Sadakierska-Chudy, A.; Suder, A.; Szumiec, L.; Mierzejewski, P.; Bienkowski, P.; Przegalinski, E.; Cryan, J.F. GABAB receptors as a therapeutic strategy in substance use disorders: Focus on positive allosteric modulators. Neuropharmacology 2015, 88, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Frankowska, M.; Przegaliński, E.; Filip, M. Targeting the GABAB Receptor for the Treatment of Substance Use Disorder. In GABAB Receptor; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 263–286. [Google Scholar] [CrossRef]
- Kumar, K.; Sharma, S.; Kumar, P.; Deshmukh, R. Therapeutic potential of GABA(B) receptor ligands in drug addiction, anxiety, depression and other CNS disorders. Pharm. Biochem. Behav. 2013, 110, 174–184. [Google Scholar] [CrossRef]
- Le Foll, B.; Wertheim, C.E.; Goldberg, S.R. Effects of baclofen on conditioned rewarding and discriminative stimulus effects of nicotine in rats. Neurosci. Lett. 2008, 443, 236–240. [Google Scholar] [CrossRef]
- Muncie, H.L., Jr.; Yasinian, Y.; Oge, L. Outpatient management of alcohol withdrawal syndrome. Am. Fam. Physician 2013, 88, 589–595. [Google Scholar]
- Prisciandaro, J.J.; Hoffman, M.; Brown, T.R.; Voronin, K.; Book, S.; Bristol, E.; Anton, R.F. Effects of Gabapentin on Dorsal Anterior Cingulate Cortex GABA and Glutamate Levels and Their Associations With Abstinence in Alcohol Use Disorder: A Randomized Clinical Trial. Am. J. Psychiatry 2021, 178, 829–837. [Google Scholar] [CrossRef]
- Prisciandaro, J.J.; Mellick, W.; Squeglia, L.M.; Hix, S.; Arnold, L.; Tolliver, B.K. Results from a randomized, double-blind, placebo-controlled, crossover, multimodal-MRI pilot study of gabapentin for co-occurring bipolar and cannabis use disorders. Addict. Biol. 2022, 27, e13085. [Google Scholar] [CrossRef]
- Prisciandaro, J.J.; Schacht, J.P.; Prescot, A.P.; Renshaw, P.F.; Brown, T.R.; Anton, R.F. Brain Glutamate, GABA, and Glutamine Levels and Associations with Recent Drinking in Treatment-Naïve Individuals with Alcohol Use Disorder Versus Light Drinkers. Alcohol. Clin. Exp. Res. 2019, 43, 221–226. [Google Scholar] [CrossRef]
- Schür, R.R.; Draisma, L.W.; Wijnen, J.P.; Boks, M.P.; Koevoets, M.G.; Joëls, M.; Klomp, D.W.; Kahn, R.S.; Vinkers, C.H. Brain GABA levels across psychiatric disorders: A systematic literature review and meta-analysis of 1H-MRS studies. Hum. Brain Mapp. 2016, 37, 3337–3352. [Google Scholar] [CrossRef]
- Prescot, A.P.; Renshaw, P.F.; Yurgelun-Todd, D.A. γ-Amino butyric acid and glutamate abnormalities in adolescent chronic marijuana smokers. Drug Alcohol Depend. 2013, 129, 232–239. [Google Scholar] [CrossRef]
- Shyu, C.; Elsaid, S.; Truong, P.; Chavez, S.; Le Foll, B. MR Spectroscopy of the Insula: Within- and between-Session Reproducibility of MEGA-PRESS Measurements of GABA+ and Other Metabolites. Brain Sci. 2021, 11, 1538. [Google Scholar] [CrossRef]
- Aufhaus, E.; Weber-Fahr, W.; Sack, M.; Tunc-Skarka, N.; Oberthuer, G.; Hoerst, M.; Meyer-Lindenberg, A.; Boettcher, U.; Ende, G. Absence of changes in GABA concentrations with age and gender in the human anterior cingulate cortex: A MEGA-PRESS study with symmetric editing pulse frequencies for macromolecule suppression. Magn. Reson. Med. 2013, 69, 317–320. [Google Scholar] [CrossRef]
- Licata, S.C.; Renshaw, P.F. Neurochemistry of drug action: Insights from Proton Magnetic Resonance Spectroscopic Imaging And Their Relevance to Addiction. Ann. N. Y. Acad. Sci. 2010, 1187, 148–171. [Google Scholar] [CrossRef]
- Owens, D.F.; Kriegstein, A.R. Is there more to GABA than synaptic inhibition? Nat. Rev. Neurosci. 2002, 3, 715–727. [Google Scholar] [CrossRef]
- Kleppner, S.R.; Tobin, A.J. GABA. In Encyclopedia of the Human Brain; Ramachandran, V.S., Ed.; Academic Press: New York, NY, USA, 2002; pp. 353–367. [Google Scholar] [CrossRef]
- Loi, B.; Maccioni, P.; Lobina, C.; Carai, M.A.; Gessa, G.L.; Thomas, A.W.; Malherbe, P.; Colombo, G. Reduction of alcohol intake by the positive allosteric modulator of the GABAB receptor, rac-BHFF, in alcohol-preferring rats. Alcohol 2013, 47, 69–73. [Google Scholar] [CrossRef]
- Maccioni, P.; Pes, D.; Orrù, A.; Froestl, W.; Gessa, G.L.; Carai, M.A.; Colombo, G. Reducing effect of the positive allosteric modulator of the GABAB receptor, GS39783, on alcohol self-administration in alcohol-preferring rats. Psychopharmacology 2007, 193, 171–178. [Google Scholar] [CrossRef]
- Liang, J.-H.; Chen, F.; Krstew, E.; Cowen, M.S.; Carroll, F.Y.; Crawford, D.; Beart, P.M.; Lawrence, A.J. The GABAB receptor allosteric modulator CGP7930, like baclofen, reduces operant self-administration of ethanol in alcohol-preferring rats. Neuropharmacology 2006, 50, 632–639. [Google Scholar] [CrossRef]
- Maccioni, P.; Fantini, N.; Froestl, W.; Carai, M.A.; Gessa, G.L.; Colombo, G. Specific reduction of alcohol’s motivational properties by the positive allosteric modulator of the GABAB receptor, GS39783—comparison with the effect of the GABAB receptor direct agonist, baclofen. Alcohol. Clin. Exp. Res. 2008, 32, 1558–1564. [Google Scholar] [CrossRef]
- Gao, H.; Xiang, Y.; Sun, N.; Zhu, H.; Wang, Y.; Liu, M.; Ma, Y.; Lei, H. Metabolic changes in rat prefrontal cortex and hippocampus induced by chronic morphine treatment studied ex vivo by high resolution 1H NMR spectroscopy. Neurochem. Int. 2007, 50, 386–394. [Google Scholar] [CrossRef]
- Fuchs, R.A.; Evans, K.A.; Ledford, C.C.; Parker, M.P.; Case, J.M.; Mehta, R.H.; See, R.E. The role of the dorsomedial prefrontal cortex, basolateral amygdala, and dorsal hippocampus in contextual reinstatement of cocaine seeking in rats. Neuropsychopharmacology 2005, 30, 296–309. [Google Scholar] [CrossRef]
- Sziráki, I.; Sershen, H.; Hashim, A.; Lajtha, A. Receptors in the ventral tegmental area mediating nicotine-induced dopamine release in the nucleus accumbens. Neurochem. Res. 2002, 27, 253–261. [Google Scholar] [CrossRef]
- Abreu-Villaça, Y.; Manhães, A.C.; Krahe, T.E.; Filgueiras, C.C.; Ribeiro-Carvalho, A. Tobacco and alcohol use during adolescence: Interactive mechanisms in animal models. Biochem. Pharmacol. 2017, 144, 1–17. [Google Scholar] [CrossRef]
- Li, X.; Semenova, S.; D’Souza, M.S.; Stoker, A.K.; Markou, A. Involvement of glutamatergic and GABAergic systems in nicotine dependence: Implications for novel pharmacotherapies for smoking cessation. Neuropharmacology 2014, 76, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.W.; Volkow, N.D. The neural basis of addiction: A pathology of motivation and choice. Am. J. Psychiatry 2005, 162, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Maccioni, P.; Bienkowski, P.; Carai, M.A.; Gessa, G.L.; Colombo, G. Baclofen attenuates cue-induced reinstatement of alcohol-seeking behavior in Sardinian alcohol-preferring (sP) rats. Drug Alcohol Depend. 2008, 95, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Slattery, D.A.; Markou, A.; Froestl, W.; Cryan, J.F. The GABAB receptor-positive modulator GS39783 and the GABAB receptor agonist baclofen attenuate the reward-facilitating effects of cocaine: Intracranial self-stimulation studies in the rat. Neuropsychopharmacology 2005, 30, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Minozzi, S.; Saulle, R.; Rosner, S. Baclofen for alcohol use disorder. Cochrane Database Syst. Rev. 2018, 11, CD012557. [Google Scholar] [CrossRef] [PubMed]
- Addolorato, G.; Leggio, L.; Ferrulli, A.; Cardone, S.; Vonghia, L.; Mirijello, A.; Abenavoli, L.; D’Angelo, C.; Caputo, F.; Zambon, A.; et al. Effectiveness and safety of baclofen for maintenance of alcohol abstinence in alcohol-dependent patients with liver cirrhosis: Randomised, double-blind controlled study. Lancet 2007, 370, 1915–1922. [Google Scholar] [CrossRef]
- De Beaurepaire, R.; Sinclair, J.M.A.; Heydtmann, M.; Addolorato, G.; Aubin, H.J.; Beraha, E.M.; Caputo, F.; Chick, J.D.; de La Selle, P.; Franchitto, N.; et al. The Use of Baclofen as a Treatment for Alcohol Use Disorder: A Clinical Practice Perspective. Front. Psychiatry 2018, 9, 708. [Google Scholar] [CrossRef]
- Morley, K.C.; Baillie, A.; Fraser, I.; Furneaux-Bate, A.; Dore, G.; Roberts, M.; Abdalla, A.; Phung, N.; Haber, P.S. Baclofen in the treatment of alcohol dependence with or without liver disease: Multisite, randomised, double-blind, placebo-controlled trial. Br. J. Psychiatry 2018, 212, 362–369. [Google Scholar] [CrossRef]
- Morley, K.C.; Lagopoulos, J.; Logge, W.; Chitty, K.; Baillie, A.; Haber, P.S. Neurometabolite Levels in Alcohol Use Disorder Patients During Baclofen Treatment and Prediction of Relapse to Heavy Drinking. Front. Psychiatry 2018, 9, 412. [Google Scholar] [CrossRef]
- Cosgrove, K.P.; Esterlis, I.; Mason, G.F.; Bois, F.; O’Malley, S.S.; Krystal, J.H. Neuroimaging insights into the role of cortical GABA systems and the influence of nicotine on the recovery from alcohol dependence. Neuropharmacology 2011, 60, 1318–1325. [Google Scholar] [CrossRef]
- Adinoff, B.; Kramer, G.L.; Petty, F. Levels of gamma-aminobutyric acid in cerebrospinal fluid and plasma during alcohol withdrawal. Psychiatry Res. 1995, 59, 137–144. [Google Scholar] [CrossRef]
- Mullins, P.G.; McGonigle, D.J.; O’Gorman, R.L.; Puts, N.A.; Vidyasagar, R.; Evans, C.J.; Edden, R.A. Current practice in the use of MEGA-PRESS spectroscopy for the detection of GABA. Neuroimage 2014, 86, 43–52. [Google Scholar] [CrossRef]
- Lukas, S.E. New perspectives on using brain imaging to study CNS stimulants. Neuropharmacology 2014, 87, 104–114. [Google Scholar] [CrossRef]
- Mason, G.F.; Krystal, J.H. MR spectroscopy: Its potential role for drug development for the treatment of psychiatric diseases. NMR Biomed. 2006, 19, 690–701. [Google Scholar] [CrossRef]
- Tapper, S.; Goransson, N.; Lundberg, P.; Tisell, A.; Zsigmond, P. A pilot study of essential tremor: Cerebellar GABA+/Glx ratio is correlated with tremor severity. Cerebellum Ataxias 2020, 7, 8. [Google Scholar] [CrossRef]
- Yanez Lopez, M.; Price, A.N.; Puts, N.A.J.; Hughes, E.J.; Edden, R.A.E.; McAlonan, G.M.; Arichi, T.; De Vita, E. Simultaneous quantification of GABA, Glx and GSH in the neonatal human brain using magnetic resonance spectroscopy. NeuroImage 2021, 233, 117930. [Google Scholar] [CrossRef]
- Bishop, J.H.; Faerman, A.; Geoly, A.; Khan, N.; Tischler, C.; Amin, H.; Maron-Katz, A.; Azeez, A.; Yeomans, D.C.; Hurd, R.; et al. Altered Neurotransmitter Ratio in the Prefrontal Cortex is Associated with Pain in Fibromyalgia Syndrome. medRxiv 2021. [Google Scholar] [CrossRef]
- Basu, S.K.; Pradhan, S.; du Plessis, A.J.; Ben-Ari, Y.; Limperopoulos, C. GABA and glutamate in the preterm neonatal brain: In-vivo measurement by magnetic resonance spectroscopy. NeuroImage 2021, 238, 118215. [Google Scholar] [CrossRef]
- Chang, L.; Cloak, C.C.; Ernst, T. Magnetic resonance spectroscopy studies of GABA in neuropsychiatric disorders. J. Clin. Psychiatry 2003, 64, 7–14. [Google Scholar]
- Martin, P.R.; Gibbs, S.J.; Nimmerrichter, A.A.; Riddle, W.R.; Welch, L.W.; Willcott, M.R. Brain proton magnetic resonance spectroscopy studies in recently abstinent alcoholics. Alcohol. Clin. Exp. Res. 1995, 19, 1078–1082. [Google Scholar] [CrossRef]
- Johnson, B.A. Recent Advances in the Development of Treatments for Alcohol and Cocaine Dependence. CNS Drugs 2005, 19, 873–896. [Google Scholar] [CrossRef]
- Barker, P.B.; Lin, D.D.M. In vivo proton MR spectroscopy of the human brain. Prog. Nucl. Magn. Reson. Spectrosc. 2006, 49, 99–128. [Google Scholar] [CrossRef]
- Bai, X.; Harris, A.D.; Gong, T.; Puts, N.A.; Wang, G.; Schär, M.; Barker, P.B.; Edden, R.A. Voxel placement precision for GABA-edited magnetic resonance spectroscopy. Open J. Radiol. 2017, 7, 35. [Google Scholar] [CrossRef]
- Deelchand, D.K.; Marjańska, M.; Henry, P.G.; Terpstra, M. MEGA-PRESS of GABA+: Influences of acquisition parameters. NMR Biomed. 2021, 34, e4199. [Google Scholar] [CrossRef] [PubMed]
- Baeshen, A.; Wyss, P.O.; Henning, A.; O’Gorman, R.L.; Piccirelli, M.; Kollias, S.; Michels, L. Test-Retest Reliability of the Brain Metabolites GABA and Glx With JPRESS, PRESS, and MEGA-PRESS MRS Sequences in vivo at 3T. J. Magn. Reson. Imaging 2020, 51, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.Y.; Andronesi, O.C.; Barker, P.; Bogner, W.; Edden, R.A.; Kaiser, L.G.; Lee, P.; Marjańska, M.; Terpstra, M.; de Graaf, R.A. Spectral editing in 1H magnetic resonance spectroscopy: Experts’ consensus recommendations. NMR Biomed. 2021, 34, e4411. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.; Bhardwaj, P.P.; Kalra, S.; Casault, C.A.; Yasmin, U.S.; Allen, P.S.; Coupland, N.J. Measurement of GABA and contaminants in gray and white matter in human brain in vivo. Magn. Reson. Med. 2007, 58, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, C.; Viant, M.R. Two-dimensional J-resolved NMR spectroscopy: Review of a key methodology in the metabolomics toolbox. Phytochem. Anal. 2010, 21, 22–32. [Google Scholar] [CrossRef]
- Ryner, L.N.; Sorenson, J.A.; Thomas, M.A. Localized 2D J-resolved 1H MR spectroscopy: Strong coupling effects in vitro and in vivo. Magn. Reson. Imaging 1995, 13, 853–869. [Google Scholar] [CrossRef]
- Jensen, J.E.; Licata, S.C.; Ongur, D.; Friedman, S.D.; Prescot, A.P.; Henry, M.E.; Renshaw, P.F. Quantification of J-resolved proton spectra in two-dimensions with LCModel using GAMMA-simulated basis sets at 4 Tesla. NMR Biomed. 2009, 22, 762–769. [Google Scholar] [CrossRef]
- Provencher, S.W. Automatic quantitation of localizedin vivo1H spectra with LCModel. NMR Biomed. 2001, 14, 260–264. [Google Scholar] [CrossRef]
- Behar, K.L.; Rothman, D.L.; Petersen, K.F.; Hooten, M.; Delaney, R.; Petroff, O.A.C.; Shulman, G.I.; Navarro, V.; Petrakis, I.L.; Charney, D.S.; et al. Preliminary Evidence of Low Cortical GABA Levels in Localized1H-MR Spectra of Alcohol-Dependent and Hepatic Encephalopathy Patients. Am. J. Psychiatry 1999, 156, 952–954. [Google Scholar] [CrossRef]
- Mon, A.; Durazzo, T.C.; Meyerhoff, D.J. Glutamate, GABA, and other cortical metabolite concentrations during early abstinence from alcohol and their associations with neurocognitive changes. Drug Alcohol Depend. 2012, 125, 27–36. [Google Scholar] [CrossRef]
- Pennington, D.L.; Abe, C.; Batki, S.L.; Meyerhoff, D.J. A preliminary examination of cortical neurotransmitter levels associated with heavy drinking in posttraumatic stress disorder. Psychiatry Res. 2014, 224, 281–287. [Google Scholar] [CrossRef]
- Prisciandaro, J.J.; Tolliver, B.K.; Prescot, A.P.; Brenner, H.M.; Renshaw, P.F.; Brown, T.R.; Anton, R.F. Unique prefrontal GABA and glutamate disturbances in co-occurring bipolar disorder and alcohol dependence. Transl. Psychiatry 2017, 7, e1163. [Google Scholar] [CrossRef]
- Prisciandaro, J.J.; Schacht, J.P.; Prescot, A.P.; Brenner, H.M.; Renshaw, P.F.; Brown, T.R.; Anton, R.F. Intraindividual changes in brain GABA, glutamate, and glutamine during monitored abstinence from alcohol in treatment-naive individuals with alcohol use disorder. Addict. Biol. 2020, 25, e12810. [Google Scholar] [CrossRef]
- Wang, G.; Weber-Fahr, W.; Frischknecht, U.; Hermann, D.; Kiefer, F.; Ende, G.; Sack, M. Cortical Glutamate and GABA Changes During Early Abstinence in Alcohol Dependence and Their Associations With Benzodiazepine Medication. Front. Psychiatry 2021, 12, 656468. [Google Scholar] [CrossRef]
- Mason, G.F.; Petrakis, I.L.; de Graaf, R.A.; Gueorguieva, R.; Guidone, E.; Coric, V.; Epperson, C.N.; Rothman, D.L.; Krystal, J.H. Cortical gamma-aminobutyric acid levels and the recovery from ethanol dependence: Preliminary evidence of modification by cigarette smoking. Biol. Psychiatry 2006, 59, 85–93. [Google Scholar] [CrossRef]
- Epperson, C.N.; O’Malley, S.; Czarkowski, K.A.; Gueorguieva, R.; Jatlow, P.; Sanacora, G.; Rothman, D.L.; Krystal, J.H.; Mason, G.F. Sex, GABA, and nicotine: The impact of smoking on cortical GABA levels across the menstrual cycle as measured with proton magnetic resonance spectroscopy. Biol. Psychiatry 2005, 57, 44–48. [Google Scholar] [CrossRef]
- Bagga, D.; Aigner, C.S.; Cecchetto, C.; Fischmeister, F.P.S.; Schopf, V. Investigating Sex-Specific Characteristics of Nicotine Addiction Using Metabolic and Structural Magnetic Resonance Imaging. Eur. Addict. Res. 2018, 24, 267–277. [Google Scholar] [CrossRef]
- Janes, A.C.; Jensen, J.E.; Farmer, S.L.; Frederick, B.D.; Pizzagalli, D.A.; Lukas, S.E. GABA levels in the dorsal anterior cingulate cortex associated with difficulty ignoring smoking-related cues in tobacco-dependent volunteers. Neuropsychopharmacology 2013, 38, 1113–1120. [Google Scholar] [CrossRef]
- Ke, Y.; Streeter, C.C.; Nassar, L.E.; Sarid-Segal, O.; Hennen, J.; Yurgelun-Todd, D.A.; Awad, L.A.; Rendall, M.J.; Gruber, S.A.; Nason, A.; et al. Frontal lobe GABA levels in cocaine dependence: A two-dimensional, J-resolved magnetic resonance spectroscopy study. Psychiatry Res. 2004, 130, 283–293. [Google Scholar] [CrossRef]
- Hulka, L.M.; Scheidegger, M.; Vonmoos, M.; Preller, K.H.; Baumgartner, M.R.; Herdener, M.; Seifritz, E.; Henning, A.; Quednow, B.B. Glutamatergic and neurometabolic alterations in chronic cocaine users measured with (1) H-magnetic resonance spectroscopy. Addict. Biol. 2016, 21, 205–217. [Google Scholar] [CrossRef]
- Ersche, K.D.; Lim, T.V.; Murley, A.G.; Rua, C.; Vaghi, M.M.; White, T.L.; Williams, G.B.; Robbins, T.W. Reduced Glutamate Turnover in the Putamen Is Linked With Automatic Habits in Human Cocaine Addiction. Biol. Psychiatry 2021, 89, 970–979. [Google Scholar] [CrossRef]
- Su, H.; Chen, T.; Zhong, N.; Jiang, H.; Du, J.; Xiao, K.; Xu, D.; Song, W.; Zhao, M. Decreased GABA concentrations in left prefrontal cortex of methamphetamine dependent patients: A proton magnetic resonance spectroscopy study. J. Clin. Neurosci. 2020, 71, 15–20. [Google Scholar] [CrossRef]
- Li, J.N.; Liu, X.L.; Li, L. Prefrontal GABA and glutamate levels correlate with impulsivity and cognitive function of prescription opioid addicts: A 1 H-magnetic resonance spectroscopy study. Psychiatry Clin. Neurosci. 2020, 74, 77–83. [Google Scholar] [CrossRef]
- Murray, D.E.; Durazzo, T.C.; Schmidt, T.P.; Abé, C.; Guydish, J.; Meyerhoff, D.J. Frontal Metabolite Concentration Deficits in Opiate Dependence Relate to Substance Use, Cognition, and Self-Regulation. J. Addict. Res. Ther. 2016, 7, 286. [Google Scholar] [CrossRef]
- Schulte, M.H.J.; Kaag, A.M.; Wiers, R.W.; Schmaal, L.; van den Brink, W.; Reneman, L.; Homberg, J.R.; van Wingen, G.A.; Goudriaan, A.E. Prefrontal Glx and GABA concentrations and impulsivity in cigarette smokers and smoking polysubstance users. Drug Alcohol Depend. 2017, 179, 117–123. [Google Scholar] [CrossRef]
- Abe, C.; Mon, A.; Durazzo, T.C.; Pennington, D.L.; Schmidt, T.P.; Meyerhoff, D.J. Polysubstance and alcohol dependence: Unique abnormalities of magnetic resonance-derived brain metabolite levels. Drug Alcohol Depend. 2013, 130, 30–37. [Google Scholar] [CrossRef]
- De Bondt, T.; De Belder, F.; Vanhevel, F.; Jacquemyn, Y.; Parizel, P.M. Prefrontal GABA concentration changes in women—Influence of menstrual cycle phase, hormonal contraceptive use, and correlation with premenstrual symptoms. Brain Res. 2015, 1597, 129–138. [Google Scholar] [CrossRef]
- Epperson, C.N.; Haga, K.; Mason, G.F.; Sellers, E.; Gueorguieva, R.; Zhang, W.; Weiss, E.; Rothman, D.L.; Krystal, J.H. Cortical γ-Aminobutyric Acid Levels Across the Menstrual Cycle in Healthy Women and Those With Premenstrual Dysphoric Disorder. Arch. Gen. Psychiatry 2002, 59, 851. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, R.L.; Michels, L.; Edden, R.A.; Murdoch, J.B.; Martin, E. In vivo detection of GABA and glutamate with MEGA-PRESS: Reproducibility and gender effects. J. Magn. Reson. Imaging 2011, 33, 1262–1267. [Google Scholar] [CrossRef]
- Pandya, M.; Palpagama, T.H.; Turner, C.; Waldvogel, H.J.; Faull, R.L.; Kwakowsky, A. Sex- and age-related changes in GABA signaling components in the human cortex. Biol. Sex Differ. 2019, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Rothman, D.L.; Petroff, O.A.; Behar, K.L.; Mattson, R.H. Localized 1H NMR measurements of gamma-aminobutyric acid in human brain in vivo. Proc. Natl. Acad. Sci. USA 1993, 90, 5662–5666. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, I.K.; Renshaw, P.F. Magnetic resonance spectroscopy: Current and future applications in psychiatric research. Biol. Psychiatry 2002, 51, 195–207. [Google Scholar] [CrossRef]
- Mullins, P.G.; Chen, H.; Xu, J.; Caprihan, A.; Gasparovic, C. Comparative reliability of proton spectroscopy techniques designed to improve detection of J-coupled metabolites. Magn. Reson. Med. 2008, 60, 964–969. [Google Scholar] [CrossRef]
- Duda, J.M.; Moser, A.D.; Zuo, C.S.; Du, F.; Chen, X.; Perlo, S.; Richards, C.E.; Nascimento, N.; Ironside, M.; Crowley, D.J.; et al. Repeatability and reliability of GABA measurements with magnetic resonance spectroscopy in healthy young adults. Magn. Reson. Med. 2021, 85, 2359–2369. [Google Scholar] [CrossRef]
- Brix, M.K.; Ersland, L.; Hugdahl, K.; Dwyer, G.E.; Gruner, R.; Noeske, R.; Beyer, M.K.; Craven, A.R. Within- and between-session reproducibility of GABA measurements with MR spectroscopy. J. Magn. Reson. Imaging 2017, 46, 421–430. [Google Scholar] [CrossRef]
- Sears, S.M.; Hewett, S.J. Influence of glutamate and GABA transport on brain excitatory/inhibitory balance. Exp. Biol. Med. 2021, 246, 1069–1083. [Google Scholar] [CrossRef]
- Mikkelsen, M.; Loo, R.S.; Puts, N.A.; Edden, R.A.; Harris, A.D. Designing GABA-edited magnetic resonance spectroscopy studies: Considerations of scan duration, signal-to-noise ratio and sample size. J. Neurosci. Methods 2018, 303, 86–94. [Google Scholar] [CrossRef]
- Mikkelsen, M.; Rimbault, D.L.; Barker, P.B.; Bhattacharyya, P.K.; Brix, M.K.; Buur, P.F.; Cecil, K.M.; Chan, K.L.; Chen, D.Y.; Craven, A.R.; et al. Big GABA II: Water-referenced edited MR spectroscopy at 25 research sites. Neuroimage 2019, 191, 537–548. [Google Scholar] [CrossRef]
- Harris, A.D.; Puts, N.A.; Edden, R.A. Tissue correction for GABA-edited MRS: Considerations of voxel composition, tissue segmentation, and tissue relaxations. J. Magn. Reson. Imaging 2015, 42, 1431–1440. [Google Scholar] [CrossRef]
- Kreis, R.; Boer, V.; Choi, I.Y.; Cudalbu, C.; de Graaf, R.A.; Gasparovic, C.; Heerschap, A.; Krššák, M.; Lanz, B.; Maudsley, A.A. Terminology and concepts for the characterization of in vivo MR spectroscopy methods and MR spectra: Background and experts’ consensus recommendations. NMR Biomed. 2021, 34, e4347. [Google Scholar] [CrossRef]
- Sandberg, S.G.; Garris, P.A. Neurochemistry of Addiction: Monitoring Essential Neurotransmitters of Addiction, 2nd ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2010. [Google Scholar]
- Lin, A.; Andronesi, O.; Bogner, W.; Choi, I.Y.; Coello, E.; Cudalbu, C.; Juchem, C.; Kemp, G.J.; Kreis, R.; Krššák, M.; et al. Minimum Reporting Standards for in vivo Magnetic Resonance Spectroscopy (MRSinMRS): Experts’ consensus recommendations. NMR Biomed. 2021, 34, e4484. [Google Scholar] [CrossRef]
- Edden, R.A.E.; Puts, N.A.J.; Harris, A.D.; Barker, P.B.; Evans, C.J. Gannet: A batch-processing tool for the quantitative analysis of gamma-aminobutyric acid–edited MR spectroscopy spectra. J. Magn. Reson. Imaging 2014, 40, 1445–1452. [Google Scholar] [CrossRef]
- Mescher, M.; Merkle, H.; Kirsch, J.; Garwood, M.; Gruetter, R. Simultaneous in vivo spectral editing and water suppression. NMR Biomed. 1998, 11, 266–272. [Google Scholar] [CrossRef]
- Ashburner, J.; Barnes, G.; Chen, C.-C.; Daunizeau, J.; Flandin, G.; Friston, K.; Kiebel, S.; Kilner, J.; Litvak, V.; Moran, R. SPM12 Manual; Wellcome Trust Centre for Neuroimaging: London, UK, 2014; Volume 2464, p. 4. [Google Scholar]
- Van Veenendaal, T.M.; Backes, W.H.; van Bussel, F.C.G.; Edden, R.A.E.; Puts, N.A.J.; Aldenkamp, A.P.; Jansen, J.F.A. Glutamate quantification by PRESS or MEGA-PRESS: Validation, repeatability, and concordance. Magn. Reson. Imaging 2018, 48, 107–114. [Google Scholar] [CrossRef]
- Bogner, W.; Gruber, S.; Doelken, M.; Stadlbauer, A.; Ganslandt, O.; Boettcher, U.; Trattnig, S.; Doerfler, A.; Stefan, H.; Hammen, T. In vivo quantification of intracerebral GABA by single-voxel (1)H-MRS-How reproducible are the results? Eur. J. Radiol. 2010, 73, 526–531. [Google Scholar] [CrossRef]
- Long, Z.; Dyke, J.P.; Ma, R.; Huang, C.C.; Louis, E.D.; Dydak, U. Reproducibility and effect of tissue composition on cerebellar γ-aminobutyric acid (GABA) MRS in an elderly population. NMR Biomed. 2015, 28, 1315–1323. [Google Scholar] [CrossRef]
- Waddell, K.W.; Zanjanipour, P.; Pradhan, S.; Xu, L.; Welch, E.B.; Joers, J.M.; Martin, P.R.; Avison, M.J.; Gore, J.C. Anterior cingulate and cerebellar GABA and Glu correlations measured by (1) H J-difference spectroscopy. Magn. Reson. Imaging 2011, 29, 19–24. [Google Scholar] [CrossRef]
- Shungu, D.C.; Mao, X.; Gonzales, R.; Soones, T.N.; Dyke, J.P.; van der Veen, J.W.; Kegeles, L.S. Brain gamma-aminobutyric acid (GABA) detection in vivo with the J-editing (1) H MRS technique: A comprehensive methodological evaluation of sensitivity enhancement, macromolecule contamination and test-retest reliability. NMR Biomed. 2016, 29, 932–942. [Google Scholar] [CrossRef]
- Near, J.; Ho, Y.-C.L.; Sandberg, K.; Kumaragamage, C.; Blicher, J.U. Long-term reproducibility of GABA magnetic resonance spectroscopy. NeuroImage 2014, 99, 191–196. [Google Scholar] [CrossRef]
- Xin, L.; Mlynarik, V.; Lei, H.; Gruetter, R. Influence of regional macromolecule baseline on the quantification of neurochemical profile in rat brain. In Proceedings of the Joint Annual Meeting ISMRM-ESMRMB 2010, Stockholm, Sweden, 1–7 May 2010. [Google Scholar]
- Harris, A.D.; Puts, N.A.J.; Barker, P.B.; Edden, R.A.E. Spectral-editing measurements of GABA in the human brain with and without macromolecule suppression. Magn. Reson. Med. 2015, 74, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Meyerhoff, D.J.; Durazzo, T.C.; Ende, G. Chronic Alcohol Consumption, Abstinence and Relapse: Brain Proton Magnetic Resonance Spectroscopy Studies in Animals and Humans. In Behavioral Neurobiology of Alcohol Addiction; Sommer, W.H., Spanagel, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 511–540. [Google Scholar] [CrossRef]
- Seo, H.S.; Jeong, E.K.; Choi, S.; Kwon, Y.; Park, H.J.; Kim, I. Changes of Neurotransmitters in Youth with Internet and Smartphone Addiction: A Comparison with Healthy Controls and Changes after Cognitive Behavioral Therapy. AJNR Am. J. Neuroradiol. 2020, 41, 1293–1301. [Google Scholar] [CrossRef]
- Dickler, M.; Lenglos, C.; Renauld, E.; Ferland, F.; Edden, R.A.; Leblond, J.; Fecteau, S. Online effects of transcranial direct current stimulation on prefrontal metabolites in gambling disorder. Neuropharmacology 2018, 131, 51–57. [Google Scholar] [CrossRef]
- Perrine, S.A.; Ghoddoussi, F.; Michaels, M.S.; Sheikh, I.S.; McKelvey, G.; Galloway, M.P. Ketamine reverses stress-induced depression-like behavior and increased GABA levels in the anterior cingulate: An 11.7 T 1H-MRS study in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 51, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Sarlo, G.L.; Holton, K.F. Brain concentrations of glutamate and GABA in human epilepsy: A review. Seizure 2021, 91, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Morley, K.C.; Lagopoulos, J.; Logge, W.; Baillie, A.; Adams, C.; Haber, P.S. Brain GABA levels are reduced in alcoholic liver disease: A proton magnetic resonance spectroscopy study. Addict. Biol. 2020, 25, e12702. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, N.H.; Bechara, A. The hidden island of addiction: The insula. Trends Neurosci. 2009, 32, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, C.; Rubin-Kahana, D.S.; Pushparaj, A.; Musiol, M.; Blumberger, D.M.; Daskalakis, Z.J.; Zangen, A.; Le Foll, B. The Insula: A Brain Stimulation Target for the Treatment of Addiction. Front. Pharmacol. 2019, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, N.H.; Gaznick, N.; Tranel, D.; Bechara, A. The insula: A critical neural substrate for craving and drug seeking under conflict and risk. Ann. N. Y. Acad. Sci. 2014, 1316, 53–70. [Google Scholar] [CrossRef]
- Droutman, V.; Read, S.J.; Bechara, A. Revisiting the role of the insula in addiction. Trends Cogn. Sci. 2015, 19, 414–420. [Google Scholar] [CrossRef]



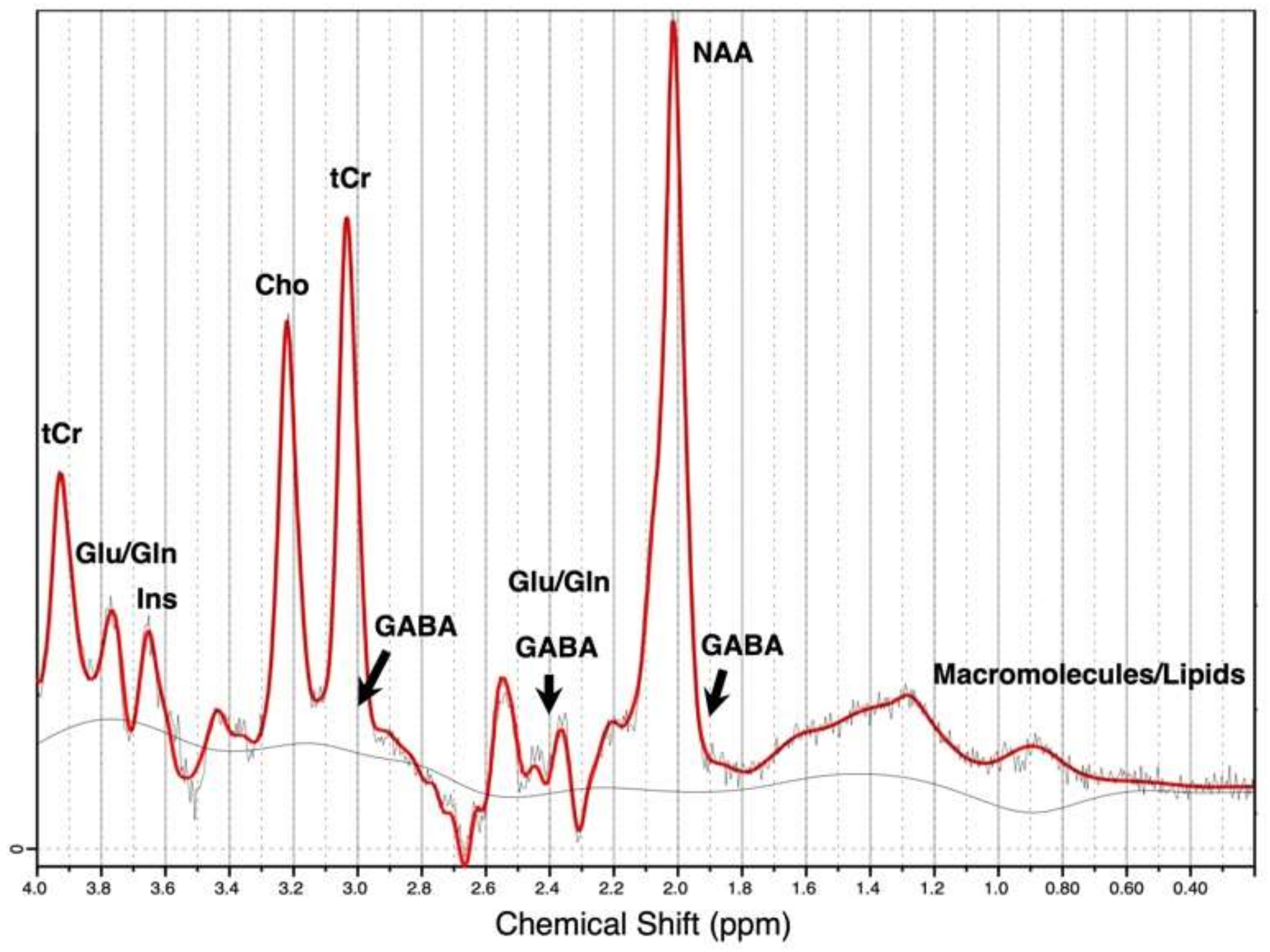{kind=link}
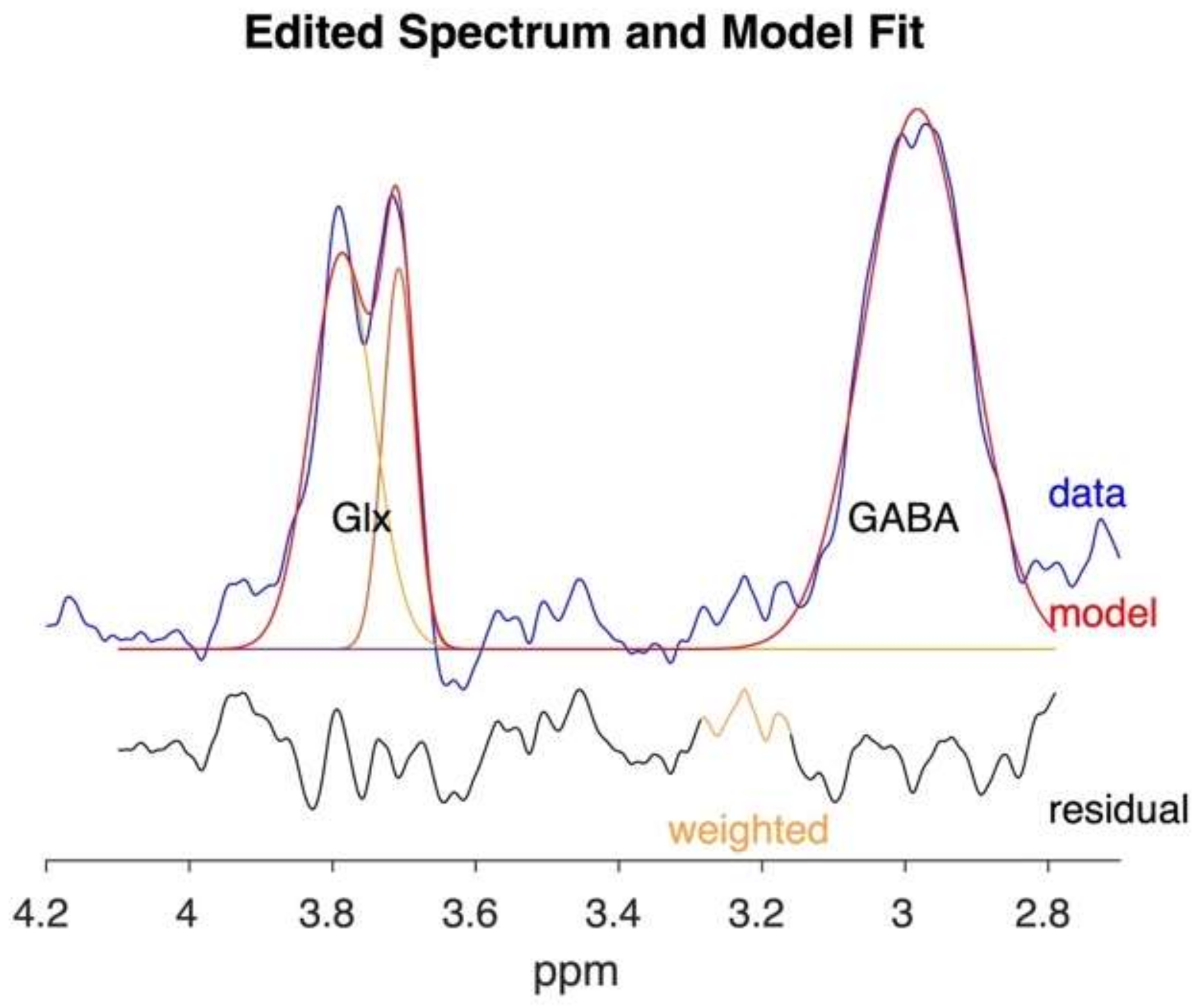{kind=link}
| ACC | dACC | pgACC | Occipital Cortex | POC | TEMP | DLPFC | PFC | OFC | Left Putamen | |
|---|---|---|---|---|---|---|---|---|---|---|
| Alcohol | -/↑ */↓ | -/↓ | ↓ | - * | - | - * | - | |||
| Nicotine | - | ↓ (Female only) | ↑ | |||||||
| Opioids | - | - | - | ↓ | - | |||||
| Cocaine | - | - | ↓ | - | ||||||
| Methamphetamine | ↓ | |||||||||
| Cannabis | ↓ | |||||||||
| Polysubstance | -/↓ | - | - | - | - |
| Substance | Study | Subjects | Sex (Male/Female) | Mean Age (SD) | Imaging Details | Region Volume (Voxel Dimensions) | Results on GABA | Clinical Correlations |
|---|---|---|---|---|---|---|---|---|
| Alcohol | Behar et al., 1999 [65] |
|
|
|
| Occipital cortex 13.5 mL |
| NA |
| Alcohol | Mon et al., 2012 [66] |
|
|
|
| ACC 17.5 mL (35 × 25 × 20 mm3) POC 16 mL (20 × 40 × 20 mm3) Right DLPFC 16 mL (40 × 20 × 20 mm3) |
| NA |
| Alcohol | Pennington et al., 2014 [67] |
|
|
|
| ACC 17.5 mL (35 × 25 × 20 mm3) POC 16 mL (20 × 40 × 20 mm3) TEMP 16 mL (20 × 40 × 20 mm3) |
|
|
| Alcohol | Prisciandaro et al., 2017 [68] |
|
|
|
| dACC 18.8 mL (25 × 25 × 30 mm3) |
|
|
| Alcohol | Prisciandaro et al., 2019 [18] |
|
|
|
| dACC 18.8 mL (25 × 25 × 30 mm3) |
| NA |
| Alcohol | Prisciandaro et al., 2020 [69] |
|
|
|
| dACC 18.8 mL (25 × 25 × 30 mm3) |
| NA |
| Alcohol | Wang et al., 2021 [70] |
|
|
|
| ACC 24 mL (20 × 30 × 40 mm3) |
| NA |
| Nicotine+ alcohol | Mason et al., 2006 [71] |
|
|
|
| Occipital cortex 13.5 mL (30 × 30 × 15 mm3) 20.5 mL (35 × 15 × 39 mm3) |
| NA |
| Nicotine | Epperson et al., 2005 [72] |
|
|
|
| Occipital cortex 13.5 mL (15 × 30 × 30 mm3) |
| NA |
| Nicotine | Baggaa et al., 2018 [73] |
|
|
|
| PFC 27 mL (30 × 30 × 30 mm3) |
| NA |
| Nicotine | Janes et al., 2013 [74] |
|
|
|
| dACC 12 mL (20 × 20 × 30 mm3) |
|
|
| Cocaine | Ke et al., 2004 [75] |
|
|
|
| Left PFC 18.75 mL |
| NA |
| Cocaine | Hulka et al., 2016 [76] |
|
|
|
| pgACC 9 mL (25 × 18 × 20 mm3) Right DLPFC 9 mL (25 × 18 × 20 mm3) |
|
|
| Cocaine | Ersche et al., 2021 [77] |
|
|
|
| Left Putamen 9 mL (16 × 16 × 35 mm3) |
| NA |
| Methamphetamine | Su et al., 2020 [78] |
|
|
|
| Left DLPFC 8 mL (20 × 20 × 20 mm3) |
|
|
| Cannabis | Prescot et al., 2013 [20] |
|
|
|
| ACC 15 mL (30 × 25 × 20 mm3) |
| NA |
| Rx opioids | Li et al., 2020 [79] |
|
|
|
| Medial PFC 12 mL (23 × 23 × 23 mm3) |
|
|
| opioids, alcohol | Murray et al., 2016 [80] |
|
|
|
| ACC 17.5 mL (35 × 25 × 20 mm3) Right DLPFC 16 mL (20 × 40 × 20 mm3) OFC 8 mL (40 × 20 × 10 mm3) POC 16 mL (40 × 20 × 20 mm3) |
| NA |
| Polysubstance | Schulte et al., 2017 [81] |
|
|
|
| dACC 10.5 mL (35 × 20 × 15 mm3) |
| NA |
| Polysubstance | Abe et al., 2013 [82] |
|
|
|
| ACC 17.5 mL (35 × 25 × 20 mm3) POC 16 mL (20 × 40 × 20 mm3) right DLPFC 16 mL (40 × 20 × 20 mm3) |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shyu, C.; Chavez, S.; Boileau, I.; Le Foll, B. Quantifying GABA in Addiction: A Review of Proton Magnetic Resonance Spectroscopy Studies. Brain Sci. 2022, 12, 918. https://doi.org/10.3390/brainsci12070918
Shyu C, Chavez S, Boileau I, Le Foll B. Quantifying GABA in Addiction: A Review of Proton Magnetic Resonance Spectroscopy Studies. Brain Sciences. 2022; 12(7):918. https://doi.org/10.3390/brainsci12070918
Chicago/Turabian StyleShyu, Claire, Sofia Chavez, Isabelle Boileau, and Bernard Le Foll. 2022. "Quantifying GABA in Addiction: A Review of Proton Magnetic Resonance Spectroscopy Studies" Brain Sciences 12, no. 7: 918. https://doi.org/10.3390/brainsci12070918