
Review of the Midbrain Ascending Arousal Network Nuclei and Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS), Gulf War Illness (GWI) and Postexertional Malaise (PEM)


Abstract
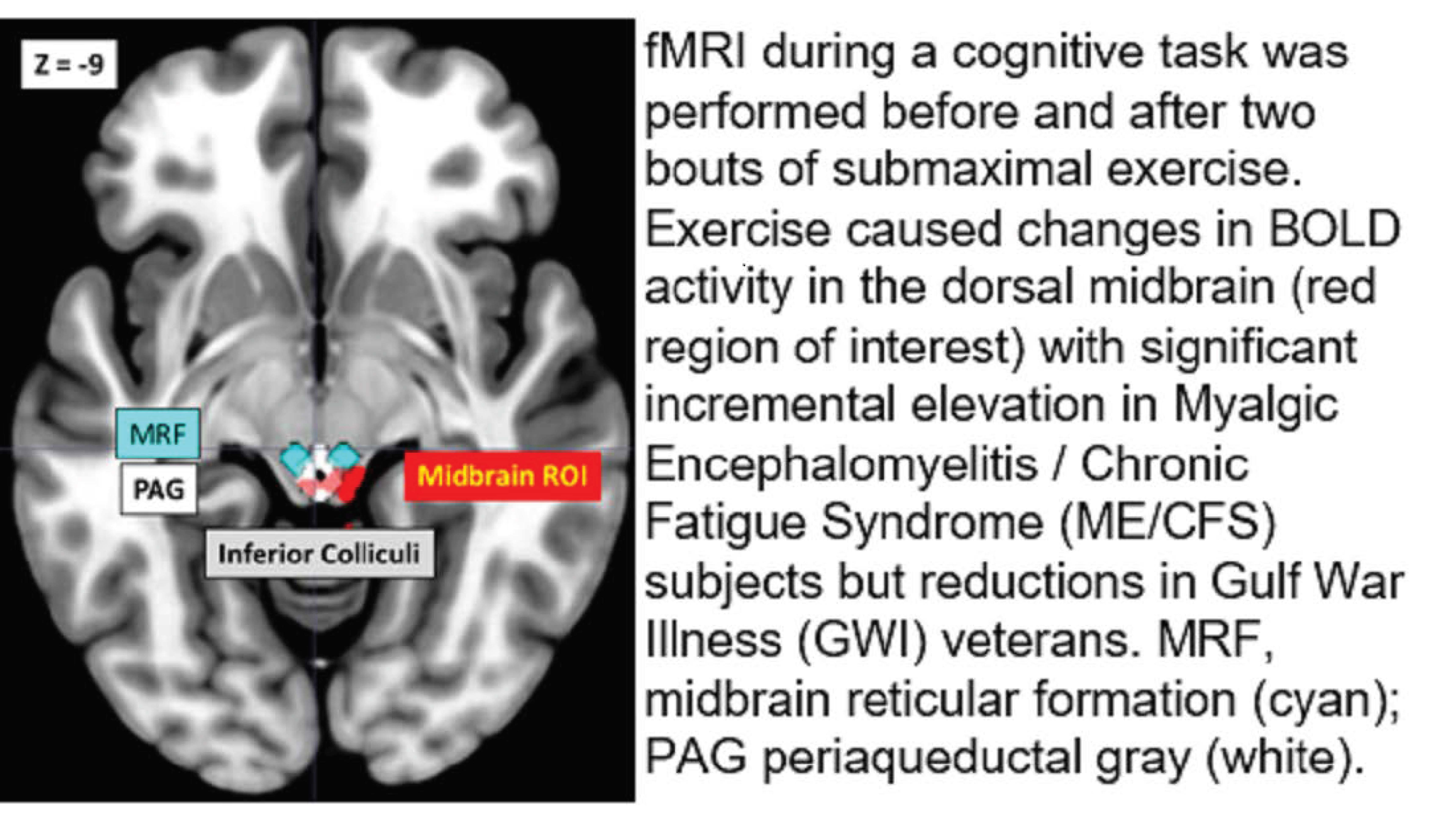
:
1. Introduction
2. Midbrain Nuclei
3. Development of the Midbrain and Isthmus
4. Periaqueductal Gray
5. Inferior Colliculus (IC)
6. Oculomotor Nerve Nuclei
7. Midbrain Reticular Formation (MRF)
8. Nuclei with Amines as Predominant Neurotransmitters
9. Locus Coeruleus (LC), Norepinephrine and Sympathetic Activation
10. Dorsal Raphe (DR), Median Raphe (MR) and Serotonin (5HT)
11. Ventral Tegmental Area (VTA) and Dopamine
12. Pedunculotegmental Nuclei (PTN), Pontis Oralis (PO), and Acetylcholine
13. Parabrachial Complex (PBC)
14. Dysfunction in ME/CFS and GWI
15. Limitations
16. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Fukuda, K.; Straus, S.E.; Hickie, I.; Sharpe, M.C.; Dobbins, J.G.; Komaroff, A.; International Chronic Fatigue Syndrome Study Group. The Chronic Fatigue Syndrome: A Comprehensive Approach to Its Definition and Study. Ann. Intern. Med. 1994, 121, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, B.M.; Jain, A.K.; De Meirleir, K.L.; Peterson, D.L.; Klimas, N.G.; Lerner, A.M.; Bested, A.C.; Flor-Henry, P.; Joshi, P.; Powles, A.C.P.; et al. Myalgic encephalomyelitis/chronic fatigue syndrome. J. Chronic Fatigue Syndr. 2003, 11, 7–115. [Google Scholar] [CrossRef]
- Fukuda, K.; Nisenbaum, R.; Stewart, G.; Thompson, W.W.; Robin, L.; Washko, R.M.; Noah, D.L.; Barrett, D.H.; Randall, B.; Herwaldt, B.L.; et al. Chronic Multisymptom Illness Affecting Air Force Veterans of the Gulf War. JAMA J. Am. Med. Assoc. 1998, 280, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Steele, L. Prevalence and Patterns of Gulf War Illness in Kansas Veterans: Association of Symptoms with Characteristics of Person, Place, and Time of Military Service. Am. J. Epidemiol. 2000, 152, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Daugherty, S.A.; Henry, B.E.; Peterson, D.L.; Swarts, R.L.; Bastien, S.; Thomas, R.S. Chronic Fatigue Syndrome in Northern Nevada. Clin. Infect. Dis. 1991, 13, S39–S44. [Google Scholar] [CrossRef] [PubMed]
- Friedberg, F.; Bateman, L.; Bested, A.; Davenport, T.; Friedman, K.; Gurwitt, A.; Jason, L.; Lappp, C.; Stevens, S.; Underhill, R.V.R. International Association for Chronic Fatigue Syndrome/Myalgic Encephalomyelitis Me/CFS: Primer for Clinical Practitioners. 2014, pp. 1–541. Available online: https://www.iacfsme.org/resources-for-healthcare-professionals/ (accessed on 1 January 2020).
- Lim, E.-J.; Ahn, Y.-C.; Jang, E.-S.; Lee, S.-W.; Lee, S.-H.; Son, C.-G. Systematic review and meta-analysis of the prevalence of chronic fatigue syndrome/myalgic encephalomyelitis (CFS/ME). J. Transl. Med. 2020, 18, 100. [Google Scholar] [CrossRef]
- Carruthers, B.M. Definitions and aetiology of myalgic encephalomyelitis: How the Canadian consensus clinical definition of myalgic encephalomyelitis works. J. Clin. Pathol. 2006, 60, 117–119. [Google Scholar] [CrossRef] [Green Version]
- Carruthers, B.M.; Van De Sande, M.I.; De Meirleir, K.L.; Klimas, N.G.; Broderick, G.; Mitchell, T.; Staines, D.; Powles, A.C.P.; Speight, N.; Vallings, R.; et al. Myalgic encephalomyelitis: International Consensus Criteria. J. Intern. Med. 2011, 270, 327–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Institute of Medicine. Beyond Myalgic Encephalomyelitis/Chronic Fatigue Syndrome; The National Academies Press: Washington, DC, USA, 2015. [Google Scholar]
- National Institute for Health Care Excellence (NICE). Myalgic Encephalomyelitis (or Encephalopathy)/Chronic Fatigue Syndrome: Diagnosis and Management. NICE Guideline [NG206]. 2021, pp. 1–87. Available online: www.nice.org.uk/guidance/ng206 (accessed on 1 November 2021).
- Bateman, L.; Bested, A.C.; Bonilla, H.F.; Chheda, B.V.; Chu, L.; Curtin, J.M.; Dempsey, T.T.; Dimmock, M.E.; Dowell, T.G.; Felsenstein, D.; et al. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: Essentials of Diagnosis and Management. Mayo Clin. Proc. 2021, 96, 2861–2878. [Google Scholar] [CrossRef]
- Rowe, P.C.; Underhill, R.A.; Friedman, K.J.; Gurwitt, A.; Medow, M.S.; Schwartz, M.S.; Speight, N.; Stewart, J.M.; Vallings, R.; Rowe, K.S. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Diagnosis and Management in Young People: A Primer. Front. Pediatr. 2017, 5, 121. [Google Scholar] [CrossRef] [Green Version]
- White, R.F.; Steele, L.; O’Callaghan, J.P.; Sullivan, K.; Binns, J.H.; Golomb, B.A.; Bloom, F.E.; Bunker, J.A.; Crawford, F.; Graves, J.C.; et al. Recent research on Gulf War illness and other health problems in veterans of the 1991 Gulf War: Effects of toxicant exposures during deployment. Cortex 2016, 74, 449–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, L.; Sastre, A.; Gerkovich, M.M.; Cook, M.R. Complex Factors in the Etiology of Gulf War Illness: Wartime Exposures and Risk Factors in Veteran Subgroups. Environ. Health Perspect. 2012, 120, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.K. Organophosphorus esters causing delayed neurotoxic effects: Mechanism of action and structure activity studies. Arch. Toxicol. 1975, 34, 259–288. [Google Scholar] [CrossRef] [PubMed]
- Namba, T.; Nolte, C.T.; Jackrel, J.; Grob, D. Poisoning due to organophosphate insecticides: Acute and chronic manifestations. Am. J. Med. 1971, 50, 475–492. [Google Scholar] [CrossRef]
- Richardson, R.J.; Fink, J.K.; Glynn, P.; Hufnagel, R.B.; Makhaeva, G.F.; Wijeyesakere, S.J. Neuropathy target esterase (NTE/PNPLA6) and organophosphorus compound-induced delayed neurotoxicity (OPIDN). Neurotox. Met. Old Issues New Dev. 2020, 4, 1–78. [Google Scholar] [CrossRef]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.-A.; Goldenberg, D.L.; Katz, R.S.; Mease, P.; Russell, A.S.; Russell, I.J.; Winfield, J.B.; Yunus, M.B. The American College of Rheumatology Preliminary Diagnostic Criteria for Fibromyalgia and Measurement of Symptom Severity. Arthritis Care Res. 2010, 62, 600–610. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.-A.; Goldenberg, D.L.; Häuser, W.; Katz, R.S.; Mease, P.; Russell, A.S.; Russell, I.J.; Winfield, J.B. Fibromyalgia Criteria and Severity Scales for Clinical and Epidemiological Studies: A Modification of the ACR Preliminary Diagnostic Criteria for Fibromyalgia. J. Rheumatol. 2011, 38, 1113–1122. [Google Scholar] [CrossRef]
- Wolfe, F.; Clauw, D.J.; Fitzcharles, M.-A.; Goldenberg, D.L.; Häuser, W.; Katz, R.L.; Mease, P.J.; Russell, A.S.; Russell, I.J.; Walitt, B. 2016 Revisions to the 2010/2011 fibromyalgia diagnostic criteria. Semin. Arthritis Rheum. 2016, 46, 319–329. [Google Scholar] [CrossRef]
- Baraniuk, J.N. Chronic fatigue syndrome prevalence is grossly overestimated using Oxford criteria compared to Centers for Disease Control (Fukuda) criteria in a U.S. population study. Fatigue Biomed. Health Behav. 2017, 5, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Rayhan, R.U.; Baraniuk, J.N. Submaximal Exercise Provokes Increased Activation of the Anterior Default Mode Network During the Resting State as a Biomarker of Postexertional Malaise in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Neurosci. 2021, 15, 748426. [Google Scholar] [CrossRef] [PubMed]
- Rayhan, R.U.; Washington, S.D.; Garner, R.; Zajur, K.; Addiego, F.M.; VanMeter, J.W.; Baraniuk, J.N. Exercise challenge alters Default Mode Network dynamics in Gulf War Illness. BMC Neurosci. 2019, 20, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washington, S.D.; Rayhan, R.U.; Garner, R.; Provenzano, D.; Zajur, K.; Addiego, F.M.; VanMeter, J.W.; Baraniuk, J.N. Exercise alters brain activation in Gulf War Illness and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Brain Commun. 2020, 2, fcaa070. [Google Scholar] [CrossRef] [PubMed]
- Edlow, B.L.; Takahashi, E.; Wu, O.; Benner, T.; Dai, G.; Bu, L.; Grant, P.E.; Greer, D.; Greenberg, S.M.; Kinney, H.C.; et al. Neuroanatomic Connectivity of the Human Ascending Arousal System Critical to Consciousness and Its Disorders. J. Neuropathol. Exp. Neurol. 2012, 71, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Horn, A.; Li, N.; Dembek, T.A.; Kappel, A.; Boulay, C.; Ewert, S.; Tietze, A.; Husch, A.; Perera, T.; Neumann, W.-J.; et al. Lead-DBS v2: Towards a comprehensive pipeline for deep brain stimulation imaging. NeuroImage 2019, 184, 293–316. [Google Scholar] [CrossRef] [PubMed]
- Rayhan, R.U.; Zheng, Y.; Uddin, E.; Timbol, C.; Adewuyi, O.; Baraniuk, J.N. Administer and collect medical questionnaires with Google documents: A simple, safe, and free system. Appl. Med. Inform. 2013, 33, 12–21. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24415903 (accessed on 1 October 2013). [PubMed]
- Garner, R.; Baraniuk, J.N. Orthostatic intolerance in chronic fatigue syndrome. J. Transl. Med. 2019, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Terem, I.; Dang, L.; Champagne, A.; Abderezaei, J.; Pionteck, A.; Almadan, Z.; Lydon, A.; Kurt, M.; Scadeng, M.; Holdsworth, S.J. 3D amplified MRI (aMRI). Magn. Reson. Med. 2021, 86, 1674–1686. [Google Scholar] [CrossRef] [PubMed]
- VanElzakker, M.B.; Brumfield, S.A.; Mejia, P.S.L. Neuroinflammation and Cytokines in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): A Critical Review of Research Methods. Front. Neurol. 2018, 9, 1033. [Google Scholar] [CrossRef]
- Zhang, Y.; Avery, T.; Vakhtin, A.A.; Mathersul, D.; Tranvinh, E.; Wintermark, M.; Massaband, P.; Ashford, J.W.; Bayley, P.J.; Furst, A.J. Brainstem atrophy in Gulf War Illness. NeuroToxicology 2020, 78, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Christova, P.; James, L.M.; Engdahl, B.E.; Lewis, S.M.; Carpenter, A.F.; Georgopoulos, A.P. Subcortical brain atrophy in Gulf War Illness. Exp. Brain Res. 2017, 235, 2777–2786. [Google Scholar] [CrossRef]
- Barnden, L.R.; Shan, Z.Y.; Staines, D.R.; Marshall-Gradisnik, S.; Finegan, K.; Ireland, T.; Bhuta, S. Hyperintense sensorimotor T1 spin echo MRI is associated with brainstem abnormality in chronic fatigue syndrome. NeuroImage Clin. 2018, 20, 102–109. [Google Scholar] [CrossRef]
- Barnden, L.R.; Crouch, B.; Kwiatek, R.; Burnet, R.; Mernone, A.; Chryssidis, S.; Scroop, G.; Del Fante, P. A brain MRI study of chronic fatigue syndrome: Evidence of brainstem dysfunction and altered homeostasis. NMR Biomed. 2011, 24, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Barnden, L.R.; Shan, Z.Y.; Staines, D.R.; Marshall-Gradisnik, S.; Finegan, K.; Ireland, T.; Bhuta, S. Intra brainstem connectivity is impaired in chronic fatigue syndrome. NeuroImage Clin. 2019, 24, 102045. [Google Scholar] [CrossRef] [PubMed]
- Athinoula, A. Martinos Center for Biomedical Imaging. Harvard Ascending Arousal Network Atlas. Available online: https://www.nmr.mgh.harvard.edu/resources/aan-atlas (accessed on 1 January 2018).
- Paxinos, G.; Furlong, T.; Watson, C. Human Brainstem: Cytoarchitecture, Chemoarchitecture, Myeloarchitecture; Revised; Academic Press Inc.: London, UK, 2020. [Google Scholar]
- Shan, Z.Y.; Barnden, L.R.; Kwiatek, R.A.; Bhuta, S.; Hermens, D.F.; Lagopoulos, J. Neuroimaging characteristics of myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS): A systematic review. J. Transl. Med. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- James, L.M.; Christova, P.; Engdahl, B.E.; Lewis, S.M.; Carpenter, A.F.; Georgopoulos, A.P. Human Leukocyte Antigen (HLA) and Gulf War Illness (GWI): HLA-DRB1*13:02 Spares Subcortical Atrophy in Gulf War Veterans. EBioMedicine 2017, 26, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, L.L.; Zhang, Y.; Buckley, S. Effects of low-level sarin and cyclosarin exposure on white matter integrity in Gulf War Veterans. NeuroToxicology 2015, 48, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracha, H.S. Human brain evolution and the “Neuroevolutionary Time-depth Principle:” Implications for the Reclassification of fear-circuitry-related traits in DSM-V and for studying resilience to warzone-related posttraumatic stress disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2006, 30, 827–853. [Google Scholar] [CrossRef]
- Puelles, L. Comments on the limits and internal structure of the mammalian midbrain. Anatomy 2016, 10, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Puelles, L. Survey of Midbrain, Diencephalon, and Hypothalamus Neuroanatomic Terms Whose Prosomeric Definition Conflicts with Columnar Tradition. Front. Neuroanat. 2019, 13, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, C.; Bartholomaeus, C.; Puelles, L. Time for Radical Changes in Brain Stem Nomenclature—Applying the Lessons From Developmental Gene Patterns. Front. Neuroanat. 2019, 13, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo-Sánchez, M.; Martínez-De-La-Torre, M.; Alvarado-Mallart, R.-M.; Puelles, L. A distinct preisthmic histogenetic domain is defined by overlap ofOtx2andPax2gene expression in the avian caudal midbrain. J. Comp. Neurol. 2005, 483, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Sato, T.; Nakamura, H. Fgf8 signaling for development of the midbrain and hindbrain. Dev. Growth Differ. 2016, 58, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Franklin, T.B. Recent Advancements Surrounding the Role of the Periaqueductal Gray in Predators and Prey. Front. Behav. Neurosci. 2019, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Linnman, C.; Moulton, E.; Barmettler, G.; Becerra, L.; Borsook, D. Neuroimaging of the periaqueductal gray: State of the field. NeuroImage 2012, 60, 505–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satpute, A.B.; Wager, T.D.; Cohen-Adad, J.; Bianciardi, M.; Choi, J.-K.; Buhle, J.T.; Wald, L.; Barrett, L.F. Identification of discrete functional subregions of the human periaqueductal gray. Proc. Natl. Acad. Sci. USA 2013, 110, 17101–17106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterthwaite, T.D.; Ciric, R.; Roalf, D.R.; Davatzikos, C.; Bassett, D.S.; Wolf, D. Motion artifact in studies of functional connectivity: Characteristics and mitigation strategies. Hum. Brain Mapp. 2019, 40, 2033–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkes, L.; Fulcher, B.; Yücel, M.; Fornito, A. An evaluation of the efficacy, reliability, and sensitivity of motion correction strategies for resting-state functional MRI. NeuroImage 2018, 171, 415–436. [Google Scholar] [CrossRef] [PubMed]
- Washington, S.D.; Rayhan, R.U.; Garner, R.; Provenzano, D.; Zajur, K.; Addiego, F.M.; VanMeter, J.W.; Baraniuk, J.N. Exercise alters cerebellar and cortical activity related to working memory in phenotypes of Gulf War Illness. Brain Commun. 2020, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rayhan, R.U.; Stevens, B.W.; Raksit, M.P.; Ripple, J.A.; Timbol, C.R.; Adewuyi, O.; VanMeter, J.W.; Baraniuk, J.N. Exercise Challenge in Gulf War Illness Reveals Two Subgroups with Altered Brain Structure and Function. PLoS ONE 2013, 8, e63903. [Google Scholar] [CrossRef]
- Garner, R.S.; Rayhan, R.U.; Baraniuk, J.N. Verification of exercise-induced transient postural tachycardia phenotype in Gulf War Illness. Am. J. Transl. Res. 2018, 10, 3254–3264. Available online: http://www.ncbi.nlm.nih.gov/pubmed/30416666 (accessed on 1 October 2018).
- Coulombe, M.-A.; Erpelding, N.; Kucyi, A.; Davis, K.D. Intrinsic functional connectivity of periaqueductal gray subregions in humans. Hum. Brain Mapp. 2016, 37, 1514–1530. [Google Scholar] [CrossRef] [PubMed]
- Faull, O.K.; Pattinson, K.T. The cortical connectivity of the periaqueductal gray and the conditioned response to the threat of breathlessness. eLife 2017, 6, e21749. [Google Scholar] [CrossRef] [PubMed]
- Canteras, N.S. The medial hypothalamic defensive system: Hodological organization and functional implications. Pharmacol. Biochem. Behav. 2002, 71, 481–491. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Griebel, G.; Pobbe, R.; Blanchard, R.J. Risk assessment as an evolved threat detection and analysis process. Neurosci. Biobehav. Rev. 2011, 35, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C. Translating dynamic defense patterns from rodents to people. Neurosci. Biobehav. Rev. 2017, 76, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Petrovic, P.; Marchant, J.L.; Hassabis, D.; Weiskopf, N.; Seymour, B.; Dolan, R.J.; Frith, C.D. When Fear Is Near: Threat Imminence Elicits Prefrontal-Periaqueductal Gray Shifts in Humans. Science 2007, 317, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- Buhle, J.T.; Kober, H.; Ochsner, K.N.; Mende-Siedlecki, P.; Weber, J.; Hughes, B.L.; Kross, E.; Atlas, L.Y.; McRae, K.; Wager, T.D. Common representation of pain and negative emotion in the midbrain periaqueductal gray. Soc. Cogn. Affect. Neurosci. 2013, 8, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Kober, H.; Barrett, L.F.; Joseph, J.; Bliss-Moreau, E.; Lindquist, K.; Wager, T.D. Functional grouping and cortical–subcortical interactions in emotion: A meta-analysis of neuroimaging studies. NeuroImage 2008, 42, 998–1031. [Google Scholar] [CrossRef] [Green Version]
- Wager, T.D.; Atlas, L.Y.; Lindquist, M.A.; Roy, M.; Woo, C.-W.; Kross, E. An fMRI-Based Neurologic Signature of Physical Pain. N. Engl. J. Med. 2013, 368, 1388–1397. [Google Scholar] [CrossRef] [Green Version]
- Tracey, I.; Ploghaus, A.; Gati, J.S.; Clare, S.; Smith, S.; Menon, R.; Matthews, P.M. Imaging Attentional Modulation of Pain in the Periaqueductal Gray in Humans. J. Neurosci. 2002, 22, 2748–2752. [Google Scholar] [CrossRef] [PubMed]
- Kinomura, S.; Larsson, J.; Gulyás, B.; Roland, P.E. Activation by Attention of the Human Reticular Formation and Thalamic Intralaminar Nuclei. Science 1996, 271, 512–515. [Google Scholar] [CrossRef]
- Bracha, H.S. Freeze, Flight, Fight, Fright, Faint: Adaptationist Perspectives on the Acute Stress Response Spectrum. CNS Spectr. 2004, 9, 679–685. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Gladwin, T.E.; De Valk, N.M.; Zhang, W.; Kaldewaij, R.; Van Ast, V.; Koch, S.B.J.; Klumpers, F.; Roelofs, K. Neural Dynamics of Shooting Decisions and the Switch from Freeze to Fight. Sci. Rep. 2019, 9, 4240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovote, P.; Esposito, M.S.; Botta, P.; Chaudun, F.; Fadok, J.P.; Markovic, M.; Wolff, S.B.E.; Ramakrishnan, C.; Fenno, L.; Deisseroth, K.; et al. Midbrain circuits for defensive behaviour. Nature 2016, 534, 206–212. [Google Scholar] [CrossRef]
- Kozlowska, K.; Walker, P.; McLean, L.; Carrive, P. Fear and the Defense Cascade. Harv. Rev. Psychiatry 2015, 23, 263–287. [Google Scholar] [CrossRef] [PubMed]
- La Cesa, S.; Tinelli, E.; Toschi, N.; Di Stefano, G.; Collorone, S.; Aceti, A.; Francia, A.; Cruccu, G.; Truini, A.; Caramia, F. fMRI pain activation in the periaqueductal gray in healthy volunteers during the cold pressor test. Magn. Reson. Imaging 2014, 32, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Naegeli, C.; Zeffiro, T.; Piccirelli, M.; Jaillard, A.; Weilenmann, A.; Hassanpour, K.; Schick, M.; Rufer, M.; Orr, S.P.; Mueller-Pfeiffer, C. Locus Coeruleus Activity Mediates Hyperresponsiveness in Posttraumatic Stress Disorder. Biol. Psychiatry 2018, 83, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Klein, L.C.; Lewis, B.P.; Gruenewald, T.L.; Gurung, R.A.R.; Updegraff, J.A. Biobehavioral responses to stress in females: Tend-and-befriend, not fight-or-flight. Psychol. Rev. 2000, 107, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Kuchenbecker, S.Y.; Pressman, S.D.; Celniker, J.; Grewen, K.M.; Sumida, K.D.; Jonathan, N.; Everett, B.; Slavich, G.M. Oxytocin, cortisol, and cognitive control during acute and naturalistic stress. Stress 2021, 24, 370–383. [Google Scholar] [CrossRef]
- Harricharan, S.; Rabellino, D.; Frewen, P.A.; Densmore, M.; Theberge, J.; McKinnon, M.C.; Schore, A.N.; Lanius, R.A. fMRI functional connectivity of the periaqueductal gray in PTSD and its dissociative subtype. Brain Behav. 2016, 6, e00579. [Google Scholar] [CrossRef]
- Lange, I.; Kasanova, Z.; Goossens, L.; Leibold, N.; De Zeeuw, C.I.; van Amelsvoort, T.; Schruers, K. The anatomy of fear learning in the cerebellum: A systematic meta-analysis. Neurosci. Biobehav. Rev. 2015, 59, 83–91. [Google Scholar] [CrossRef]
- Yoshiuchi, K.; Farkas, J.; Natelson, B.H. Patients with chronic fatigue syndrome have reduced absolute cortical blood flow. Clin. Physiol. Funct. Imaging 2006, 26, 83–86. [Google Scholar] [CrossRef]
- Van Campen, C.M.; Verheugt, F.W.; Rowe, P.C.; Visser, F.C. Cerebral blood flow is reduced in ME/CFS during head-up tilt testing even in the absence of hypotension or tachycardia: A quantitative, controlled study using Doppler echography. Clin. Neurophysiol. Pract. 2020, 5, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Finkelmeyer, A.; He, J.; MacLachlan, L.; Blamire, A.; Newton, J.L. Intracranial compliance is associated with symptoms of orthostatic intolerance in chronic fatigue syndrome. PLoS ONE 2018, 13, e0200068. [Google Scholar] [CrossRef] [PubMed]
- Falvo, M.; Lindheimer, J.B.; Serrador, J.M. Dynamic cerebral autoregulation is impaired in Veterans with Gulf War Illness: A case-control study. PLoS ONE 2018, 13, e0205393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Aslan, S.; Li, X.; Buhner, D.M.; Spence, J.S.; Briggs, R.W.; Haley, R.; Lu, H. Perfusion deficit to cholinergic challenge in veterans with Gulf War Illness. NeuroToxicology 2011, 32, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, H.H.; Holstege, G. Periaqueductal Gray Control of Breathing. Adv. Exp. Med. Biol. 2010, 669, 353–358. [Google Scholar] [CrossRef]
- Holstege, G. The Periaqueductal Gray Controls Brainstem Emotional Motor Systems Including Respiration. Prog. Brain Res. 2014, 209, 379–405. [Google Scholar]
- Ravindran, M.K.; Adewuyi, O.; Zheng, Y.; Rayhan, R.U.; Le, U.; Timbol, C.R.; Merck, S.; Esteitie, R.; Cooney, M.; Read, C.; et al. Dyspnea in Chronic Fatigue Syndrome (CFS): Comparison of Two Prospective Cross-Sectional Studies. Glob. J. Health Sci. 2012, 5, 94–110. [Google Scholar] [CrossRef]
- Baraniuk, J.N.; Zheng, Y. Relationships among rhinitis, fibromyalgia, and chronic fatigue. Allergy Asthma Proc. 2010, 31, 169–178. [Google Scholar] [CrossRef]
- Heinricher, M.; Tavares, I.; Leith, J.; Lumb, B. Descending control of nociception: Specificity, recruitment and plasticity. Brain Res. Rev. 2009, 60, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loyd, D.R.; Murphy, A.Z. The Role of the Periaqueductal Gray in the Modulation of Pain in Males and Females: Are the Anatomy and Physiology Really that Different? Neural Plast. 2009, 2009, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, N.F.; Chaplain, M.A.J.; Skevington, S.M. The role of N-methyl-D-aspartate (NMDA) receptors in wind-up: A mathematical model. Math. Med. Biol. 1996, 13, 193–205. [Google Scholar] [CrossRef]
- Surian, A.A.; Baraniuk, J.N. Systemic Hyperalgesia in Females with Gulf War Illness, Chronic Fatigue Syndrome and Fibromyalgia. Sci. Rep. 2020, 10, 5751. [Google Scholar] [CrossRef] [Green Version]
- Naranch, K.; Park, Y.-J.; Repka-Ramirez, M.; Velarde, A.; Clauw, D.; Baraniuk, J. A Tender Sinus does not Always Mean Rhinosinusitis. Otolaryngol. Neck Surg. 2002, 127, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Sims-Williams, H.; Matthews, J.; Talbot, P.; Love-Jones, S.; Brooks, J.; Patel, N.K.; Pickering, A.E. Deep brain stimulation of the periaqueductal gray releases endogenous opioids in humans. NeuroImage 2017, 146, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Bordia, T.; Zahr, N.M. The Inferior Colliculus in Alcoholism and Beyond. Front. Syst. Neurosci. 2020, 14, 606345. [Google Scholar] [CrossRef]
- Latash, L.P. Orienting reaction, organizing for action, and emotional processes. Pavlov. J. Biol. Sci. 1990, 25, 123–131. [Google Scholar] [CrossRef]
- Brandao, M.; Melo, L.; Cardoso, S. Mechanisms of defense in the inferior colliculus. Behav. Brain Res. 1993, 58, 49–55. [Google Scholar] [CrossRef]
- Parham, K.; Willott, J.F. Effects of inferior colliculus lesions on the acoustic startle response. Behav. Neurosci. 1990, 104, 831–840. [Google Scholar] [CrossRef]
- Heeringa, A.N.; Van Dijk, P. The immediate effects of acoustic trauma on excitation and inhibition in the inferior colliculus: A Wiener-kernel analysis. Hear. Res. 2016, 331, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Orr, S.P.; Lasko, N.B.; Shalev, A.Y.; Pitman, R.K. Physiologic responses to loud tones in Vietnam veterans with posttraumatic stress disorder. J. Abnorm. Psychol. 1995, 104, 75–82. [Google Scholar] [CrossRef]
- Kevanishvili, Z.S. Sources of the Human Brainstem Auditory Evoked Potential. Scand. Audiol. 1980, 9, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Fluge, Ø.; Mella, O.; Bruland, O.; Risa, K.; Dyrstad, S.E.; Alme, K.; Rekeland, I.G.; Sapkota, D.; Røsland, G.V.; Fosså, A.; et al. Metabolic profiling indicates impaired pyruvate dehydrogenase function in myalgic encephalopathy/chronic fatigue syndrome. JCI Insight 2016, 1, e89376. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.-C.; Chanraud, S.; Sullivan, E.V. Neuroimaging of Wernicke’s Encephalopathy and Korsakoff’s Syndrome. Neuropsychol. Rev. 2012, 22, 170–180. [Google Scholar] [CrossRef]
- Neary, J.P.; Roberts, A.D.W.; Leavins, N.; Harrison, M.F.; Croll, J.C.; Sexsmith, J.R. Prefrontal cortex oxygenation during incremental exercise in chronic fatigue syndrome. Clin. Physiol. Funct. Imaging 2008, 28, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.L.; Paterson, K.B.; McGowan, V.; Hutchinson, C.V. Visual Aspects of Reading Performance in Myalgic Encephalomyelitis (ME). Front. Psychol. 2018, 9, 1468. [Google Scholar] [CrossRef] [Green Version]
- Godts, D.; Moorkens, G.; Mathysen, D.G. Binocular Vision in Chronic Fatigue Syndrome. Am. Orthopt. J. 2016, 66, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Barnden, L.R.; Kwiatek, R.; Crouch, B.; Burnet, R.; Del Fante, P. Autonomic correlations with MRI are abnormal in the brainstem vasomotor centre in Chronic Fatigue Syndrome. NeuroImage Clin. 2016, 11, 530–537. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.S.; McCall, J.G.; Charney, D.S.; Murrough, J.W. The role of the locus coeruleus in the generation of pathological anxiety. Brain Neurosci. Adv. 2020, 4, 2398212820930321. [Google Scholar] [CrossRef]
- Samuels, E.; Szabadi, E.R.S.A.E. Functional Neuroanatomy of the Noradrenergic Locus Coeruleus: Its Roles in the Regulation of Arousal and Autonomic Function Part II: Physiological and Pharmacological Manipulations and Pathological Alterations of Locus Coeruleus Activity in Humans. Curr. Neuropharmacol. 2008, 6, 254–285. [Google Scholar] [CrossRef] [Green Version]
- De Cicco, V.; Fantozzi, M.P.T.; Cataldo, E.; Barresi, M.; Bruschini, L.; Faraguna, U.; Manzoni, D. Trigeminal, Visceral and Vestibular Inputs May Improve Cognitive Functions by Acting through the Locus Coeruleus and the Ascending Reticular Activating System: A New Hypothesis. Front. Neuroanat. 2017, 11, 130. [Google Scholar] [CrossRef]
- Bracha, H.S. Postmortem Locus Coeruleus Neuron Count in Three American Veterans with Probable or Possible War-Related PTSD. J. Neuropsychiatr. 2005, 17, 503–509. [Google Scholar] [CrossRef]
- Hökfelt, T.; Barde, S.; Xu, Z.-Q.D.; Kuteeva, E.; Rüegg, J.; Le Maitre, E.; Risling, M.; Kehr, J.; Ihnatko, R.; Theodorsson, E.; et al. Neuropeptide and Small Transmitter Coexistence: Fundamental Studies and Relevance to Mental Illness. Front. Neural Circuits 2018, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Porges, S.W. The polyvagal perspective. Biol. Psychol. 2007, 74, 116–143. [Google Scholar] [CrossRef]
- Porges, S.W. A phylogenetic journey through the vague and ambiguous Xth cranial nerve: A commentary on contemporary heart rate variability research. Biol. Psychol. 2007, 74, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, K.; Prickaerts, J.; Steinbusch, H. The dorsal raphe nucleus and serotonin: Implications for neuroplasticity linked to major depression and Alzheimer’s disease. Prog. Brain Res. 2008, 172, 233–264. [Google Scholar] [CrossRef]
- Paul, E.D.; Lowry, C.A. Functional topography of serotonergic systems supports the Deakin/Graeff hypothesis of anxiety and affective disorders. J. Psychopharmacol. 2013, 27, 1090–1106. [Google Scholar] [CrossRef]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef]
- Ranaldi, R. Dopamine and reward seeking: The role of ventral tegmental area. Rev. Neurosci. 2014, 25, 621–630. [Google Scholar] [CrossRef]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 8, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spindler, L.R.B.; Luppi, A.I.; Adapa, R.M.; Craig, M.M.; Coppola, P.; Peattie, A.R.D.; Manktelow, A.E.; Finoia, P.; Sahakian, B.J.; Williams, G.B.; et al. Dopaminergic brainstem disconnection is common to pharmacological and pathological consciousness perturbation. Proc. Natl. Acad. Sci. USA 2021, 118, e2026289118. [Google Scholar] [CrossRef] [PubMed]
- Edlow, B.L. Dopaminergic modulation of human consciousness via default mode network connectivity. Proc. Natl. Acad. Sci. USA 2021, 118, e2111268118. [Google Scholar] [CrossRef]
- Hassan, A.; Benarroch, E.E. Heterogeneity of the midbrain dopamine system: Implications for Parkinson disease. Neurology 2015, 85, 1795–1805. [Google Scholar] [CrossRef]
- Chao, L. Do Gulf War veterans with high levels of deployment-related exposures display symptoms suggestive of Parkinson’s disease? Int. J. Occup. Med. Environ. Health 2019, 32, 503–526. [Google Scholar] [CrossRef] [PubMed]
- Mena-Segovia, J.; Bolam, J.P. Rethinking the Pedunculopontine Nucleus: From Cellular Organization to Function. Neuron 2017, 94, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Xi, M.-C.; Morales, F.R.; Chase, M.H. Interactions between GABAergic and Cholinergic Processes in the Nucleus Pontis Oralis: Neuronal Mechanisms Controlling Active (Rapid Eye Movement) Sleep and Wakefulness. J. Neurosci. 2004, 24, 10670–10678. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rill, E.; Tackett, A.J.; Byrum, S.D.; Lan, R.S.; Mackintosh, S.G.; Hyde, J.R.; Bisagno, V.; Urbano, F.J. Local and Relayed Effects of Deep Brain Stimulation of the Pedunculopontine Nucleus. Brain Sci. 2019, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Herrera, V.; Trujillo-Ferrara, J.G.; Miranda-Páez, A.; De La Cruz, F.; Zamudio, S.R. Corticosterone microinjected into nucleus pontis oralis increases tonic immobility in rats. Horm. Behav. 2011, 60, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.J.; McGill, M.M.; Shehab, S.A.S. Neurokinin 1 receptor expression by neurons in laminae I, III and IV of the rat spinal dorsal horn that project to the brainstem. Eur. J. Neurosci. 2000, 12, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.C.; Bowen, A.; Schier, L.A.; Tupone, D.; Uddin, O.; Heinricher, M.M. Parabrachial Complex: A Hub for Pain and Aversion. J. Neurosci. 2019, 39, 8225–8230. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liu, R.; Guo, F.; Wen, M.-Q.; Ma, X.-L.; Li, K.-Y.; Sun, H.; Xu, C.-L.; Li, Y.-Y.; Wu, M.-Y.; et al. Parabrachial nucleus circuit governs neuropathic pain-like behavior. Nat. Commun. 2020, 11, 5974. [Google Scholar] [CrossRef]
- Kelley, K.W.; Bluthé, R.-M.; Dantzer, R.; Zhou, J.-H.; Shen, W.; Johnson, R.; Broussard, S.R. Cytokine-induced sickness behavior. Brain, Behav. Immun. 2003, 17, 112–118. [Google Scholar] [CrossRef]
- Morrison, S.; Nakamura, K. Central Mechanisms for Thermoregulation. Annu. Rev. Physiol. 2019, 81, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D. The Parabrachial Nucleus: CGRP Neurons Function as a General Alarm. Trends Neurosci. 2018, 41, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies—Successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Rayhan, R.R.; Ravindran, M.K.; Baraniuk, J.N. Migraine in gulf war illness and chronic fatigue syndrome: Prevalence, potential mechanisms, and evaluation. Front. Physiol. 2013, 4, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, M.K.; Zheng, Y.; Timbol, C.; Merck, S.J.; Baraniuk, J.N. Migraine headaches in Chronic Fatigue Syndrome (CFS): Comparison of two prospective cross-sectional studies. BMC Neurol. 2011, 11, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Montagna, P.; Pierangeli, G.; Cortelli, P. The Primary Headaches as a Reflection of Genetic Darwinian Adaptive Behavioral Responses. Headache 2010, 50, 273–289. [Google Scholar] [CrossRef]
- Mingam, R.; Moranis, A.; Bluthé, R.-M.; De Smedt-Peyrusse, V.; Kelley, K.W.; Guesnet, P.; Lavialle, M.; Dantzer, R.; Layé, S. Uncoupling of interleukin-6 from its signalling pathway by dietary n-3-polyunsaturated fatty acid deprivation alters sickness behaviour in mice. Eur. J. Neurosci. 2008, 28, 1877–1886. [Google Scholar] [CrossRef] [Green Version]
- Borsini, A.; Stangl, D.; Jeffries, A.R.; Pariante, C.M.; Thuret, S. The role of omega-3 fatty acids in preventing glucocorticoid-induced reduction in human hippocampal neurogenesis and increase in apoptosis. Transl. Psychiatry 2020, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Borsini, A.; Nicolaou, A.; Camacho-Muñoz, D.; Kendall, A.C.; Di Benedetto, M.G.; Giacobbe, J.; Su, K.-P.; Pariante, C.M. Omega-3 polyunsaturated fatty acids protect against inflammation through production of LOX and CYP450 lipid mediators: Relevance for major depression and for human hippocampal neurogenesis. Mol. Psychiatry 2021, 26, 6773–6788. [Google Scholar] [CrossRef]
- Kalkman, H.; Hersberger, M.; Walitza, S.; Berger, G. Disentangling the Molecular Mechanisms of the Antidepressant Activity of Omega-3 Polyunsaturated Fatty Acid: A Comprehensive Review of the Literature. Int. J. Mol. Sci. 2021, 22, 4393. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-P.; Tseng, P.-T.; Lin, P.-Y.; Okubo, R.; Chen, T.-Y.; Chen, Y.-W.; Matsuoka, Y.J. Association of Use of Omega-3 Polyunsaturated Fatty Acids with Changes in Severity of Anxiety Symptoms: A Systematic Review and Meta-analysis. JAMA Netw. Open 2018, 1, e182327. [Google Scholar] [CrossRef]
- Kalinić, D.; Štefanović, L.B.; Jeroncic, A.; Mimica, N.; Dodig, G.; Delaš, I. Eicosapentaenoic acid in serum lipids could be inversely correlated with severity of clinical symptomatology in Croatian war veterans with posttraumatic stress disorder. Croat. Med. J. 2014, 55, 27–37. Available online: http://www.ncbi.nlm.nih.gov/pubmed/24577824 (accessed on 1 April 2020).
- Matsuoka, Y.J.; Hamazaki, K.; Nishi, D.; Hamazaki, T. Change in blood levels of eicosapentaenoic acid and posttraumatic stress symptom: A secondary analysis of data from a placebo-controlled trial of omega3 supplements. J. Affect. Disord. 2016, 205, 289–291. [Google Scholar] [CrossRef]
- Appleton, K.M.; Voyias, P.D.; Sallis, H.M.; Dawson, S.; Ness, A.R.; Churchill, R.; Perry, R. Omega-3 fatty acids for depression in adults. Cochrane Database Syst. Rev. 2021, 11, CD004692. [Google Scholar] [CrossRef]
- Castro-Marrero, J.; Zaragozá, M.C.; Domingo, J.C.; Martinez-Martinez, A.; Alegre, J.; von Schacky, C. Low omega-3 index and polyunsaturated fatty acid status in patients with chronic fatigue syndrome/myalgic encephalomyelitis. Prostaglandins Leukot. Essent. Fat. Acids 2018, 139, 20–24. [Google Scholar] [CrossRef]
- Marsland, A.L.; Kuan, D.C.-H.; Sheu, L.K.; Krajina, K.; Kraynak, T.; Manuck, S.B.; Gianaros, P.J. Systemic inflammation and resting state connectivity of the default mode network. Brain Behav. Immun. 2017, 62, 162–170. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FM 1990 | Oxford 1991 | CFS “Fukuda” 1994 | Chronic Multi-Symptom Illness (CMI) 1998 | GWI “Kansas” 2000 | Canadian Consensus Criteria ME/CFS 2003, 2007 | FM 2010, 2011, 2016 | SEID 2015 |
|---|---|---|---|---|---|---|---|
| Disability/ Reduced Quality of Life | Disability/Reduced Quality of Life | Disability/ Reduced Quality of Life | |||||
| Tenderness | |||||||
| Widespread Pain | Myalgia Arthralgia | Musculoskeletal pain | Pain | Widespread Pain | Widespread Pain | ||
| Fatigue | Fatigue | Fatigue | Fatigue | Fatigue | Fatigue | Fatigue | |
| Sleep | Sleep | Waking unrefreshed | Waking unrefreshed | Waking unrefreshed | |||
| Post-exertional malaise | Post-exertional malaise | Post-exertional malaise | Post-exertional malaise | ||||
| Cognition | Cognition | Cognition | Cognition | Cognition | Cognition | ||
| Mood | Mood | ||||||
| Interoceptive symptoms | |||||||
| Headache Sore throat Sore lymph nodes | Neurological Gastrointestinal Respiratory Skin | Neurological Autonomic Neuroendocrine Immune | Somatic symptoms | Orthostatic Intolerance | |||
| Key features for diagnosis | |||||||
| Pain + Tenderness | No exclusion * | Fatigue plus ≥4 of 8 | ≥1 chronic symptom in ≥2 categories | ≥3 of 6 categories | Severity scores | Moderate or severe >50% of time | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baraniuk, J.N. Review of the Midbrain Ascending Arousal Network Nuclei and Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS), Gulf War Illness (GWI) and Postexertional Malaise (PEM). Brain Sci. 2022, 12, 132. https://doi.org/10.3390/brainsci12020132
Baraniuk JN. Review of the Midbrain Ascending Arousal Network Nuclei and Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS), Gulf War Illness (GWI) and Postexertional Malaise (PEM). Brain Sciences. 2022; 12(2):132. https://doi.org/10.3390/brainsci12020132
Chicago/Turabian StyleBaraniuk, James N. 2022. "Review of the Midbrain Ascending Arousal Network Nuclei and Implications for Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS), Gulf War Illness (GWI) and Postexertional Malaise (PEM)" Brain Sciences 12, no. 2: 132. https://doi.org/10.3390/brainsci12020132