Identification of Copy Number Variation in Domestic Chicken Using Whole-Genome Sequencing Reveals Evidence of Selection in the Genome

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Chicken Whole-Genome Sequencing Data
2.2. Sequence Alignment to Reference Genome
2.3. CNV and CNVR Detection
2.4. CNVR Annotation
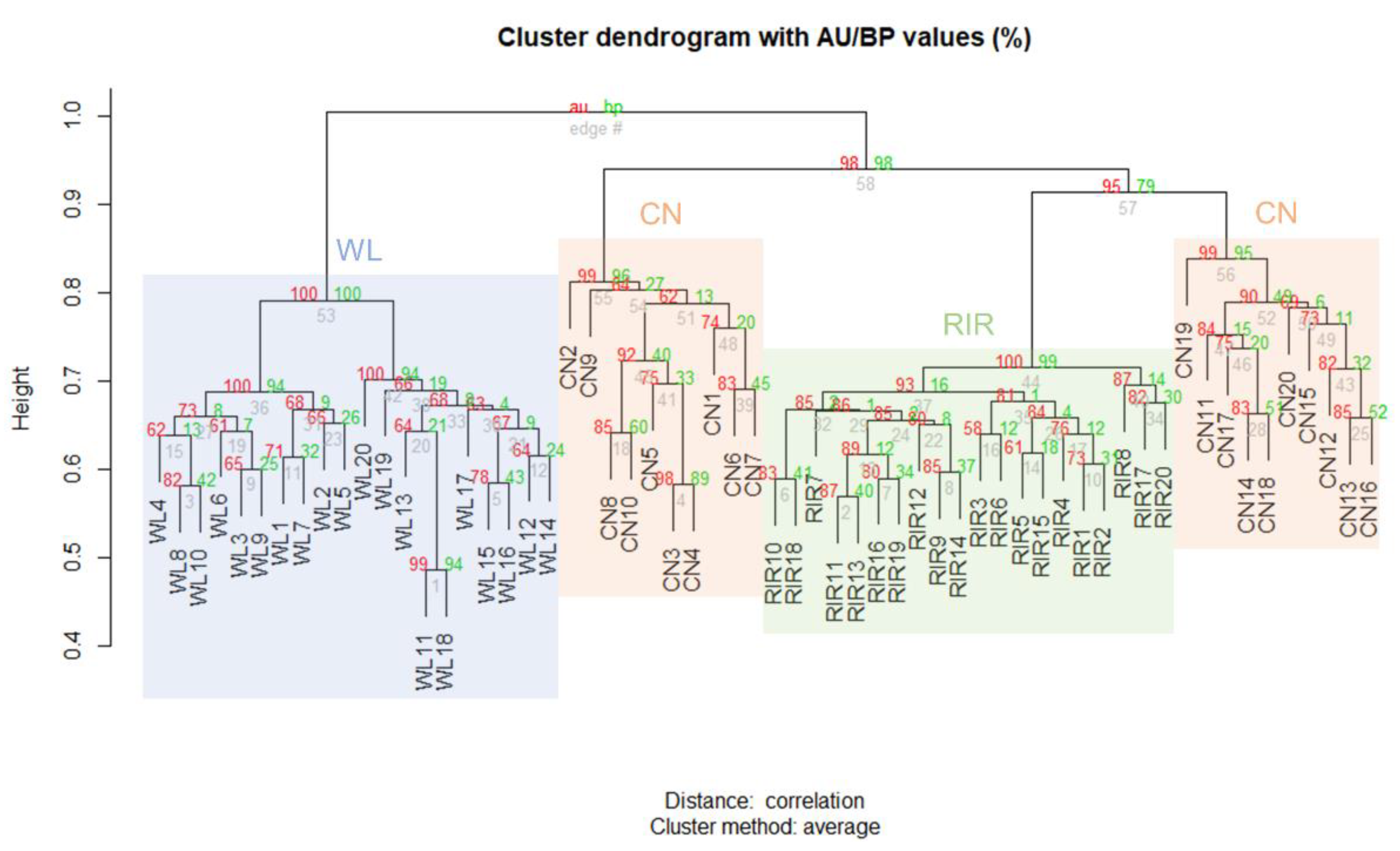
2.5. Breed Differentiation
3. Results
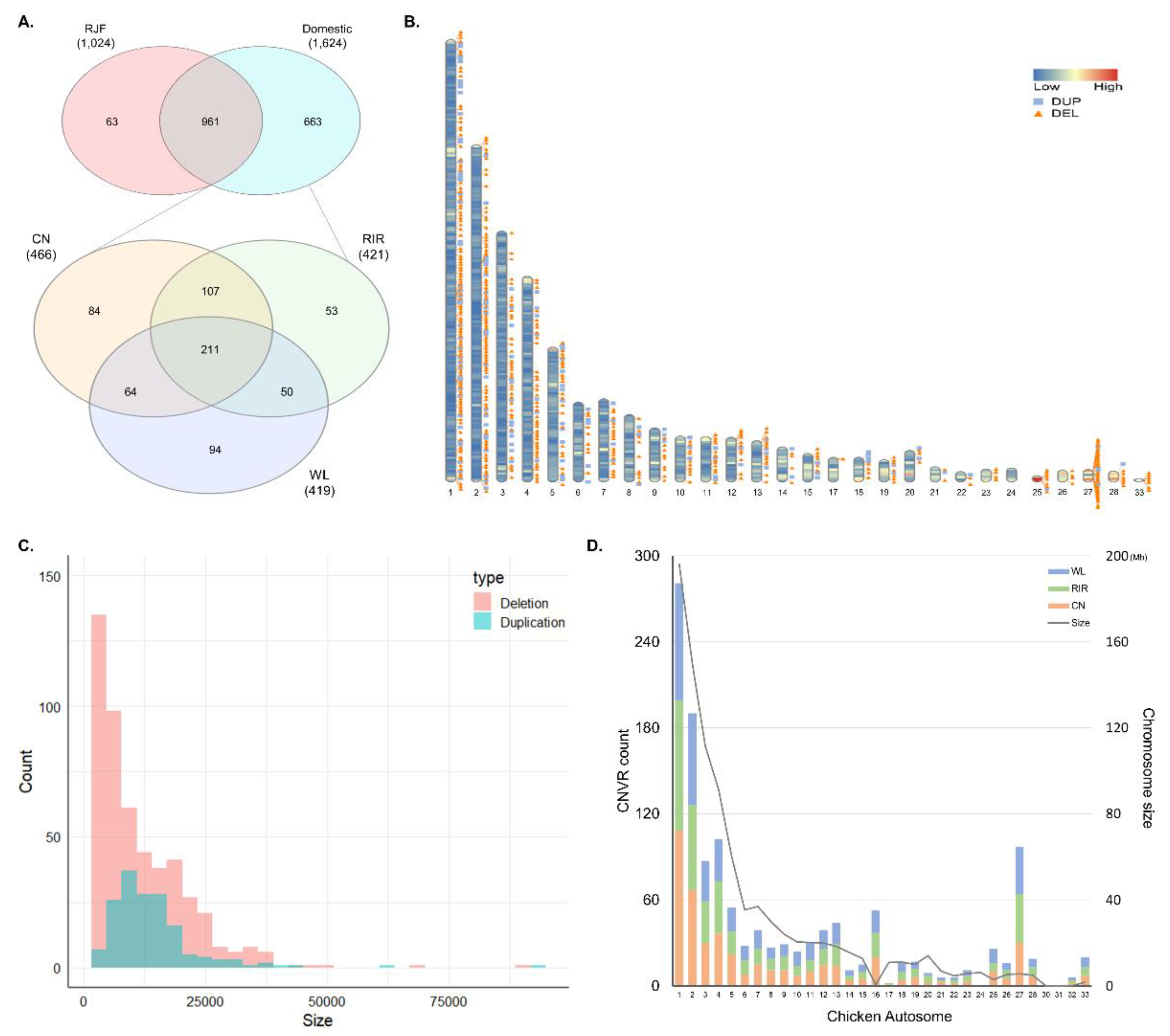
3.1. Identification of CNVs and CNVRs
3.2. Annotation of CNVR
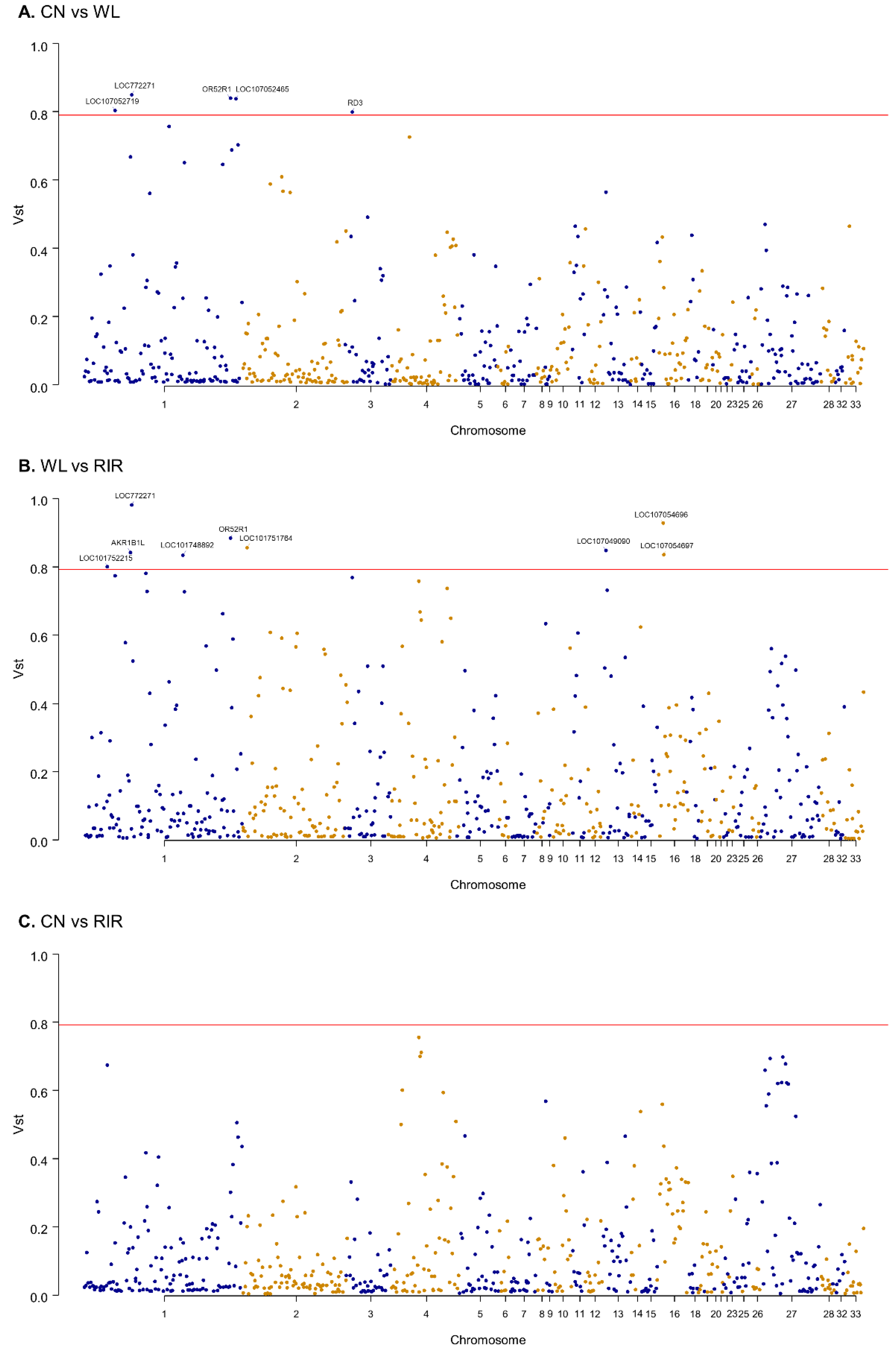
3.3. Breed Differentiation of CNVR
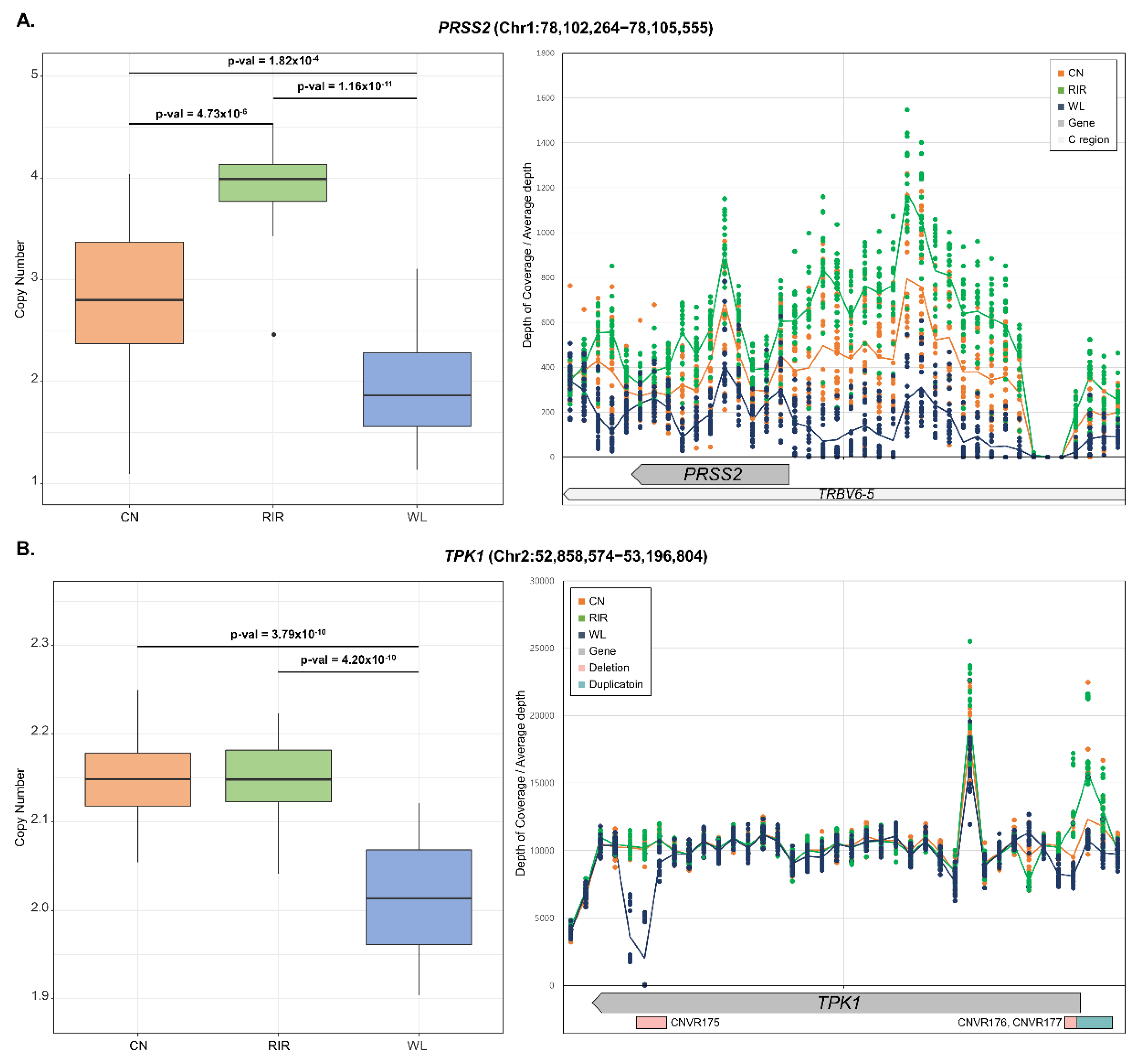
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Feuk, L.; Carson, A.R.; Scherer, S.W. Structural variation in the human genome. Nat. Rev. Genet. 2006, 7, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and functional impact of copy number variation in the human genome. Nature 2010, 464, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lemos, B.; Dopman, E.B.; Hartl, D.L. Copy-number variation: The balance between gene dosage and expression in Drosophila melanogaster. Genome Biol. Evol. 2011, 3, 1014–1024. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.E.; Walter, K.; Stewart, C.; Handsaker, R.E.; Chen, K.; Alkan, C.; Abyzov, A.; Yoon, S.C.; Ye, K.; Cheetham, R.K.; et al. Mapping copy number variation by population-scale genome sequencing. Nature 2011, 470, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Keel, B.N.; Lindholm-Perry, A.K.; Snelling, W.M. Evolutionary and functional features of copy number variation in the cattle genome. Front. Genet. 2016, 7, 207. [Google Scholar] [CrossRef] [PubMed]
- Poptsova, M.; Banerjee, S.; Gokcumen, O.; Rubin, M.A.; Demichelis, F. Impact of constitutional copy number variants on biological pathway evolution. BMC Evol. Biol. 2013, 13, 19. [Google Scholar] [CrossRef]
- Poole, A.C.; Goodrich, J.K.; Youngblut, N.D.; Luque, G.G.; Ruaud, A.; Sutter, J.L.; Waters, J.L.; Shi, Q.; El-Hadidi, M.; Johnson, L.M.; et al. Human salivary Amylase gene copy number impacts oral and gut microbiomes. Cell Host Microbe 2019, 25, 553–564. [Google Scholar] [CrossRef]
- Levy-Sakin, M.; Pastor, S.; Mostovoy, Y.; Li, L.; Leung, A.K.Y.; McCaffrey, J.; Young, E.; Lam, E.T.; Hastie, A.R.; Wong, K.H.Y.; et al. Genome maps across 26 human populations reveal population-specific patterns of structural variation. Nat. Commun. 2019, 10, 1025. [Google Scholar] [CrossRef]
- Mathieson, S.; Mathieson, I. FADS1 and the timing of human adaptation to agriculture. Mol. Biol. Evol. 2018, 35, 2957–2970. [Google Scholar] [CrossRef]
- Kommadath, A.; Grant, J.R.; Krivushin, K.; Butty, A.M.; Baes, C.F.; Carthy, T.R.; Berry, D.P.; Stothard, P. A large interactive visual database of copy number variants discovered in taurine cattle. GigaScience 2019, 8, giz073. [Google Scholar] [CrossRef]
- Liu, M.; Fang, L.; Liu, S.; Pan, M.G.; Seroussi, E.; Cole, J.B.; Ma, L.; Chen, H.; Liu, G.E. Array CGH-based detection of CNV regions and their potential association with reproduction and other economic traits in Holsteins. BMC Genom. 2019, 20, 181. [Google Scholar] [CrossRef] [PubMed]
- Geistlinger, L.; da Silva, V.H.; Cesar, A.S.M.; Tizioto, P.C.; Waldron, L.; Zimmer, R.; Regitano, L.C.D.A.; Coutinho, L.L. Widespread modulation of gene expression by copy number variation in skeletal muscle. Sci. Rep. 2018, 8, 1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revilla, M.; Puig-Oliveras, A.; Castelló, A.; Crespo-Piazuelo, D.; Paludo, E.; Fernández, A.I.; Ballester, M.; Folch, J.M. A global analysis of CNVs in swine using whole genome sequence data and association analysis with fatty acid composition and growth traits. PLoS ONE 2017, 12, e0177014. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.-Q.; Pan, H.; Huang, S.-H.; Liu, C.; Niu, X.; Li, S.; Wang, J.-F. Copy number variations of MTHFSD gene across pig breeds and its association with litter size traits in Chinese indigenous Xiang pig. J. Anim. Physiol. Anim. Nutr. (Berl) 2018, 102, 1320–1327. [Google Scholar] [CrossRef]
- Dong, K.; Pu, Y.; Yao, N.; Shu, G.; Liu, X.; He, X.; Zhao, Q.; Guan, W.; Ma, Y. Copy number variation detection using SNP genotyping arrays in three Chinese pig breeds. Anim. Genet. 2015, 46, 101–109. [Google Scholar] [CrossRef]
- Gorla, E.; Cozzi, M.C.; Román-Ponce, S.I.; Ruiz López, F.J.; Vega-Murillo, V.E.; Cerolini, S.; Bagnato, A.; Strillacci, M.G. Genomic variability in Mexican chicken population using copy number variants. BMC Genet. 2017, 18, 61. [Google Scholar] [CrossRef]
- Lin, S.; Lin, X.; Zhang, Z.; Jiang, M.; Rao, Y.; Nie, Q.; Zhang, X. Copy number variation in SOX6 contributes to chicken muscle development. Genes 2018, 9, 42. [Google Scholar] [CrossRef]
- Rao, Y.S.; Li, J.; Zhang, R.; Lin, X.R.; Xu, J.G.; Xie, L.; Xu, Z.Q.; Wang, L.; Gan, J.K.; Xie, X.J.; et al. Copy number variation identification and analysis of the chicken genome using a 60K SNP BeadChip. Poult. Sci. 2016, 95, 1750–1756. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, Y.; Rosen, B.D.; Van Tassell, C.P.; Stella, A.; Tosser-Klopp, G.; Rupp, R.; Palhière, I.; Colli, L.; Sayre, B.; et al. Diversity of copy number variation in the worldwide goat population. Heredity 2019, 122, 636–646. [Google Scholar] [CrossRef]
- Serres-Armero, A.; Povolotskaya, I.S.; Quilez, J.; Ramirez, O.; Santpere, G.; Kuderna, L.F.K.; Hernandez-Rodriguez, J.; Fernandez-Callejo, M.; Gomez-Sanchez, D.; Freedman, A.H.; et al. Similar genomic proportions of copy number variation within gray wolves and modern dog breeds inferred from whole genome sequencing. BMC Genom. 2017, 18, 977. [Google Scholar] [CrossRef]
- Genova, F.; Longeri, M.; Lyons, L.A.; Bagnato, A.; Gandolfi, B.; Aberdein, D.; Alves, P.C.; Barsh, G.S.; Beale, H.C.; Bergström, T.F.; et al. First genome-wide CNV mapping in FELIS CATUS using next generation sequencing data. BMC Genom. 2018, 19, 895. [Google Scholar] [CrossRef] [PubMed]
- Lauer, S.; Avecilla, G.; Spealman, P.; Sethia, G.; Brandt, N.; Levy, S.F.; Gresham, D. Single-cell copy number variant detection reveals the dynamics and diversity of adaptation. PLoS Biol. 2018, 16, e3000069. [Google Scholar] [CrossRef] [PubMed]
- Hosgood, H.D., III; Hu, W.; Rothman, N.; Klugman, M.; Weinstein, S.J.; Virtamo, J.R.; Albanes, D.; Cawthon, R.; Lan, Q. Variation in ribosomal DNA copy number is associated with lung cancer risk in a prospective cohort study. Carcinogenesis 2019, 40, 975–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic hallmarks and structural variation in metastatic prostate cancer. Cell 2018, 174, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Warland, A.; Kendall, K.M.; Rees, E.; Kirov, G.; Caseras, X. Schizophrenia-associated genomic copy number variants and subcortical brain volumes in the UK Biobank. Mol. Psychiatry 2019. [Google Scholar] [CrossRef]
- Kendall, K.M.; Rees, E.; Bracher-Smith, M.; Legge, S.; Riglin, L.; Zammit, S.; O’Donovan, M.C.; Owen, M.J.; Jones, I.; Kirov, G.; et al. Association of rare copy number variants with risk of depression. JAMA Psychiatry 2019, 76, 818–825. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Chhotaray, S.; Pruthviraj, D.R.; Rawat, C.; Karthikeyan, A.; Panigrahi, M. Copy number variation in livestock: A mini review. Vet. World 2018, 11, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Bickhart, D.; Liu, G. The challenges and importance of structural variation detection in livestock. Front. Genet. 2014, 5, 37. [Google Scholar] [CrossRef]
- Clop, A.; Vidal, O.; Amills, M. Copy number variation in the genomes of domestic animals. Anim. Genet. 2012, 43, 503–517. [Google Scholar] [CrossRef]
- Al-Nasser, A.; Al-Khalaifa, H.; Al-Saffar, A.; Khalil, F.; Albahouh, M.; Ragheb, G.; Al-Haddad, A.; Mashaly, M. Overview of chicken taxonomy and domestication. World’s Poult. Sci. J. 2007, 63, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Herron, L.R.; Pridans, C.; Turnbull, M.L.; Smith, N.; Lillico, S.; Sherman, A.; Gilhooley, H.J.; Wear, M.; Kurian, D.; Papadakos, G.; et al. A chicken bioreactor for efficient production of functional cytokines. BMC Biotechnol. 2018, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Schock, E.N.; Chang, C.-F.; Youngworth, I.A.; Davey, M.G.; Delany, M.E.; Brugmann, S.A. Utilizing the chicken as an animal model for human craniofacial ciliopathies. Dev. Biol. 2016, 415, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.-J.; Zody, M.C.; Eriksson, J.; Meadows, J.R.S.; Sherwood, E.; Webster, M.T.; Jiang, L.; Ingman, M.; Sharpe, T.; Ka, S.; et al. Whole-genome resequencing reveals loci under selection during chicken domestication. Nature 2010, 464, 587. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Robertson, L.B.; Tempest, H.G.; Vignal, A.; Fillon, V.; Crooijmans, R.P.M.A.; Groenen, M.A.M.; Deryusheva, S.; Gaginskaya, E.; Carré, W.; et al. Whole genome comparative studies between chicken and turkey and their implications for avian genome evolution. BMC Genom. 2008, 9, 168. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Boije, H.; Meadows, J.R.S.; Bed’hom, B.; Gourichon, D.; Vieaud, A.; Tixier-Boichard, M.; Rubin, C.-J.; Imsland, F.; Hallböök, F.; et al. Copy number variation in Intron 1 of SOX5 causes the Pea-comb phenotype in chickens. PLoS Genet. 2009, 5, e1000512. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Chen, S.; Zhou, H.; Li, D.; Liu, W.; Yang, N. Copy number variations identified in the chicken using a 60K SNP BeadChip. Anim. Genet. 2013, 44, 276–284. [Google Scholar] [CrossRef]
- Escaramís, G.; Docampo, E.; Rabionet, R. A decade of structural variants: Description, history and methods to detect structural variation. Brief. Funct. Genom. 2015, 14, 305–314. [Google Scholar] [CrossRef]
- Hayes, J.L.; Tzika, A.; Thygesen, H.; Berri, S.; Wood, H.M.; Hewitt, S.; Pendlebury, M.; Coates, A.; Willoughby, L.; Watson, C.M.; et al. Diagnosis of copy number variation by Illumina next generation sequencing is comparable in performance to oligonucleotide array comparative genomic hybridisation. Genomics 2013, 102, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Kerstens, H.H.D.; Crooijmans, R.P.M.A.; Dibbits, B.W.; Vereijken, A.; Okimoto, R.; Groenen, M.A.M. Structural variation in the chicken genome identified by paired-end next-generation DNA sequencing of reduced representation libraries. BMC Genom. 2011, 12, 94. [Google Scholar] [CrossRef]
- Fan, W.-L.; Ng, C.S.; Chen, C.-F.; Lu, M.-Y.J.; Chen, Y.-H.; Liu, C.-J.; Wu, S.-M.; Chen, C.-K.; Chen, J.-J.; Mao, C.-T.; et al. Genome-wide patterns of genetic variation in two domestic chickens. Genome Biol. Evol. 2013, 5, 1376–1392. [Google Scholar] [CrossRef]
- Yi, G.; Qu, L.; Liu, J.; Yan, Y.; Xu, G.; Yang, N. Genome-wide patterns of copy number variation in the diversified chicken genomes using next-generation sequencing. BMC Genom. 2014, 15, 962. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Yang, N.; Cheng, H.H.; Song, J.; Qu, L. Genome-wide identification of copy number variations between two chicken lines that differ in genetic resistance to Marek’s disease. BMC Genom. 2015, 16, 843. [Google Scholar] [CrossRef] [PubMed]
- Sohrabi, S.S.; Mohammadabadi, M.; Wu, D.-D.; Esmailizadeh, A. Detection of breed-specific copy number variations in domestic chicken genome. Genome 2017, 61, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Cutler, D.J.; Jensen, J.D. To pool, or not to pool? Genetics 2010, 186, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Abyzov, A.; Urban, A.E.; Snyder, M.; Gerstein, M. CNVnator: An approach to discover, genotype, and characterize typical and atypical CNVs from family and population genome sequencing. Genome Res. 2011, 21, 974–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.H.; Lee, J.H.; Kong, H.S. Assessment of genetic diversity and phylogenetic relationships of Korean native chicken breeds using microsatellite markers. Asian Australas. J. Anim. Sci. 2017, 30, 1365–1371. [Google Scholar] [CrossRef]
- Seol, Y.-J.; Lee, T.-H.; Park, D.-S.; Kim, C.-K. NABIC: A new access portal to search, visualize, and share agricultural genomics data. Evol. Bioinform. 2016, 12, EBO.S34493. [Google Scholar] [CrossRef]
- Yan, Y.; Yi, G.; Sun, C.; Qu, L.; Yang, N. Genome-wide characterization of insertion and deletion variation in chicken using next generation sequencing. PLoS ONE 2014, 9, e104652. [Google Scholar] [CrossRef]
- Wang, M.-S.; Zhang, R.-W.; Su, L.-Y.; Li, Y.; Peng, M.-S.; Liu, H.-Q.; Zeng, L.; Irwin, D.M.; Du, J.-L.; Yao, Y.-G.; et al. Positive selection rather than relaxation of functional constraint drives the evolution of vision during chicken domestication. Cell Res. 2016, 26, 556. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef] [PubMed]
- Trost, B.; Walker, S.; Wang, Z.; Thiruvahindrapuram, B.; MacDonald, J.R.; Sung, W.W.L.; Pereira, S.L.; Whitney, J.; Chan, A.J.S.; Pellecchia, G.; et al. A comprehensive workflow for read depth-based identification of copy-number variation from whole-genome sequence data. Am. J. Hum. Genet. 2018, 102, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Pierce, M.D.; Dzama, K.; Muchadeyi, F.C. Genetic diversity of seven cattle breeds inferred using copy number variations. Front. Genet. 2018, 9, 163. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; He, Q.-Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Hu, Z.L.; Park, C.A.; Reecy, J.M. Building a livestock genetic and genomic information knowledgebase through integrative developments of Animal QTLdb and CorrDB. Nucleic Acids Res. 2019, 47, D701–D710. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Suzuki, R.; Shimodaira, H. Pvclust: An R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 2006, 22, 1540–1542. [Google Scholar] [CrossRef]
- Redon, R.; Ishikawa, S.; Fitch, K.R.; Feuk, L.; Perry, G.H.; Andrews, T.D.; Fiegler, H.; Shapero, M.H.; Carson, A.R.; Chen, W.; et al. Global variation in copy number in the human genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.S.; Quinlan, A.R. Mosdepth: Quick coverage calculation for genomes and exomes. Bioinformatics 2017, 34, 867–868. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Ho, S.S.; Zhang, X.; Pattni, R.; Haraksingh, R.R.; Urban, A.E. Whole-genome sequencing analysis of CNV using low-coverage and paired-end strategies is efficient and outperforms array-based CNV analysis. J. Med. Genet. 2018, 55, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Byers, S. Copy Number Variation in chickens: A review and future prospects. Microarrays 2014, 3, 24–38. [Google Scholar] [CrossRef]
- Zarrei, M.; MacDonald, J.R.; Merico, D.; Scherer, S.W. A copy number variation map of the human genome. Nat. Rev. Genet. 2015, 16, 172. [Google Scholar] [CrossRef]
- Lawal, R.A.; Al-Atiyat, R.M.; Aljumaah, R.S.; Silva, P.; Mwacharo, J.M.; Hanotte, O. Whole-genome resequencing of red Junglefowl and indigenous village chicken reveal new insights on the genome dynamics of the species. Front. Genet. 2018, 9, 264. [Google Scholar] [CrossRef]
- Miller, M.M.; Taylor, R.L., Jr. Brief review of the chicken major histocompatibility complex: The genes, their distribution on chromosome 16, and their contributions to disease resistance. Poult. Sci. 2016, 95, 375–392. [Google Scholar] [CrossRef]
- Moiseyeva, I.G.; Romanov, M.N.; Nikiforov, A.A.; Sevastyanova, A.A.; Semyenova, S.K. Evolutionary relationships of Red Jungle Fowl and chicken breeds. Genet. Sel. Evol. 2003, 35, 403. [Google Scholar] [CrossRef]
- Khawaja, T.; Khan, S.H.; Mukhtar, N.; Ullah, N.; Parveen, A. Production performance, egg quality and biochemical parameters of Fayoumi, Rhode Island Red and their reciprocal crossbred chickens. J. Appl. Anim. Res. 2013, 41, 208–217. [Google Scholar] [CrossRef]
- Qanbari, S.; Rubin, C.-J.; Maqbool, K.; Weigend, S.; Weigend, A.; Geibel, J.; Kerje, S.; Wurmser, C.; Peterson, A.T.; Brisbin, I.L., Jr.; et al. Genetics of adaptation in modern chicken. PLoS Genet. 2019, 15, e1007989. [Google Scholar] [CrossRef]
- Ma, J.; Yan, R.; Zu, X.; Cheng, J.-M.; Rao, K.; Liao, D.-F.; Cao, D. Aldo-keto reductase family 1 B10 affects fatty acid synthesis by regulating the stability of Acetyl-CoA Carboxylase-α in breast cancer cells. J. Biol. Chem. 2008, 283, 3418–3423. [Google Scholar] [CrossRef] [PubMed]
- Claire D’Andre, H.; Paul, W.; Shen, X.; Jia, X.; Zhang, R.; Sun, L.; Zhang, X. Identification and characterization of genes that control fat deposition in chickens. J. Anim. Sci. Biotechnol. 2013, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Nahashon, S.; Feaster, T.K.; Bohannon-Stewart, A.; Adefope, N. An initial map of chromosomal segmental copy number variations in the chicken. BMC Genom. 2010, 11, 351. [Google Scholar] [CrossRef] [PubMed]
- Paudel, Y.; Madsen, O.; Megens, H.-J.; Frantz, L.A.F.; Bosse, M.; Bastiaansen, J.W.M.; Crooijmans, R.P.M.A.; Groenen, M.A.M. Evolutionary dynamics of copy number variation in pig genomes in the context of adaptation and domestication. BMC Genom. 2013, 14, 449. [Google Scholar] [CrossRef]
- Upadhyay, M.; da Silva, V.H.; Megens, H.-J.; Visker, M.H.P.W.; Ajmone-Marsan, P.; Bâlteanu, V.A.; Dunner, S.; Garcia, J.F.; Ginja, C.; Kantanen, J.; et al. Distribution and functionality of copy number variation across european cattle populations. Front. Genet. 2017, 8, 108. [Google Scholar] [CrossRef]
- Krause, E.T.; Schrader, L.; Caspers, B.A. Olfaction in chicken (Gallus gallus): A neglected mode of social communication? Front. Ecol. Evol. 2016, 4, 94. [Google Scholar] [CrossRef]
- Roth, T.C.; Cox, J.G.; Lima, S.L. Can foraging birds assess predation risk by scent? Anim. Behav. 2008, 76, 2021–2027. [Google Scholar] [CrossRef]
- Steiger Silke, S.; Fidler Andrew, E.; Valcu, M.; Kempenaers, B. Avian olfactory receptor gene repertoires: evidence for a well-developed sense of smell in birds? Proc. Biol. Sci. 2008, 275, 2309–2317. [Google Scholar] [CrossRef] [Green Version]
- Azadi, S.; Molday, L.L.; Molday, R.S. RD3, the protein associated with Leber congenital amaurosis type 12, is required for guanylate cyclase trafficking in photoreceptor cells. Proc. Natl. Acad. Sci. USA 2010, 107, 21158–21163. [Google Scholar] [CrossRef] [Green Version]
- Renema, R.A.; Robinson, F.E.; Feddes, J.J.R.; Fasenko, G.M.; Zuidhoft, M.J. Effects of light intensity from photostimulation in four strains of commercial egg layers: 2. egg production parameters. Poult. Sci. 2001, 80, 1121–1131. [Google Scholar] [CrossRef]
- Jácome, I.; Rossi, L.; Borille, R. Influence of artificial lighting on the performance and egg quality of commercial layers: A review. Braz. J. Poult. Sci. 2014, 16, 337–344. [Google Scholar] [CrossRef]
- Dalgin, G.S.; Holloway, D.T.; Liou, L.S.; DeLisi, C. Identification and characterization of renal cell carcinoma gene markers. Cancer Inform. 2007, 3, 65–92. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Temperley, N.D.; Ren, L.; Smith, J.; Li, N.; Burt, D.W. Molecular evolution of the vertebrate TLR1 gene family—A complex history of gene duplication, gene conversion, positive selection and co-evolution. BMC Evol. Biol. 2011, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.K.; Wu, Y.H.; An, J.; Cui, D.F.; Li, H.R.; Zheng, S.J. Toll-like receptor 2 type 1 and type 2 polymorphisms in different chicken breeds. Poult. Sci. 2012, 91, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.M.; Masson, E.; Le Maréchal, C.; Férec, C. Copy number variations in chronic pancreatitis. Cytogenet. Genome Res. 2008, 123, 102–107. [Google Scholar] [CrossRef]
- Luo, S.; Yu, J.A.; Li, H.; Song, Y.S. Worldwide genetic variation of the IGHV and TRBV immune receptor gene families in humans. Life Sci. Alliance 2019, 2, e201800221. [Google Scholar] [CrossRef] [Green Version]
- Muroya, S.; Oe, M.; Ojima, K. Thiamine accumulation and thiamine triphosphate decline occur in parallel with ATP exhaustion during postmortem aging of pork muscles. Meat Sci. 2018, 137, 228–234. [Google Scholar] [CrossRef]
- Bettendorff, L.; Wins, P. Thiamin diphosphate in biological chemistry: New aspects of thiamin metabolism, especially triphosphate derivatives acting other than as cofactors. FEBS J. 2009, 276, 2917–2925. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | CNVs | Deletion | Size_Del (bp) | Duplication | Size_Dup (bp) |
|---|---|---|---|---|---|
| Red Jungle Fowl | 714 | 584 | 20,377.4 | 130 | 16,019.1 |
| Cornish | 443 | 327 | 27,572.3 | 116 | 16,156.9 |
| Rhode Island Red | 471 | 346 | 26,778.9 | 125 | 16,394.3 |
| White Leghorn | 467 | 347 | 26,451.8 | 120 | 15,823 |
| Category | ID | Term | Count | p-Value | Genes |
|---|---|---|---|---|---|
| BP | GO:2000251 | Positive regulation of actin cytoskeleton Reorganization | 2 | 1.88E-02 | NTRK3, HRAS |
| BP | GO:0002224 | Toll-like receptor signaling pathway | 2 | 3.90E-02 | TLR2B, TLR2A |
| BP | GO:0050852 | T cell receptor signaling pathway | 2 | 4.81E-02 | HRAS, CD247 |
| BP | GO:0060041 | Retina development in camera-type eye | 2 | 4.99E-02 | LPCAT1, RD3 |
| CC | GO:0031226 | Intrinsic component of plasma membrane | 2 | 1.58E-02 | TLR2B, TLR2A |
| MF | GO:0016491 | Oxidoreductase activity | 5 | 3.44E-05 | LOC425137, LOC107053269, AKR1B1L, LOC772271, ADH6 |
| MF | GO:0004888 | Transmembrane signaling receptor activity | 3 | 2.57E-03 | CD247, TLR2B, TLR2A |
| KEGG PATHWAY | gga01100 | Metabolic pathways | 7 | 3.10E-02 | TPK1, LOC425137, LPCAT1, AKR1B1L, ADH6, TWISTNB, ALDH3B1 |
| KEGG PATHWAY | gga00040 | Pentose and glucuronate interconversions | 2 | 4.25E-02 | LOC425137, AKR1B1L |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seol, D.; Ko, B.J.; Kim, B.; Chai, H.-H.; Lim, D.; Kim, H. Identification of Copy Number Variation in Domestic Chicken Using Whole-Genome Sequencing Reveals Evidence of Selection in the Genome. Animals 2019, 9, 809. https://doi.org/10.3390/ani9100809
Seol D, Ko BJ, Kim B, Chai H-H, Lim D, Kim H. Identification of Copy Number Variation in Domestic Chicken Using Whole-Genome Sequencing Reveals Evidence of Selection in the Genome. Animals. 2019; 9(10):809. https://doi.org/10.3390/ani9100809
Chicago/Turabian StyleSeol, Donghyeok, Byung June Ko, Bongsang Kim, Han-Ha Chai, Dajeong Lim, and Heebal Kim. 2019. "Identification of Copy Number Variation in Domestic Chicken Using Whole-Genome Sequencing Reveals Evidence of Selection in the Genome" Animals 9, no. 10: 809. https://doi.org/10.3390/ani9100809