Association of Twelve Candidate Gene Polymorphisms with the Intramuscular Fat Content and Average Backfat Thickness of Chinese Suhuai Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
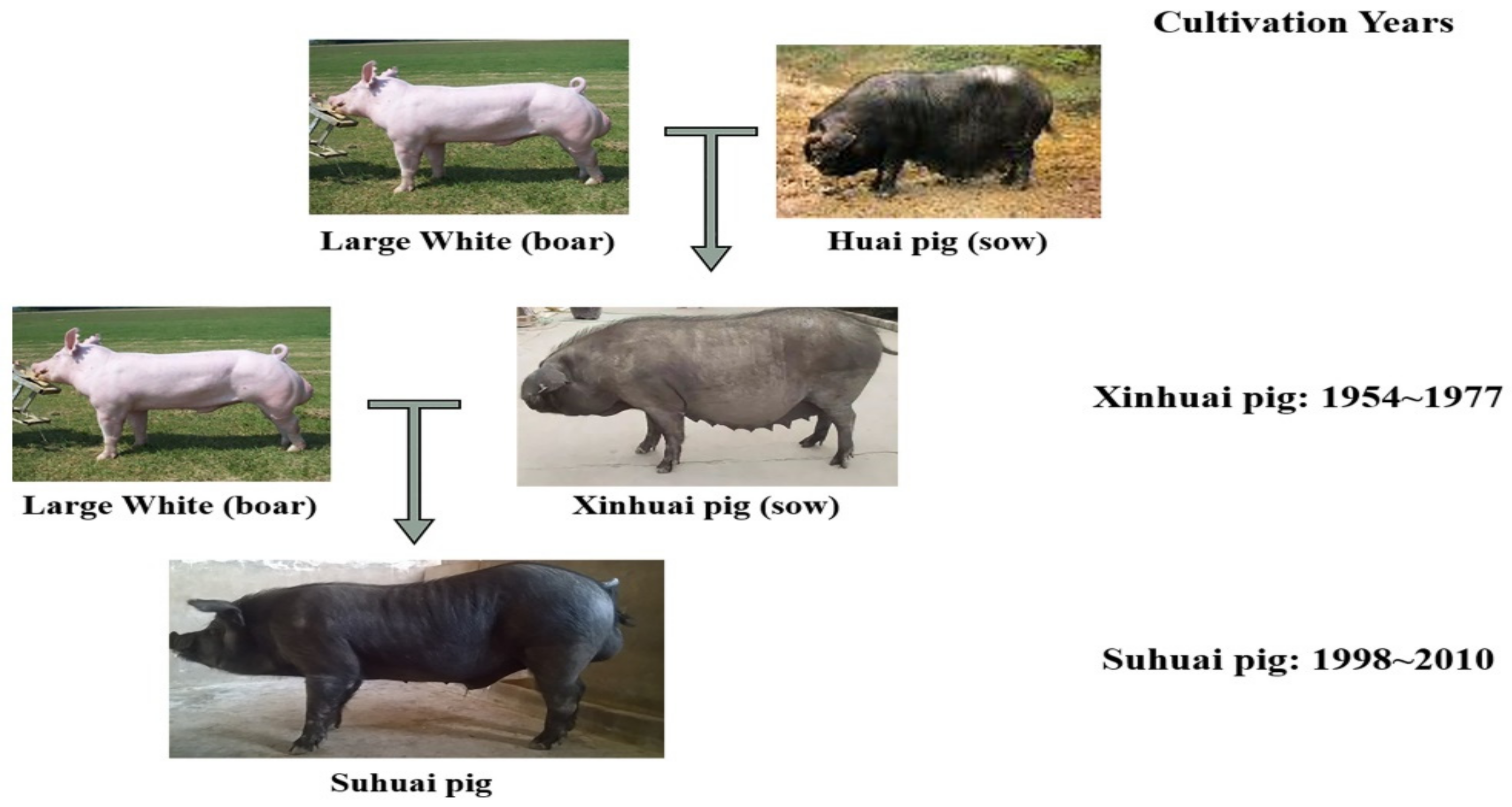
2.2. Animals and Phenotype Measurements
2.3. Isolation of Genomic DNA and SNPs Genotyping
2.4. Statistical Analysis
3. Results
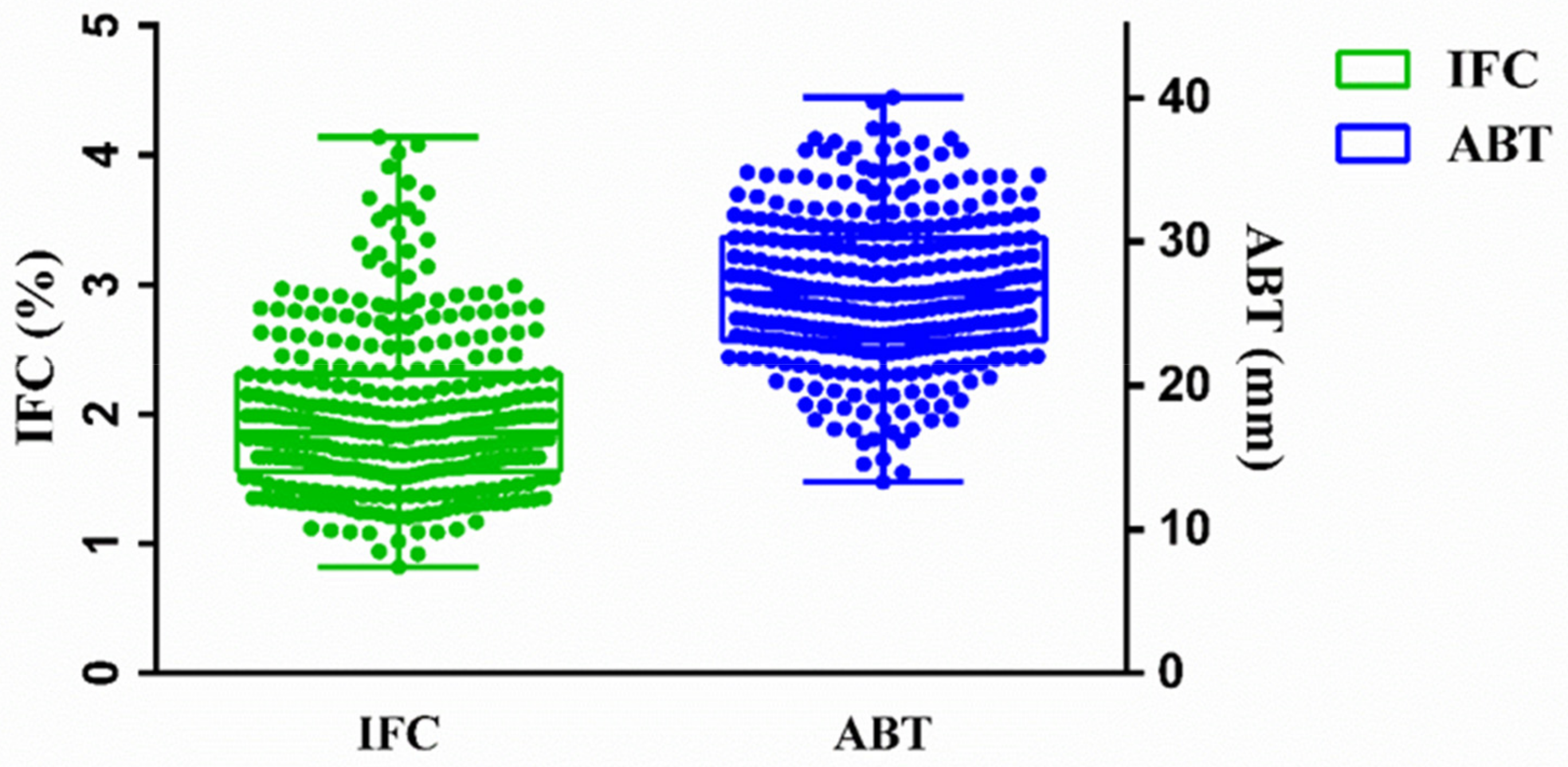
3.1. Descriptive Statistics for IFC and ABT
3.2. Estimates of Phenotypic and Genetic Correlations of Suhuai Pigs
3.3. Genetic Parameters of These SNPs
3.4. Association Analysis between SNPs and IFC and ABT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- He, Y.; Yang, X.; Xia, J.; Zhao, L.; Yang, Y. Consumption of meat and dairy products in China: A review. Proc. Nutr. Soc. 2016, 75, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D. Consequences for meat quality of reducing carcass fatness. In Reducing Fat in Meat Animals; Elsevier Applied Science Press: New York, NY, USA, 1990; pp. 344–397. [Google Scholar]
- Li, B.; Weng, Q.; Dong, C.; Zhang, Z.; Li, R.; Liu, J.; Jiang, A.; Li, Q.; Jia, C.; Wu, W. A Key Gene, PLIN1, Can Affect Porcine Intramuscular Fat Content Based on Transcriptome Analysis. Genes 2018, 9, 194. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Font-I-Furnols, M.; Guerrero, L. Consumer preference, behavior and perception about meat and meat products: An overview. Meat Sci. 2014, 98, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.Z.; Zhao, S.M. Physiology, affecting factors and strategies for control of pig meat intramuscular fat. Recent Pat. Food Nutr. Agric. 2009, 1, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Cesar, A.S.; Regitano, L.C.; Koltes, J.E.; Fritz-Waters, E.R.; Lanna, D.P.; Gasparin, G.; Mourão, G.B.; Oliveira, P.S.; Reecy, J.M.; Coutinho, L.L. Putative regulatory factors associated with intramuscular fat content. PLoS ONE 2015, 10, e0128350. [Google Scholar] [CrossRef]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat—1. Composition of the lipid fraction and sensory characteristics of m. longissimus lumborum. Meat Sci. 1999, 53, 59–65. [Google Scholar] [CrossRef]
- Bergeron, K.; Julien, P.; Davis, T.A.; Myre, A.; Thivierge, M.C. Long-chain n-3 fatty acids enhance neonatal insulin-regulated protein metabolism in piglets by differentially altering muscle lipid composition. J. Lipid Res. 2007, 48, 2396–2410. [Google Scholar] [CrossRef] [Green Version]
- Ramayo-Caldas, Y.; Fortes, M.R.S.; Hudson, N.J.; Porto-Neto, L.R.; Bolormaa, S.; Barendse, W.; Kelly, M.; Moore, S.S.; Goddard, M.E.; Lehnert, S.A.; et al. A marker-derived gene network reveals the regulatory role of PPARGC1A, HNF4G, and FOXP3 in intramuscular fat deposition of beef cattle. J. Anim. Sci. 2014, 92, 2832–2845. [Google Scholar] [CrossRef]
- Verbeke, W.; Liu, R. The impacts of information about the risks and benefits of pork consumption on Chinese consumers’ perceptions towards, and intention to eat, pork. Meat Sci. 2014, 98, 766–772. [Google Scholar] [CrossRef]
- Suzuki, K.; Irie, M.; Kadowaki, H.; Shibata, T.; Kumagai, M.; Nishida, A. Genetic parameter estimates of meat quality traits in Duroc pigs selected for average daily gain, longissimus muscle area, backfat thickness, and intramuscular fat content. J. Anim. Sci. 2005, 83, 2058–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbens, F.; Rettenberger, G.; Lenstra, J.A.; Veerkamp, J.H.; te Pas, M.F. Characterization, chromosomal localization, and genetic variation of the porcine heart fatty acid-binding protein gene. Mamm. Genome 1997, 8, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.M.; Ren, L.J.; Chen, L.; Zhang, X.; Cheng, M.L.; Li, W.Z.; Zhang, Y.Y.; Gao, S.Z. Differential Expression of Lipid Metabolism Related Genes in Porcine Muscle Tissue Leading to Different Intramuscular Fat Deposition. Lipids 2009, 44, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.; Isabel, B.; López-Bote, C.J.; Ayuso, M.; Fernández, A.; Óvilo, C.; Benítez, R.; Daza, A. Long term vitamin A restriction improves meat quality parameters and modifies gene expression in Iberian pigs1. J. Anim. Sci. 2015, 93, 2730–2744. [Google Scholar] [CrossRef]
- Burgos, C.; Galve, A.; Moreno, C.; Altarriba, J.; Reina, R.; García, C.; López-Buesa, P. The effects of two alleles of IGF2 on fat content in pig carcasses and pork. Meat Sci. 2011, 90, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Gardan, D.; Gondret, F.; Louveau, I. Lipid metabolism and secretory function of porcine intramuscular adipocytes compared with subcutaneous and perirenal adipocytes. Ajp. Endocrinol. Metab. 2006, 291, E372–E380. [Google Scholar] [CrossRef] [PubMed]
- Rosfreixedes, R.; Gol, S.; Pena, R.N.; Tor, M.; Ibáñezescriche, N.; Dekkers, J.C.M.; Estany, J. Genome-Wide Association Study Singles Out SCD and LEPR as the Two Main Loci Influencing Intramuscular Fat Content and Fatty Acid Composition in Duroc Pigs. PLoS ONE 2016, 11, e0152496. [Google Scholar] [CrossRef]
- Choi, J.S.; Jin, S.K.; Jeong, Y.H.; Jung, Y.C.; Jung, J.H.; Shim, K.S.; Choi, Y.I. Relationships between Single Nucleotide Polymorphism Markers and Meat Quality Traits of Duroc Breeding Stocks in Korea. Asian Australas. J. Anim. Sci. 2016, 29, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yang, J.; Zhou, L.; Ren, J.; Liu, X.; Zhang, H.; Yang, B.; Zhang, Z.; Ma, H.; Xie, X.; et al. A splice mutation in the PHKG1 gene causes high glycogen content and low meat quality in pig skeletal muscle. PLoS Genet. 2014, 10, e1004710. [Google Scholar] [CrossRef]
- Cieslak, J.; Nowacka-Woszuk, J.; Bartz, M.; Fijak-Nowak, H.; Grzes, M.; Szydlowski, M.; Switonski, M. Association studies on the porcine RETN, UCP1, UCP3 and ADRB3 genes polymorphism with fatness traits. Meat Sci. 2009, 83, 551–554. [Google Scholar] [CrossRef]
- Rojas, J.E.; Wilches, M.A.; Cepeda, L.A.; Garcés, M.F.; Suarez, M.A.; Baldrich, R.M.; Vélez, C.A.; Guerrero, M.F.; García, M.R.; Moreno, I.H. Molecular Diagnostics of Porcine Stress Syndrome Susceptibility Associated with the Arg615Cys Mutation Using Real-Time PCR with Fluorescent Hybridization Probes. Rev. Colomb. Anestesiol. 2008, 36, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Supakankul, P.; Mekchay, S. Effect of UBE3C polymorphisms on intramuscular fat content and fatty acid composition in Duroc pigs. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Weng, Q. Comparison of Determination Methods of Porcine Intramuscular Fat Content and Correlation Analysis with Carcass and Meat Quality Traits; Nanjing Agric. Univ.: Nanjing, China, 2017. [Google Scholar]
- Yang, J.; Zhou, L.; Liu, X.; Ma, H.; Xie, X.; Xiong, X.; Hou, L.; Huang, Y.; Chen, C.; Ma, J. A comparative study of meat quality traits between Laiwu and DLY pigs. Acta Vet. Zootech. Sin. 2014, 45, 1752–1759. [Google Scholar] [CrossRef]
- Wei-Hua, X.U.; Kang, L.Z.; Lin, X.J.; Yang, Z.S.; Gao, W.P.; Wang, Z.C. Polymorphism Analysis of H-FABP Gene and Its Correlation with IMF Content in Huai Pig. J. Fujian Norm. Univ. 2014, 30, 76–80. [Google Scholar]
- Liu, Y.; Yang, X.; Jing, X.; He, X.; Wang, L.; Liu, Y.; Liu, D. Transcriptomics Analysis on Excellent Meat Quality Traits of Skeletal Muscles of the Chinese Indigenous Min Pig Compared with the Large White Breed. Int. J. Mol. Sci. 2017, 19, 21. [Google Scholar] [CrossRef]
- Wang, B.; Wang, H.; Xiang, M.A.; Guang, P.U.; Niu, P.; Zhang, Z.; Zhou, J.; Pinghua, L.I.; Huang, R. Analysis of intramuscular fat content in Suhuai pigs and its correlation with polymorphisms of IGF2 and SCD genes. Anim. Husb. Vet. Med. J. 2018, 50, 20–25. [Google Scholar]
- Supakankul, P.; Mekchay, S. Association of NLK polymorphisms with intramuscular fat content and fatty acid composition traits in pigs. Meat Sci. 2016, 118, 61–65. [Google Scholar] [CrossRef]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 49, pp. 895–909. [Google Scholar]
- Xie, B.; Zhang, P.; Liu, M.; Zeng, W.; Yang, J.; Liu, H. Deltex1 Polymorphisms Are Associated with Hepatitis B Vaccination Non-Response in Southwest China. PLoS ONE 2016, 11, e0149199. [Google Scholar] [CrossRef]
- Madsen, P.; Sørensen, P.; Su, G.; Damgaard, L.H.; Thomsen, H.; Labouriau, R. DMU—A package for analyzing multivariate mixed models. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Brazil, 13–18 August 2006. [Google Scholar]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Gong, H.; Cui, L.; Zhang, W.; Ma, J.; Chen, C.; Ai, H.; Xiao, S.; Huang, L.; et al. Genetic correlation of fatty acid composition with growth, carcass, fat deposition and meat quality traits based on GWAS data in six pig populations. Meat Sci. 2019, 150, 47–55. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, L.; Zhang, L.; Liu, X.; Hou, X.; Gao, H.; Yan, H.; Zhao, F.; Wang, L. NTN1 Affects Porcine Intramuscular Fat Content by Affecting the Expression of Myogenic Regulatory Factors. Animals 2019, 9, 609. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gong, J.; Wang, L.; Hou, X.; Gao, H.; Yan, H.; Zhao, F.; Zhang, L.; Wang, L. Genome-Wide Profiling of the Microrna Transcriptome Regulatory Network to Identify Putative Candidate Genes Associated with Backfat Deposition in Pigs. Animals 2019, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Gol, S.; González-Prendes, R.; Bosch, L.; Tor, M.; Reixach, J.; Pena, R.N.; Estany, J. Linoleic acid metabolic pathway allows for an efficient increase of intramuscular fat content in pigs. J. Anim. Sci. Biotechnol. 2019, 10, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jacyno, E.; Pietruszka, A.; Kawęcka, M.; Biel, W.; Kołodziej-Skalska, A. Phenotypic Correlations of Backfat Thickness with Meatiness Traits, Intramuscular Fat, Longissimus Muscle Cholesterol and Fatty Acid Composition in Pigs. S. Afr. J. Anim. Sci. 2015, 45, 122–128. [Google Scholar] [CrossRef]
- Fujii, J.; Otsu, K.; Zorzato, F.; de Leon, S.; Khanna, V.K.; Weiler, J.E.; O’Brien, P.J.; MacLennan, D.H. Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science 1991, 253, 448–451. [Google Scholar] [CrossRef]
- Veerkamp, J.H.; Maatman, R.G. Cytoplasmic fatty acid-binding proteins: Their structure and genes. Prog. Lipid Res. 1995, 34, 17–52. [Google Scholar] [CrossRef]
- Chen, J.N.; Jiang, Y.Z.; Cen, W.M.; Xing, S.H.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Lou, P.E.; Wen, A.X.; et al. Distribution of H-FABP and ACSL4 gene polymorphisms and their associations with intramuscular fat content and backfat thickness in different pig populations. Genet. Mol. Res. 2014, 13, 6759–6772. [Google Scholar] [CrossRef]
- Serao, N.V.L.; Veroneze, R.; Ribeiro, A.M.F.; Verardo, L.L.; Braccini Neto, J.; Gasparino, E.; Campos, C.F.; Lopes, P.S.; Guimaraes, S.E.F. Candidate gene expression and intramuscular fat content in pigs. J. Anim. Breed. Genet. 2011, 128, 28–34. [Google Scholar] [CrossRef]
- Matika, O.; Robledo, D.; Pong-Wong, R.; Bishop, S.C.; Riggio, V.; Finlayson, H.; Lowe, N.R.; Hoste, A.E.; Walling, G.A.; Del-Pozo, J.; et al. Balancing selection at a premature stop mutation in the myostatin gene underlies a recessive leg weakness syndrome in pigs. PLoS Genet. 2019, 15, e1007759. [Google Scholar] [CrossRef]
- Lim, K.S.; Lee, K.T.; Park, J.E.; Chung, W.H.; Jang, G.W.; Choi, B.H.; Hong, K.C.; Kim, T.H. Identification of differentially expressed genes in longissimus muscle of pigs with high and low intramuscular fat content using RNA sequencing. Anim. Genet. 2017, 48, 166–174. [Google Scholar] [CrossRef]
- Henriquez-Rodriguez, E.; Tor, M.; Pena, R.N.; Estany, J. A polymorphism in the stearoyl-CoA desaturase gene promoter increases monounsaturated fatty acid content in dry-cured ham. Meat Sci. 2015, 106, 38–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.P.; Sangwook, K.; Jungsuck, C.; Yoonmi, L.; Cheolkoo, L.; Bonghwan, C.; Taehun, K.; Yangil, C.; Jongjoo, K.; Kwansuk, K. Investigation of porcine FABP3 and LEPR gene polymorphisms and mRNA expression for variation in intramuscular fat content. Mol. Biol. Rep. 2010, 37, 3931–3939. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Suzuki, K. Effects of porcine leptin receptor gene polymorphisms on backfat thickness, fat area ratios by image analysis, and serum leptin concentrations in a Duroc purebred population. Anim. Sci. J. 2012, 83, 375–385. [Google Scholar] [CrossRef]
- Souza, F.R.P.; Mercadante, M.E.Z.; Fonseca, L.F.S.; Ferreira, L.M.S.; Regatieri, I.C.; Ayres, D.R.; Tonhati, H.; Silva, S.L.; Razook, A.G.; Albuquerque, L.G. Assessment of DGAT1 and LEP gene polymorphisms in three Nelore (Bos indicus) lines selected for growth and their relationship with growth and carcass traits. J. Anim. Sci. 2010, 88, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Cepica, S.; Ovilo, C.; Masopust, M.; Knoll, A.; Fernandez, A.; Lopez, A.; Rohrer, G.A.; Nonneman, D. Four genes located on a SSC2 meat quality QTL region are associated with different meat quality traits in Landrace x Chinese-European crossbred population. Anim. Genet. 2012, 43, 333–336. [Google Scholar] [CrossRef]
- Holst, L.S.; Langin, D.; Mulder, H.; Laurell, H.; Grober, J.; Bergh, A.; Mohrenweiser, H.W.; Edgren, G.; Holm, C. Molecular Cloning, Genomic Organization, and Expression of a Testicular Isoform of Hormone-Sensitive Lipase. Genomics 1996, 35, 441–447. [Google Scholar] [CrossRef]
- Aslan, O.; Hamill, R.M.; Davey, G.; Mcbryan, J.; Mullen, A.M.; Gispert, M.; Sweeney, T. Variation in the IGF2 gene promoter region is associated with intramuscular fat content in porcine skeletal muscle. Mol. Biol. Rep. 2012, 39, 4101–4110. [Google Scholar] [CrossRef]
- Uemoto, Y.; Soma, Y.; Sato, S.; Shibata, T.; Kadowaki, H.; Katoh, K.; Kobayashi, E.; Suzuki, K. Mapping QTL for fat area ratios and serum leptin concentrations in a Duroc purebred population. Anim. Sci. J. 2012, 83, 187–193. [Google Scholar] [CrossRef]
- Davoli, R.; Braglia, S.; Valastro, V.; Annarratone, C.; Comella, M.; Zambonelli, P.; Nisi, I.; Gallo, M.; Buttazzoni, L.; Russo, V. Analysis of MC4R polymorphism in Italian Large White and Italian Duroc pigs: Association with carcass traits. Meat Sci. 2012, 90, 887–892. [Google Scholar] [CrossRef]
- Loos, R.J.F.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat. Genet. 2008, 40, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Hao, C.L.; Li, W.H.; Lin, J.L.; Wu, F.R.; Bu-gen, S.; Xie, Z.; Li, Q.F. Association of the MC4R gene expression with intramuscular fat content in Hu sheep muscles. J. Nanjing Agric. Univ. 2011, 34, 143–146. [Google Scholar]
- Lyadskiy, I.K.; Getya, A.A.; Pochernyaev, K.F. Association of the Asp298Asn polymorphism in the mc4r gene with back fat thickness in pigs of the large white breed. Cytol. Genet. 2011, 45, 106–109. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Number | Mean ± SE | Max | Min | CV% |
|---|---|---|---|---|---|
| IFC | 330 | 1.99 ± 0.03% | 4.14% | 0.82% | 31.21 |
| ABT | 330 | 26.68 ± 0.28 mm | 40.02 mm | 13.13 mm | 19.36 |
| Gene (SNP) | Chromosome | Genotype (Number) | Frequency | HE | HO | PIC | |
|---|---|---|---|---|---|---|---|
| Genotype | Allele | ||||||
| FABP3 rs1110770079 | 6 | GG (82) | 0.25 | 0.44 (G) | 0.36 | 0.64 | 0.37 |
| GT (117) | 0.36 | ||||||
| TT (123) | 0.38 | 0.56 (T) | |||||
| LIPE rs328830166 | 6 | AA (53) | 0.16 | 0.40 (A) | 0.48 | 0.52 | 0.36 |
| AG (157) | 0.48 | ||||||
| GG (120) | 0.36 | 0.60 (G) | |||||
| IGF1 rs341412920 | 5 | CC (8) | 0.02 | 0.16 (C) | 0.28 | 0.72 | 0.24 |
| CT (90) | 0.28 | ||||||
| TT (228) | 0.70 | 0.84 (T) | |||||
| IGF1 rs322131043 | 5 | AG (32) | 0.10 | 0.05 (A) | 0.10 | 0.90 | 0.09 |
| GG (298) | 0.90 | 0.95 (G) | |||||
| IGF2 g.3072G > A | 2 | AA (62) | 0.19 | 0.42 (A) | 0.46 | 0.54 | 0.37 |
| AG (151) | 0.46 | ||||||
| GG (117) | 0.35 | 0.58 (G) | |||||
| LEP rs45431504 | 18 | CC (1) | 0.01 | 0.08 (C) | 0.15 | 0.85 | 0.14 |
| CT (51) | 0.15 | ||||||
| TT (278) | 0.84 | 0.92 (T) | |||||
| LEPR rs45435518 | 6 | CC (2) | 0.01 | 0.09 (C) | 0.16 | 0.84 | 0.15 |
| CT (53) | 0.16 | ||||||
| TT (274) | 0.83 | 0.91 (T) | |||||
| MC4R rs81219178 | 1 | AA (109) | 0.33 | 0.60 (A) | 0.53 | 0.47 | 0.37 |
| AG (175) | 0.53 | ||||||
| GG (45) | 0.14 | 0.40 (G) | |||||
| PHKG1 rs697732005 | 3 | AA (184) | 0.57 | 0.70 (A) | 0.26 | 0.74 | 0.33 |
| AG (85) | 0.26 | ||||||
| GG (53) | 0.16 | 0.30 (G) | |||||
| RETN rs327132149 | 2 | AA (1) | 0.00 | 0.03 (A) | 0.06 | 0.94 | 0.06 |
| AG (20) | 0.06 | ||||||
| GG (309) | 0.94 | 0.97 (G) | |||||
| RYR1 rs344435545 | 6 | CC (309) | 0.94 | 0.97 (C) | 0.06 | 0.94 | 0.06 |
| CT (20) | 0.06 | ||||||
| TT (1) | 0.00 | 0.03 (T) | |||||
| SCD rs80912566 | 14 | CC (59) | 0.18 | 0.41 (C) | 0.47 | 0.53 | 0.37 |
| CT (154) | 0.47 | ||||||
| TT (117) | 0.35 | 0.59 (T) | |||||
| UBE3C rs81329544 | 18 | AA (14) | 0.04 | 0.22 (A) | 0.35 | 0.65 | 0.28 |
| AG (117) | 0.35 | ||||||
| GG (199) | 0.60 | 0.78 (G) | |||||
| Gene (SNP) | Genotype | Number | IFC (%) | ABT (mm) | ||
|---|---|---|---|---|---|---|
| Mean ± SE | p Value | Mean ± SE | p Value | |||
| FABP3 rs1110770079 | GG | 82 | 2.19 ± 0.09 a | 0.0230 | 26.89 ± 0.57 | 0.6040 |
| GT | 117 | 2.13 ± 0.07 ab | 26.18 ± 0.49 | |||
| TT | 123 | 1.93 ± 0.07 b | 26.49 ± 0.52 | |||
| LIPE rs328830166 | AA | 53 | 2.05 ± 0.08 | 0.4204 | 26.82 ± 0.70 | 0.9068 |
| AG | 157 | 1.95 ± 0.05 | 28.14 ± 4.53 | |||
| GG | 120 | 2.03 ± 0.06 | 27.82 ± 1.03 | |||
| IGF1 rs341412920 | CC | 8 | 1.92 ± 0.20 | 0.5286 | 26.67 ± 1.61 | 0.9857 |
| CT | 90 | 1.94 ± 0.06 | 26.52 ± 0.51 | |||
| TT | 228 | 2.02 ± 0.04 | 26.61 ± 0.35 | |||
| IGF1 rs322131043 | AG | 32 | 2.00 ± 0.10 | 0.9981 | 26.50 ± 0.31 | 0.5349 |
| GG | 298 | 2.00 ± 0.04 | 27.22 ± 1.22 | |||
| IGF2 g.3072G > A | AA | 62 | 2.01 ± 0.07 | 0.9340 | 26.50 ± 0.44 | 0.4130 |
| AG | 151 | 2.00 ± 0.05 | 26.59 ± 0.37 | |||
| GG | 117 | 1.98 ± 0.06 | 26.21 ± 3.20 | |||
| LEP rs45431504 | CT | 51 | 1.98 ± 0.08 | 0.7460 | 26.11 ± 0.65 | 0.3260 |
| TT | 278 | 2.01 ± 0.04 | 26.78 ± 0.32 | |||
| LEPR rs45435518 | CT | 53 | 1.88 ± 0.09 | 0.1070 | 26.10 ± 0.67 | 0.4410 |
| TT | 274 | 2.02 ± 0.04 | 26.63 ± 0.32 | |||
| MC4R rs81219178 | AA | 109 | 2.04 ± 0.05 | 0.2504 | 26.12 ± 0.67 | 0.7776 |
| AG | 175 | 1.95 ± 0.05 | 26.65 ± 0.32 | |||
| GG | 45 | 2.10 ± 0.09 | 26.82 ± 0.70 | |||
| PHKG1 rs697732005 | AA | 187 | 1.98 ± 0.07 | 0.2221 | 26.10 ± 0.41 | 0.1650 |
| AG | 89 | 2.14 ± 0.08 | 27.18 ± 0.52 | |||
| GG | 36 | 2.02 ± 0.12 | 26.74 ± 0.77 | |||
| RETN rs327132149 | AG | 20 | 1.96 ± 0.13 | 0.7730 | 27.81 ± 1.03 | 0.2080 |
| GG | 309 | 2.00 ± 0.04 | 26.50 ± 0.31 | |||
| RYR1 rs344435545 | CC | 309 | 2.01 ± 0.04 | 0.0850 | 26.61 ± 0.31 | 0.9550 |
| CT | 20 | 1.78 ± 0.13 | 26.56 ± 1.02 | |||
| SCD rs80912566 | CC | 59 | 2.09 ± 0.08 | 0.0510 | 27.01 ± 0.46 | 0.1666 |
| CT | 154 | 1.88 ± 0.05 | 26.76 ± 0.48 | |||
| TT | 117 | 2.01 ± 0.06 | 26.42 ± 0.37 | |||
| UBE3C rs81329544 | AA | 14 | 1.79 ± 0.15 | 0.1659 | 27.22 ± 1.22 | 0.8540 |
| AG | 117 | 1.95 ± 0.06 | 26.50 ± 0.44 | |||
| GG | 199 | 2.04 ± 0.05 | 26.59 ± 0.37 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Li, P.; Zhou, W.; Gao, C.; Liu, H.; Li, H.; Niu, P.; Zhang, Z.; Li, Q.; Zhou, J.; et al. Association of Twelve Candidate Gene Polymorphisms with the Intramuscular Fat Content and Average Backfat Thickness of Chinese Suhuai Pigs. Animals 2019, 9, 858. https://doi.org/10.3390/ani9110858
Wang B, Li P, Zhou W, Gao C, Liu H, Li H, Niu P, Zhang Z, Li Q, Zhou J, et al. Association of Twelve Candidate Gene Polymorphisms with the Intramuscular Fat Content and Average Backfat Thickness of Chinese Suhuai Pigs. Animals. 2019; 9(11):858. https://doi.org/10.3390/ani9110858
Chicago/Turabian StyleWang, Binbin, Pinghua Li, Wuduo Zhou, Chen Gao, Hang Liu, Huixia Li, Peipei Niu, Zongping Zhang, Qiang Li, Juan Zhou, and et al. 2019. "Association of Twelve Candidate Gene Polymorphisms with the Intramuscular Fat Content and Average Backfat Thickness of Chinese Suhuai Pigs" Animals 9, no. 11: 858. https://doi.org/10.3390/ani9110858