Comprehensive Analysis of Porcine Prox1 Gene and Its Relationship with Meat Quality Traits

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Committee
2.2. Animals and Sample Collection
2.3. RNA Extraction
2.4. Cloning of Full-Length cDNA and Proximal Promoter
2.5. Sequence Analysis
2.6. Gene Expression and Correlation Analysis
2.7. Promoter Activity Analysis
2.8. Identification of Variations
2.9. Traits and Genotyping
2.10. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of Porcine Prox1 Sequences
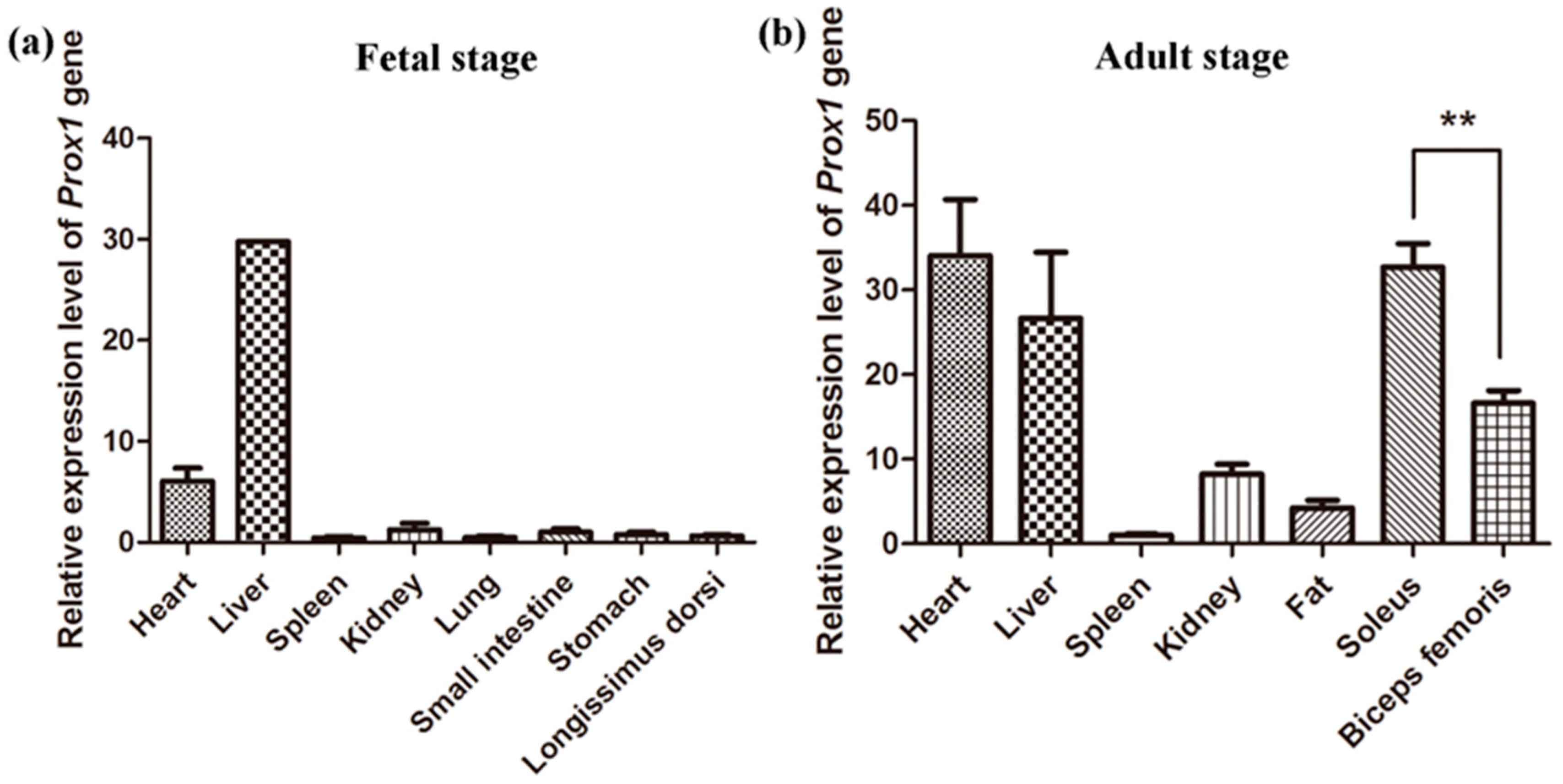
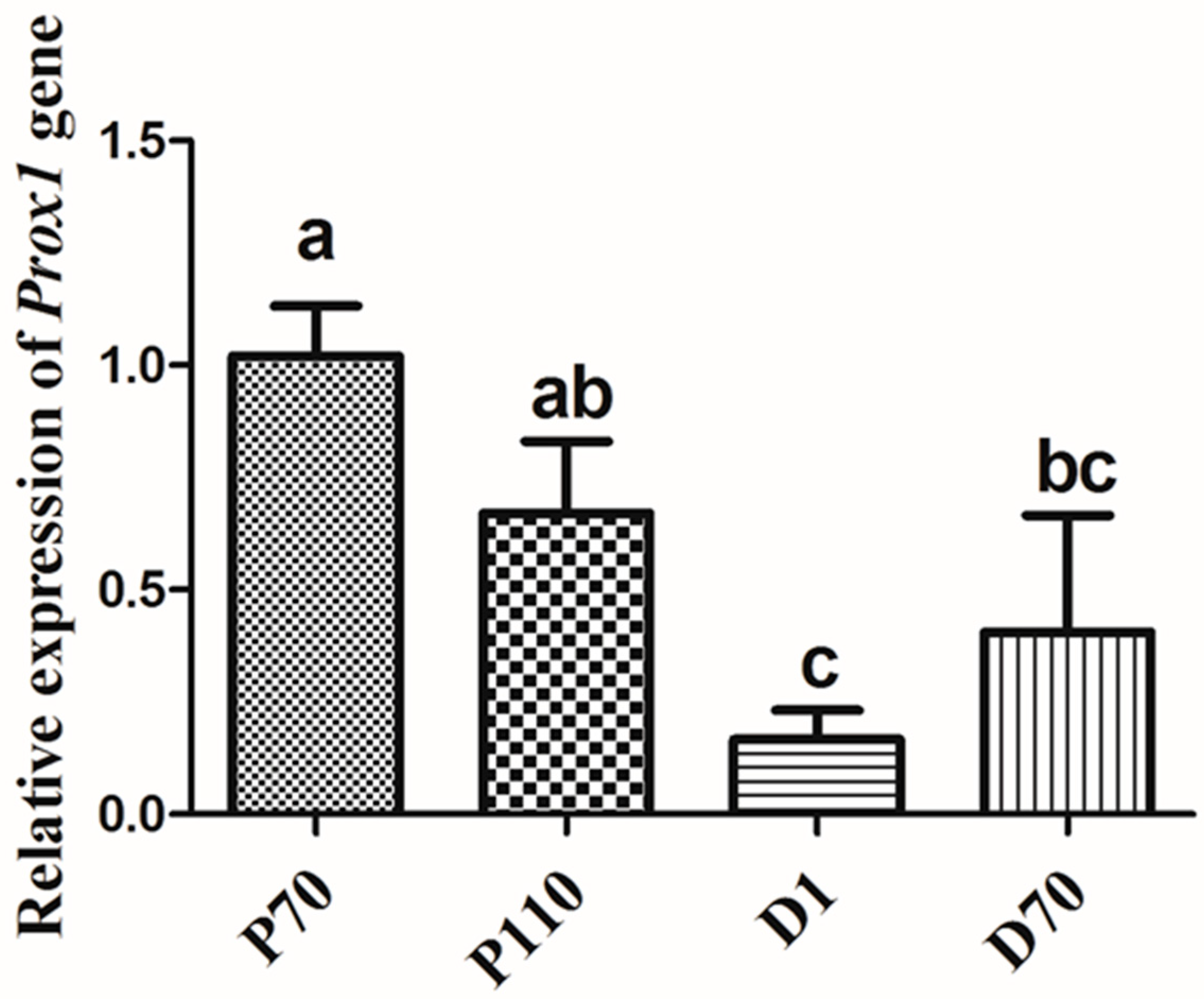
3.2. Expression Patterns of Porcine Prox1
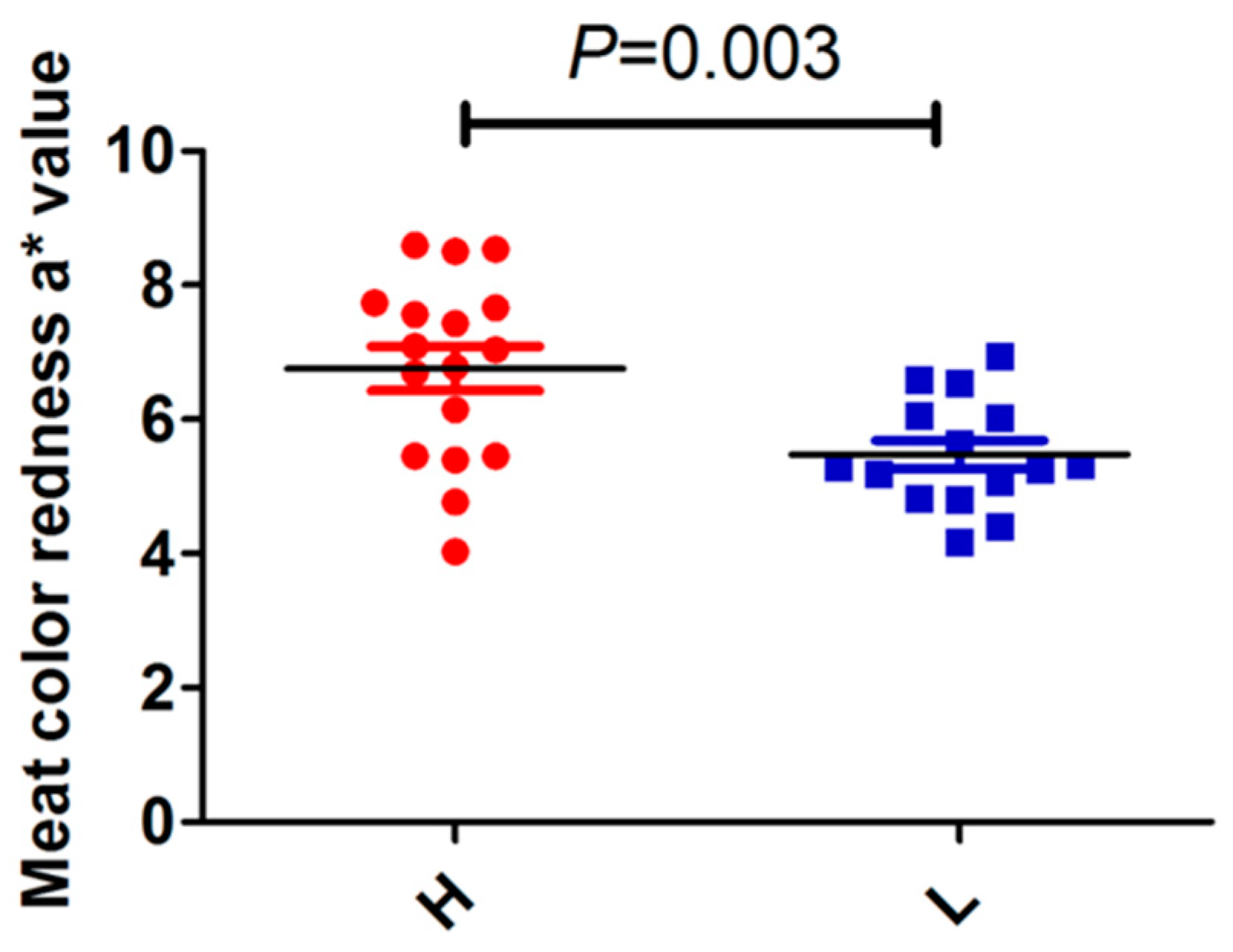
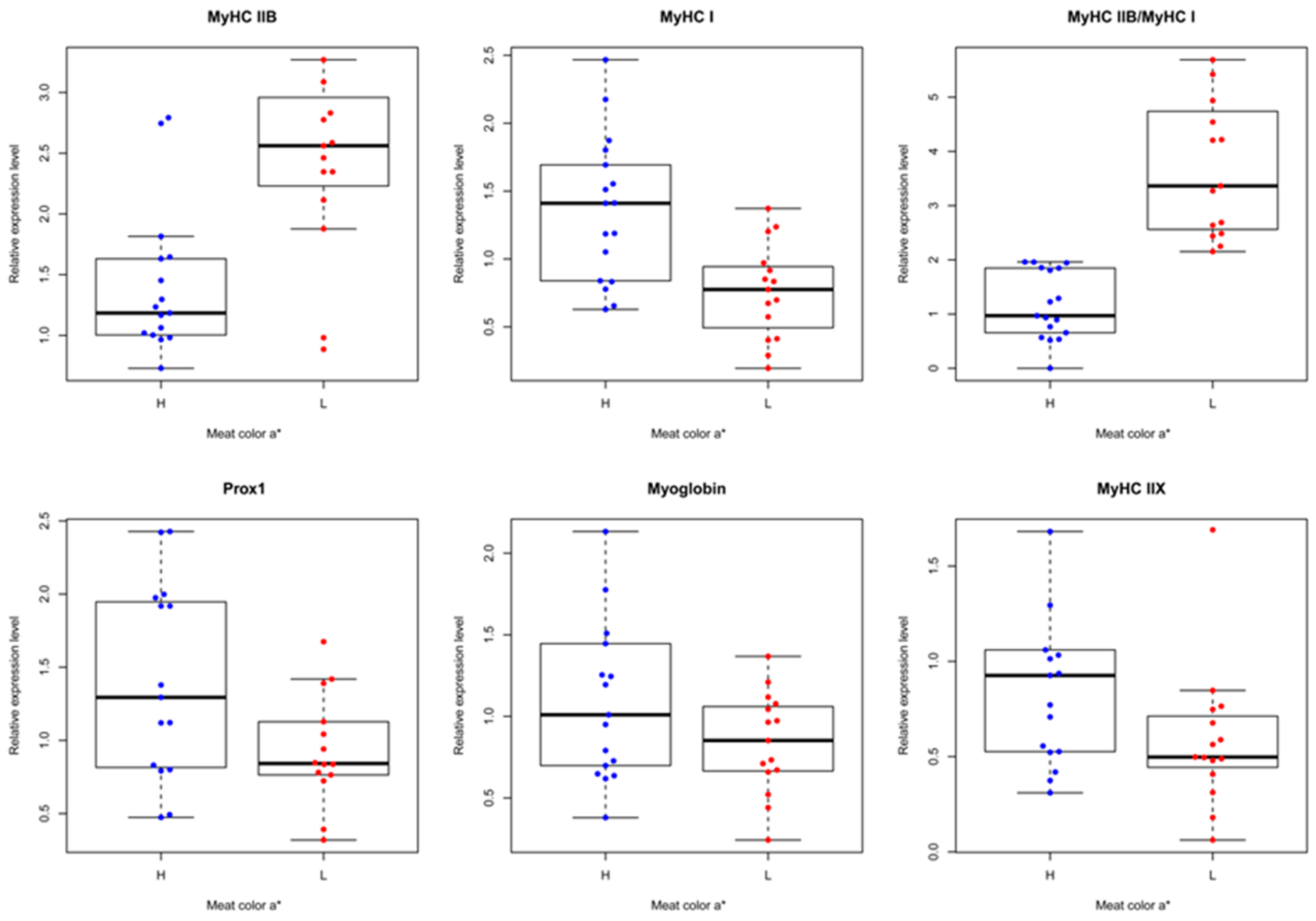
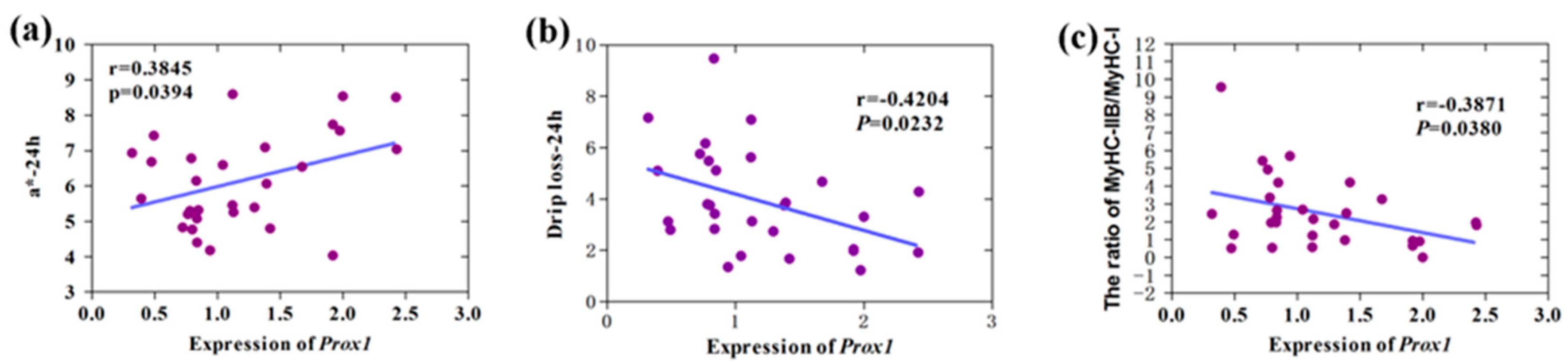
3.3. Correlation Analysis of Prox1 Expression with Meat Quality Traits
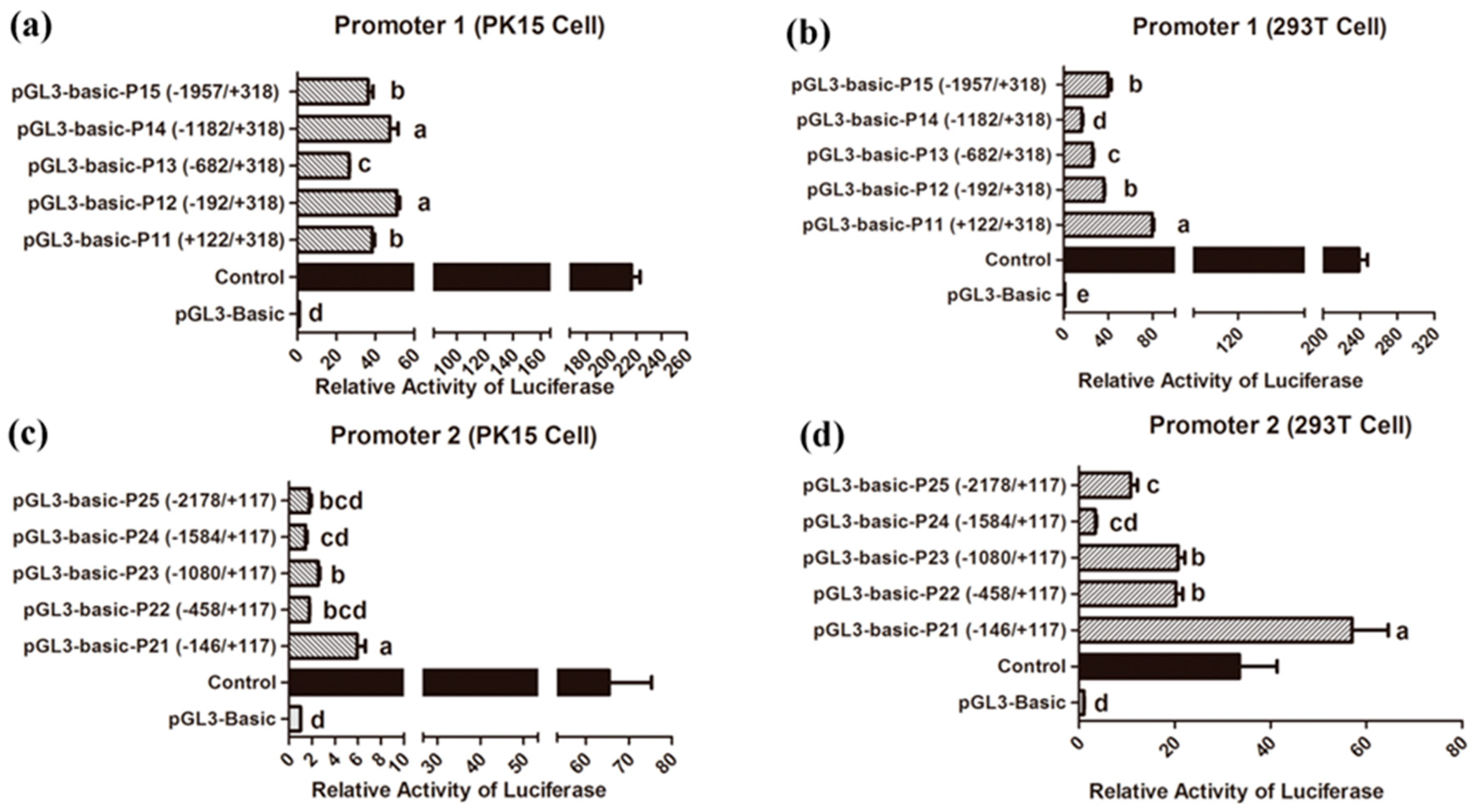
3.4. Identification of Core Promoter Region
3.5. Genetic Diversity of Sequence Variations
3.6. Trait Association Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sosa-Pineda, B.; Wigle, J.T.; Oliver, G. Hepatocyte migration during liver development requires Prox1. Nat. Genet. 2000, 25, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.L.; Srinivasan, R.S.; Dillard, M.E.; Johnson, N.C.; Witte, M.H.; Boyd, K.; Sleeman, M.W.; Oliver, G. Lymphatic vascular defects promoted by Prox1 haploinsufficiency cause adult-onset obesity. Nat. Genet. 2005, 37, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Wigle, J.T.; Oliver, G. Prox1 function is required for the development of the murine lymphatic system. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef]
- Wang, J.; Kilic, G.; Aydin, M.; Burke, Z.; Oliver, G.; Sosa-Pineda, B. Prox1 activity controls pancreas morphogenesis and participates in the production of “secondary transition” pancreatic endocrine cells. Dev. Biol. 2005, 286, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Wigle, J.T.; Chowdhury, K.; Gruss, P.; Oliver, G. Prox1 function is crucial for mouse lens-fibre elongation. Nat. Genet. 1999, 21, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Dyer, M.A.; Livesey, F.J.; Cepko, C.L.; Oliver, G. Prox1 function controls progenitor cell proliferation and horizontal cell genesis in the mammalian retina. Nat. Genet. 2003, 34, 53–58. [Google Scholar] [CrossRef]
- Armour, S.M.; Remsberg, J.R.; Damle, M.; Sidoli, S.; Ho, W.Y.; Li, Z.; Garcia, B.A.; Lazar, M.A. An HDAC3-PROX1 corepressor module acts on HNF4 alpha to control hepatic triglycerides. Nat. Commun. 2017, 8, 549. [Google Scholar] [CrossRef] [PubMed]
- Risebro, C.A.; Risebro, C.A.; Searles, R.G.; Melville, A.A.; Ehler, E.; Jina, N.; Shah, S.; Pallas, J.; Hubank, M.; Dillard, M.; et al. Prox1 maintains muscle structure and growth in the developing heart. Development 2009, 136, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Petchey, L.K.; Risebro, C.A.; Vieira, J.M.; Roberts, T.; Bryson, J.B.; Greensmith, L.; Lythgoe, M.F.; Riley, P.R. Loss of Prox1 in striated muscle causes slow to fast skeletal muscle fiber conversion and dilated cardiomyopathy. Proc. Natl. Acad. Sci. USA 2014, 111, 9515–9520. [Google Scholar] [CrossRef]
- An, C.I.; Dong, Y.; Hagiwara, N. Genome-wide mapping of Sox6 binding sites in skeletal muscle reveals both direct and indirect regulation of muscle terminal differentiation by Sox6. BMC Dev. Biol. 2011, 11, 59. [Google Scholar] [CrossRef]
- Kivela, R.; Salmela, I.; Nguyen, Y.H.; Petrova, T.V.; Koistinen, H.A.; Wiener, Z.; Alitalo, K. The transcription factor Prox1 is essential for satellite cell differentiation and muscle fibre-type regulation. Nat. Commun. 2016, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Li, B.J.; Dong, C.; Li, P.; Ren, Z.; Wang, H.; Yu, F.; Ning, C.; Liu, K.; Wei, W.; Huang, R.; et al. Identification of candidate genes associated with porcine meat color traits by genome-wide transcriptome analysis. Sci. Rep. 2016, 6, 35224. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.Q.; Chu-LaGraff, Q.; Wright, D.M.; Scott, M.P. The prospero gene specifies cell fates in the Drosophila central nervous system. Cell 1991, 65, 451–464. [Google Scholar] [CrossRef]
- Oliver, G.; Sosa-Pineda, B.; Geisendorf, S.; Spana, E.P.; Doe, C.Q.; Gruss, P. Prox 1, a prospero-related homeobox gene expressed during mouse development. Mech. Dev. 1993, 44, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Zinovieva, R.D.; Duncan, M.K.; Johnson, T.R.; Torres, R.; Polymeropoulos, M.H.; Tomarev, S.I. Structure and chromosomal localization of the human homeobox gene Prox 1. Genomics 1996, 35, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Elsir, T.; Smits, A.; Lindström, M.S.; Nistér, M. Transcription factor PROX1: Its role in development and cancer. Cancer Metastasis Rev. 2012, 31, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Liu, K.Q.; Li, B.J.; Dong, C.; Zhang, Z.K.; Li, P.H.; Huang, R.H.; Wei, W.; Chen, J.; Liu, H.L. Identification of an (AC)n microsatellite in the Six1 gene promoter and its effect on production traits in Pietrain x Duroc x Landrace x Yorkshire pigs. J. Anim. Sci. 2018, 96, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, X.; Zhang, J.B.; Ouyang, H.; Shen, Z.; Wu, Y.; Wang, W.; Wu, J.; Tao, S.; Yang, X. PROX1 promotes hepatocellular carcinoma proliferation and sorafenib resistance by enhancing beta-catenin expression and nuclear translocation. Oncogene 2015, 34, 5524–5535. [Google Scholar] [CrossRef] [PubMed]
- Nakamori, D.; Takayama, K.; Nagamoto, Y.; Mitani, S.; Sakurai, F.; Tachibana, M.; Mizuguchi, H. Hepatic maturation of human iPS cell-derived hepatocyte-like cells by ATF5, c/EBPalpha, and PROX1 transduction. Biochem. Biophys. Res. Commun. 2016, 469, 424–429. [Google Scholar] [CrossRef]
- Roy, S.; Wolff, C.; Ingham, P.W. The u-boot mutation identifies a Hedgehog-regulated myogenic switch for fiber-type diversification in the zebrafish embryo. Genes Dev. 2001, 15, 1563–1576. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, K. Excitation-transcription coupling in skeletal muscle: The molecular pathways of exercise. Biol. Rev. 2011, 86, 564–600. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.C.; Kim, B.C. The relationship between muscle fiber characteristics, postmortem metabolic rate, and meat quality of pig longissimus dorsi muscle. Meat Sci. 2005, 71, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Joo, S.T.; Ryu, Y.C. Skeletal muscle fiber type and myofibrillar proteins in relation to meat quality. Meat Sci. 2010, 86, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J.; Erdos, M.R.; Huyghe, J.R.; Welch, R.P.; Beck, A.T.; Wolford, B.N.; Chines, P.S.; Didion, J.P.; Narisu, N.; Stringham, H.M.; et al. The genetic regulatory signature of type 2 diabetes in human skeletal muscle. Nat. Commun. 2016, 7, 11764. [Google Scholar] [CrossRef] [Green Version]
- Hamet, P.; Haloui, M.; Harvey, F.; Marois-Blanchet, F.C.; Sylvestre, M.P.; Tahir, M.R.; Simon, P.H.G.; Kanzki, B.S.; Raelson, J.; Long, C.; et al. PROX1 gene CC genotype as a major determinant of early onset of type 2 diabetes in slavic study participants from Action in Diabetes and Vascular Disease: Preterax and Diamicron MR Controlled Evaluation study. J. Hypertens. 2017, 35 (Suppl. 1), S24–S32. [Google Scholar] [CrossRef] [PubMed]
- Kretowski, A.; Adamska, E.; Maliszewska, K.; Wawrusiewicz-Kurylonek, N.; Citko, A.; Goscik, J.; Bauer, W.; Wilk, J.; Golonko, A.; Waszczeniuk, M.; et al. The rs340874 PROX1 type 2 diabetes mellitus risk variant is associated with visceral fat accumulation and alterations in postprandial glucose and lipid metabolism. Genes Nutr. 2015, 10, 4. [Google Scholar] [CrossRef]
- Lecompte, S.; Pasquetti, G.; Hermant, X.; Grenier-Boley, B.; Gonzalez-Gross, M.; De Henauw, S.; Molnar, D.; Stehle, P.; Béghin, L.; Moreno, L.A.; et al. Genetic and Molecular Insights In to the Role of PROX1 in Glucose Metabolism. Diabetes 2013, 62, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Kauffman, R.G.; Kim, B.C.; Park, G.B. The relationship of sarcoplasmic and myofibrillar protein solubility to colour and water-holding capacity in porcine longissimus muscle. Meat Sci. 1999, 52, 291–297. [Google Scholar] [CrossRef]
- Van Laack, R.L.J.M.; Lane, J.L. Denaturation of myofibrillar proteins from chicken as affected by pH, temperature, and adenosine triphosphate concentration. Poult. Sci. 2000, 79, 105–109. [Google Scholar] [CrossRef]
- Choi, Y.M.; Lee, S.H.; Choe, J.H.; Rhee, M.S.; Lee, S.K.; Joo, S.T.; Kim, B.C. Protein solubility is related to myosin isoforms, muscle fiber types, meat quality traits, and postmortem protein changes in porcine longissimus dorsi muscle. Livestig. Sci. 2010, 131, 148. [Google Scholar] [CrossRef]
- Chan, J.T.Y.; Omana, D.A.; Betti, M. Effect of ultimate pH and freezing on the biochemical properties of proteins in turkey breast meat. Food Chem. 2011, 127, 109–117. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, W.G.; Zhou, G.H.; Guo, B. Comparison of protein degradation, protein oxidation, and mu-calpain activation between pale, soft, and exudative and red, firm, and nonexudative pork during postmortem aging. J. Anim. Sci. 2014, 92, 3745–3752. [Google Scholar] [CrossRef]
- O’Neill, D.J.; Lynch, P.B.; Troy, D.J.; Buckley, D.J.; Kerry, J.P. Influence of the time of year on the incidence of PSE and DFD in Irish pigmeat. Meat Sci. 2003, 64, 105–111. [Google Scholar] [CrossRef]
- Paul, L.; Walker, E.M.; Drosos, Y.; Cyphert, H.A.; Neale, G.; Stein, R.; South, J.; Grosveld, G.; Herrera, P.L.; Sosa-Pineda, B. Lack of Prox1 Downregulation Disrupts the Expansion and Maturation of Postnatal Murine beta-Cells. Diabetes 2016, 65, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Elbahrawy, A.; Furuyama, K.; Horiguchi, M.; Hosokawa, S.; Aoyama, Y.; Tsuboi, K.; Sakikubo, M.; Hirata, K.; Masui, T.; et al. Liver-specific Prox1 inactivation causes hepatic injury and glucose intolerance in mice. FEBS Lett. 2017, 591, 624–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassel-Duby, R.; Olson, E.N. Signaling pathways in skeletal muscle remodeling. Annu. Rev. Biochem. 2006, 75, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Bee, G.; Anderson, A.L.; Lonergan, S.M.; Huff-Lonergan, E. Rate and extent of pH decline affect proteolysis of cytoskeletal proteins and water-holding capacity in pork. Meat Sci. 2007, 76, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Bowker, B.; Zhuang, H. Relationship between water-holding capacity and protein denaturation in broiler breast meat. Poult. Sci. 2015, 94, 1657–1664. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Localization | Polymorphism | Chromosome | Reference Genome Position 1 |
|---|---|---|---|
| g. +123 | G/A | 9 | 142477946 |
| g. −268 | T/G | 9 | 142478335 |
| g. −414 | C/T | 9 | 142478482 |
| g. −427 | C/A | 9 | 142478495 |
| g. −502 | G/A | 9 | 142478570 |
| g. −516 | A/C | 9 | 142478584 |
| g. −759 | C/T | 9 | 142478827 |
| g. −780 | AGA/- | 9 | 142478846-142478848 |
| g. −848 | -/A | 9 | 142478913-142478913 |
| g. −924 | T/C | 9 | 142478992 |
| g. −930 | C/A | 9 | 142478998 |
| g. −1143 | G/T | 9 | 142479211 |
| g. −1179 | C/G | 9 | 142479247 |
| g. −1421 | A/G | 9 | 142479489 |
| g. −1448 | AC/- | 9 | 142479515-142479516 |
| g. −1452 | ACACAC/- | 9 | 142479515-142479520 |
| g. −1454 | ACACACAC/- | 9 | 142479515-142479522 |
| g. −1573 | C/G | 9 | 142479641 |
| Trait 1 | N 2 | Mean ± SD 3 | CV% 4 |
|---|---|---|---|
| Age (d) | 279 | 176.92 ± 4.14 | 2.34 |
| CW (kg) | 279 | 82.50 ± 11.10 | 13.46 |
| BF (mm) | 279 | 1.77 ± 0.48 | 27.13 |
| pH45min | 279 | 6.22 ± 0.30 | 4.78 |
| pH24h | 279 | 5.58 ± 0.18 | 3.28 |
| IMF (%) | 279 | 2.64 ± 0.91 | 34.55 |
| L*45min | 129 | 45.37 ± 2.29 | 5.04 |
| a*45min | 129 | 3.85 ± 1.500 | 38.92 |
| b*45min | 129 | 7.67 ± 0.93 | 12.14 |
| L* 24h | 279 | 51.49 ± 3.00 | 5.81 |
| a*24h | 279 | 6.32 ± 1.15 | 18.14 |
| b*24h | 279 | 5.88 ± 1.26 | 21.42 |
| DL24h (%) | 279 | 1.75 ± 0.97 | 55.31 |
| DL48h (%) | 279 | 3.89 ± 1.54 | 39.66 |
| CL (%) | 279 | 30.88 ± 3.48 | 11.26 |
| SF (N) | 279 | 47.23 ± 12.58 | 26.65 |
| MG (μmol/g) | 279 | 5.28 ± 1.71 | 32.44 |
| G (μmol/g) | 279 | 3.33 ± 0.84 | 25.23 |
| G6P (μmol/g) | 279 | 1.48 ± 0.19 | 12.69 |
| LA (μmol/g) | 279 | 130.82 ± 23.41 | 17.90 |
| GP (μmol/g) | 279 | 150.03 ± 26.27 | 17.50 |
| Traits | Genotype (g. −930 bp) | Genotype (g. −1421 bp) | Genotype (g. −1573 bp) | Number | Least Squares Means (LSM) ± SE | p Value |
|---|---|---|---|---|---|---|
| pH24h | CA | AG | CG | 87 | 5.55 ± 0.02 | 0.022 |
| CC | AA | CC | 184 | 5.60 ± 0.01 | ||
| Drip loss24h (%) | CA | AG | CG | 87 | 1.95 ± 0.12 | 0.070 |
| CC | AA | CC | 184 | 1.71 ± 0.09 | ||
| Drip loss48h (%) | CA | AG | CG | 87 | 5.04 ± 0.33 | 0.193 |
| CC | AA | CC | 184 | 4.78 ± 0.31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, C.; Zhang, X.; Liu, K.; Li, B.; Chao, Z.; Jiang, A.; Li, R.; Li, P.; Liu, H.; Wu, W. Comprehensive Analysis of Porcine Prox1 Gene and Its Relationship with Meat Quality Traits. Animals 2019, 9, 744. https://doi.org/10.3390/ani9100744
Dong C, Zhang X, Liu K, Li B, Chao Z, Jiang A, Li R, Li P, Liu H, Wu W. Comprehensive Analysis of Porcine Prox1 Gene and Its Relationship with Meat Quality Traits. Animals. 2019; 9(10):744. https://doi.org/10.3390/ani9100744
Chicago/Turabian StyleDong, Chao, Xiying Zhang, Kaiqing Liu, Bojiang Li, Zhe Chao, Aiwen Jiang, Rongyang Li, Pinghua Li, Honglin Liu, and Wangjun Wu. 2019. "Comprehensive Analysis of Porcine Prox1 Gene and Its Relationship with Meat Quality Traits" Animals 9, no. 10: 744. https://doi.org/10.3390/ani9100744