
Detection of Mannheimia haemolytica-Specific IgG, IgM and IgA in Sera and Their Relationship to Respiratory Disease in Cattle

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. M. haemolytica A1 Strain P1148 Culture
2.3. Development of M. haemolytica IgG, IgM, and IgA Isotype-Specific Indirect ELISAs
2.4. Study Population and Sampling for the Test Validation
2.5. Test Validation
2.6. Test Repeatability Determination
2.7. Statistical Analyses
3. Results
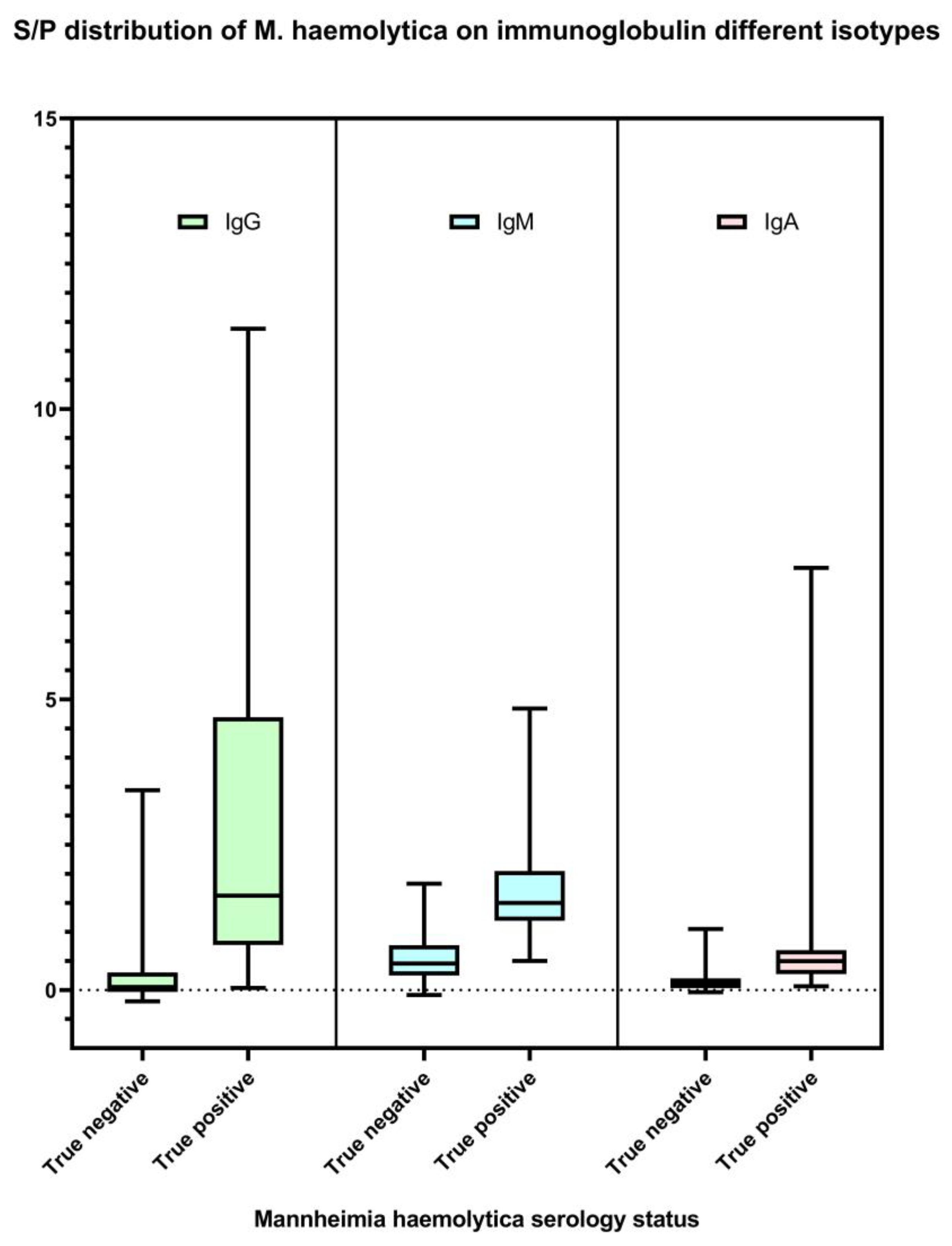
3.1. M. haemolytica IgG, IgM, and IgA ELISAs
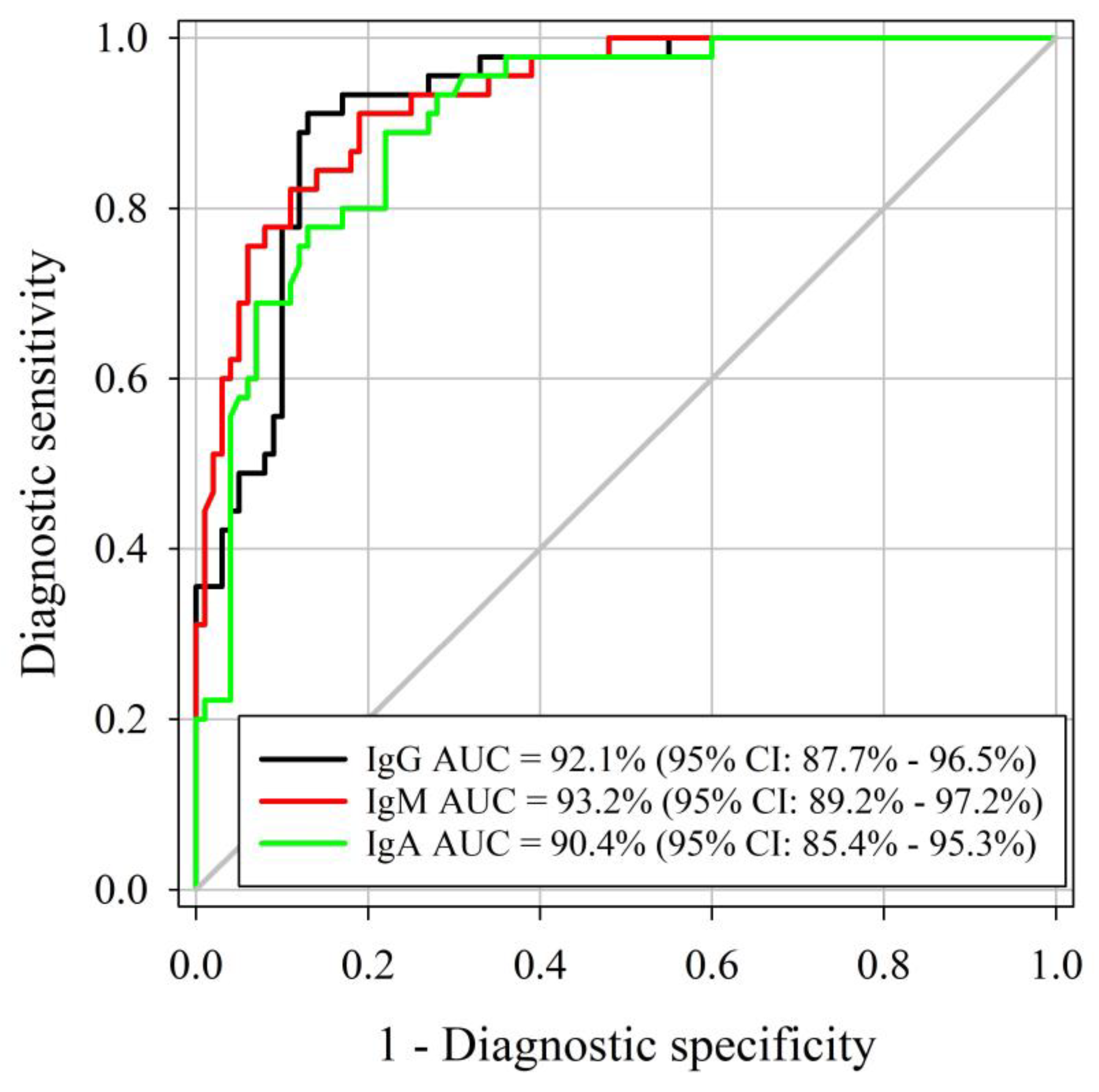
3.2. Determination of Specificity and Sensitivity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, T.A. Control methods for bovine respiratory disease for feedlot cattle. Vet. Clin. Food Anim. Pract. 2010, 26, 273–284. [Google Scholar] [CrossRef]
- Snowder, G.D.; Van Vleck, L.D.; Cundiff, L.V.; Bennett, G.L.; Koohmaraie, M.; Dikeman, M.E. Bovine respiratory disease in feedlot cattle: Phenotypic, environmental, and genetic correlations with growth, carcass, and longissimus muscle palatability traits. J. Anim. Sci. 2007, 85, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Booker, C.W.; Abutarbush, S.M.; Morley, P.S.; Jim, G.K.; Pittman, T.J.; Schunicht, O.C.; Perrett, T.; Wildman, B.K.; Fenton, R.K.; Guichon, P.T.; et al. Microbiological and histopathological findings in cases of fatal bovine respiratory disease of feedlot cattle in western Canada. Can. Vet. J. 2008, 49, 473. [Google Scholar] [PubMed]
- Blakebrough-Hall, C.; McMeniman, J.P.; González, L.A. An evaluation of the economic effects of bovine respiratory disease on animal performance, carcass traits, and economic outcomes in feedlot cattle defined using four BRD diagnosis methods. J. Anim. Sci. 2020, 98, skaa005. [Google Scholar] [CrossRef] [PubMed]
- ASAS Editorial Infographic. Preview: Economic effects of bovine respiratory disease. J. Anim. Sci. 2020, 98, 42. [Google Scholar]
- Whiteley, L.O.; Maheswaran, S.K.; Weiss, D.J.; Ames, T.R.; Kannan, M.S. Pasteurella haemolytica A1 and bovine respiratory disease: Pathogenesis. J. Vet. Intern. Med. 1992, 6, 11–22. [Google Scholar] [CrossRef]
- Klima, C.L.; Zaheer, R.; Cook, S.R.; Booker, C.W.; Hendrick, S.; Alexander, T.W.; McAllister, T.A. Pathogens of bovine respiratory disease in North American feedlots conferring multidrug resistance via integrative conjugative elements. J. Clin. Microbiol. 2014, 52, 438–448. [Google Scholar] [CrossRef]
- Fulton, R.W.; Blood, K.S.; Panciera, R.J.; Payton, M.E.; Ridpath, J.F.; Confer, A.W.; Saliki, J.T.; Burge, L.T.; Welsh, R.D.; Johnson, B.J.; et al. Lung pathology and infectious agents in fatal feedlot pneumonias and relationship with mortality, disease onset, and treatments. J. Vet. Diagn. Investig. 2009, 21, 464–477. [Google Scholar] [CrossRef]
- Lamm, C.G.; Love, B.C.; Krehbiel, C.R.; Johnson, N.J.; Step, D.L. Comparison of antemortem antimicrobial treatment regimens to antimicrobial susceptibility patterns of postmortem lung isolates from feedlot cattle with bronchopneumonia. J. Vet. Diagn. Investig. 2012, 24, 277–282. [Google Scholar] [CrossRef]
- Woolums, A.R. Pasteurellaceae: Avibacterium, Bibersteinia, Mannheimia, and Pasteurella. In Veterinary Microbiology, 3rd ed.; Wiley-Blackwell: Ames, IA, USA, 2013; pp. 101–114. [Google Scholar]
- Kiorpes, A.L. Pulmonary changes in rats following administration of 3-methylindole in cremophore EL. Histol. Histopathol. 1988, 3, 43–48. [Google Scholar]
- Van Donkersgoed, J.; Ribble, C.S.; Boyer, L.G.; Townsend, H.G. Epidemiological study of enzootic pneumonia in dairy calves in Saskatchewan. Can. J. Vet. Res. 1993, 57, 247–252. [Google Scholar]
- Ames, T.R. Dairy calf pneumonia: The disease and its impact. Vet. Clin. Food Anim. Pract. 1997, 13, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Frank, G.H.; Adlam, C.; Rutter, J.M. Pasteurella and pasteurellosis. In Pasteurellosis of Cattle, 1st ed.; Adlam, C., Rutter, J.M., Eds.; Academic Press Limited: London, UK, 1989; pp. 197–222. [Google Scholar]
- Carter, G.R.; Chengappa, M.M.; Roberts, A.W.; Claus, G.W.; Rikihisa, Y. Essentials of Veterinary Microbiology, 1st ed.; Williams & Wilkins: Baltimore, MD, USA, 1995. [Google Scholar]
- Rice, J.A.; Carrasco-Medina, L.; Hodgins, D.C.; Shewen, P.E. Mannheimia haemolytica and bovine respiratory disease. Anim. Health Res. Rev. 2007, 8, 117–128. [Google Scholar] [CrossRef]
- Haines, D.M.; Martin, K.M.; Clark, E.G.; Jim, G.K.; Janzen, E.D. The immunohistochemical detection of Mycoplasma bovis and bovine viral diarrhea virus in tissues of feedlot cattle with chronic, unresponsive respiratory disease and/or arthritis. Can. Vet. J. 2001, 42, 857–862. [Google Scholar] [PubMed]
- Taylor, J.D.; Holland, B.P.; Step, D.L.; Payton, M.E.; Confer, A.W. Nasal isolation of Mannheimia haemolytica and Pasteurella multocida as predictors of respiratory disease in shipped calves. Res. Vet. Sci. 2015, 99, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Snyder, E.; Credille, B. Mannheimia haemolytica and Pasteurella multocida in bovine respiratory disease: How are they changing in response to efforts to control them? Vet. Clin. Food Anim. Pract. 2020, 36, 253–268. [Google Scholar] [CrossRef]
- Kumar, J.; Dixit, S.K.; Kumar, R. Rapid detection of Mannheimia haemolytica in lung tissues of sheep and from bacterial culture. Vet. World 2015, 8, 1073–1077. [Google Scholar] [CrossRef]
- Fulton, R.W.; Confer, A.W. Laboratory test descriptions for bovine respiratory disease diagnosis and their strengths and weaknesses: Gold standards for diagnosis, do they exist? Can. Vet. J. 2012, 53, 754. [Google Scholar]
- Virtala, A.M.; Gröhn, Y.T.; Mechor, G.D.; Erb, H.N. The effect of maternally derived immunoglobulin G on the risk of respiratory disease in heifers during the first 3 months of life. Prev. Vet. Med. 1999, 39, 25–37. [Google Scholar] [CrossRef]
- Ellis, J.A. The immunology of the bovine respiratory disease complex. Vet. Clin. Food Anim. Pract. 2001, 17, 535–550. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. The distribution and functions of immunoglobulin isotypes. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Zamri-Saad, M.; Effendy, A.W.M.; Israf, D.A.; Azmi, M.L. Cellular and humoral responses in the respiratory tract of goats following intranasal stimulation using formalin-killed Pasteurella haemolytica A2. Vet. Microbiol. 1999, 65, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Huang, T.; Hammarström, L.; Zhao, Y. The immunoglobulins: New insights, implications, and applications. Annu. Rev. Anim. Biosci. 2020, 8, 145–169. [Google Scholar] [CrossRef] [PubMed]
- Sathe, A.; Cusick, J.K. Biochemistry, Immunoglobulin M (IgM). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Butler, J.E. Bovine immunoglobulins: A review. J. Dairy Sci. 1969, 52, 1895–1909. [Google Scholar] [CrossRef]
- Gapper, L.W.; Copestake, D.E.; Otter, D.E.; Indyk, H.E. Analysis of bovine immunoglobulin G in milk, colostrum and dietary supplements: A review. Anal. Bioanal. Chem. 2007, 389, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W., Jr.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. The mucosal immune system. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. Available online: https://www.ncbi.nlm.nih.gov/books/NBK27169/ (accessed on 22 February 2023).
- Prado, M.E.; Prado, T.M.; Payton, M.E.; Confer, A.W. Maternally and naturally acquired antibodies to Mannheimia haemolytica and Pasteurella multocida in beef calves. Vet. Immunol. Immunopathol. 2006, 111, 301–307. [Google Scholar] [CrossRef]
- Brennan, R.E.; Corstvet, R.E.; Paulson, D.B. Antibody responses to Pasteurella haemolytica 1: A and three of its outer membrane proteins in serum, nasal secretions, and bronchoalveolar lavage fluid from calves. Am. J. Vet. Res. 1998, 59, 727–732. [Google Scholar]
- Lainson, F.A.; Murray, J.; Davies, R.C.; Donachie, W. Characterization of epitopes involved in the neutralization of Pasteurella haemolytica serotype A1 leukotoxin. Microbiology 1996, 142, 2499–2507. [Google Scholar] [CrossRef]
- Moore, R.N.; Walker, R.D.; Shaw, G.A.; Hopkins, F.M.; Shull, E.P. Antileukotoxin antibody produced in the bovine lung after aerosol exposure to viable Pasteurella haemolytica. Am. J. Vet. Res. 1985, 46, 1949–1952. [Google Scholar]
- Townsend, E.L.; Maheswaran, S.K.; Leininger, J.R.; Ames, T.R. Detection of immunoglobulin G to Pasteurella haemolytica capsular polysaccharide by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1987, 25, 242–247. [Google Scholar] [CrossRef]
- Confer, A.W.; Ayalew, S. Mannheimia haemolytica in bovine respiratory disease: Immunogens, potential immunogens, and vaccines. Anim. Health Res. Rev. 2018, 19, 79–99. [Google Scholar] [CrossRef]
- Roier, S.; Fenninger, J.C.; Leitner, D.R.; Rechberger, G.N.; Reidl, J.; Schild, S. Immunogenicity of Pasteurella multocida and Mannheimia haemolytica outer membrane vesicles. Int. J. Med. Microbiol. 2013, 303, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Hille, M.M.; Spangler, M.L.; Clawson, M.L.; Heath, K.D.; Vu, H.L.; Rogers, R.E.; Loy, J.D. A Five Year Randomized Controlled Trial to Assess the Efficacy and Antibody Responses to a Commercial and Autogenous Vaccine for the Prevention of Infectious Bovine Keratoconjunctivitis. Vaccines 2022, 10, 916. [Google Scholar] [CrossRef]
- Confer, A.W.; Simons, K.R.; Panciera, R.J.; Mort, A.J.; Mosier, D.A. Serum antibody response to carbohydrate antigens of Pasteurella haemolytica serotype 1: Relation to experimentally induced bovine pneumonic pasteurellosis. Am. J. Vet. Res. 1989, 50, 98–105. [Google Scholar] [PubMed]
- Gorden, P.J.; Plummer, P. Control, management, and prevention of bovine respiratory disease in dairy calves and cows. Vet. Clin. Food Anim. Pract. 2010, 26, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Woolums, A.R.; Karisch, B.B.; Frye, J.G.; Epperson, W.; Smith, D.R.; Blanton, J., Jr.; Austin, F.; Kaplan, R.; Hiott, L.; Woodley, T.; et al. Multidrug resistant Mannheimia haemolytica isolated from high-risk beef stocker cattle after antimicrobial metaphylaxis and treatment for bovine respiratory disease. Vet. Microbiol. 2018, 221, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.Y.; Zimmerman, J.J.; Giménez-Lirola, L.G. Internal reference genes with the potential for normalizing quantitative PCR results for oral fluid specimens. Anim. Health Res. Rev. 2022, 23, 147–156. [Google Scholar] [CrossRef]
- Koo, T.K.; Li, M.Y. A guideline of selecting and reporting intraclass correlation coefficients for reliability research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef]
- Carpenter, J.; Bithell, J. Bootstrap confidence intervals: When, which, what? A practical guide for medical statisticians. Stat. Med. 2000, 19, 1141–1164. [Google Scholar] [CrossRef]
- Youden, W.J. Index for rating diagnostic tests. Cancer 1950, 3, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Rico, G.R.; Martínez-Castillo, M.; González-Ruíz, C.; Luna-Castro, S.; de la Garza, M. Mannheimia haemolytica A2 secretes different proteases into the culture medium and in outer membrane vesicles. Microb. Pathog. 2017, 113, 276–281. [Google Scholar] [CrossRef]
- Singh, K.; Ritchey, J.W.; Confer, A.W. Mannheimia haemolytica: Bacterial–host interactions in bovine pneumonia. Vet. Pathol. 2011, 48, 338–348. [Google Scholar] [CrossRef]
- Larson, R.L.; Step, D.L. Evidence-based effectiveness of vaccination against Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni in feedlot cattle for mitigating the incidence and effect of bovine respiratory disease complex. Vet. Clin. Food Anim. Pract. 2012, 28, 97–106. [Google Scholar] [CrossRef]
- Czuprynski, C.J.; Leite, F.; Sylte, M.; Kuckleburg, C.; Schultz, R.; Inzana, T.; Behling-Kelly, E.; Corbeil, L. Complexities of the pathogenesis of Mannheimia haemolytica and Haemophilus somnus infections: Challenges and potential opportunities for prevention? Anim. Health Res. Rev. 2004, 5, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Biberstein, E.L.; Gills, M.; Knight, H. Serological types of Pasteurella hemolytica. Cornell Vet. 1960, 50, 283–300. [Google Scholar]
- Biberstein, E.L.; Gills, M.G. The relation of the antigenic types to the A and T types of Pasteurella haemolytica. J. Comp. Pathol. Ther. 1962, 72, 316–320. [Google Scholar] [CrossRef]
- Fodor, L.; Varga, J.; Hajtos, J.; Donachie, W.; Gilmour, N.J. Characterisation of a new serotype of P. haemolytica isolated in Hungary. Res. Vet. Sci. 1988, 44, 399. [Google Scholar] [CrossRef]
- Frank, G.H.; Wessman, G.E. Rapid plate agglutination procedure for serotyping Pasteurella haemolytica. J. Clin. Microbiol. 1978, 7, 142–145. [Google Scholar] [CrossRef]
- Pardon, B.; Alliët, J.; Boone, R.; Roelandt, S.; Valgaeren, B.; Deprez, P. Prediction of respiratory disease and diarrhea in veal calves based on immunoglobulin levels and the serostatus for respiratory pathogens measured at arrival. Prev. Vet. Med. 2015, 120, 169–176. [Google Scholar] [CrossRef]
- Opuda-Asibo, J.; Maheswaran, S.K.; Leininger, J.R. Measurement of Pasteurella haemolytica-specific lung and serum antibodies by ELISA. Vet. Microb. 1986, 12, 337–351. [Google Scholar] [CrossRef]
- Klima, C.L.; Zaheer, R.; Briggs, R.E.; McAllister, T.A. A multiplex PCR assay for molecular capsular serotyping of Mannheimia haemolytica serotypes 1, 2, and 6. J. Microbiol. Methods 2017, 139, 155–160. [Google Scholar] [CrossRef]
- Blberstein, E.L. Chapter IX Biotyping and Serotyping of Pasteurella haemolytica. In Methods in Microbiology; Academic Press: Cambridge, MA, USA, 1978; Volume 10, pp. 253–269. [Google Scholar]
- Fraser, J.; Donachie, W.; Quirie, M.; Gilmour, N.J.L. Rapid indirect hemagglutination test for serotyping Pasteurella haemolytica. J. Clin. Microbiol. 1983, 18, 206–207. [Google Scholar] [CrossRef]
- Fodor, L.; Pénzes, Z.; Varga, J. Coagglutination test for serotyping Pasteurella haemolytica. J. Clin. Microbiol. 1996, 34, 393–397. [Google Scholar] [CrossRef]
- Quirie, M.; Donachie, W.; Gilmour, N.J. Serotypes of Pasteurella haemolytica from cattle. Vet. Rec. 1986, 119, 93–94. [Google Scholar] [CrossRef] [PubMed]
- Angen, Ø.; Mutters, R.; Caugant, D.A.; Olsen, J.E.; Bisgaard, M. Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridizations and 16S rRNA sequencing with proposal of Mannheimia haemolytica gen. nov., comb. nov., Mannheimia granulomatis comb. nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 67–86. [Google Scholar]
- Clawson, M.L.; Murray, R.W.; Sweeney, M.T.; Apley, M.D.; DeDonder, K.D.; Capik, S.F.; Larson, R.L.; Lubbers, B.V.; White, B.J.; Kalbfleisch, T.S.; et al. Genomic signatures of Mannheimia haemolytica that associate with the lungs of cattle with respiratory disease, an integrative conjugative element, and antibiotic resistance genes. BMC Genom. 2016, 17, 982. [Google Scholar] [CrossRef]
- Loy, J.D.; Clawson, M.L. Rapid typing of Mannheimia haemolytica major genotypes 1 and 2 using MALDI-TOF mass spectrometry. J. Microbiol. Methods 2017, 136, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Clawson, M.L.; Schuller, G.; Dickey, A.M.; Bono, J.L.; Murray, R.W.; Sweeney, M.T.; Apley, M.D.; DeDonder, K.D.; Capik, S.F.; Larson, R.L.; et al. Differences between predicted outer membrane proteins of genotype 1 and 2 Mannheimia haemolytica. BMC Microbiol. 2020, 20, 250. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, V.; Ciriaco-Solano, L.; Rodríguez-Patiño, G.; López-Arellano, R.; Revilla-Vázquez, A.; Morales-Álvarez, J. Respuesta serológica contra Mannheimia haemolytica y su leucotoxina en conejos suplementados con selenio. Abanico Vet. 2020, 10, 1–15. [Google Scholar]
- Sabri, M.Y.; Zamri-Saad, M.; Mutalib, A.R.; Israf, D.A.; Muniandy, N. Efficacy of an outer membrane protein of Pasteurella haemolytica A2, A7 or A9-enriched vaccine against intratracheal challenge exposure in sheep. Vet. Microbiol. 2000, 73, 13–23. [Google Scholar] [CrossRef]
- Effendy, A.W.M.; Zamri-Saad, M.; Puspa, R.; Rosiah, S. Efficacy of intranasal administration of formalin-killed Pasteurella haemolytica A2 against intratracheal challenge in goats. Vet. Rec. 1998, 142, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Srinand, S.; Hsuan, S.L.; Yoo, H.S.; Maheswaran, S.K.; Ames, T.R.; Werdin, R.E. Comparative evaluation of antibodies induced by commercial Pasteurella haemolytica vaccines using solid phase immunoassays. Vet. Microbiol. 1996, 49, 181–195. [Google Scholar] [CrossRef]
- Zheng, T.; Gupta, S.K.; McCarthy, A.R.; Moffat, J.; Buddle, B.M. Cross-protection study of a Mannheimia haemolytica serotype 1 vaccine against acute pasteurellosis in lambs induced by a serotype 2 strain. Vet. Microbiol. 2015, 177, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Rotolo, M.L.; Giménez-Lirola, L.; Ji, J.; Magtoto, R.; Henao-Díaz, Y.A.; Wang, C.; Baum, D.H.; Harmon, K.M.; Main, R.G.; Zimmerman, J.J. Detection of porcine reproductive and respiratory syndrome virus (PRRSV)-specific IgM-IgA in oral fluid samples reveals PRRSV infection in the presence of maternal antibody. Vet. Microbiol. 2018, 214, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Goi, A.; Penasa, M.; Nardino, G.; Posenato, L.; De Marchi, M. Variation of immunoglobulins G, A, and M and bovine serum albumin concentration in Holstein cow colostrum. Animal 2021, 15, 100299. [Google Scholar] [CrossRef]
- Ahmann, J.; Steinhoff-Wagner, J.; Büscher, W. Determining immunoglobulin content of bovine colostrum and factors affecting the outcome: A review. Animals 2021, 11, 3587. [Google Scholar] [CrossRef]
- Hodgins, D.C.; Shewen, P.E. Passive immunity to Pasteurella haemolytica A1 in dairy calves: Effects of preparturient vaccination of the dams. Can. J. Vet. Res. 1994, 58, 31. [Google Scholar]



{kind=link}
{kind=link}
| Inter-Assay Repeatability (30 Samples, 4 Rounds) | Intra-Assay Repeatability (8 Samples, 12 Rounds) | ||
|---|---|---|---|
| ICC (95% CI) | ICC (95% CI) | ||
| Antibody isotype | IgG | 97.6% (95.5%, 98.8%) | 99.9% (99.8%, 100.0%) |
| IgM | 87.2% (77.7%, 93.3%) | 99.9% (99.7%, 100.0%) | |
| IgA | 98.6% (97.6%, 99.3%) | 99.6% (99.1%, 99.9%) |
| IgG | IgM | IgA | ||||
|---|---|---|---|---|---|---|
| Cutoff (S/P) | Diagnostic Sensitivity (95% CI) | Diagnostic Specificity (95% CI) | Diagnostic Sensitivity (95% CI) | Diagnostic Specificity (95% CI) | Diagnostic Sensitivity (95% CI) | Diagnostic Specificity (95% CI) |
| 0.1 | 97.8 (93.3, 100.0) | 59.0 (49.0, 69.0) | 100.0 (100.0, 100.0) | 6.0 (2.0, 11.0) | 97.8 (93.3, 100.0) | 50.0 (40.0, 60.0) |
| 0.2 | 95.6 (88.9, 100.0) | 67.0 (58.0, 76.0) | 100.0 (100.0, 100.0) | 20.0 (13.0, 28.0) | 88.9 (77.8, 97.8) | 74.0 (65.0, 82.0) |
| 0.3 | 93.3 (84.4, 100.0) | 75.0 (66.0, 83.0) | 100.0 (100.0, 100.0) | 31.0 (22.0, 40.0) | 68.9 (55.6, 82.2) | 89.0 (82.0, 95.0) |
| 0.4 | 91.1 (82.2, 97.8) | 85.0 (78.0, 91.0) | 100.0 (100.0, 100.0) | 45.0 (36.0, 55.0) | 57.8 (44.4, 71.1) | 94.0 (89.0, 98.0) |
| 0.5 | 91.1 (82.2, 97.8) | 87.0 (80.0, 93.0) | 100.0 (100.0, 100.0) | 52.0 (42.0, 62.0) | 46.7 (33.3, 62.2) | 96.0 (92.0, 99.0) |
| 0.6 | 86.7 (75.6, 95.6) | 88.0 (81.0, 94.0) | 97.8 (93.3, 100.0) | 61.0 (51.0, 70.0) | 35.6 (22.2, 48.9) | 96.0 (92.0, 99.0) |
| 0.7 | 77.8 (64.4, 88.9) | 90.0 (84.0, 95.0) | 93.3 (84.4, 100.0) | 66.0 (57.0, 75.0) | 24.4 (13.3, 37.8) | 96.0 (92.0, 99.0) |
| 0.8 | 73.3 (60.0, 86.7) | 90.0 (84.0, 95.0) | 91.1 (82.2, 97.8) | 77.0 (69.0, 85.0) | 20.0 (8.9, 31.1) | 99.0 (97.0, 100.0) |
| 0.9 | 73.3 (60.0, 86.7) | 90.0 (84.0, 95.0) | 84.4 (73.3, 93.3) | 82.0 (74.0, 89.0) | 20.0 (8.9, 31.1) | 99.0 (97.0, 100.0) |
| 1.0 | 66.7 (53.3, 80.0) | 90.0 (84.0, 95.0) | 82.2 (71.1, 93.3) | 89.0 (82.0, 95.0) | 20.0 (8.9, 31.1) | 99.0 (97.0, 100.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poonsuk, K.; Kordik, C.; Hille, M.; Cheng, T.-Y.; Crosby, W.B.; Woolums, A.R.; Clawson, M.L.; Chitko-McKown, C.; Brodersen, B.; Loy, J.D. Detection of Mannheimia haemolytica-Specific IgG, IgM and IgA in Sera and Their Relationship to Respiratory Disease in Cattle. Animals 2023, 13, 1531. https://doi.org/10.3390/ani13091531
Poonsuk K, Kordik C, Hille M, Cheng T-Y, Crosby WB, Woolums AR, Clawson ML, Chitko-McKown C, Brodersen B, Loy JD. Detection of Mannheimia haemolytica-Specific IgG, IgM and IgA in Sera and Their Relationship to Respiratory Disease in Cattle. Animals. 2023; 13(9):1531. https://doi.org/10.3390/ani13091531
Chicago/Turabian StylePoonsuk, Korakrit, Carita Kordik, Matthew Hille, Ting-Yu Cheng, William B. Crosby, Amelia R. Woolums, Michael L. Clawson, Carol Chitko-McKown, Bruce Brodersen, and John Dustin Loy. 2023. "Detection of Mannheimia haemolytica-Specific IgG, IgM and IgA in Sera and Their Relationship to Respiratory Disease in Cattle" Animals 13, no. 9: 1531. https://doi.org/10.3390/ani13091531